
The Common Bean Small Heat Shock Protein Nodulin 22 from Phaseolus vulgaris L. Assembles into Functional High-Molecular-Weight Oligomers
 , , , , and
, , , , and 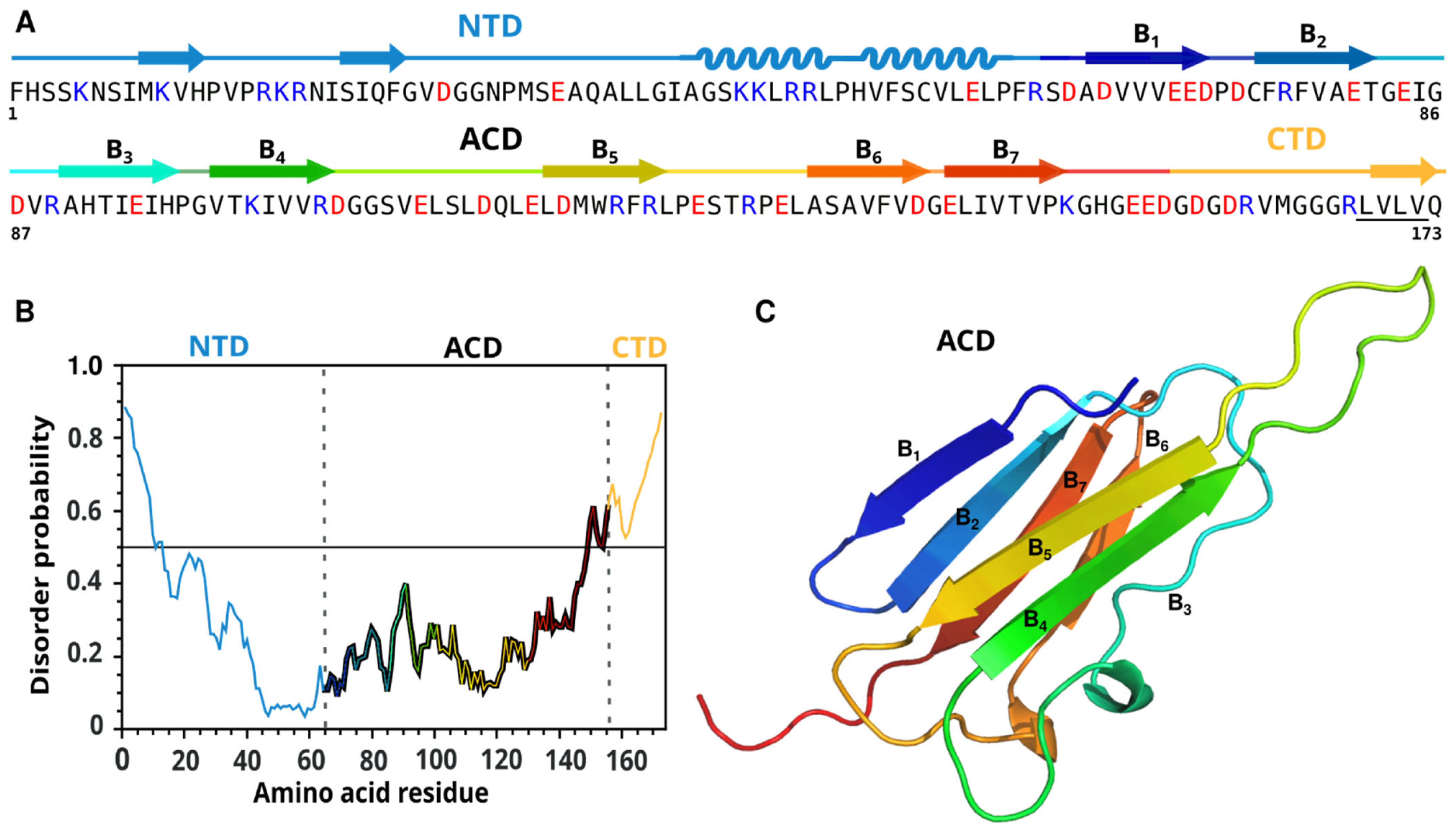{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Bioinformatic Analysis and Molecular Modeling of PvNod22
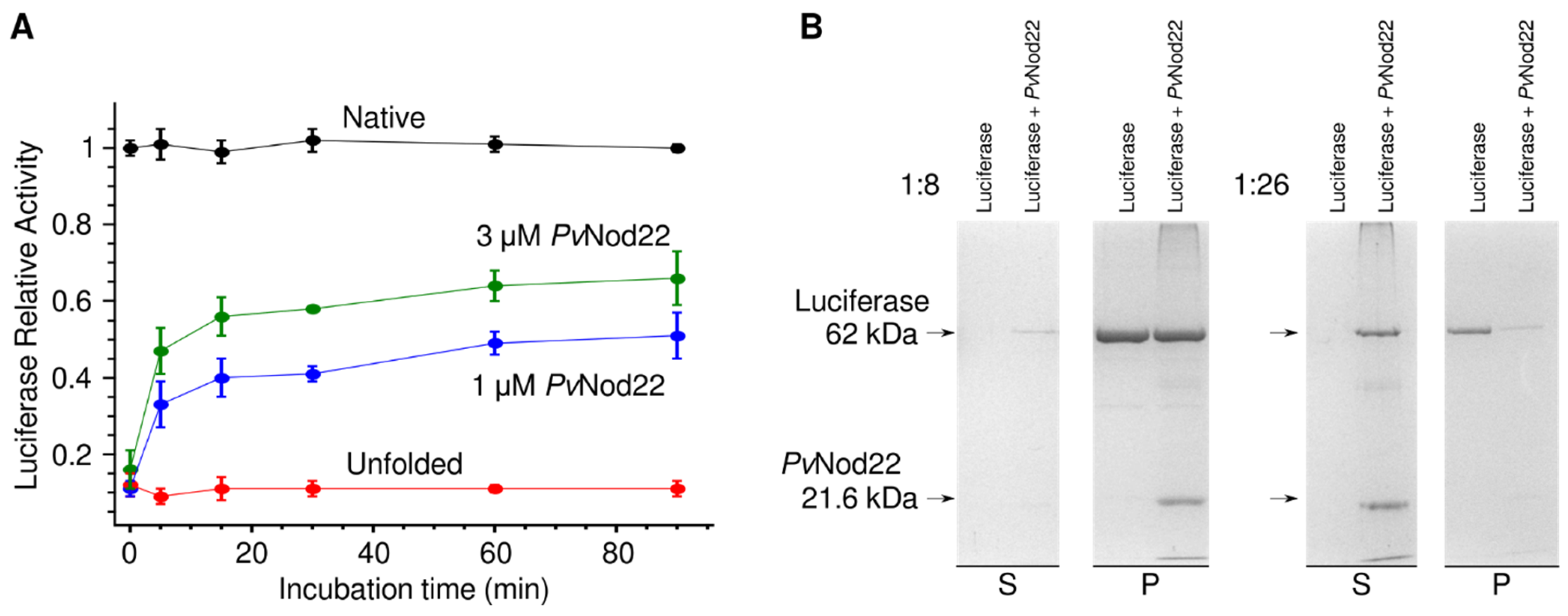
2.2. PvNod22 Has a Chaperone Function In Vitro
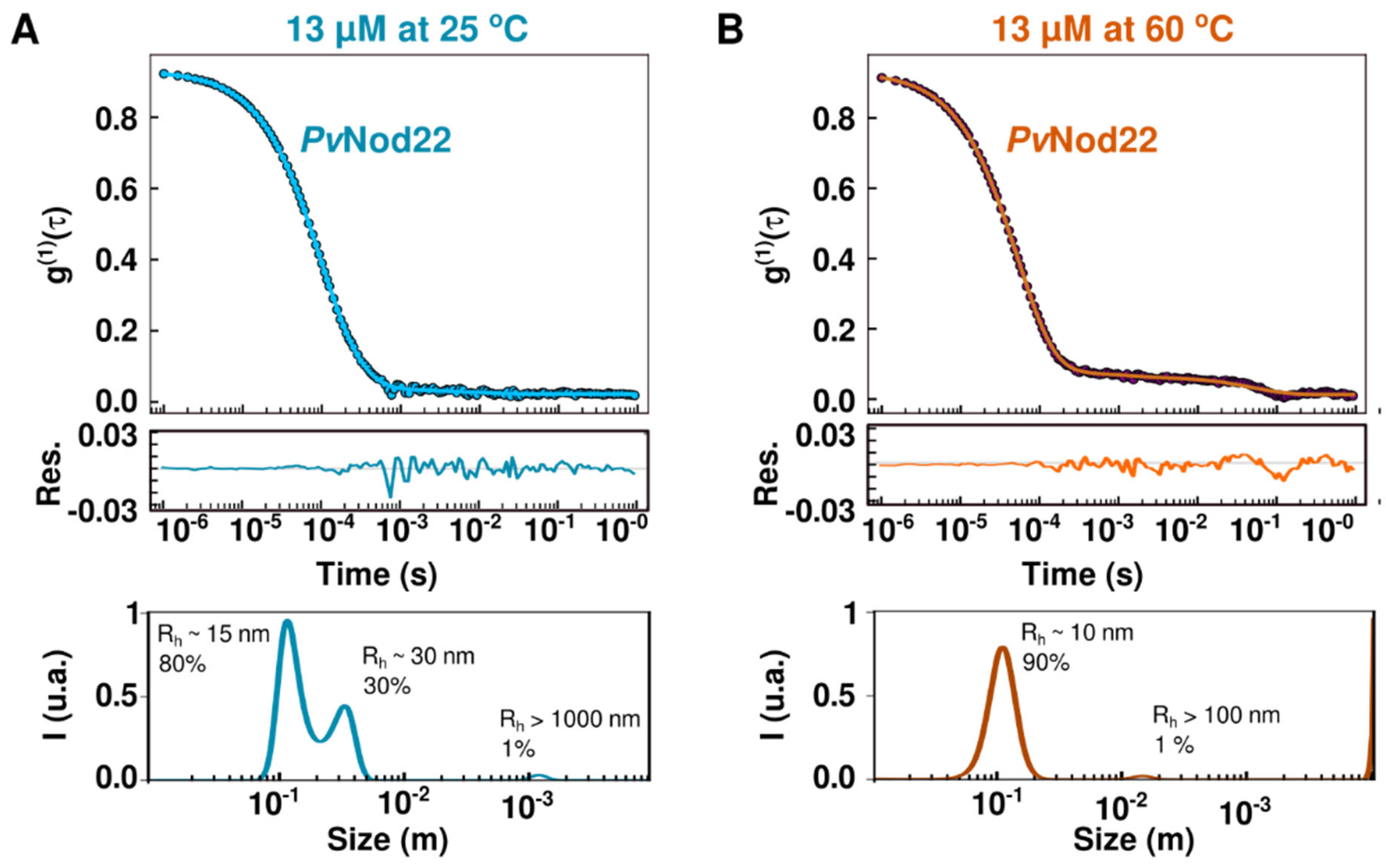
2.3. PvNod22 Exists as HMW Oligomers under Non-Stress Conditions
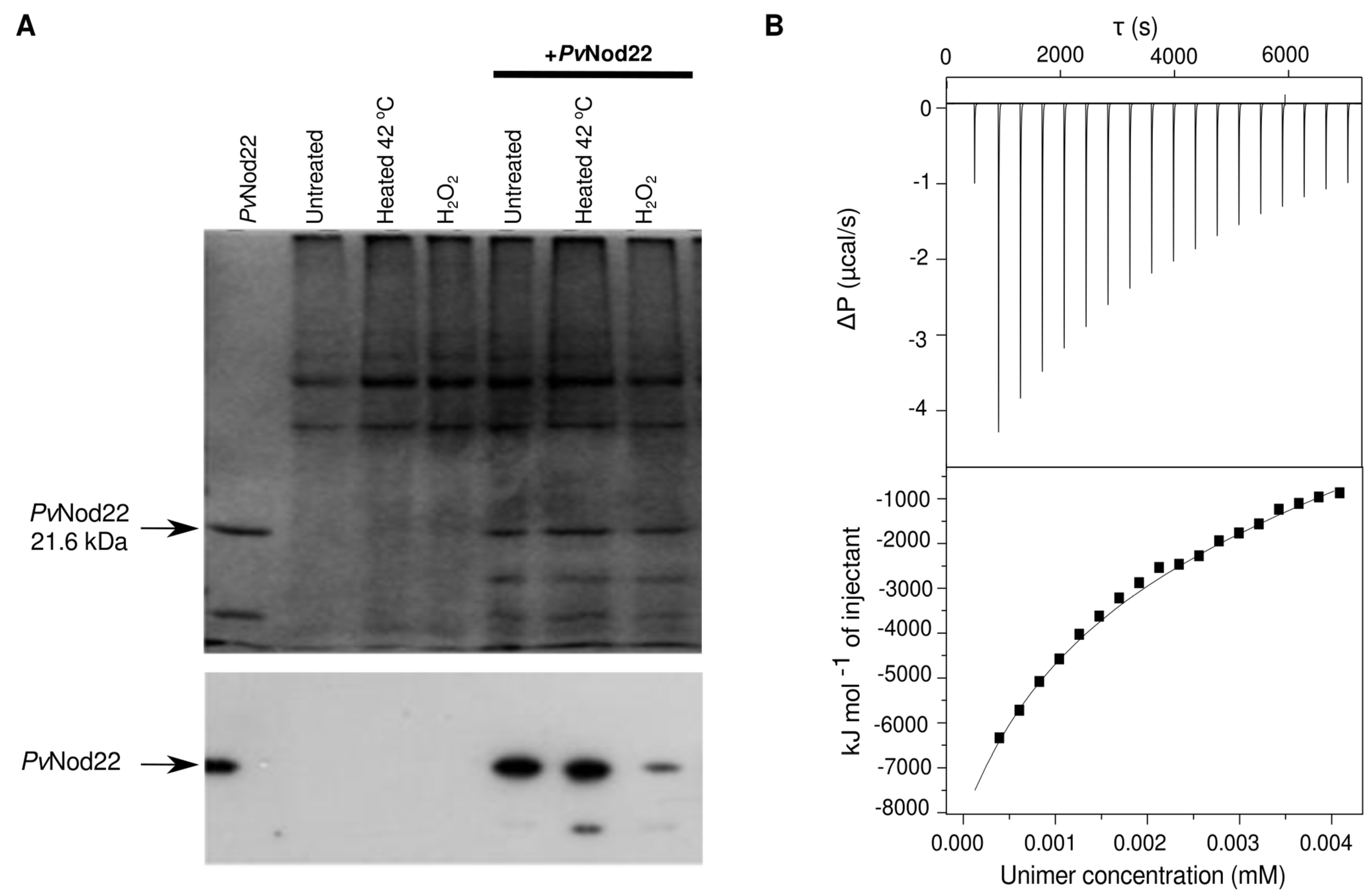
2.4. Dynamics of Recombinant PvNod22 Oligomeric States under Stress Conditions
2.5. PvNod22–PvNod22 Interactions
3. Discussion
4. Materials and Methods
4.1. Expression and Purification
4.2. Anti-Precipitation Activity Assay
4.3. In Silico Modeling and Identification of Intrinsically Disordered Regions in the Protein
4.4. Cross-Linking
4.5. Affinity Chromatography
4.6. PvNod22 Antibody and Immunoblot Analysis
4.7. Electron Microscopy
4.8. Dynamic Light Scattering
4.9. Isothermal Titration Calorimetry
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haslbeck, M.; Vierling, E. A First Line of Stress Defense: Small Heat Shock Proteins and Their Function in Protein Homeostasis. J. Mol. Biol. 2015, 427, 1537–1548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, E.R.; Aevermann, B.D.; Sanders-Reed, Z. Comparative Analysis of the Small Heat Shock Proteins in Three Angiosperm Genomes Identifies New Subfamilies and Reveals Diverse Evolutionary Patterns. Cell Stress Chaperones 2008, 13, 127–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddique, M.; Gernhard, S.; Von Koskull-Döring, P.; Vierling, E.; Scharf, K.D. The Plant SHSP Superfamily: Five New Members in Arabidopsis Thaliana with Unexpected Properties. Cell