
Structural Prediction and Characterization of Canavalia grandiflora (ConGF) Lectin Complexed with MMP1: Unveiling the Antiglioma Potential of Legume Lectins
 ,
,  , ,
, ,
Abstract
:1. Introduction
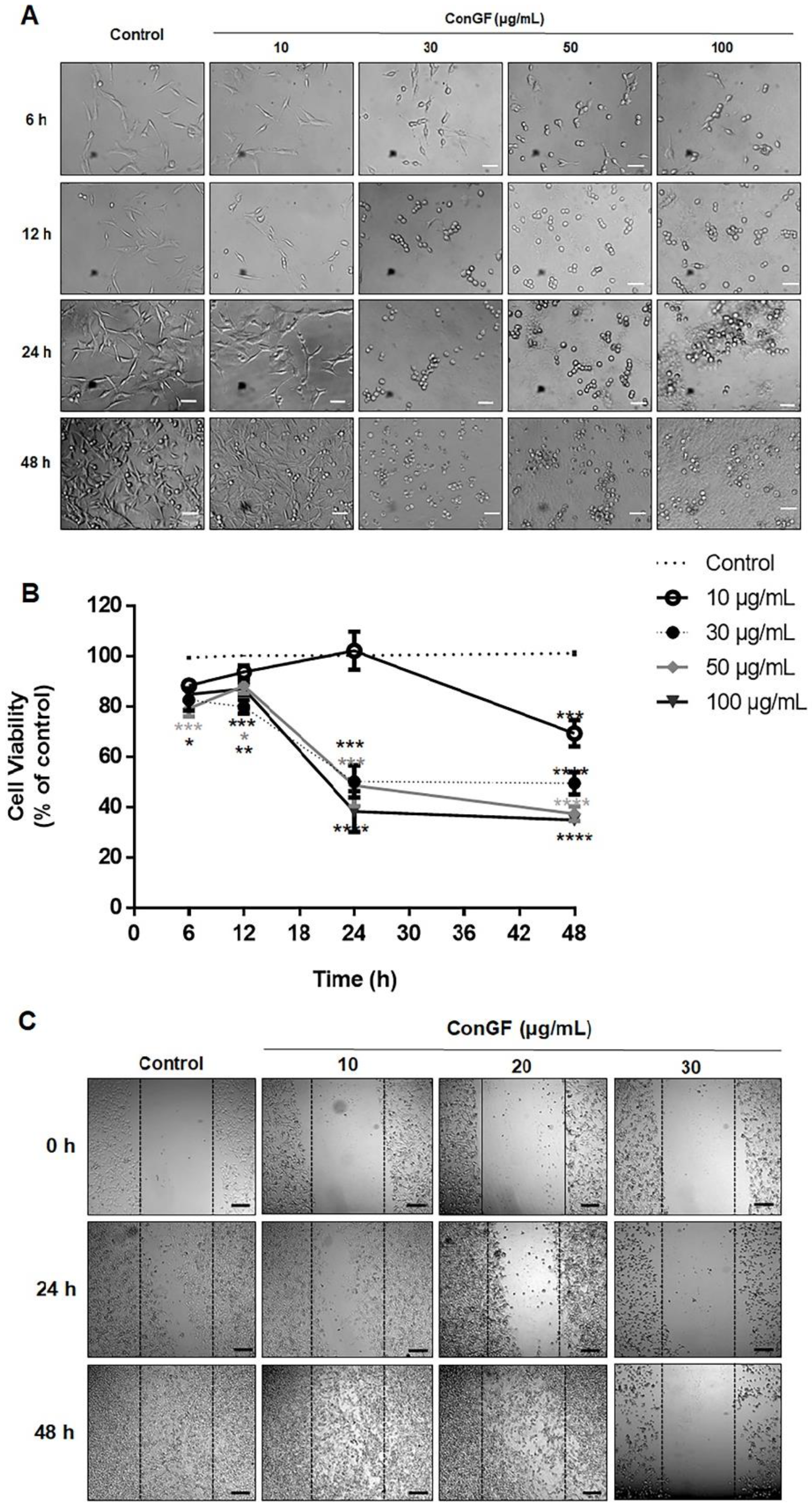
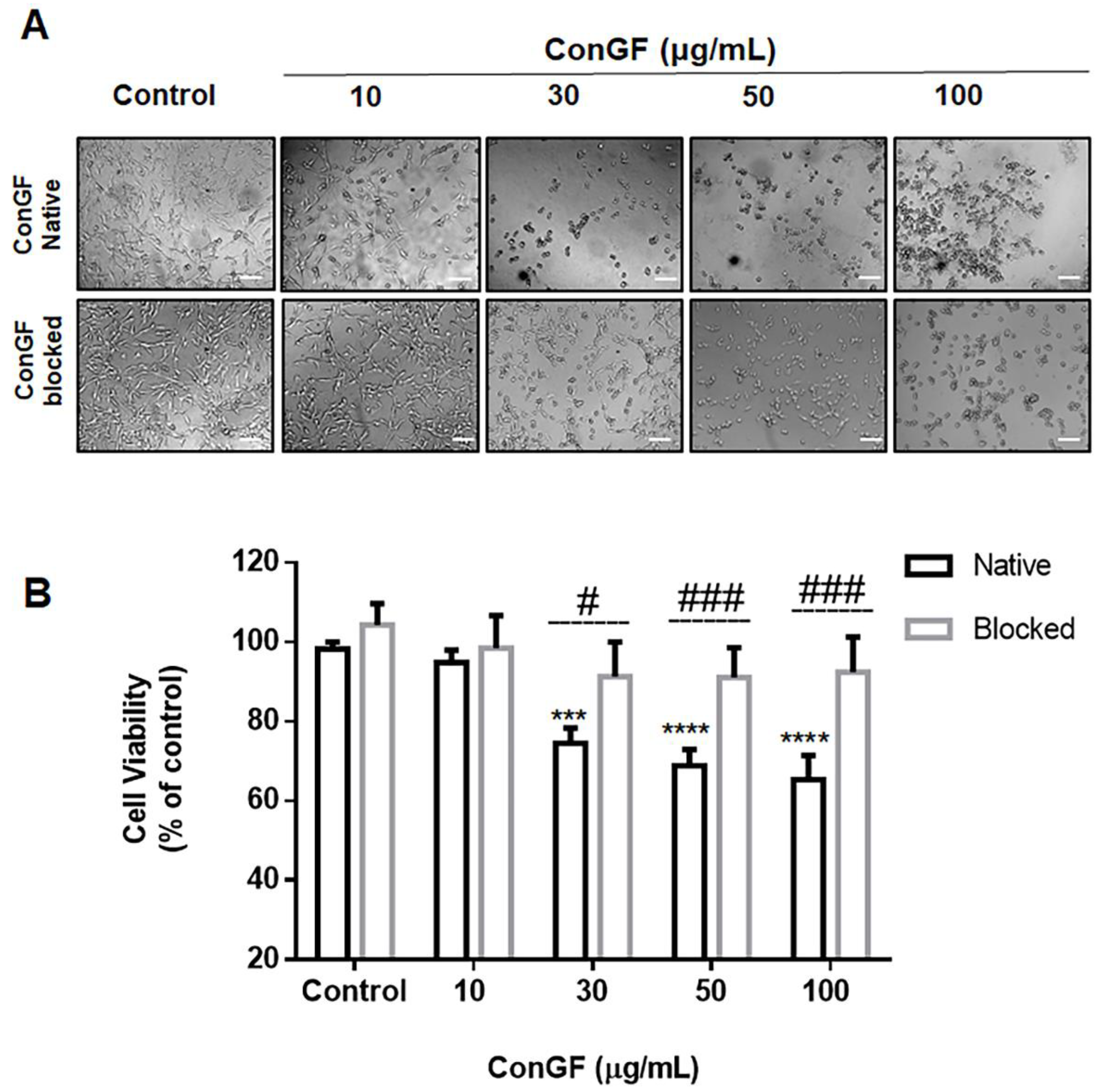
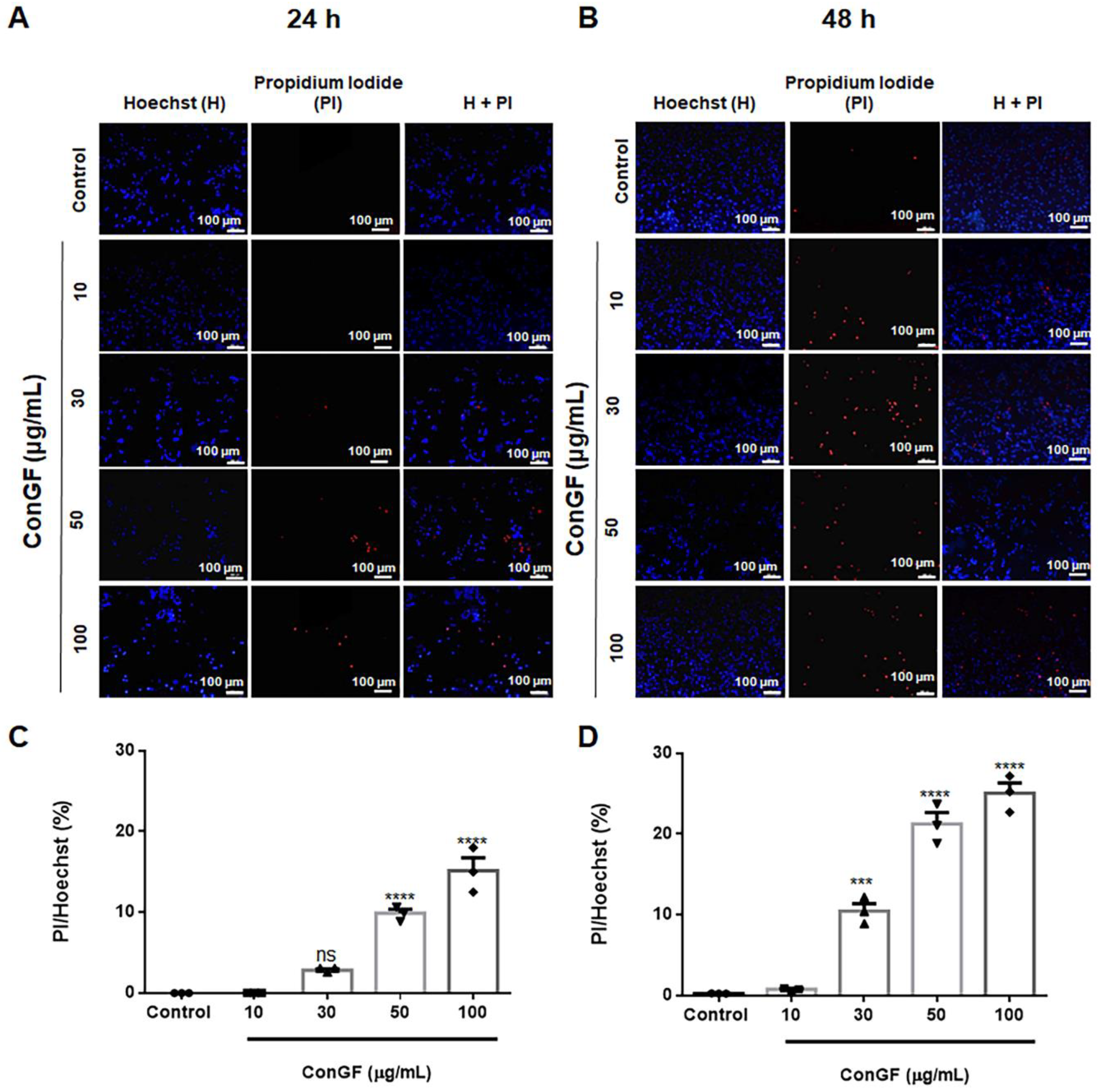
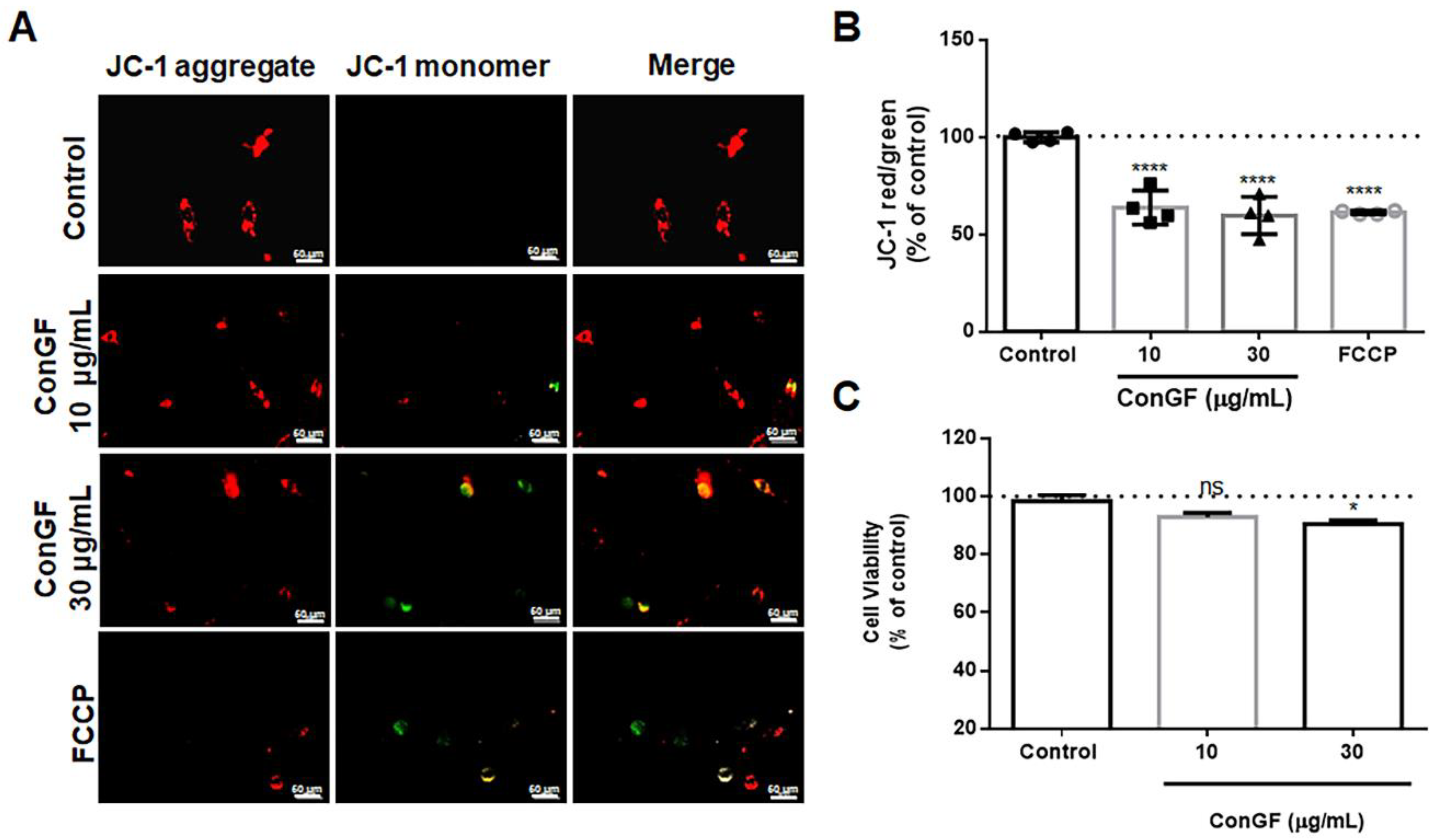
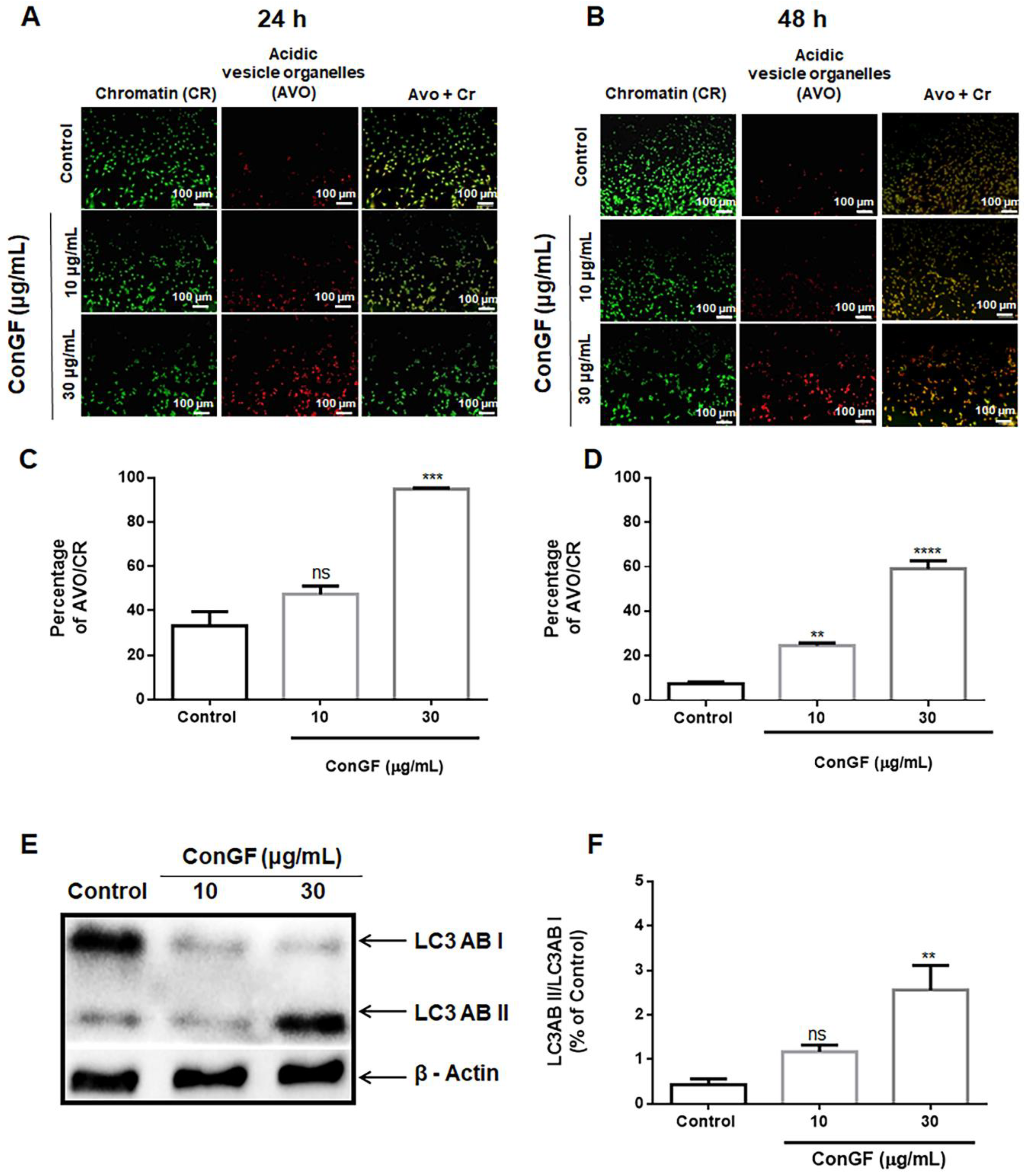
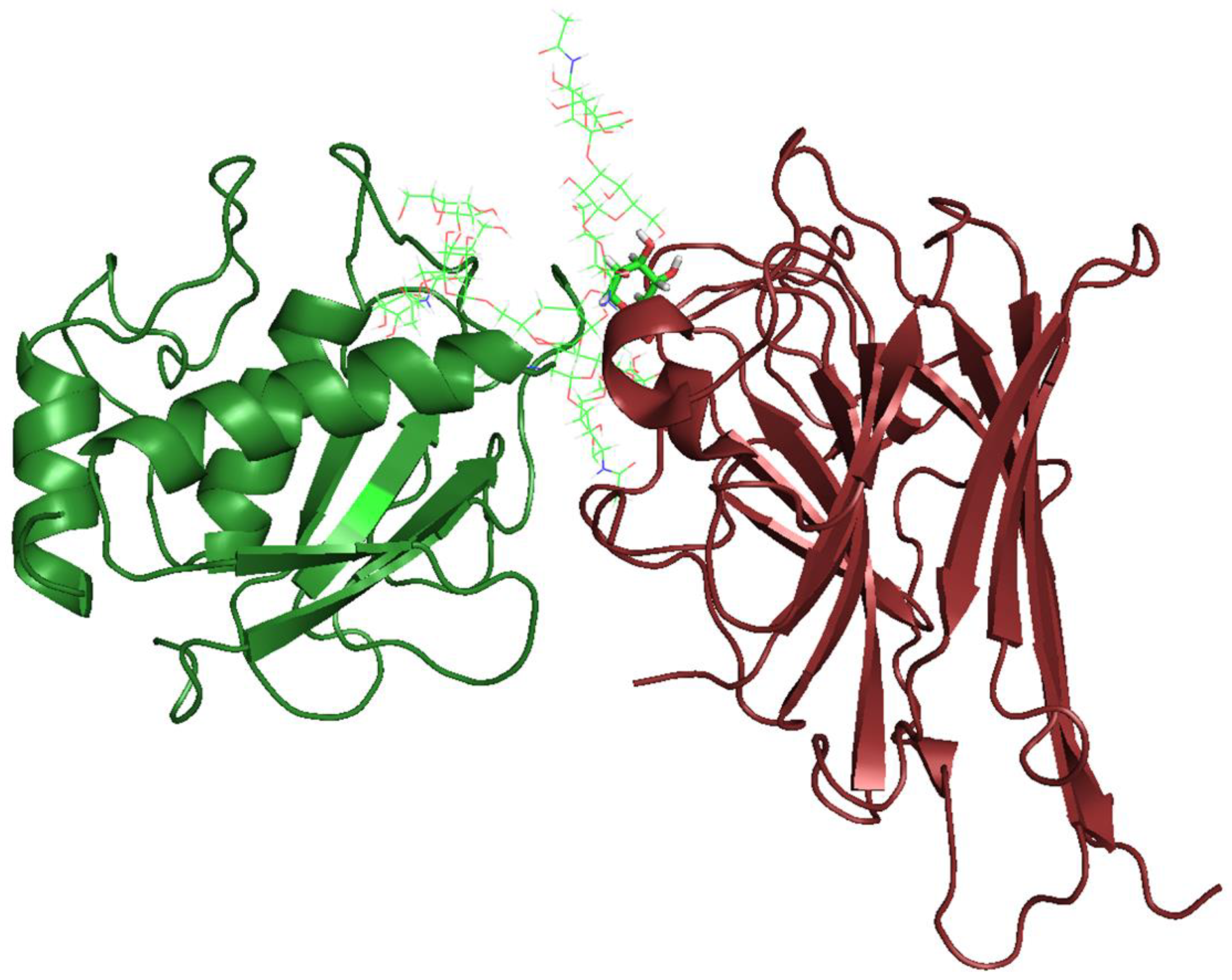
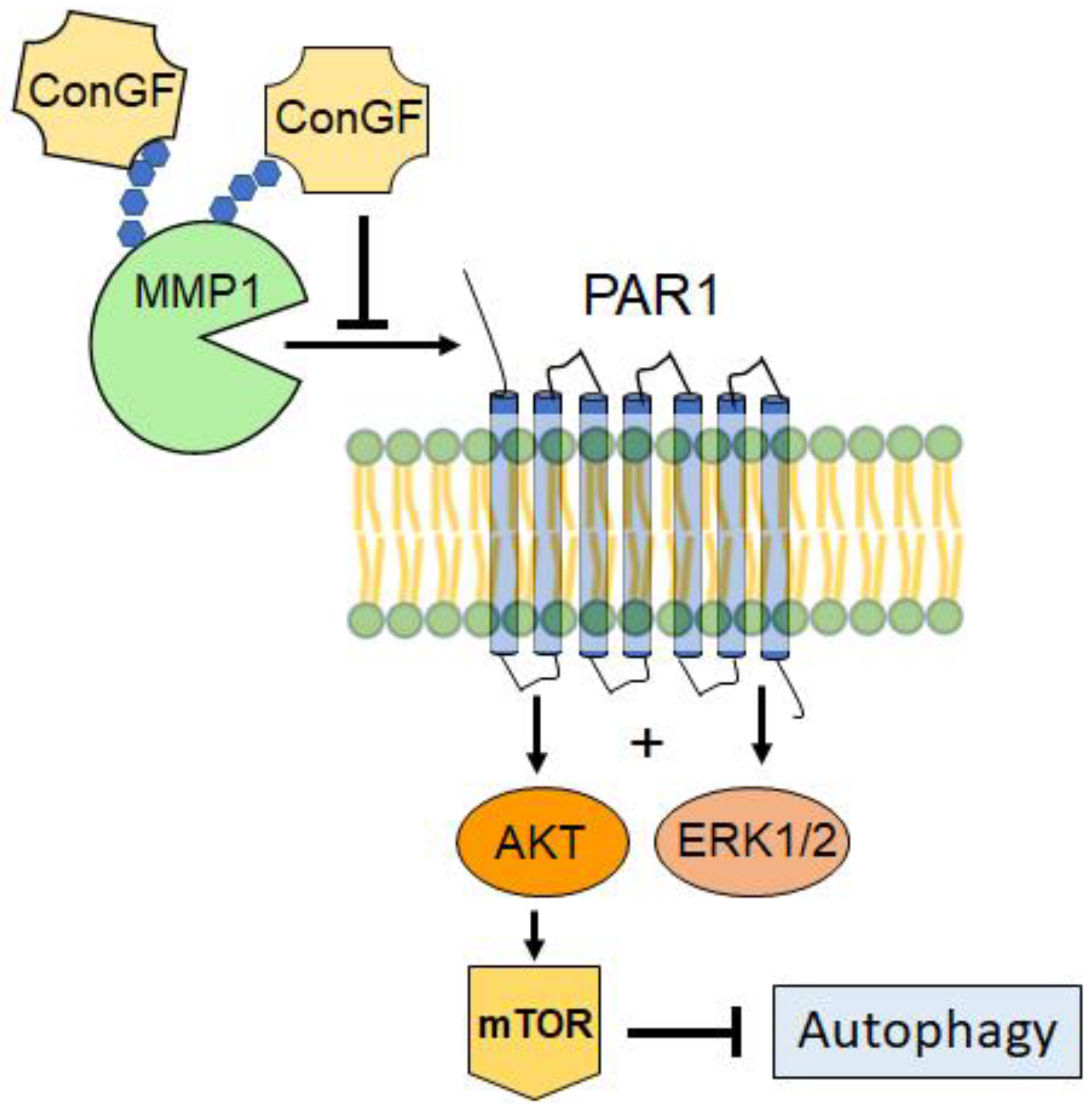
2. Results and Discussion
3. Methodology
3.1. Purification of Lectin ConGF
3.2. Cell Culture of Rat Glioblastoma C6 and Treatment
3.3. MTT Assay
3.4. Light Microscopy
3.5. Cell Migration (Scratch Assay)
3.6. JC-1 Assay
3.7. Propidium Iodide (PI) Staining
3.8. Acridine Orange Staining of Acidic Vesicular Organelle
3.9. Western Blot
3.10. Molecular Docking
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Horbinski, C.; Berger, T.; Packer, R.J.; Wen, P.Y. Clinical Implications of the 2021 Edition of the WHO Classification of Central Nervous System Tumours. Nat. Rev. Neurol. 2022, 18, 515–529. [Google Scholar] [CrossRef] [PubMed]
- Tucker-Burden, C.; Chappa, P.; Krishnamoorthy, M.; Gerwe, B.A.; Scharer, C.D.; Heimburg-Molinaro, J.; Harris, W.; Usta, S.N.; Eilertson, C.D.; Hadjipanayis, C.G.; et al. Lectins Identify Glycan Biomarkers on Glioblastoma-Derived Cancer Stem Cells. Stem Cells Dev. 2012, 21, 2374–2386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A Summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guidotti, G.; Brambilla, L.; Rossi, D. Exploring Novel Molecular Targets for the Treatment of High-Grade Astrocytomas Using Peptide Therapeutics: An Overview. Cells 2020, 9, 490. [Google Scholar] [CrossRef] [Green Version]
- Mazalovska, M.; Kouokam, J.C. Plant-Derived Lectins as Potential Cancer Therapeutics and Diagnostic Tools. Biomed. Res. Int. 2020, 2020, 1631394. [Google Scholar] [CrossRef] [PubMed]
- De Mejía, E.G.; Prisecaru, V.I. Lectins as Bioactive Plant Proteins: A Potential in Cancer Treatment. Crit. Rev. Food Sci. Nutr. 2005, 45, 425–445. [Google Scholar] [CrossRef] [PubMed]
