
Ethanolic Extract from Pteris wallichiana Alleviates DSS-Induced Intestinal Inflammation and Intestinal Barrier Dysfunction by Inhibiting the TLR4/NF-κB Pathway and Regulating Tight Junction Proteins

 and
and
Abstract
:1. Introduction
2. Results
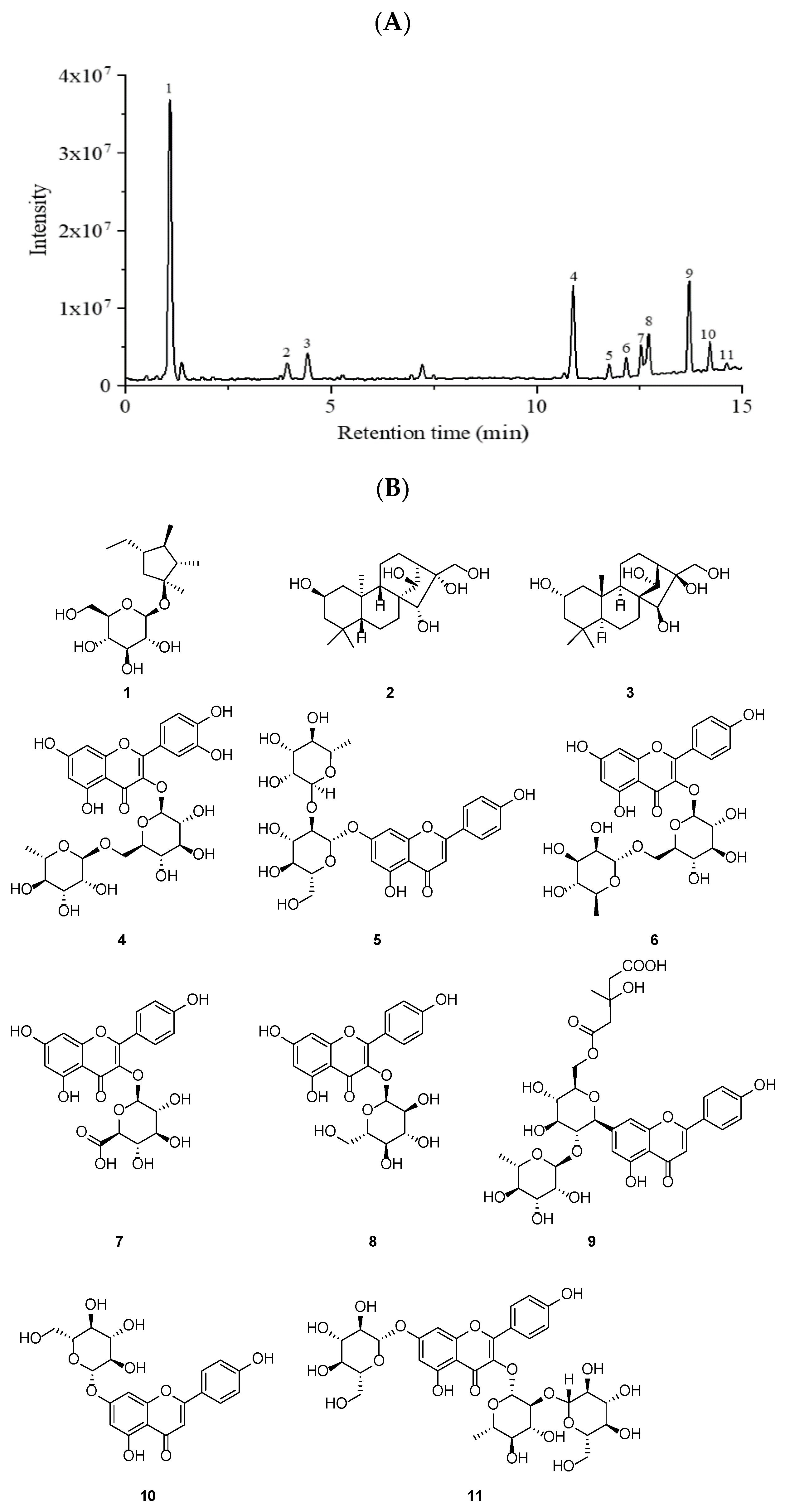
2.1. Chemical Composition of PWE
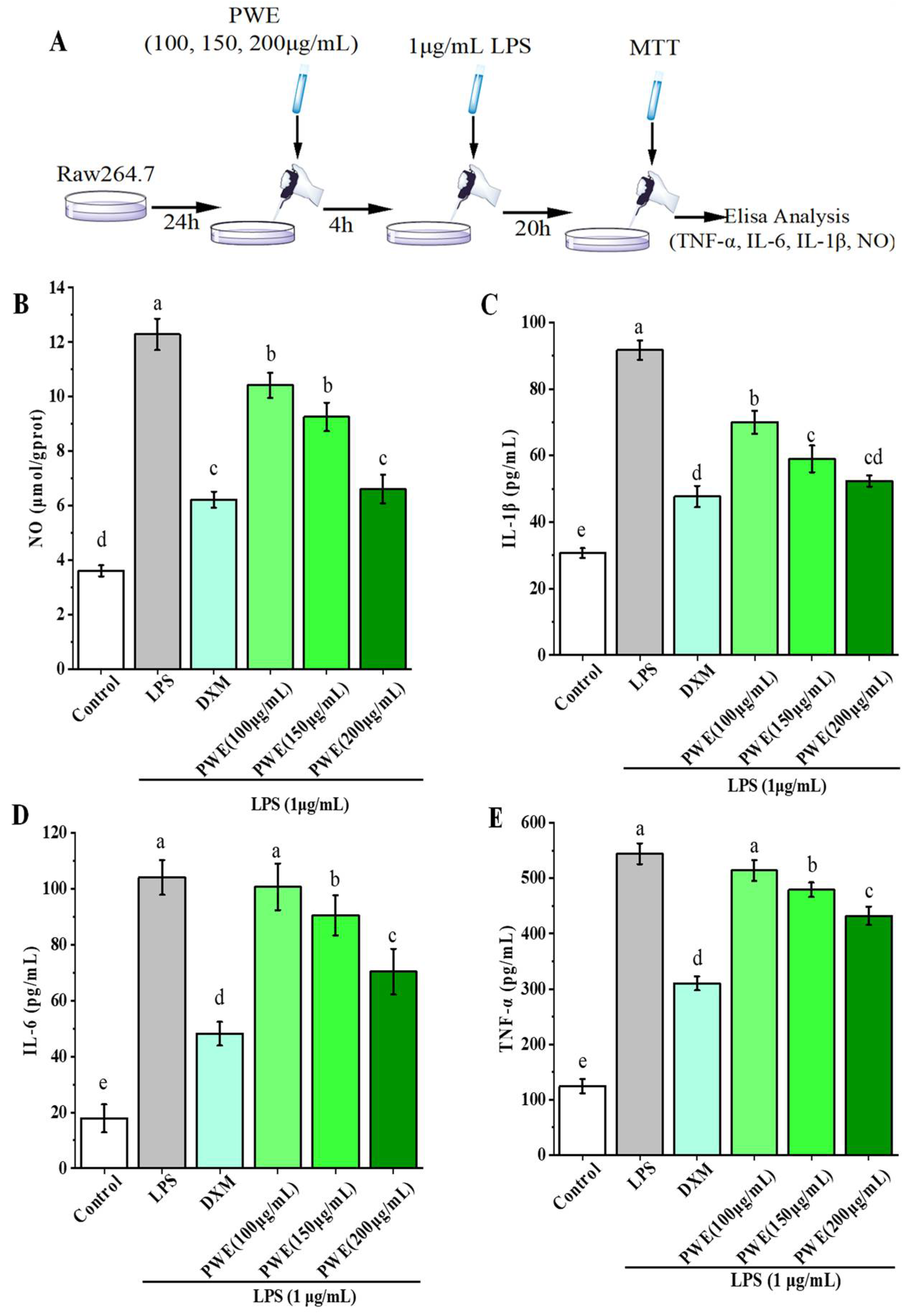
2.2. PWE Suppressed Inflammation Response in LPS-Induced RAW264.7 Cells
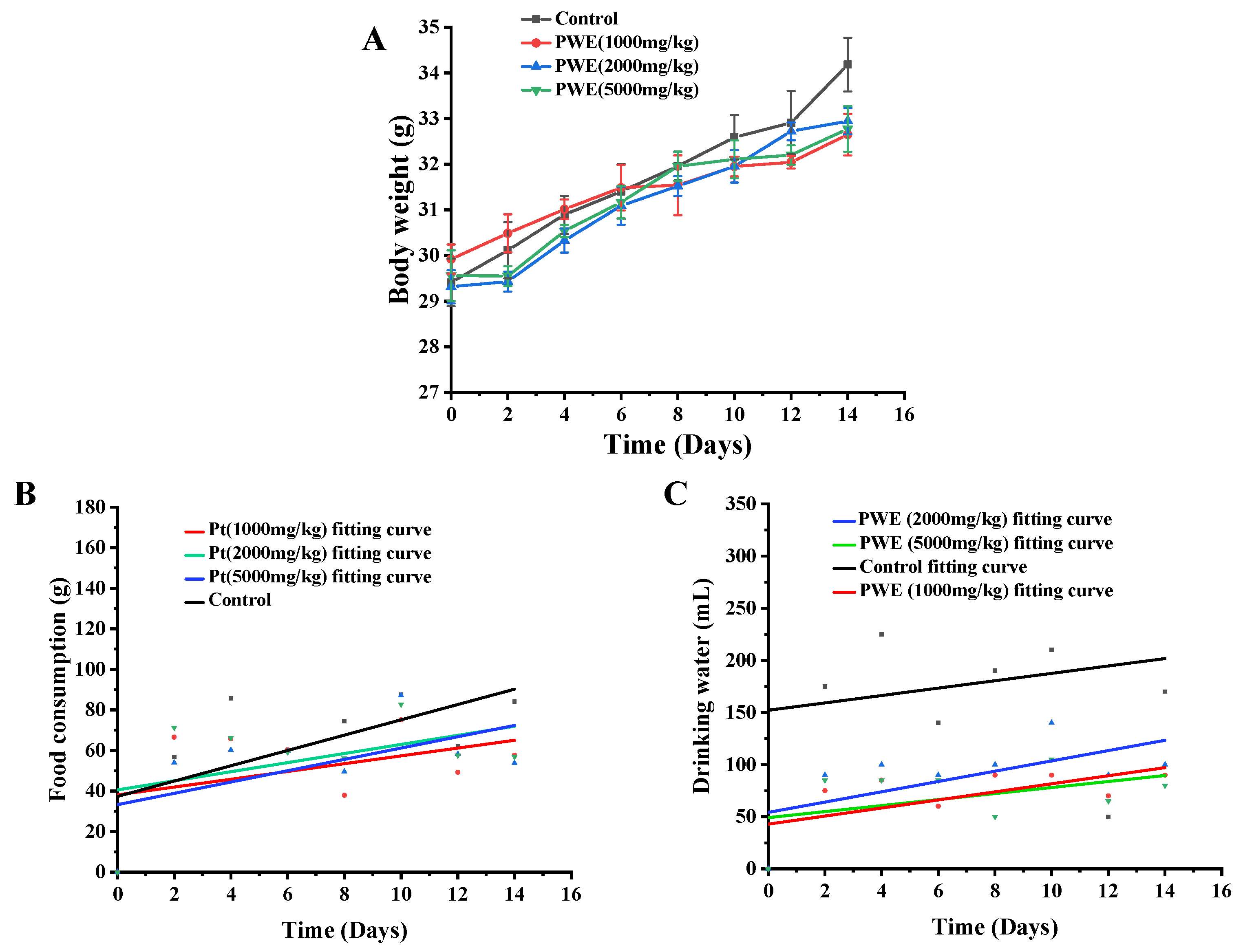
2.3. Acute Toxicity Evaluation of PWE
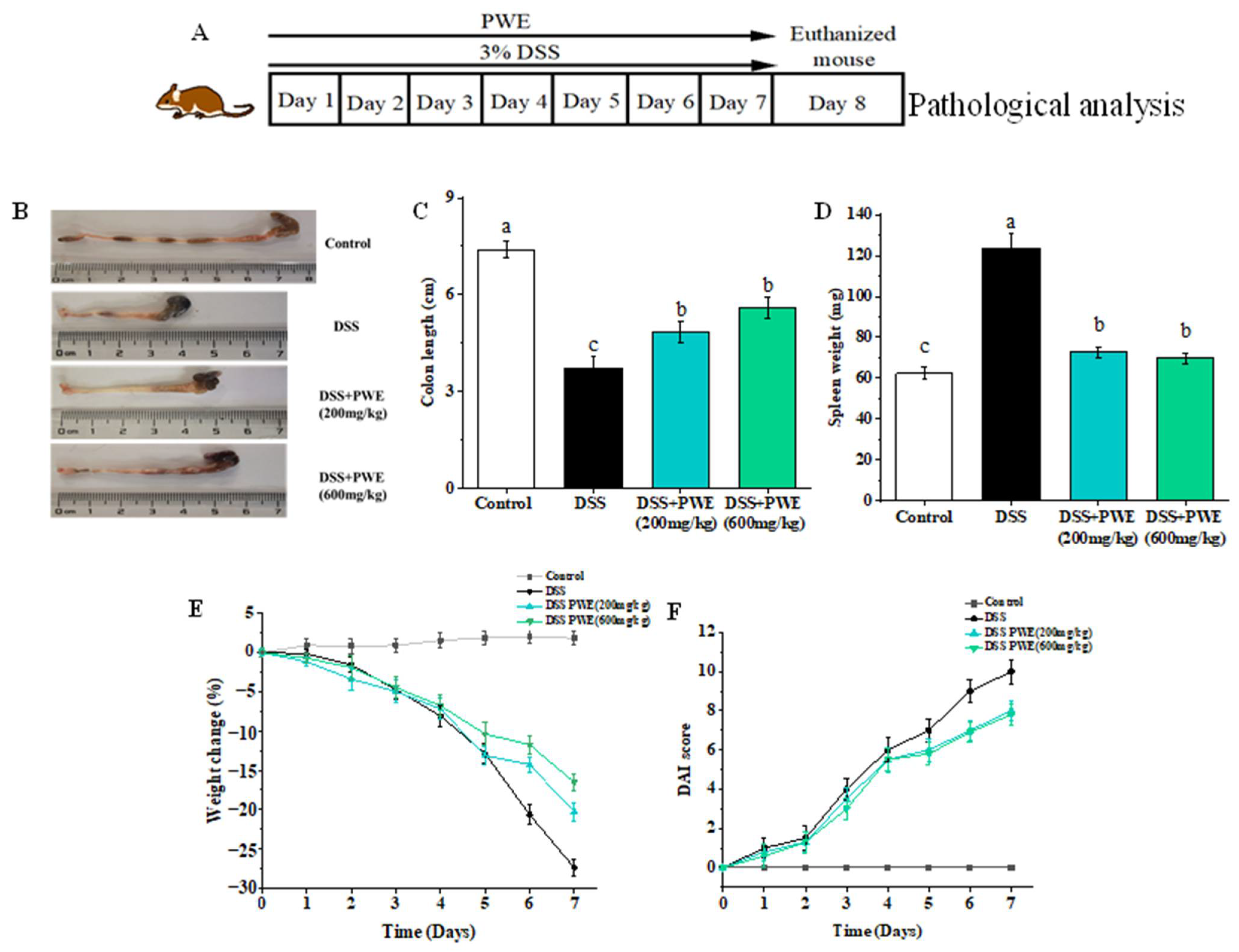
2.4. PWE Administration Attenuated DSS-Induced Ulcerative Colitis in Mice
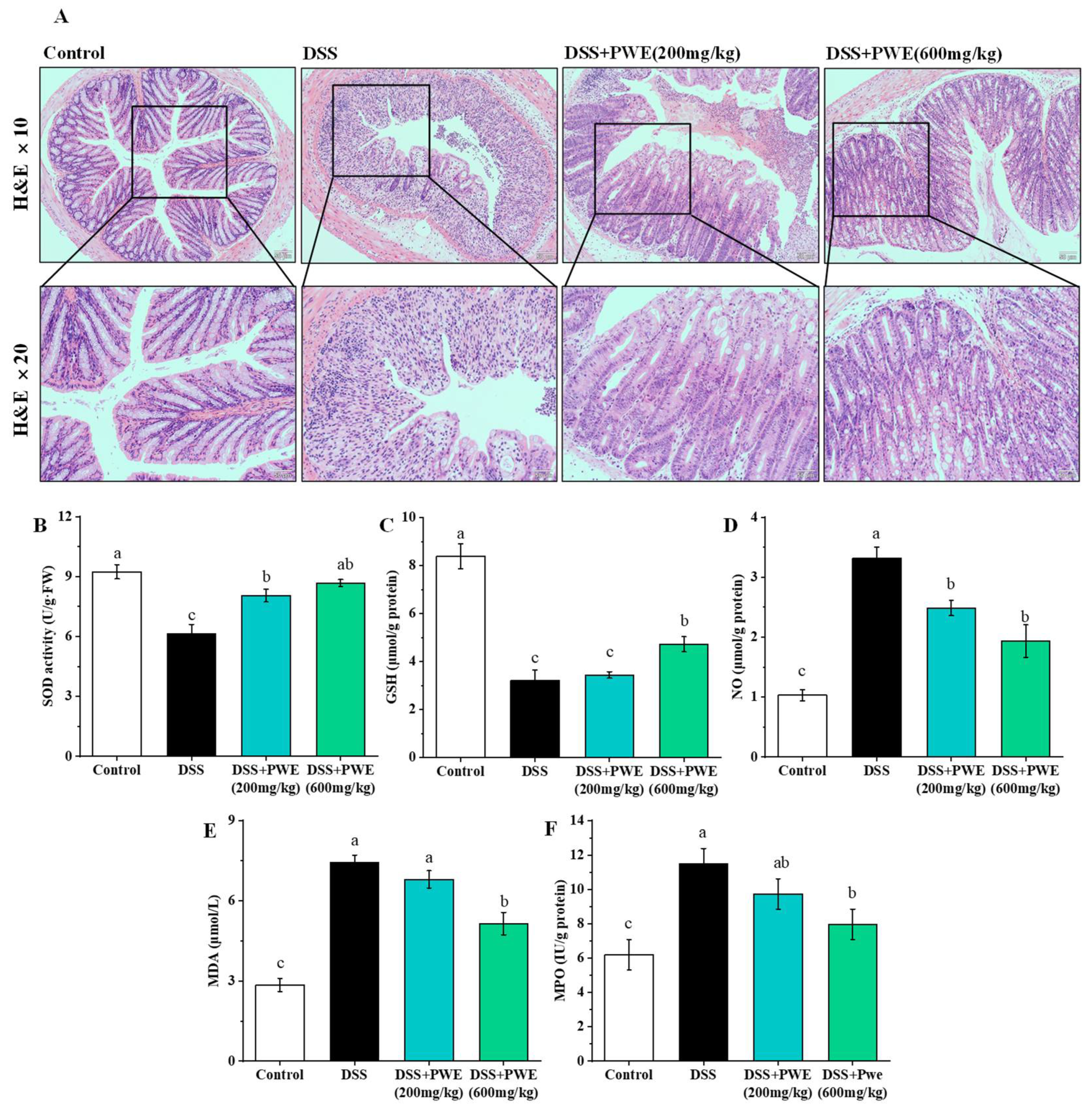
2.5. PWE Administration Attenuated Histopathological Changes in Colonic Tissues
2.6. PWE Ameliorated Oxidative Stress in the Colonic Tissues
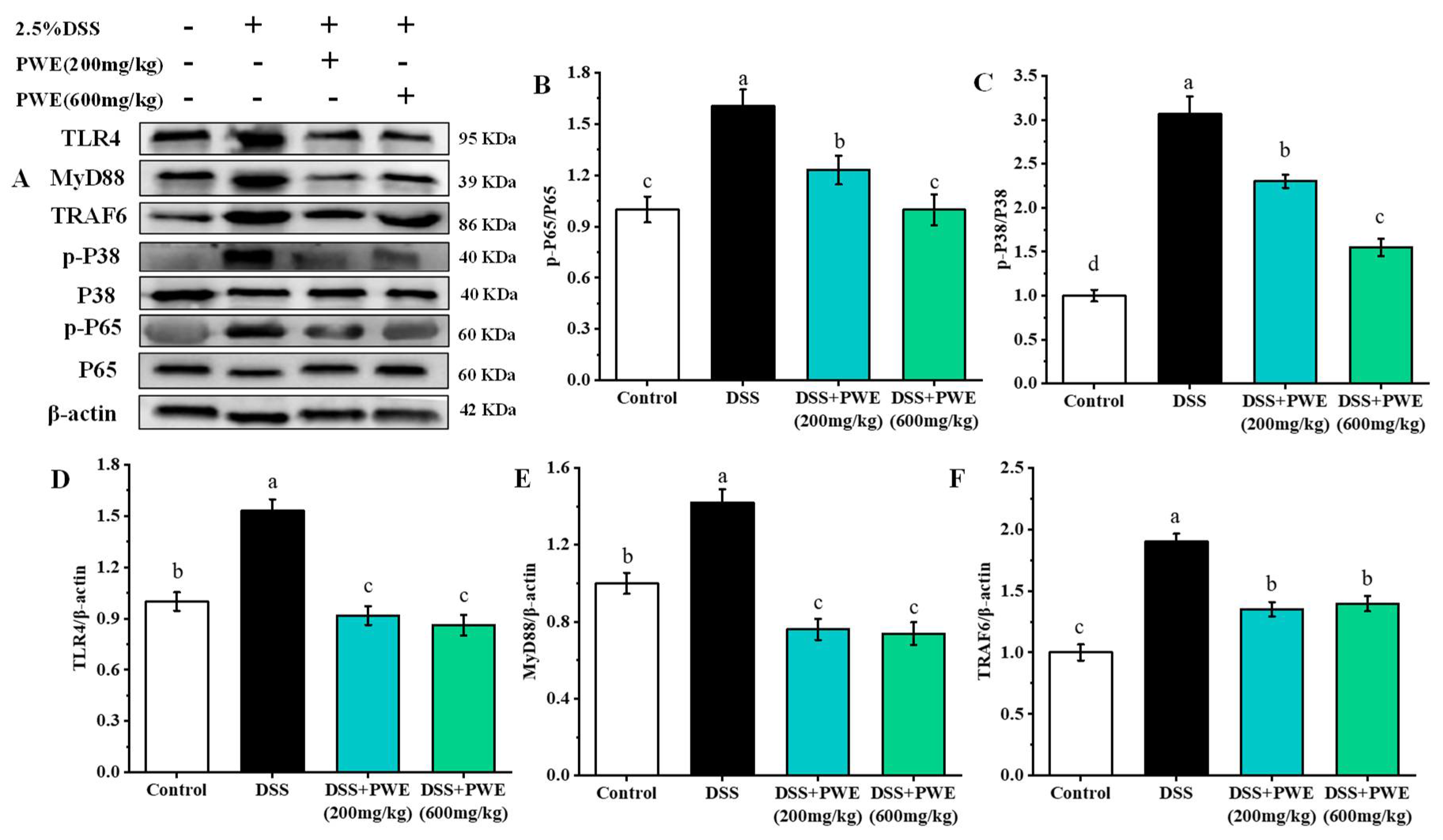
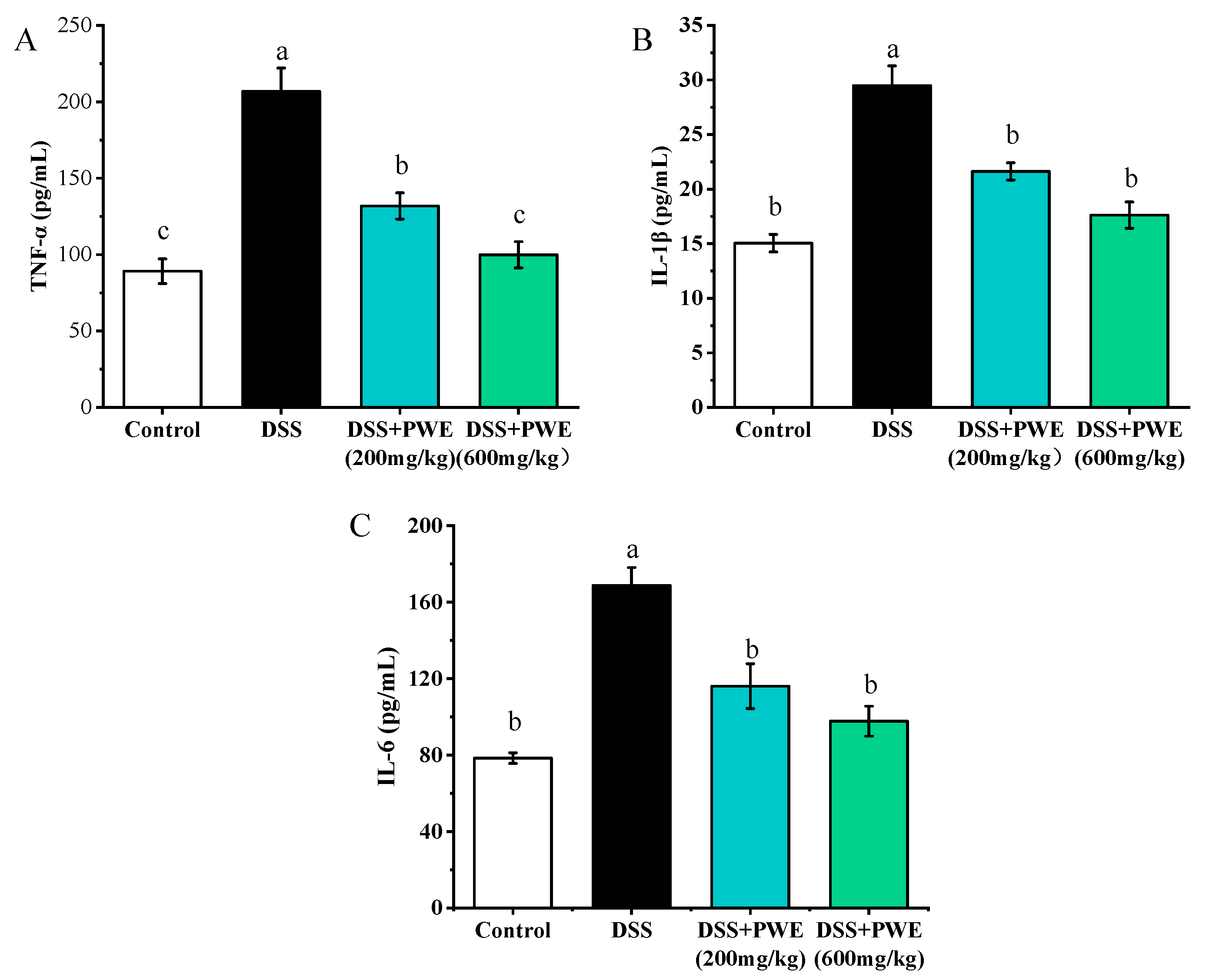
2.7. PWE Administration Inhibited the Inflammatory Response by TLR-4 Signaling Pathway
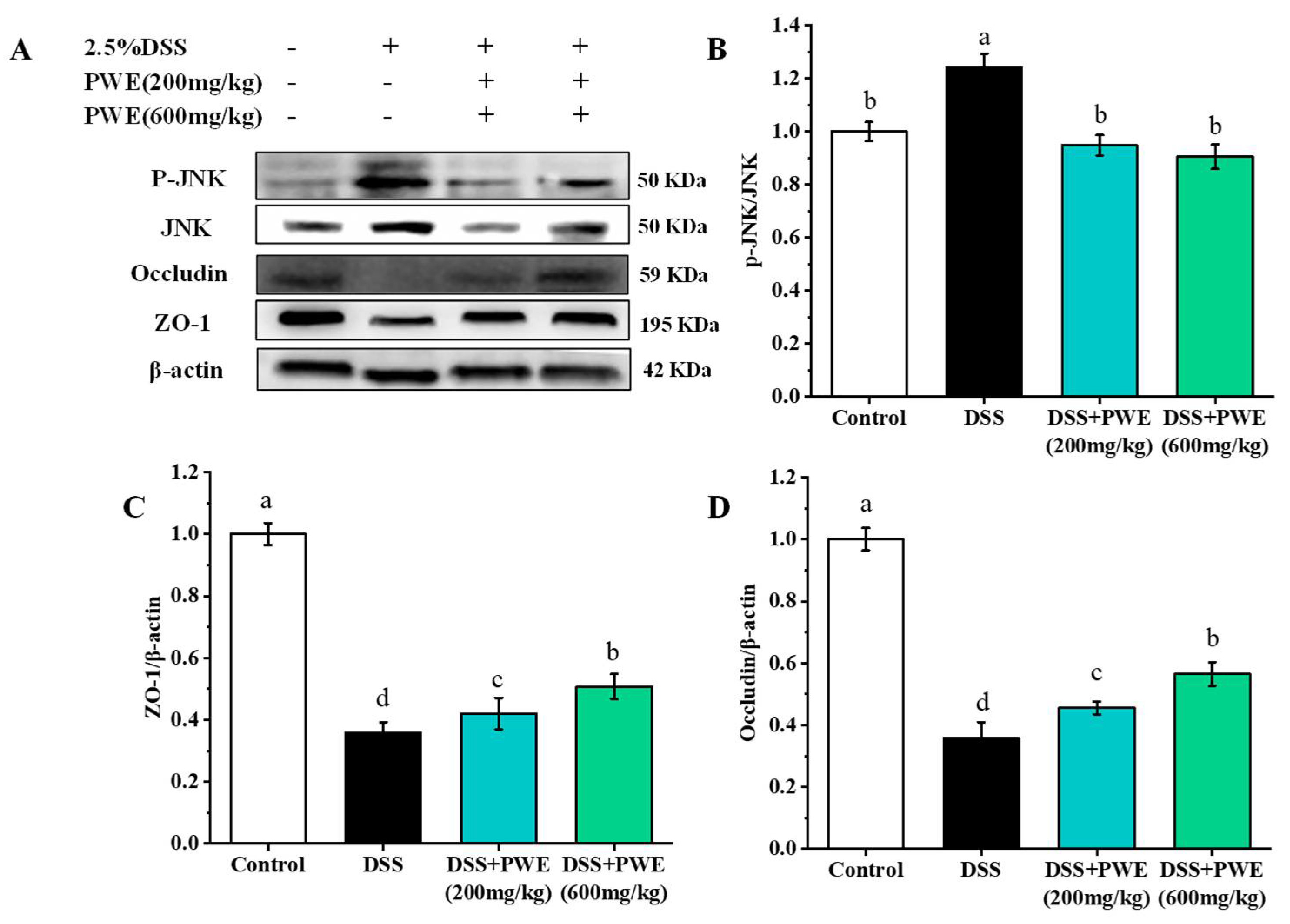
2.8. PWE Maintained Intestinal Barrier Function
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Extraction
4.2. Determination of the Contents of Total Phenolics, Flavonoids, and Terpenoids
4.3. Identification of Chemical Constituents in PWE by UHPLC-ESI-HRMS/MS
4.4. Cell Culture and Viability Assay
4.5. Cytokine Assays
4.6. Animals
4.7. Acute Toxicity Assays
4.8. Animal Experiment Design of DSS-Induced Ulcerative Colitis
4.9. Hematoxylin and Eosin (H&E) Staining
4.10. Determination of GSH, SOD, MPO, MDA, and NO Content in Colon Tissues
4.11. Western Blotting Analysis
4.12. Statistics Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ganesan, K.; Quiles, J.; Daglia, M.; Xiao, J.; Xu, B. Dietary phytochemicals modulate intestinal epithelial barrier dysfunction and autoimmune diseases. Food Front. 2021, 2, 357–382. [Google Scholar] [CrossRef]
- Yu, W.; Li, Z.; Long, F.; Chen, W.; Geng, Y.; Xie, Z.; Yao, M.; Han, B.; Liu, T. A systems pharmacology approach to determine active compounds and action mechanisms of Xipayi KuiJie’an enema for treatment of ulcerative colitis. Sci. Rep. 2017, 7, 4255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roncoroni, L.; Gori, R.; Elli, L.; Tontini, G.E.; Doneda, L.; Norsa, L.; Cuomo, M.; Lombardo, V.; Scricciolo, A.; Caprioli, F.; et al. Nutrition in patients with inflammatory bowel diseases: A narrative review. Nutrients 2022, 14, 751. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Yang, L.; Zhang, X.; Ji, P.; Hua, Y.; Wei, Y. Huang-lian-jie-du decoction ameliorates acute ulcerative colitis in mice via regulating NF-κB and Nrf2 signaling pathways and enhancing intestinal barrier function. Front. Pharmacol. 2019, 10, 1354. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.; Auyeung, K. Inflammatory bowel disease: Etiology, pathogenesis and current therapy. Curr. Pharm. Des. 2013, 20, 263–274. [Google Scholar] [CrossRef]
- Baumgart, D.C.; Sandborn, W.J. Inflammatory bowel disease: Clinical aspects and established and evolving therapies. Lancet 2007, 396, 1641–1657. [Google Scholar] [CrossRef]
- Zeng, X.; Qiu, H. New Information of Fern in South China. J. Fujian For. Sci. Tech. 2015, 42, 166–168. [Google Scholar] [CrossRef]
- Pei, J. Chinese Medicinal Plants; Science Press: Beijing, China, 1955; Volume 1, pp. 547–548. [Google Scholar]
- Hou, M.; Hu, W.; Hao, K.; Xiu, Z.; Cao, S. Chemical constituents from Pteris ensiformis Burm. (Pteridaceae). Biochem. Syst. Ecol. 2020, 92, 104107. [Google Scholar] [CrossRef]
- Wang, R.; Wan, Z.; Gao, F.; Khan, A.; Fan, J.; Yang, X.; Cao, J.; Cheng, G.; Zhao, T.R. Chemical constituents of Pteris wallichiana J. Agardh (Pteridaceae). Biochem. Syst. Ecol. 2017, 71, 225–229. [Google Scholar] [CrossRef]
- González, R.; Ballester, I.; López-Posadas, R.; Suárez, M.; Medina, F. Effects of flavonoids and other polyphenols on inflammation. Crit. Rev. Food Sci. 2011, 51, 331–362. [Google Scholar] [CrossRef]
- Joseph, N.; Anjum, N.; Tripathi, Y. Phytochemical screening and evaluation of polyphenols, flavonoids and antioxidant activity of prunus cerasoides D. don leaves. J. Res. Pharm. 2016, 10, 502–508. [Google Scholar]
- Li, F.; Wang, Y.; He, S.; Khan, A.; Xue, Q.; Cui, Q.; Liu, L.; Cheng, G. Targeted isolation of terpenoid indole alkaloids from Melodinus cochinchinensis (Lour.) Merr. using molecular networking and their biological activities. Ind. Crops Prod. 2020, 157, 112922. [Google Scholar] [CrossRef]