Stress Chaperones 2008, 13, 183–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basha, E.; O’Neill, H.; Vierling, E.; O’Neill, H.; Vierling, E. Small Heat Shock Proteins and α-Crystallins: Dynamic Proteins with Flexible Functions. Trends Biochem. Sci. 2012, 37, 106–117. [Google Scholar] [CrossRef] [Green Version]
- Waters, E.R.; Vierling, E. Plant Small Heat Shock Proteins—Evolutionary and Functional Diversity. New Phytol. 2020, 227, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Van Montagu, M.; Verbruggen, N. Small Heat Shock Proteins and Stress Tolerance in Plants. Biochim. Biophys. Acta-Gene Struct. Expr. 2002, 1577, 1–9. [Google Scholar] [CrossRef]
- Sudnitsyna, M.V.; Seit-Nebi, A.S.; Gusev, N.B. Cofilin Weakly Interacts with 14-3-3 and Therefore Can Only Indirectly Participate in Regulation of Cell Motility by Small Heat Shock Protein HspB6 (Hsp20). Arch. Biochem. Biophys. 2012, 521, 62–70. [Google Scholar] [CrossRef]
- Kim, D.H.; Xu, Z.Y.; Na, Y.J.; Yoo, Y.J.; Lee, J.; Sohn, E.J.; Hwang, I. Small Heat Shock Protein Hsp17.8 Functions as an AKR2A Cofactor in the Targeting of Chloroplast Outer Membrane Proteins in Arabidopsis. Plant Physiol. 2011, 157, 132–146. [Google Scholar] [CrossRef] [Green Version]
- Balogi, Z.; Cheregi, O.; Giese, K.C.; Juhász, K.; Vierling, E.; Vass, I.; Vígh, L.; Horváth, I. A Mutant Small Heat Shock Protein with Increased Thylakoid Association Provides an Elevated Resistance Against UV-B Damage in Synechocystis 6803. J. Biol. Chem. 2008, 283, 22983–22991. [Google Scholar] [CrossRef] [Green Version]
- Török, Z.; Goloubinoff, P.; Horváth, I.; Tsvetkova, N.M.; Glatz, A.; Balogh, G.; Varvasovszki, V.; Los, D.A.; Vierling, E.; Crowe, J.H.; et al. Synechocystis HSP17 Is an Amphitropic Protein That Stabilizes Heat-Stressed Membranes and Binds Denatured Proteins for Subsequent Chaperone-Mediated Refolding. Proc. Natl. Acad. Sci. USA 2001, 98, 3098–3103. [Google Scholar] [CrossRef]