- Yau, T.; Dan, X.; Ng, C.C.W.; Ng, T.B. Lectins with Potential for Anti-Cancer Therapy. Molecules 2015, 20, 3791–3810. [Google Scholar] [CrossRef] [Green Version]
- Cavada, B.S.; Pinto-Junior, V.R.; Osterne, V.J.S.; Nascimento, K.S. ConA-Like Lectins: High Similarity Proteins as Models to Study Structure/Biological Activities Relationships. Int. J. Mol. Sci. 2018, 20, 30. [Google Scholar] [CrossRef] [Green Version]
- Pratt, J.; Annabi, B. Induction of Autophagy Biomarker BNIP3 Requires a JAK2/STAT3 and MT1-MMP Signaling Interplay in Concanavalin-A-Activated U87 Glioblastoma Cells. Cell. Signal. 2014, 26, 917–924. [Google Scholar] [CrossRef]
- Nascimento, A.P.M.; Knaut, J.L.; Rieger, D.K.; Wolin, I.A.V.; Heinrich, I.A.; Mann, J.; Juarez, A.V.; del Sosa, L.V.; De Paul, A.L.; Moreira, C.G.; et al. Anti-Glioma Properties of DVL, a Lectin Purified from Dioclea Violacea. Int. J. Biol. Macromol. 2018, 120, 566–577. [Google Scholar] [CrossRef]
- Nascimento, A.P.M.; Wolin, I.A.V.; Welter, P.G.; Heinrich, I.A.; Zanotto-Filho, A.; Osterne, V.J.S.; Lossio, C.F.; Silva, M.T.L.; Nascimento, K.S.; Cavada, B.S.; et al. Lectin from Dioclea Violacea Induces Autophagy in U87 Glioma Cells. Int. J. Biol. Macromol. 2019, 134, 660–672. [Google Scholar] [CrossRef] [PubMed]
- Wolin, I.A.V.; Heinrich, I.A.; Nascimento, A.P.M.; Welter, P.G.; Sosa, L.D.V.; De Paul, A.L.; Zanotto-Filho, A.; Nedel, C.B.; Lima, L.D.; Osterne, V.J.S.; et al. ConBr Lectin Modulates MAPKs and Akt Pathways and Triggers Autophagic Glioma Cell Death by a Mechanism Dependent upon Caspase-8 Activation. Biochimie 2021, 180, 186–204. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Fabián, S.; Arreola, R.; Becerril-Villanueva, E.; Torres-Romero, J.C.; Arana-Argáez, V.; Lara-Riegos, J.; Ramírez-Camacho, M.A.; Alvarez-Sánchez, M.E. Role of Matrix Metalloproteinases in Angiogenesis and Cancer. Front. Oncol. 2019, 9, 1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sounni, N.E.; Devy, L.; Hajitou, A.; Frankenne, F.; Munaut, C.; Gilles, C.; Deroanne, C.; Thompson, E.W.; Foidart, J.M.; Noel, A. MT1-MMP Expression Promotes Tumor Growth and Angiogenesis through an up-Regulation of Vascular Endothelial Growth Factor Expression. FASEB J. 2002, 16, 555–564. [Google Scholar] [CrossRef] [Green Version]
