
The Discovery of Actinospene, a New Polyene Macrolide with Broad Activity against Plant Fungal Pathogens and Pathogenic Yeasts

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
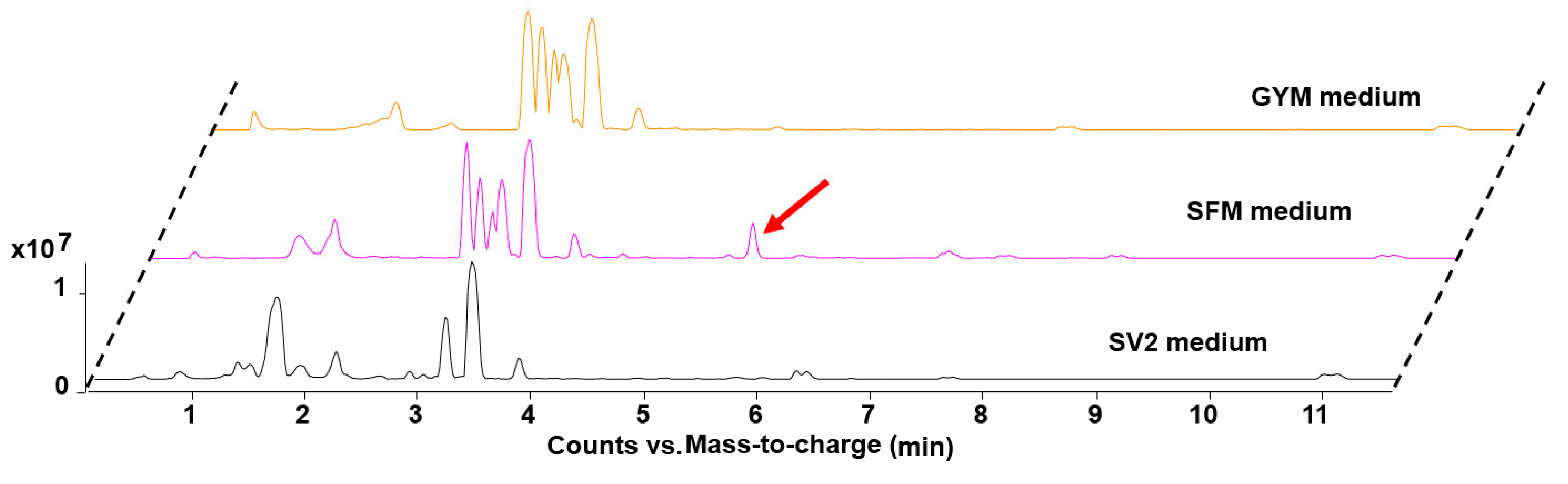
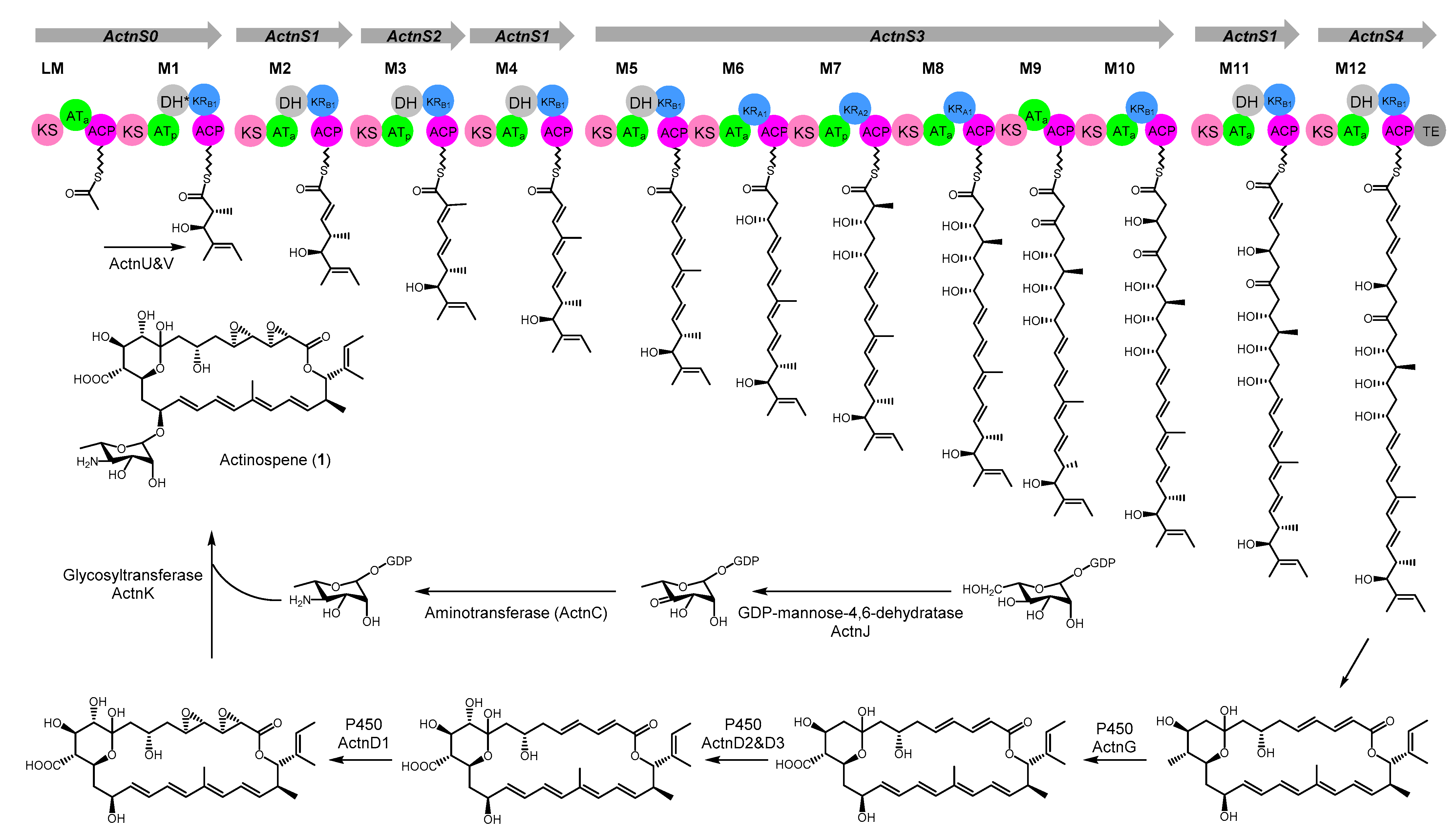