- Haslbeck, M.; Weinkauf, S.; Buchner, J. Small Heat Shock Proteins: Simplicity Meets Complexity. J. Biol. Chem. 2019, 294, 2121–2132. [Google Scholar] [CrossRef] [Green Version]
- Jehle, S.; Vollmar, B.S.; Bardiaux, B.; Dove, K.K.; Rajagopal, P.; Gonen, T.; Oschkinat, H.; Klevit, R.E. N-Terminal Domain of AB-Crystallin Provides a Conformational Switch for Multimerization and Structural Heterogeneity. Proc. Natl. Acad. Sci. USA 2011, 108, 6409–6414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carver, J.A.; Grosas, A.B.; Ecroyd, H.; Quinlan, R.A. The Functional Roles of the Unstructured N- and C-Terminal Regions in AB-Crystallin and Other Mammalian Small Heat-Shock Proteins. Cell Stress Chaperones 2017, 22, 627–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriehuber, T.; Rattei, T.; Weinmaier, T.; Bepperling, A.; Haslbeck, M.; Buchner, J. Independent Evolution of the Core Domain and Its Flanking Sequences in Small Heat Shock Proteins. FASEB J. 2010, 24, 3633–3642. [Google Scholar] [CrossRef] [PubMed]
- Avelange-Macherel, M.H.; Rolland, A.; Hinault, M.P.; Tolleter, D.; Macherel, D. The Mitochondrial Small Heat Shock Protein HSP22 from Pea Is a Thermosoluble Chaperone Prone to Co-Precipitate with Unfolding Client Proteins. Int. J. Mol. Sci. 2020, 21, 97. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, G.M.S.; Ramos, C.H.I. Initial Characterization of Newly Identified Mitochondrial and Chloroplast Small HSPs from Sugarcane Shows That These Chaperones Have Different Oligomerization States and Substrate Specificities. Plant Physiol. Biochem. 2018, 129, 285–294. [Google Scholar] [CrossRef]