- Barroso-Neto, I.L.; Simões, R.C.; Rocha, B.A.M.; Bezerra, M.J.B.; Pereira-Junior, F.N.; Silva Osterne, V.J.; Nascimento, K.S.; Nagano, C.S.; Delatorre, P.; Pereira, M.G.; et al. Vasorelaxant Activity of Canavalia Grandiflora Seed Lectin: A Structural Analysis. Arch. Biochem. Biophys. 2014, 543, 31–39. [Google Scholar] [CrossRef]
- Nunes, B.S.; Rensonnet, N.S.; Dal-Secco, D.; Vieira, S.M.; Cavada, B.S.; Teixeira, E.H.; Moura, T.R.; Teixeira, C.S.; Clemente-Napimoga, J.T.; Cunha, F.Q.; et al. Lectin Extracted from Canavalia Grandiflora Seeds Presents Potential Anti-Inflammatory and Analgesic Effects. Naunyn Schmiedebergs Arch. Pharmacol. 2009, 379, 609–616. [Google Scholar] [CrossRef]
- Ly, J.D.; Grubb, D.R.; Lawen, A. The Mitochondrial Membrane Potential (deltapsi(m)) in Apoptosis: An Update. Apoptosis 2003, 8, 115–128. [Google Scholar] [CrossRef]
- Thomé, M.P.; Filippi-Chiela, E.C.; Villodre, E.S.; Migliavaca, C.B.; Onzi, G.R.; Felipe, K.B.; Lenz, G. Ratiometric Analysis of Acridine Orange Staining in the Study of Acidic Organelles and Autophagy. J. Cell Sci. 2016, 129, 4622–4632. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-P.; Yang, M.-C.; Liu, H.-S.; Lin, Y.-S.; Lei, H.-Y. Concanavalin A Induces Autophagy in Hepatoma Cells and Has a Therapeutic Effect in a Murine in Situ Hepatoma Model. Hepatology 2007, 45, 286–296. [Google Scholar] [CrossRef]
- Roy, B.; Pattanaik, A.K.; Das, J.; Bhutia, S.K.; Behera, B.; Singh, P.; Maiti, T.K. Role of PI3K/Akt/mTOR and MEK/ERK Pathway in Concanavalin A Induced Autophagy in HeLa Cells. Chem. Biol. Interact. 2014, 210, 96–102. [Google Scholar] [CrossRef]
