
Associations of Region and Lactation Stage with Odd-Chain Fatty Acid Profile in Triglycerides of Breast Milk in China
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
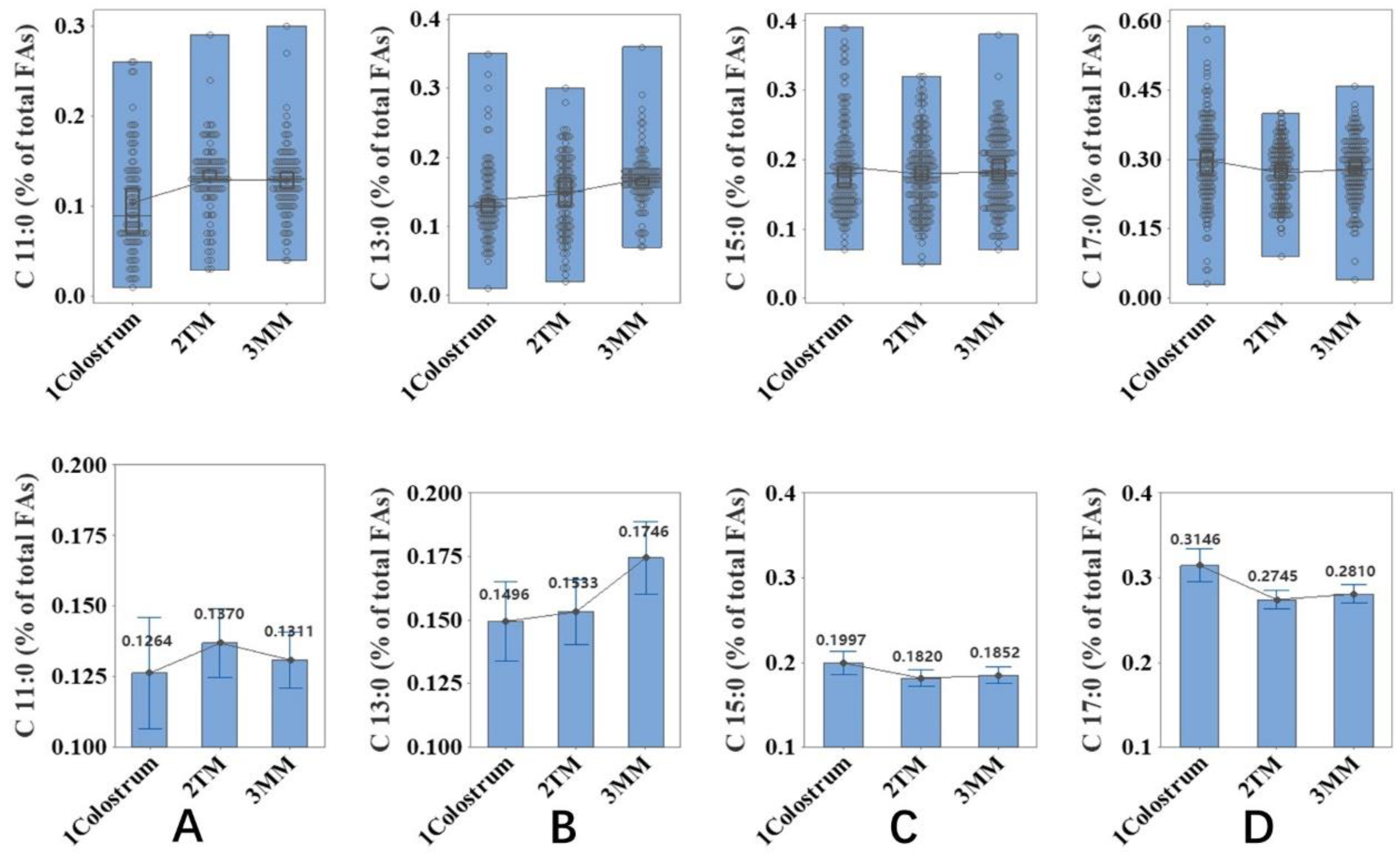
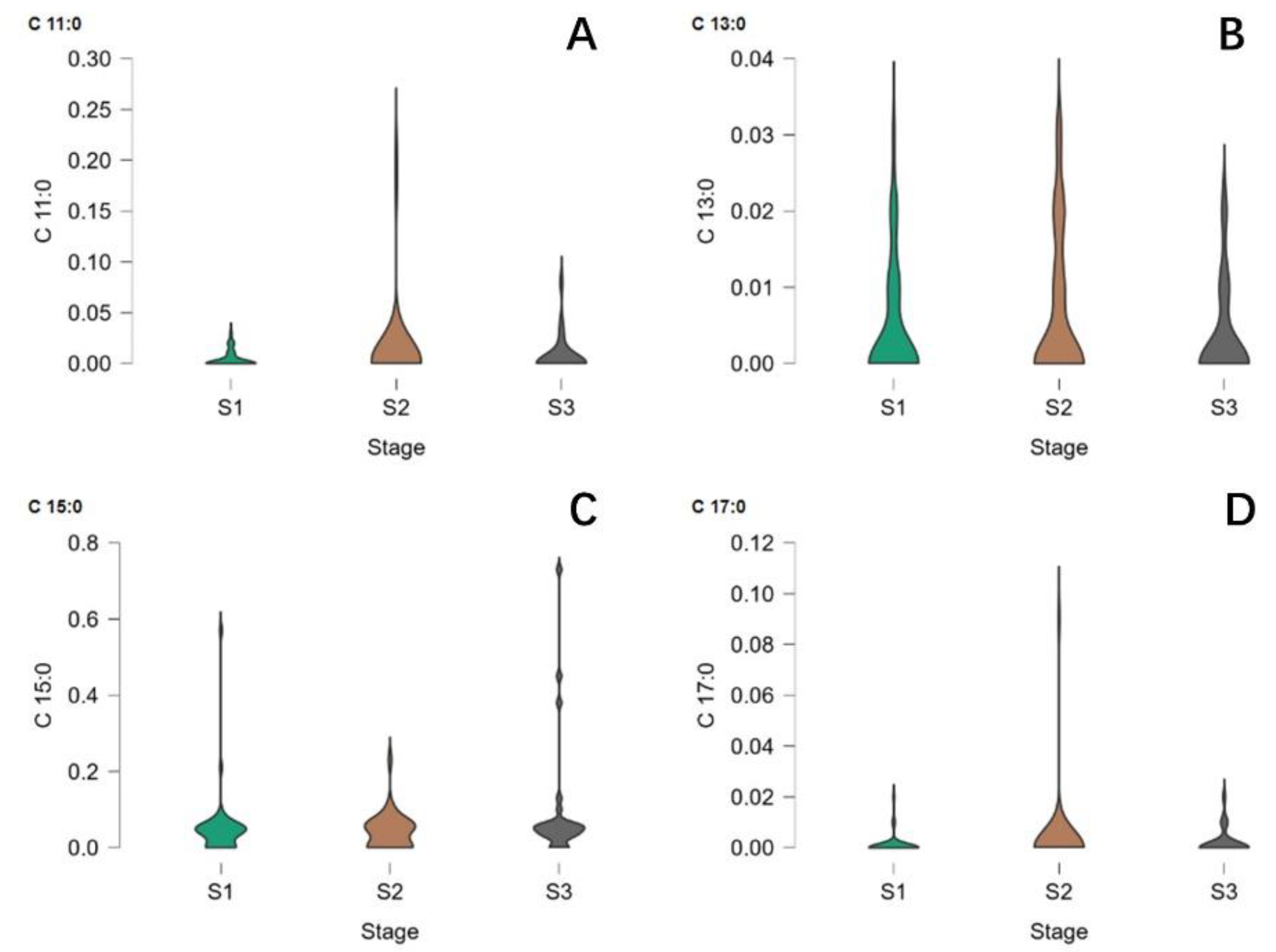
2.1. Associationof Lactation Stage with the Content of OCFAs
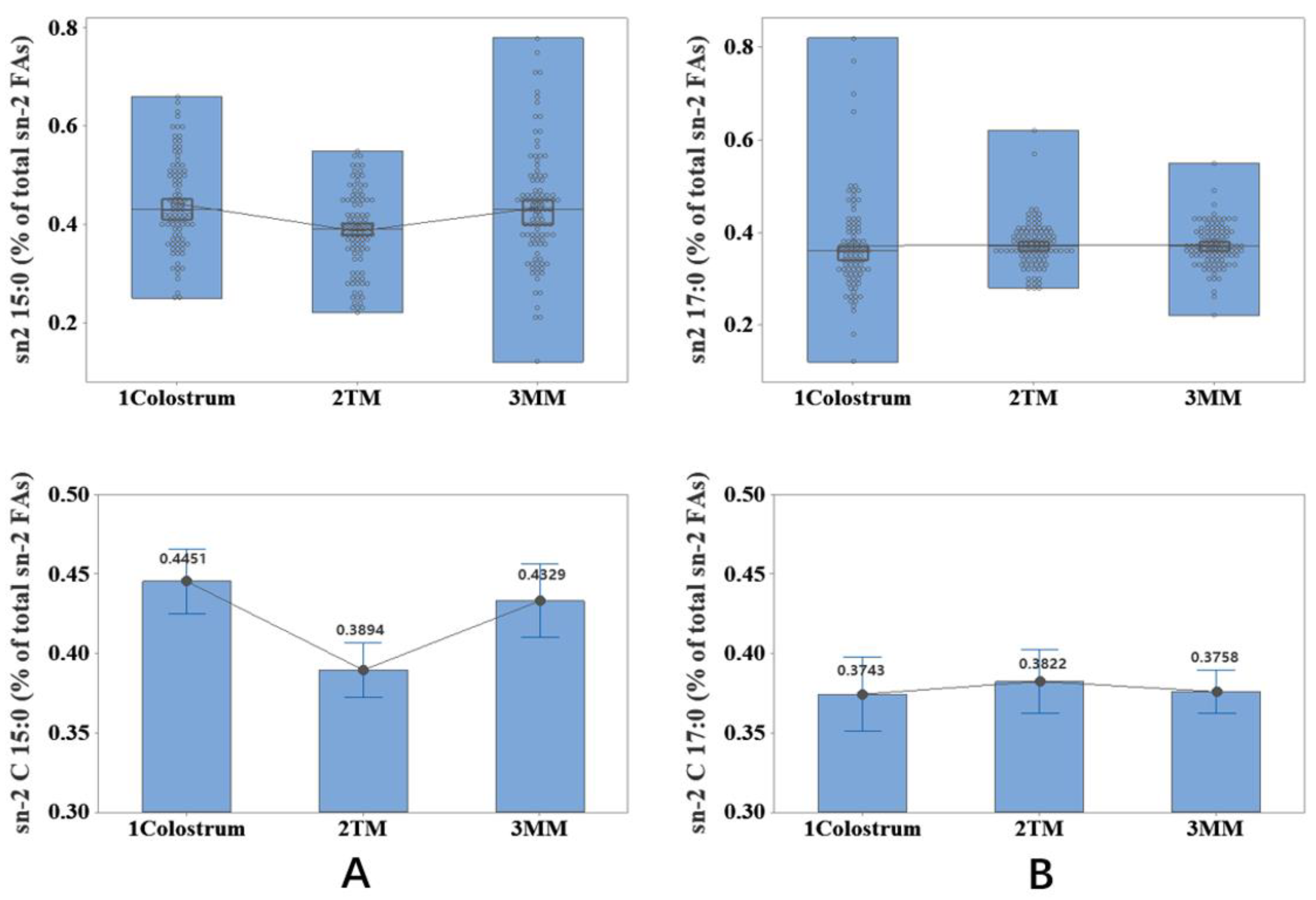
2.2. Association of Lactation Stage with the Content of sn-2 OCFAs
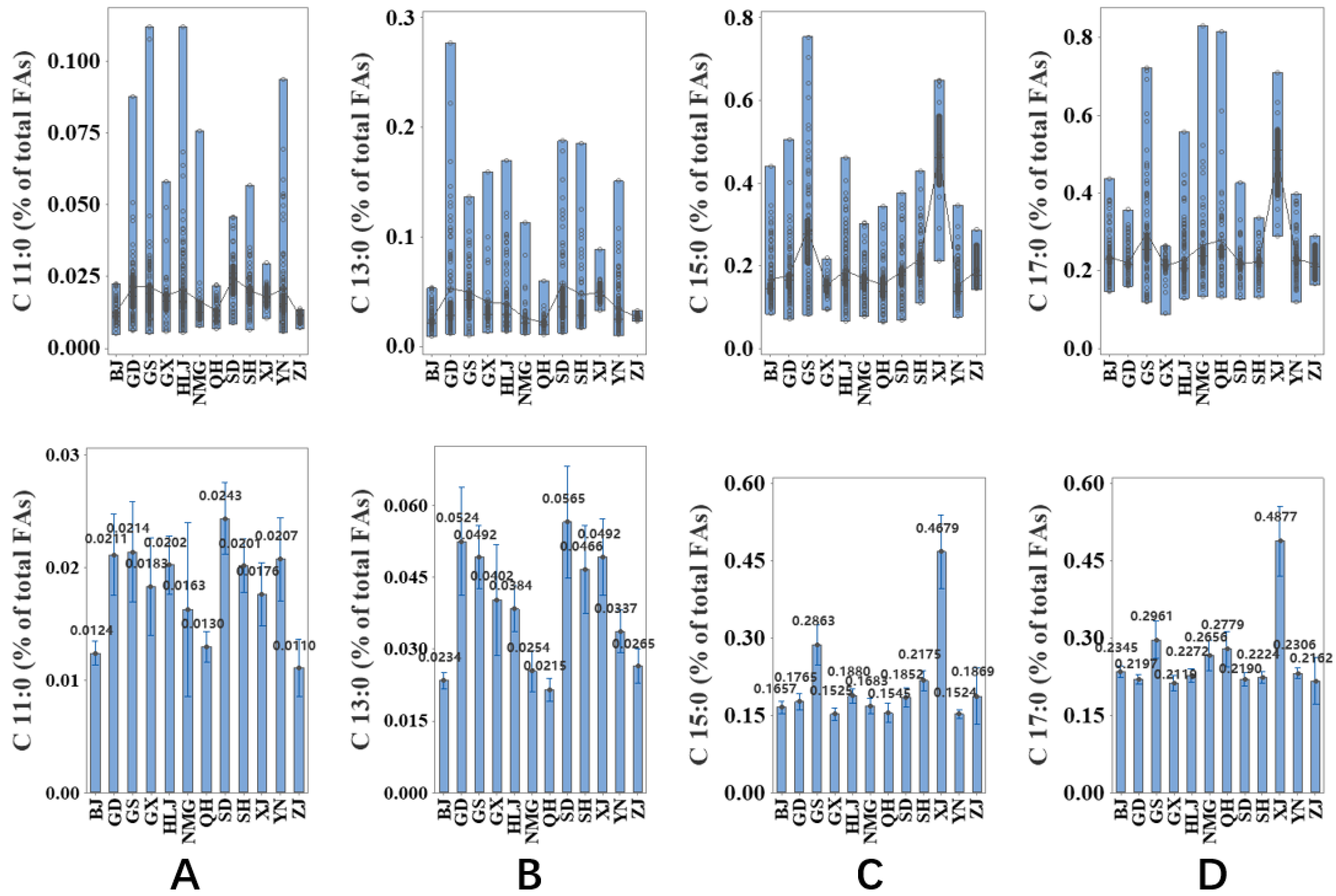
2.3. Associationof Milk Origins with the Content of OCFAs
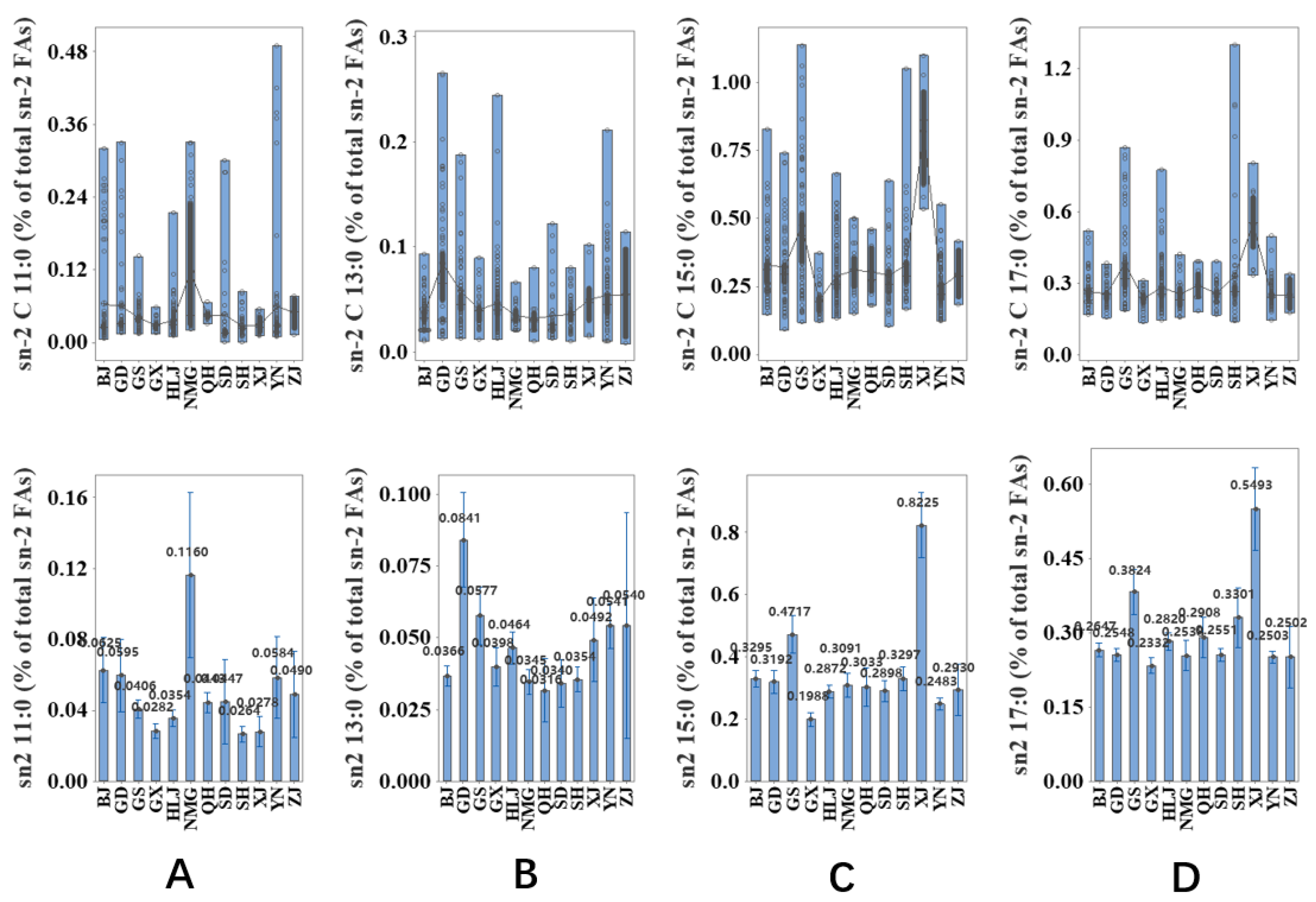
2.4. Associationof Milk Origins with the Content of sn-2 OCFAs
2.5. Analysis of the OCFAs in Infant Formula
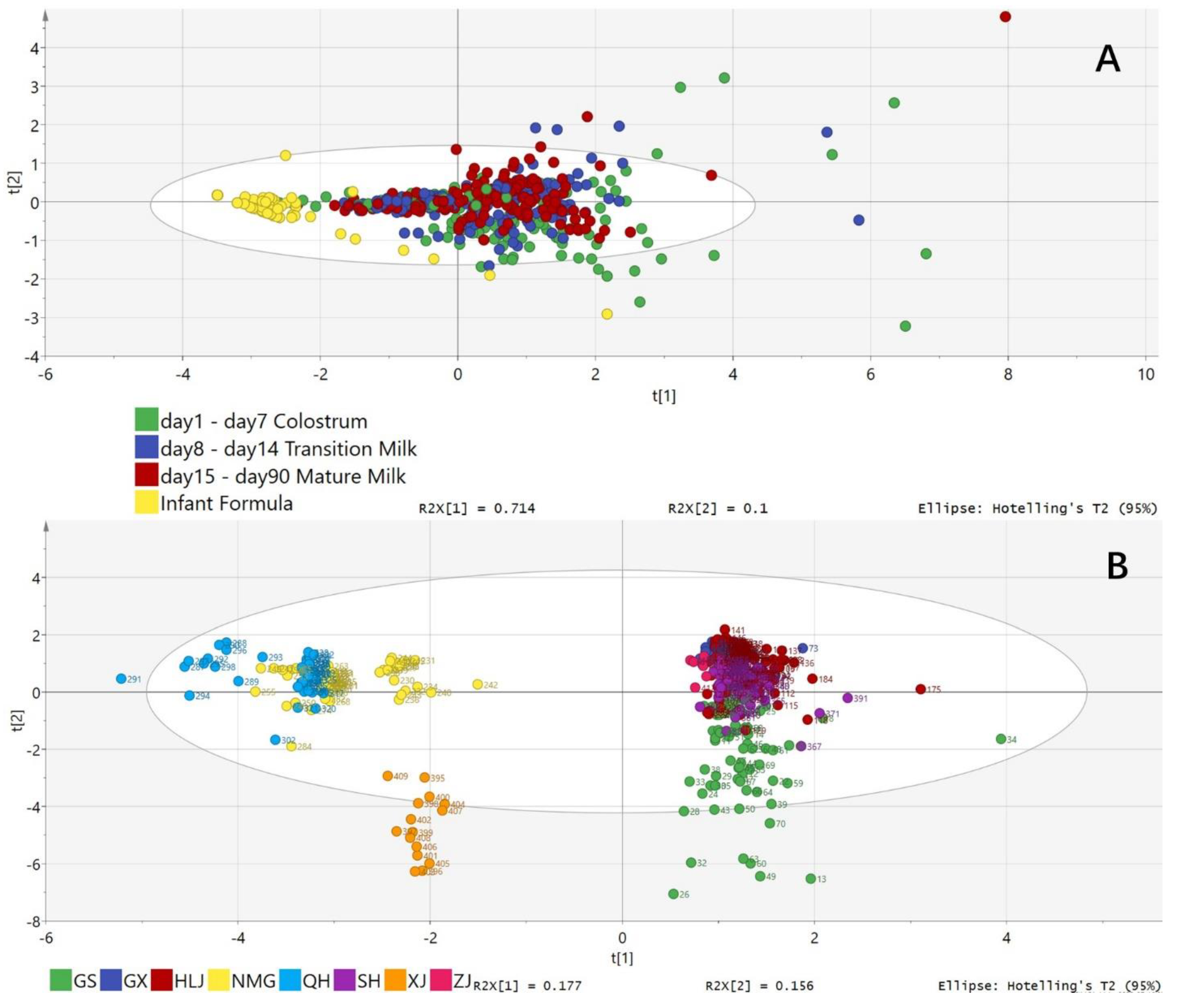
2.6. PDS-LA Analysis of the OCFA Profile
3. Discussion
4. Materials and Methods
4.1. Data Collection and Statements
4.1.1. Infant Formula Collection
4.1.2. Human Milk Sample Collection
4.2. Method for Data Collection
4.2.1. Total Lipid Extraction from Human Milk and Infant Formula Milk