- Wang, Y.; Tian, L.; Wang, Y.; Zhao, T.; Khan, A.; Wang, Y.; Cao, J.; Cheng, G. Protective effect of Que Zui tea hot-water and aqueous ethanol extract against acetaminophen-induced liver injury in mice via inhibition of oxidative stress, inflammation, and apoptosis. Food Funct. 2021, 12, 2468–2480. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Ni, Z.; Thakur, K.; Zhang, F.; Wei, Z.J. Natural compounds play therapeutic roles in various human pathologies via regulating endoplasmic reticulum pathway. Med. Drug Discov. 2020, 8, 100065. [Google Scholar] [CrossRef]
- Ge, X.; Ye, G.; Li, P.; Wang, W.; Gao, J.; Zhao, W. Cytotoxic Diterpenoids and Sesquiterpenoids from Pteris multifida. J. Nat. Prod. 2008, 71, 227–231. [Google Scholar] [CrossRef]
- Wei, Z.; Wang, Y.; Liu, H.; Luo, Y.; Su, W. Melitidin: A flavanone glycoside from Citrus grandis ‘Tomentosa’. Nat. Prod. Commun. 2013, 8, 457–458. [Google Scholar] [CrossRef] [Green Version]
- Pereira, O.; Silva, A.; Domingues, M.; Domingues, M.; Cardoso, S. Identification of phenolic constituents of Cytisus multiflorus. Food Chem. 2012, 131, 652–659. [Google Scholar] [CrossRef]
- Carmona, M.; Sánchez, A.M.; Ferreres, F.; Zalacain, A.; Tomás-Barberán, F.; Alonso, G.L. Identification of the flavonoid fraction in saffron spice by LC/DAD/MS/MS: Comparative study of samples from different geographical origins. Food Chem. 2007, 100, 445–450. [Google Scholar] [CrossRef]
- Hu, W.; Hou, M.; Hao, K.; Zhang, X.; Chen, Y. Phytochemical and chemotaxonomic studies on Pteris wallichiana. J. Agardh. Biochem. Syst. Ecol. 2021, 95, 104242. [Google Scholar] [CrossRef]
- Duan, L.; Cheng, S.; Li, L.; Liu, Y.; Liu, G. Natural anti-Inflammatory compounds as drug candidates for inflammatory bowel disease. Front. Pharmacol. 2021, 12, 684486. [Google Scholar] [CrossRef]
- Gizem, C.; Kone, V.; Luigi, L.; Gabriele, R.; Dominique, D.; Xiao, J.; Esre, C. Interaction of dietary polyphenols and gut microbiota: Microbial metabolism of polyphenols, influence on the gut microbiota, and implications on host health. Food Front. 2020, 1, 109–133. [Google Scholar] [CrossRef]
- Mascaraque, C.; Aranda, C.; Ocón, B.; Monte, M.J.; Suárez, M.D.; Zarzuelo, A.; Marín, J.J.G.; Martínez-Augustin, O.; de Medina, F.S. Rutin has intestinal antiinflammatory effects in the CD4+ CD62L+ T cell transfer model of colitis. Pharmacol. Res. 2014, 90, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.; Shin, H.; Cho, J.; Kang, S.; Moon, J. Isolation and structural determination of free radical scavenging compounds from korean fermented red Pepper Paste (Kochujang). Food Sci. Biotechnol. 2009, 18, 463–470. [Google Scholar] [CrossRef]