- Bhutia, S.K.; Panda, P.K.; Sinha, N.; Praharaj, P.P.; Bhol, C.S.; Panigrahi, D.P.; Mahapatra, K.K.; Saha, S.; Patra, S.; Mishra, S.R.; et al. Plant Lectins in Cancer Therapeutics: Targeting Apoptosis and Autophagy-Dependent Cell Death. Pharmacol. Res. 2019, 144, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Leal, R.B.; Pinto-Junior, V.R.; Osterne, V.J.S.; Wolin, I.A.V.; Nascimento, A.P.M.; Neco, A.H.B.; Araripe, D.A.; Welter, P.G.; Neto, C.C.; Correia, J.L.A.; et al. Crystal Structure of DlyL, a Mannose-Specific Lectin from Dioclea Lasiophylla Mart. Ex Benth Seeds That Display Cytotoxic Effects against C6 Glioma Cells. Int. J. Biol. Macromol. 2018, 114, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, C.; Anacker, J.; Ernestus, R.-I.; Vince, G.H. A Complete Compilation of Matrix Metalloproteinase Expression in Human Malignant Gliomas. World J. Clin. Oncol. 2012, 3, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar]
- Stojic, J.; Hagemann, C.; Haas, S.; Herbold, C.; Kühnel, S.; Gerngras, S.; Roggendorf, W.; Roosen, K.; Vince, G.H. Expression of Matrix Metalloproteinases MMP-1, MMP-11 and MMP-19 Is Correlated with the WHO-Grading of Human Malignant Gliomas. Neurosci. Res. 2008, 60, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Kubota, T.; Kabuto, M.; Sato, K.; Kawano, H.; Hayakawa, T.; Okada, Y. Production of Matrix Metalloproteinases and Tissue Inhibitor of Metalloproteinases-1 by Human Brain Tumors. J. Neurosurg. 1994, 81, 69–77. [Google Scholar] [CrossRef]
- McCready, J.; Broaddus, W.C.; Sykes, V.; Fillmore, H.L. Association of a Single Nucleotide Polymorphism in the Matrix Metalloproteinase-1 Promoter with Glioblastoma. Int. J. Cancer 2005, 117, 781–785. [Google Scholar] [CrossRef]
- Ruhul Amin, A.R.M.; Oo, M.L.; Senga, T.; Suzuki, N.; Feng, G.-S.; Hamaguchi, M. SH2 Domain Containing Protein Tyrosine Phosphatase 2 Regulates Concanavalin A-Dependent Secretion and Activation of Matrix Metalloproteinase 2 via the Extracellular Signal-Regulated Kinase and p38 Pathways. Cancer Res. 2003, 63, 6334–6339. [Google Scholar]
- Gingras, D.; Pagé, M.; Annabi, B.; Béliveau, R. Rapid Activation of Matrix Metalloproteinase-2 by Glioma Cells Occurs through a Posttranslational MT1-MMP-Dependent Mechanism. Biochim. Biophys. Acta 2000, 1497, 341–350. [Google Scholar] [CrossRef] [Green Version]