4.2.2. Preparation of Fatty Acid Methyl Esters (FAMEs) for Chromatographic Analysis
4.2.3. Preparation of 2-Monoacylglycerol and its Methyl Esters
4.2.4. GC Analysis
4.3. Statistical Analysis Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Dai, X.; Yuan, T.; Zhang, X.; Zhou, Q.; Bi, H.; Yu, R.; Wei, W.; Wang, X. Short-chain fatty acid (SCFA) and medium-chain fatty acid (MCFA) concentrations in human milk consumed by infants born at different gestational ages and the variations in concentration during lactation stages. Food Funct. 2020, 11, 1869–1880. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Zhou, X.H.; Han, B.; Li, S.M.; Xu, T.; Yi, H.X.; Liu, P.; Zhang, L.W.; Li, Y.Y.; Jiang, S.L.; et al. Composition analysis of fatty acids and stereo-distribution of triglycerides in human milk from three regions of China. Food Res. Int. 2020, 133, 109196. [Google Scholar] [CrossRef] [PubMed]
- Hodson, L.; Eyles, H.C.; McLachlan, K.J.; Bell, M.L.; Green, T.J.; Skeaff, C.M. Plasma and Erythrocyte Fatty Acids Reflect Intakes of Saturated and n-6 PUFA within a Similar Time Frame. J. Nutr. 2014, 144, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Řezanka, T.; Sigler, K. Odd-numbered very-long-chain fatty acids from the microbial, animal and plant kingdoms. Prog. Lipid Res. 2009, 48, 206–238. [Google Scholar] [CrossRef] [PubMed]
- Imamura, F.; Fretts, A.; Marklund, M.; Ardisson Korat, A.V.; Yang, W.-S.; Lankinen, M.; Qureshi, W.; Helmer, C.; Chen, T.-A.; Wong, K.; et al. Fatty acid biomarkers of dairy fat consumption and incidence of type 2 diabetes: A pooled analysis of prospective cohort studies. PLoS Med. 2018, 15, e1002670. [Google Scholar] [CrossRef]
- Dąbrowski, G.; Konopka, I. Update on food sources and biological activity of odd-chain, branched and cyclic fatty acids—A review. Trends Food Sci. Technol. 2022, 119, 514–529. [Google Scholar] [CrossRef]
- Moltó-Puigmartí, C.; Castellote, A.I.; Carbonell-Estrany, X.; López-Sabater, M.C. Differences in fat content and fatty acid proportions among colostrum, transitional, and mature milk from women delivering very preterm, preterm, and term infants. Clin. Nutr. 2011, 30, 116–123. [Google Scholar] [CrossRef]