- Van Montfort, R.L.M.M.; Basha, E.; Friedrich, K.L.; Slingsby, C.; Vierling, E. Crystal Structure and Assembly of a Eukaryotic Small Heat Shock Protein. Nat. Struct. Biol. 2001, 8, 1025–1030. [Google Scholar] [CrossRef]
- Santhanagopalan, I.; Degiacomi, M.T.; Shepherd, D.A.; Hochberg, G.K.A.; Benesch, J.L.P.; Vierling, E. It Takes a Dimer to Tango: Oligomeric Small Heat Shock Proteins Dissociate to Capture Substrate. J. Biol. Chem. 2018, 293, 19511–19521. [Google Scholar] [CrossRef] [Green Version]
- Stengel, F.; Baldwin, A.J.; Painter, A.J.; Jaya, N.; Basha, E.; Kay, L.E.; Vierling, E.; Robinson, C.V.; Benesch, J.L.P. Quaternary Dynamics and Plasticity Underlie Small Heat Shock Protein Chaperone Function. Proc. Natl. Acad. Sci. USA 2010, 107, 2007–2012. [Google Scholar] [CrossRef] [Green Version]
- Shashidharamurthy, R.; Koteiche, H.A.; Dong, J.; Mchaourab, H.S. Mechanism of Chaperone Function in Small Heat Shock Proteins: Dissociation of the HSP27 Oligomer Is Required for Recognition and Binding of Destabilized T4 Lysozyme. J. Biol. Chem. 2005, 280, 5281–5289. [Google Scholar] [CrossRef]
- Hilario, E.; Martin, F.J.M.; Bertolini, M.C.; Fan, L.; Javier, F.; Martin, M.; Bertolini, M.C.; Fan, L. Crystal Structures of Xanthomonas Small Heat Shock Protein Provide a Structural Basis for an Active Molecular Chaperone Oligomer. J. Mol. Biol. 2011, 408, 74–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, R.D.; Chidawanyika, T.; Tims, H.S.; Meulia, T.; Bouchard, R.A.; Pett, V.B. Chaperone Function of Two Small Heat Shock Proteins from Maize. Plant Sci. 2014, 221–222, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, A.; Miranda-Ríos, J.; Navarrete, G.E.; Quinto, C.; Olivares, J.E.; García-Ponce, B.; Sánchez, F. Nodulin 22 from Phaseolus Vulgaris Protects Escherichia Coli Cells from Oxidative Stress. Planta 2004, 219, 993–1002. [Google Scholar] [CrossRef]