- Sina, A.; Proulx-Bonneau, S.; Roy, A.; Poliquin, L.; Cao, J.; Annabi, B. The Lectin Concanavalin-A Signals MT1-MMP Catalytic Independent Induction of COX-2 through an IKKgamma/NF-kappaB-Dependent Pathway. J. Cell Commun. Signal. 2010, 4, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Pratt, J.; Roy, R.; Annabi, B. Concanavalin-A-Induced Autophagy Biomarkers Requires Membrane Type-1 Matrix Metalloproteinase Intracellular Signaling in Glioblastoma Cells. Glycobiology 2012, 22, 1245–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, M.; Van Meter, T.E.; Fillmore, H.L. Epidermal Growth Factor Induces Matrix Metalloproteinase-1 (MMP-1) Expression and Invasion in Glioma Cell Lines via the MAPK Pathway. J. Neurooncol. 2011, 104, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhan, H.; Xu, W.; Yuan, Z.; Lu, P.; Zhan, L.; Li, Q. Upregulation of Matrix Metalloproteinase-1 and Proteinase-Activated Receptor-1 Promotes the Progression of Human Gliomas. Pathol. Res. Pract. 2011, 207, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Saarinen, J.; Welgus, H.G.; Flizar, C.A.; Kalkkinen, N.; Helin, J. N-Glycan Structures of Matrix Metalloproteinase-1 Derived from Human Fibroblasts and from HT-1080 Fibrosarcoma Cells. Eur. J. Biochem. 1999, 259, 829–840. [Google Scholar] [CrossRef] [Green Version]
- Ghazarian, H.; Idoni, B.; Oppenheimer, S.B. A Glycobiology Review: Carbohydrates, Lectins and Implications in Cancer Therapeutics. Acta Histochem. 2011, 113, 236–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagae, M.; Yamaguchi, Y. Function and 3D Structure of the N-Glycans on Glycoproteins. Int. J. Mol. Sci. 2012, 13, 8398–8429. [Google Scholar] [CrossRef] [Green Version]
- Pinho, S.S.; Reis, C.A. Glycosylation in Cancer: Mechanisms and Clinical Implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef]
- Bieberich, E. Synthesis, Processing, and Function of N-Glycans in N-Glycoproteins. Adv. Neurobiol. 2014, 9, 47–70. [Google Scholar]