- Fang, J.; Cao, Z.; Song, X.; Zhang, X.; Xiao, F. Rhoifolin alleviates inflammation of acute inflammation animal models and LPS-Induced RAW264.7 cells via IKKβ/NF-κB signaling pathway. Inflammation 2020, 43, 2191–2201. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Ji, G.; Sung, M. Dietary kaempferol suppresses inflammation of dextran sulfate sodium-induced colitis in mice. Digest. Dis. Sci. 2012, 57, 355–363. [Google Scholar] [CrossRef]
- Zou, H.; Ye, H.; Kamaraj, R.; Zhang, T.; Zhang, J.; Pavek, P. A review on pharmacological activities and synergistic effect of quercetin with small molecule agents. Phytomedicine 2021, 92, 153736. [Google Scholar] [CrossRef]
- Wan, F.; Han, H.; Zhong, R.; Wang, M.; Tang, S.; Zhang, S.; Hou, F.; Yi, B.; Zhang, H. Dihydroquercetin supplement alleviates colonic inflammation potentially through improved gut microbiota community in mice. Food Funct. 2021, 12, 11420–11434. [Google Scholar] [CrossRef]
- Annaházi, A.; Molnár, T.; Rosztóczy, A.; Gecse, K.; Inczefi, O.; Nagy, F.; Földesi, I.; Szűcs, M.; Dabek, M.; Ferrier, L. Fecal MMP-9: A new noninvasive differential diagnostic and activity marker in ulcerative colitis. J. Crohn’s Colitis 2013, 19, 316–320. [Google Scholar] [CrossRef]
- Kawanishi, S.; Hiraku, Y.; Pinlaor, S.; Ma, N. Oxidative and nitrative DNA damage in animals and patients with inflammatory diseases in relation to inflammation-related carcinogenesis. Biol. Chem. 2006, 98, 117–372. [Google Scholar] [CrossRef]
- Thakur, K.; Wei, Z.; Tomar, S. Chapter 5—Riboflavin-enriched fermented soy milk for redox-mediated gut modulation: In the search of novel prebiotics. In Molecular Nutrition; Patel, V.B., Ed.; Academic Press: Amsterdam, The Netherlands, 2020; pp. 91–103. [Google Scholar]
- Rana, S.; Sharma, S.; Prasad, K.; Sinha, S.; Singh, K. Role of oxidative stress & antioxidant defence in ulcerative colitis patients from north India. Indian J. Med. Res. 2014, 139, 568–571. [Google Scholar]