2.1. Genome Mining and Activation of the Actinospene Biosynthetic Gene Cluster
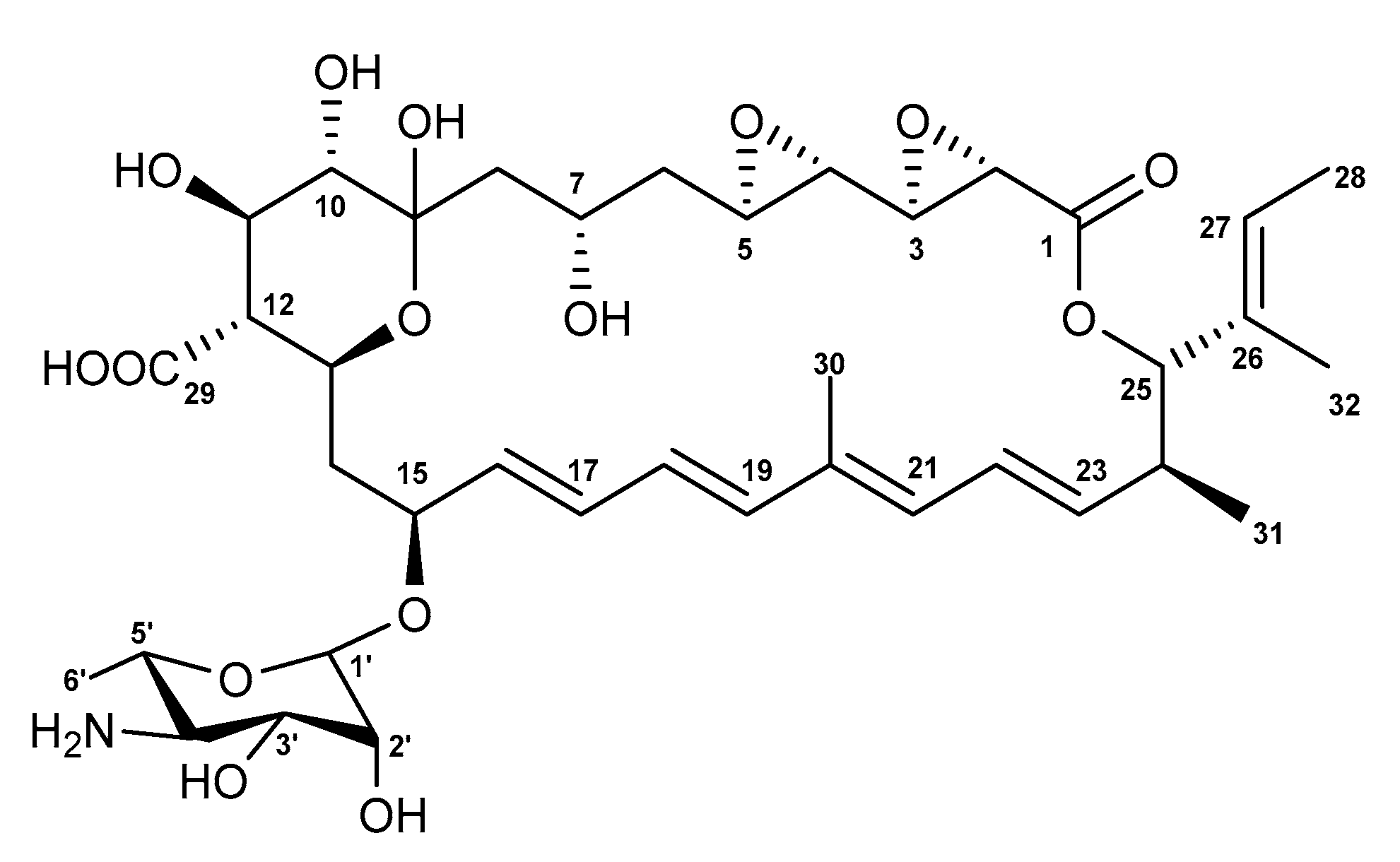
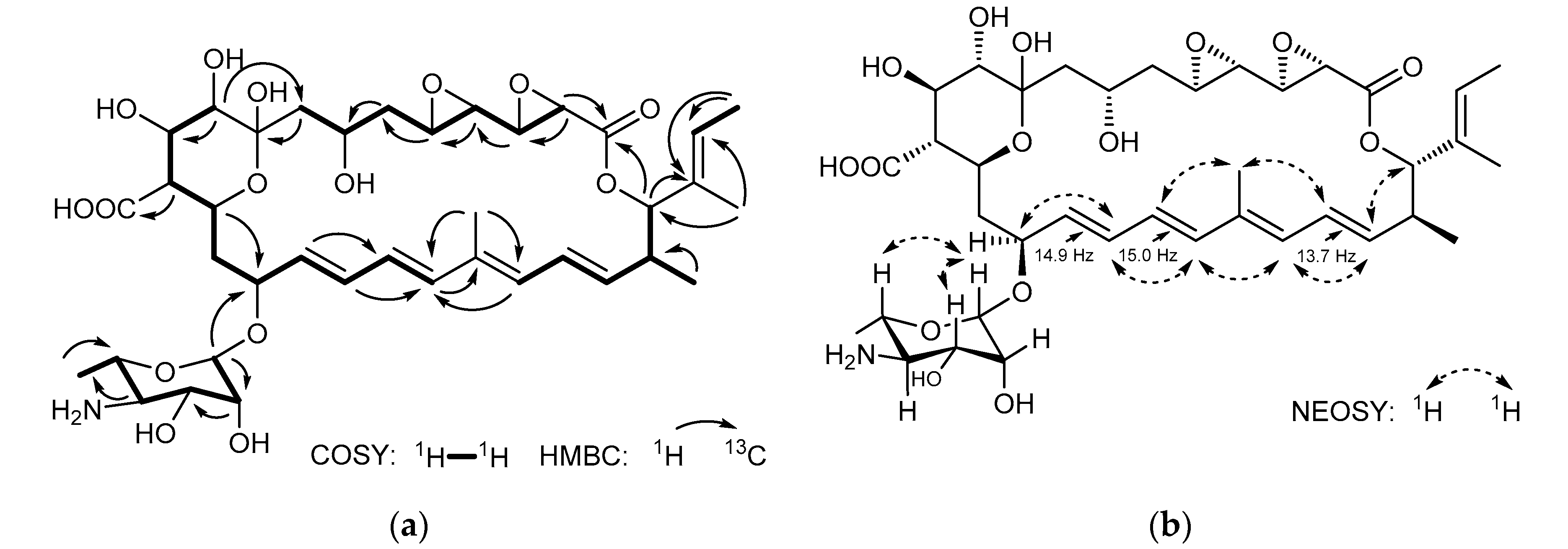