- Wu, K.; Gao, R.; Tian, F.; Mao, Y.; Wang, B.; Zhou, L.; Shen, L.; Guan, Y.; Cai, M. Fatty acid positional distribution (sn-2 fatty acids) and phospholipid composition in Chinese breast milk from colostrum to mature stage. Br. J. Nutr. 2019, 121, 65–73. [Google Scholar] [CrossRef]
- Qi, C.; Sun, J.; Xia, Y.; Yu, R.; Wei, W.; Xiang, J.; Jin, Q.; Xiao, H.; Wang, X. Fatty Acid Profile and the sn-2 Position Distribution in Triacylglycerols of Breast Milk during Different Lactation Stages. J. Agric. Food Chem. 2018, 66, 3118–3126. [Google Scholar] [CrossRef]
- Wang, J.; Liu, L.; Liu, L.; Sun, L.; Li, C. Absorption of 1,3-dioleyl-2-palmitoylglycerol and intestinal flora profiles changes in mice. Int. J. Food Sci. Nutr. 2020, 71, 296–306. [Google Scholar] [CrossRef]
- Jenkins, B.; West, J.A.; Koulman, A. A review of odd-chain fatty acid metabolism and the role of pentadecanoic acid (C15: 0) and heptadecanoic acid (C17:0) in health and disease. Molecules 2015, 20, 2425–2444. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.V.; Lee, B.-C.; Hsieh, T.C.-Y.; Laine, R.A. Occurrence of an unusual amount of an odd-numbered fatty acid in glycosphingolipids from human cataracts. Curr. Eye Res. 1987, 6, 1361–1367. [Google Scholar] [CrossRef] [PubMed]
- Yuhas, R.; Pramuk, K.; Lien, E.L. Human milk fatty acid composition from nine countries varies most in DHA. Lipids 2006, 41, 851–858. [Google Scholar] [CrossRef]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4578–4585. [Google Scholar] [CrossRef]
- Robert, K.Y.; Nakatani, Y.; Yanagisawa, M. The role of glycosphingolipid metabolism in the developing brain. J. Lipid Res. 2009, 50, S440–S445. [Google Scholar]
- Innis, S.M.; Dyer, R.; Nelson, C.M. Evidence that palmitic acid is absorbed as sn-2 monoacylglycerol from human milk by breast-fed infants. Lipids 1994, 29, 541–545. [Google Scholar] [CrossRef]