- Boon, L.; Ugarte-Berzal, E.; Vandooren, J.; Opdenakker, G. Glycosylation of Matrix Metalloproteases and Tissue Inhibitors: Present State, Challenges and Opportunities. Biochem. J. 2016, 473, 1471–1482. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, H.; Swoger, J.; Greene, S.; Saito, T.; Hurh, J.; Sweeley, C.; Leestma, J.; Mkrdichian, E.; Cerullo, L.; Nishikawa, A.; et al. Beta1,6-N-Acetylglucosamine-Bearing N-Glycans in Human Gliomas: Implications for a Role in Regulating Invasivity. Cancer Res. 2000, 60, 134–142. [Google Scholar]
- Stowell, S.R.; Ju, T.; Cummings, R.D. Protein Glycosylation in Cancer. Annu. Rev. Pathol. 2015, 10, 473–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pullen, N.A.; Anand, M.; Cooper, P.S.; Fillmore, H.L. Matrix Metalloproteinase-1 Expression Enhances Tumorigenicity as Well as Tumor-Related Angiogenesis and Is Inversely Associated with TIMP-4 Expression in a Model of Glioblastoma. J. Neurooncol. 2012, 106, 461–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Zhong, Z.; Yuan, J.; Zhang, Z.; Wei, Q.; Song, W.; Chen, H. Collaborative Overexpression of Matrix Metalloproteinase-1 and Vascular Endothelial Growth Factor-C Predicts Adverse Prognosis in Patients with Gliomas. Cancer Epidemiol. 2013, 37, 697–702. [Google Scholar] [CrossRef] [PubMed]
- Austin, K.M.; Covic, L.; Kuliopulos, A. Matrix Metalloproteases and PAR1 Activation. Blood 2013, 121, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Goerge, T.; Barg, A.; Schnaeker, E.-M.; Poppelmann, B.; Shpacovitch, V.; Rattenholl, A.; Maaser, C.; Luger, T.A.; Steinhoff, M.; Schneider, S.W. Tumor-Derived Matrix Metalloproteinase-1 Targets Endothelial Proteinase-Activated Receptor 1 Promoting Endothelial Cell Activation. Cancer Res. 2006, 66, 7766–7774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomko, N.; Kluever, M.; Wu, C.; Zhu, J.; Wang, Y.; Salomon, R.G. 4-Hydroxy-7-Oxo-5-Heptenoic Acid Lactone Is a Potent Inducer of Brain Cancer Cell Invasiveness That May Contribute to the Failure of Anti-Angiogenic Therapies. Free Radic. Biol. Med. 2020, 146, 234–256. [Google Scholar] [CrossRef]