- Rudolph, V.; Andrié, R.; Rudolph, T.; Friedrichs, K.; Klinke, A.; Hirsch-Hoffmann, B.; Schwoerer, A.; Lau, D.; Fu, X.; Klingel, K. Myeloperoxidase acts as a profibrotic mediator of atrial fibrillation. Nat. Med. 2010, 16, 470–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Lu, C.; Fan, Z.; Zhao, T.; Cheng, G.; Wang, Y.; Cao, J.; Liu, Y. Hypoglycemic and hypolipidemic effects of Epigynum auritum in high fat diet and streptozotocin-induced diabetic rats. J. Ethnopharmacol. 2022, 288, 114986. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wang, Y.; Patel, G.; Xue, Q.; Njateng, G.S.S.; Cai, S.; Cheng, G.; Kai, G. In vitro and in vivo anti-inflammatory effects of different extracts from Epigynum auritum through down-regulation of NF-κB and MAPK signaling pathways. J. Ethnopharmacol. 2020, 261, 113105. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wang, Y.; Fan, Z.; Xue, Q.; Njateng, G.; Liu, Y.; Cao, J.; Khan, A.; Cheng, G. Chemical constituents and anti-inflammatory activity of the total alkaloid extract from Melodinus cochinchinensis (Lour.) Merr. and its inhibition of the NF-κB and MAPK signaling pathways. Phytomedicine 2021, 91, 153684. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Wang, Y.; Tian, L.; Yang, M.; He, S.; Liu, Y.; Khan, A.; Li, Y.; Cao, J.; Cheng, G. Anneslea fragrans Wall. ameliorates ulcerative colitis via inhibiting NF-κB and MAPK activation and mediating intestinal barrier integrity. J. Ethnopharmacol. 2021, 278, 114304. [Google Scholar] [CrossRef]
- Vallabhapurapu, S.; Karin, M. Regulation and function of NF-κB transcription factors in the immune system. Annu. Rev. Immunol. 2009, 27, 693–733. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, Y.; Gong, X.; Cheng, G.; Pu, S.; Cai, S. The preventive effect of phenolic-rich extracts from Chinese sumac fruits against nonalcoholic fatty liver disease in rats induced by a high-fat diet. Food Funct. 2020, 11, 799–812. [Google Scholar] [CrossRef]
- Huang, S.; Fu, Y.; Xu, B.; Liu, C.; Wang, Q.; Luo, S.; Nong, F.; Wang, X.; Huang, S.; Chen, J. Wogonoside alleviates colitis by improving intestinal epithelial barrier function via the MLCK/pMLC2 pathway—ScienceDirect. Phytomedicine 2020, 68, 153179. [Google Scholar] [CrossRef]
- Sánchez-Fidalgo, S.; Villegas, I.; Rosillo, M.; Aparicio-Soto, M.; Alarcón, D. Dietary squalene supplementation improves DSS-induced acute colitis by downregulating p38 MAPK and NFkB signaling pathways. Mol. Nutr. Food Res. 2015, 59, 284–292. [Google Scholar] [CrossRef]
- Poritz, L.; Garver, K.; Tilberg, A.; Koltun, W. Tumor necrosis factor alpha disrupts tight junction assembly. J. Surg. Res. 2004, 116, 14–18. [Google Scholar] [CrossRef]