- Sun, W.; Luo, Y.; Wang, D.H.; Kothapalli, K.S.D.; Brenna, J.T. Branched chain fatty acid composition of yak milk and manure during full-lactation and half-lactation. Prostaglandins Leukot. Essent. Fat. Acids 2019, 150, 16–20. [Google Scholar] [CrossRef]
- Markiewicz-Kęszycka, M.; Czyżak-Runowska, G.; Lipińska, P.; Wójtowski, J. Fatty acid profile of milk-a review. Bull. Vet. Inst. Pulawy 2013, 57, 135–139. [Google Scholar] [CrossRef]
- Schwendel, B.H.; Morel, P.C.H.; Wester, T.J.; Tavendale, M.H.; Deadman, C.; Fong, B.; Shadbolt, N.M.; Thatcher, A.; Otter, D.E. Fatty acid profile differs between organic and conventionally produced cow milk independent of season or milking time. J. Dairy Sci. 2015, 98, 1411–1425. [Google Scholar] [CrossRef]
- Dong, S.; Long, R.; Kang, M. Milking and milk processing: Traditional technologies in the yak farming system of the Qinghai-Tibetan Plateau, China. Int. J. Dairy Technol. 2003, 56, 86–93. [Google Scholar] [CrossRef]
- Wallingford, J.C.; Yuhas, R.; Du, S.; Zhai, F.; Popkin, B.M. Fatty Acids in Chinese Edible Oils: Value of Direct Analysis as a Basis for Labeling. Food Nutr. Bull. 2004, 25, 330–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuming, L.; Fang, W.; Heigang, X.; Zhaoguo, W.; Baofu, L. A study on classification and zoning of Chinese geoheritage resources in national geoparks. Geoheritage 2016, 8, 247–261. [Google Scholar] [CrossRef]
- Junqin, M. The present situation and characteristics of contemporary Islam in China. Doshisha Univ. Cent. Interdiscip. Study Monoth. Relig. (CISMOR) 2013, 8, 26–36. [Google Scholar]
- Kornsteiner, M.; Singer, I.; Elmadfa, I. Very low n-3 long-chain polyunsaturated fatty acid status in Austrian vegetarians and vegans. Ann. Nutr. Metab. 2008, 52, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B.; Zetterström, R. Protein content of infant formula—How much and from what age? Acta Paediatr. 1988, 77, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Nadaoka, I.; Hatakeyama, E.; Tanada, C.; Sakamoto, T.; Fukaya, S.; Akiba, T.; Inoue, K.; Yamano, Y.; Toyo’oka, T. Quantitative determination of total L-carnitine in infant formula, follow-up formula, and raw materials by liquid chromatography with tandem mass spectrometry. Anal. Sci. 2014, 30, 839–844. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhang, H.; Karrar, E.; Wei, W.; Zeng, W.; Jin, Q.; Wang, X. Associations of Region and Lactation Stage with Odd-Chain Fatty Acid Profile in Triglycerides of Breast Milk in China. Molecules 2022, 27, 6324. https://doi.org/10.3390/molecules27196324
Zhang Y, Zhang H, Karrar E, Wei W, Zeng W, Jin Q, Wang X. Associations of Region and Lactation Stage with Odd-Chain Fatty Acid Profile in Triglycerides of Breast Milk in China. Molecules. 2022; 27(19):6324. https://doi.org/10.3390/molecules27196324
Chicago/Turabian StyleZhang, Yiren, Hui Zhang, Emad Karrar, Wei Wei, Wei Zeng, Qingzhe Jin, and Xingguo Wang. 2022. "Associations of Region and Lactation Stage with Odd-Chain Fatty Acid Profile in Triglycerides of Breast Milk in China" Molecules 27, no. 19: 6324. https://doi.org/10.3390/molecules27196324