- Rodríguez-López, J.; Martínez-Centeno, C.; Padmanaban, A.; Guillén, G.; Olivares, J.E.; Stefano, G.; Lledías, F.; Ramos, F.; Ghabrial, S.A.; Brandizzi, F.; et al. Nodulin 22, a Novel Small Heat-Shock Protein of the Endoplasmic Reticulum, Is Linked to the Unfolded Protein Response in Common Bean. Mol. Plant. Microbe Interact. 2014, 27, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-López, J.; López, A.H.; Estrada-Navarrete, G.; Sánchez, F.; Díaz-Camino, C. The Noncanonical Heat Shock Protein PvNOD22 Is Essential for Infection Thread Progression during Rhizobial Endosymbiosis in Common Bean. Mol. Plant. Microbe Interact. 2019, 32, 939–948. [Google Scholar] [CrossRef]
- Sun, Y.; MacRae, T.H. The Small Heat Shock Proteins and Their Role in Human Disease. FEBS J. 2005, 272, 2613–2627. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Camino, C.; Annamalai, P.; Sanchez, F.; Kachroo, A.; Ghabrial, S.A. An Effective Virus-Based Gene Silencing Method for Functional Genomics Studies in Common Bean. Plant Methods 2011, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Bakthisaran, R.; Tangirala, R.; Rao, C.M. Small Heat Shock Proteins: Role in Cellular Functions and Pathology. Biochim. Biophys. Acta-Proteins Proteom. 2015, 1854, 291–319. [Google Scholar] [CrossRef] [Green Version]
- Van Montfort, R.; Slingsby, C.; Vierling, E. Structure and Function of the Small Heat Shock Protein/Alpha-Crystallin Family of Molecular Chaperones. Adv. Protein Chem. 2001, 59, 105–156. [Google Scholar] [CrossRef]