- Amasheh, S. Claudin-2 expression induces cation-selective channels in tight junctions of epithelial cells. J. Cell Sci. 2002, 115, 4969–4976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummins, P.M. Occludin: One protein, many forms. Cell Mol. Biol. 2011, 32, 242–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinsaku, T.; Tomohito, H.; Mikio, F.; Zegers, M. ZO-1 Knockout by TALEN-Mediated Gene Targeting in MDCK Cells: Involvement of ZO-1 in the Regulation of Cytoskeleton and Cell Shape. PLoS ONE 2014, 9, e104994. [Google Scholar] [CrossRef] [Green Version]
- Kojima, T.; Fuchimoto, J.; Yamaguchi, H.; Ito, T. c-Jun N-terminal kinase is largely involved in the regulation of tricellular tight junctions via tricellulin in human pancreatic duct epithelial cells. J. Cell. Physiol. 2010, 225, 720–733. [Google Scholar] [CrossRef]
- Cui, Q.; Wang, Y.; Zhou, W.; He, S.; Yang, M.; Cao, J.; Khan, A.; Cheng, G. Phenolic composition, antioxidant and cytoprotective effects of aqueous-methanol extract from Anneslea fragrans leaves as affected by drying methods. Int. J. Food Sci. Tech. 2021, 56, 4807–4819. [Google Scholar] [CrossRef]
- Fan, Z.; Wang, Y.; Yang, M.; Cao, J.; Cheng, G. UHPLC-ESI-HRMS/MS analysis on phenolic compositions of different E Se tea extracts and their antioxidant and cytoprotective activities. Food Chem. 2020, 318, 126512. [Google Scholar] [CrossRef]
- Yang, M.; Ma, Y.; Wang, Z.; Khan, A.; Zhou, W.; Zhao, T.; Cao, J.; Cheng, G.; Cai, S. Phenolic constituents, antioxidant and cytoprotective activities of crude extract and fractions from cultivated artichoke inflorescence. Ind. Crops Prod. 2020, 143, 11433. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Y.-D.; Xue, Q.-W.; Zhao, T.-R.; Khan, A.; Wang, Y.-F.; Liu, Y.-P.; Cao, J.-X.; Cheng, G.-G. The effect of ultra-high pretreatment on free, esterified and insoluble-bound phenolics from mango leaves and their antioxidant and cytoprotective activities. Food Chem. 2021, 368, 130864. [Google Scholar] [CrossRef]
- Gao, S.; Zhao, T.; Liu, Y.; Wang, Y.; Cheng, G.; Cao, J. Phenolic constituents, antioxidant activity and neuroprotective effects of ethanol extracts of fruits, leaves and flower buds from Vaccinium dunalianum Wight. Food Chem. 2022, 374, 131752. [Google Scholar] [CrossRef]