- Bode, M.F.; Schmedes, C.M.; Egnatz, G.J.; Bharathi, V.; Hisada, Y.M.; Martinez, D.; Kawano, T.; Weithauser, A.; Rosenfeldt, L.; Rauch, U.; et al. Cell Type-Specific Roles of PAR1 in Coxsackievirus B3 Infection. Sci. Rep. 2021, 11, 14264. [Google Scholar] [CrossRef]
- Ceccatto, V.M.; Cavada, B.S.; Nunes, E.P.; Nogueira, N.A.P.; Grangeiro, M.B.; Moreno, F.B.M.B.; Teixeira, E.H.; Sampaio, A.H.; Alves, M.A.O.; Ramos, M.V.; et al. Purification and Partial Characterization of a Lectin from Canavalia Grandiflora Benth. Seeds. Protein Pept. Lett. 2002, 9, 67–73. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Liang, C.-C.; Park, A.Y.; Guan, J.-L. In Vitro Scratch Assay: A Convenient and Inexpensive Method for Analysis of Cell Migration in Vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [Green Version]
- De Ávila, M.B.; Xavier, M.M.; Pintro, V.O.; de Azevedo, W.F., Jr. Supervised Machine Learning Techniques to Predict Binding Affinity. A Study for Cyclin-Dependent Kinase 2. Biochem. Biophys. Res. Commun. 2017, 494, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and Validation of a Genetic Algorithm for Flexible Docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korb, O.; Stützle, T.; Exner, T.E. Empirical Scoring Functions for Advanced Protein-Ligand Docking with PLANTS. J. Chem. Inf. Model. 2009, 49, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Heck, G.S.; Pintro, V.O.; Pereira, R.R.; de Ávila, M.B.; Levin, N.M.B.; de Azevedo, W.F. Supervised Machine Learning Methods Applied to Predict Ligand- Binding Affinity. Curr. Med. Chem. 2017, 24, 2459–2470. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, K.N.; Yongye, A.B.; Tschampel, S.M.; González-Outeiriño, J.; Daniels, C.R.; Foley, B.L.; Woods, R.J. GLYCAM06: A Generalizable Biomolecular Force Field. Carbohydrates. J. Comput. Chem. 2008, 29, 622–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. PatchDock and SymmDock: Servers for Rigid and Symmetric Docking. Nucleic Acids Res. 2005, 33, W363–W367. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbohydrate | Score |