- Aquilina, J.A.; Benesch, J.L.P.; Bateman, O.A.; Slingsby, C.; Robinson, C.V. Polydispersity of a Mammalian Chaperone: Mass Spectrometry Reveals the Population of Oligomers in AB-Crystallin. Proc. Natl. Acad. Sci. USA 2003, 100, 10611–10616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.M.; Li, J.; He, J.; Liu, H.; Zhang, H.M. A Class I Cytosolic HSP20 of Rice Enhances Heat and Salt Tolerance in Different Organisms. Sci. Rep. 2020, 10, 1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haslbeck, M.; Kastenmüller, A.; Buchner, J.; Weinkauf, S.; Braun, N. Structural Dynamics of Archaeal Small Heat Shock Proteins. J. Mol. Biol. 2008, 378, 362–374. [Google Scholar] [CrossRef]
- Zhang, K.; Ezemaduka, A.N.; Wang, Z.; Hu, H.; Shi, X.; Liu, C.; Lu, X.; Fu, X.; Chang, Z.; Yin, C.C. A Novel Mechanism for Small Heat Shock Proteins to Function as Molecular Chaperones. Sci. Rep. 2015, 5, 8811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernova, L.S.; Bogachev, M.I.; Chasov, V.V.; Vishnyakov, I.E.; Kayumov, A.R. N- And C-Terminal Regions of the Small Heat Shock Protein IbpA from: Acholeplasma Laidlawii Competitively Govern Its Oligomerization Pattern and Chaperone-like Activity. RSC Adv. 2020, 10, 8364–8376. [Google Scholar] [CrossRef] [Green Version]
- The UniProt Consortium; Bateman, A.; Martin, M.-J.; Orchard, S.; Magrane, M.; Agivetova, R.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; et al. UniProt: The Universal Protein Knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Pundir, S.; Martin, M.J.; O’Donovan, C. UniProt Tools. Curr. Protoc. Bioinforma. 2016, 53, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Buchan, D.W.A.; Jones, D.T. The PSIPRED Protein Analysis Workbench: 20 Years On. Nucleic Acids Res. 2019, 47, W402–W407. [Google Scholar] [CrossRef] [Green Version]