- Bor, J.; Chen, H.; Yen, G. Evaluation of antioxidant activity and inhibitory effect on nitric oxide production of some common vegetables. J. Agric. Food Chem. 2006, 54, 1680–1686. [Google Scholar] [CrossRef]
- Yu, G.; Wang, X.; Jing-Hong, H. Optimization of total terpenoids determination in volatile oil by response surface methodology. Mod. Chin. Med. 2018, 20, 1262–1269. [Google Scholar] [CrossRef]
- Yang, M.; Sun, P.; Fan, Z.; Khan, A.; Xue, Q.; Wang, Y.; Cao, J.; Cheng, G. Safety evaluation and hypolipidemic effect of aqueous-ethanol and hot-water extracts from E Se tea in rats. Food Chem. Toxicol. 2021, 156, 112506. [Google Scholar] [CrossRef] [PubMed]
- Zolotarev, K.; Belyaeva, N.; Mikhailov, A.; Mikhailova, M. Dependence between LD50 for Rodents and LC50 for Adult Fish and Fish Embryos. Bull. Exp. Biol. Med. 2017, 162, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Jin, X.; Xiao, X.; Qiu, B. PI3K/Akt signaling pathway is involved in the pathogenesis of ulcerative colitis. Inflamm. Res. 2011, 60, 727–734. [Google Scholar] [CrossRef]
- Peng, Y.; Yan, Y.; Wan, P.; Chen, D.; Ding, Y.; Ran, L.; Mi, J.; Lu, L.; Zhang, Z.; Li, X. Gut microbiota modulation and anti-inflammatory properties of anthocyanins from the fruits of Lycium ruthenicum Murray in dextran sodium sulfate-induced colitis in mice. Free Radic. Biol. Med. 2019, 136, 96–108. [Google Scholar] [CrossRef]
- Zhai, K.; Duan, H.; Fang, X. Protective effects of paeonol on inflammatory response in IL-1β-induced human fibroblast-like synoviocytes and rheumatoid arthritis progression via modulating NF-κB pathway. Inflammopharmacology 2017, 25, 523–532. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Rt # (Min) | Compounds | Molecular Formula | [M-H]- m/z | MS/MS Fragment Ions | Ref. |
|---|---|---|---|---|---|---|
| 1 | 1.08 | sucrose | C12H22O11 | 341.1096 | 59.0126, 71.0126, 101.0232, 323.4808 | HMDB * |
| 2 | 3.94 | 2β,14β,15α,16α,17-pentahydroxy-ent-kaurane | C20H34O5 | 353.0886 | 85.0283, 135.0443, 173.0449, 179.0345 | [16] |
| 3 | 4.43 | 2α,14α,15β,16β,17-pentahydroxykaurane | C20H34O5 | 353.0887 | 93.0334, 135.0443, 173.0449, 191.0557 | [16] |
| 4 | 10.89 | Rutin | C27H30O16 | 609.1476 | 151.0026, 301.0353, 343.0476, 609.1475 | standard |
| 5 | 11.75 | Rhoifolin | C27H30O14 | 577.1578 | 102.9558, 285.0410, 431.0981,577.1572 | HMDB * |