|---|---|
| Xman | −45.26 |
| MMP1 N-glycans in normal cells | |
| Glycan | Score |
| N001 | −64.29 |
| N002 | −55.18 |
| N003 | −51.11 |
| N004 | −58.72 |
| N005 | −63.28 |
| N006 | −52.70 |
| N007 | −50.25 |
| N008 | −64.64 |
| N009 | −64.68 |
| N010 | −49.00 |
| N011 | - |
| MMP1 N-glycans in malignant cells | |
| Glycan | Score |
| T001 | −50.62 |
| T002 | −63.96 |
| T003 | −57.95 |
| T004 | - |
| T005 | −59.12 |
| T006 | −47.63 |
| T007 | −52.33 |
| T008 | −60.83 |
| T009 | −53.75 |
| T010 | −59.61 |
| T011 | −45.05 |
| T012 | −44.67 |
| T013 | - |
| T014 | −54.20 |
| T015 | −57.97 |
| T016 | −47.46 |
| T017 | −53.89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leal, R.B.; Mann, J.; Pinto-Junior, V.R.; Oliveira, M.V.; Osterne, V.J.S.; Wolin, I.A.V.; Nascimento, A.P.M.; Welter, P.G.; Ferreira, V.M.S.; Silva, A.A.; et al. Structural Prediction and Characterization of Canavalia grandiflora (ConGF) Lectin Complexed with MMP1: Unveiling the Antiglioma Potential of Legume Lectins. Molecules 2022, 27, 7089. https://doi.org/10.3390/molecules27207089
Leal RB, Mann J, Pinto-Junior VR, Oliveira MV, Osterne VJS, Wolin IAV, Nascimento APM, Welter PG, Ferreira VMS, Silva AA, et al. Structural Prediction and Characterization of Canavalia grandiflora (ConGF) Lectin Complexed with MMP1: Unveiling the Antiglioma Potential of Legume Lectins. Molecules. 2022; 27(20):7089. https://doi.org/10.3390/molecules27207089
Chicago/Turabian StyleLeal, Rodrigo Bainy, Josiane Mann, Vanir Reis Pinto-Junior, Messias Vital Oliveira, Vinicius Jose Silva Osterne, Ingrid Alessandra Victoria Wolin, Ana Paula Machado Nascimento, Priscilla Gomes Welter, Valeria Maria Sousa Ferreira, Alice Araujo Silva, and et al. 2022. "Structural Prediction and Characterization of Canavalia grandiflora (ConGF) Lectin Complexed with MMP1: Unveiling the Antiglioma Potential of Legume Lectins" Molecules 27, no. 20: 7089. https://doi.org/10.3390/molecules27207089