- Dosztányi, Z.; Csizmok, V.; Tompa, P.; Simon, I. IUPred: Web Server for the Prediction of Intrinsically Unstructured Regions of Proteins Based on Estimated Energy Content. Bioinformatics 2005, 21, 3433–3434. [Google Scholar] [CrossRef] [Green Version]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Cassarino, T.G.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling Protein Tertiary and Quaternary Structure Using Evolutionary Information. Nucleic Acids Res. 2014, 42, 252–258. [Google Scholar] [CrossRef]
- Gabrielson, J.P.; Randolph, T.W.; Kendrick, B.S.; Stoner, M.R. Sedimentation Velocity Analytical Ultracentrifugation and SEDFIT/c(s): Limits of Quantitation for a Monoclonal Antibody System. Anal. Biochem. 2007, 361, 24–30. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Silva, A.; Lledías, F.; Rodríguez-López, J.; Olivares, J.E.; French-Pacheco, L.; Treviño, M.; Amero, C.; Díaz-Camino, C. The Common Bean Small Heat Shock Protein Nodulin 22 from Phaseolus vulgaris L. Assembles into Functional High-Molecular-Weight Oligomers. Molecules 2022, 27, 8681. https://doi.org/10.3390/molecules27248681
Fernández-Silva A, Lledías F, Rodríguez-López J, Olivares JE, French-Pacheco L, Treviño M, Amero C, Díaz-Camino C. The Common Bean Small Heat Shock Protein Nodulin 22 from Phaseolus vulgaris L. Assembles into Functional High-Molecular-Weight Oligomers. Molecules. 2022; 27(24):8681. https://doi.org/10.3390/molecules27248681
Chicago/Turabian StyleFernández-Silva, Arline, Fernando Lledías, Jonathan Rodríguez-López, Juan E. Olivares, Leidys French-Pacheco, Marcela Treviño, Carlos Amero, and Claudia Díaz-Camino. 2022. "The Common Bean Small Heat Shock Protein Nodulin 22 from Phaseolus vulgaris L. Assembles into Functional High-Molecular-Weight Oligomers" Molecules 27, no. 24: 8681. https://doi.org/10.3390/molecules27248681