| 6 | 12.17 | Kaempferol 3-rutinoside | C27H30O15 | 593.1527 | 151.0027, 285.0408, 327.0510, 593.1530 | standard |
| 7 | 12.52 | Kaempferol 3-O-β-D-glucuronide | C21H18O12 | 461.0737 | 85.0283, 113.0234, 285.0410, 389.3503 | [17] |
| 8 | 12.71 | Kaempferol 3-O-β-D-glucoside | C21H20O11 | 447.0944 | 151.0028, 284.0329, 300.0280, 447.0925 | HMDB * |
| 9 | 13.71 | Melitidin | C33H40O17 | 7235045 | 73.7354, 347.8025, 451.3281, 677.4988 | [17] |
| 10 | 14.21 | Apigenin 7-O-β-D-glucopyranoside | C21H20O10 | 431.0992 | 227.0351, 255.0302, 284.0331, 431.0987 | [18] |
| 11 | 14.62 | Kaempferol-3-sophoroside-7-glucoside | C33H40O21 | 771.1798 | 151.0038, 301.0359, 609.1474, 771.1782 | [19] |
| Organ | Control (g/100 g) | PWE (1000 mg/kg) (g/100 g) | PWE (2000 mg/kg) (g/100 g) | PWE (5000 mg/kg) (g/100 g) |
|---|---|---|---|---|
| Heart | 0.54 ± 0.04 | 0.53 ± 0.03 | 0.54 ± 0.06 | 0.55 ± 0.02 |
| Liver | 4.48 ± 0.45 | 4.52 ± 0.56 | 4.47 ± 0.43 | 4.50 ± 0.38 |
| Spleen | 0.35 ± 0.02 | 0.36 ± 0.05 | 0.34 ± 0.03 | 0.36 ± 0.07 |
| Lung | 0.69 ± 0.06 | 0.70 ± 0.03 | 0.71 ± 0.06 | 0.68 ± 0.12 |
| Kidney | 1.08 ± 0.09 | 1.12 ± 0.02 | 1.10 ± 0.01 | 1.12 ± 0.04 |
| Thymus | 0.46 ± 0.04 | 0.48 ± 0.05 | 0.45 ± 0.05 | 0.47 ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, J.; Huang, Z.; Wang, Y.; Liu, Y.; Zhao, T.; Wang, Y.; Tian, L.; Cheng, G. Ethanolic Extract from Pteris wallichiana Alleviates DSS-Induced Intestinal Inflammation and Intestinal Barrier Dysfunction by Inhibiting the TLR4/NF-κB Pathway and Regulating Tight Junction Proteins. Molecules 2022, 27, 3093. https://doi.org/10.3390/molecules27103093
Tao J, Huang Z, Wang Y, Liu Y, Zhao T, Wang Y, Tian L, Cheng G. Ethanolic Extract from Pteris wallichiana Alleviates DSS-Induced Intestinal Inflammation and Intestinal Barrier Dysfunction by Inhibiting the TLR4/NF-κB Pathway and Regulating Tight Junction Proteins. Molecules. 2022; 27(10):3093. https://doi.org/10.3390/molecules27103093
Chicago/Turabian StyleTao, Junhong, Zhihua Huang, Yudan Wang, Yaping Liu, Tianrui Zhao, Yifen Wang, Lei Tian, and Guiguang Cheng. 2022. "Ethanolic Extract from Pteris wallichiana Alleviates DSS-Induced Intestinal Inflammation and Intestinal Barrier Dysfunction by Inhibiting the TLR4/NF-κB Pathway and Regulating Tight Junction Proteins" Molecules 27, no. 10: 3093. https://doi.org/10.3390/molecules27103093