2.2. Isolation and Structure Elucidation
2.3. Proposed Model for Biosynthetic Pathway of Actinospene
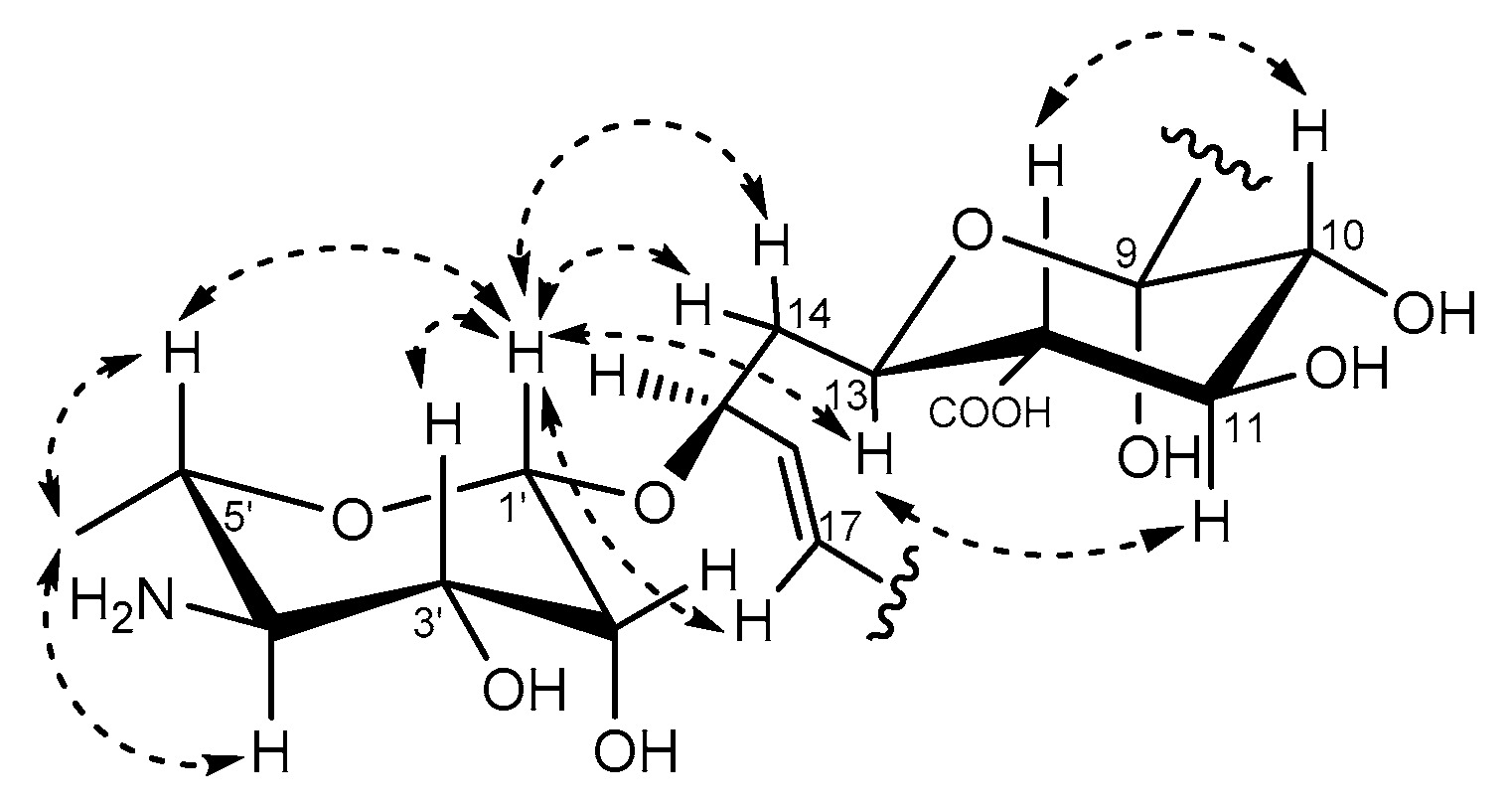
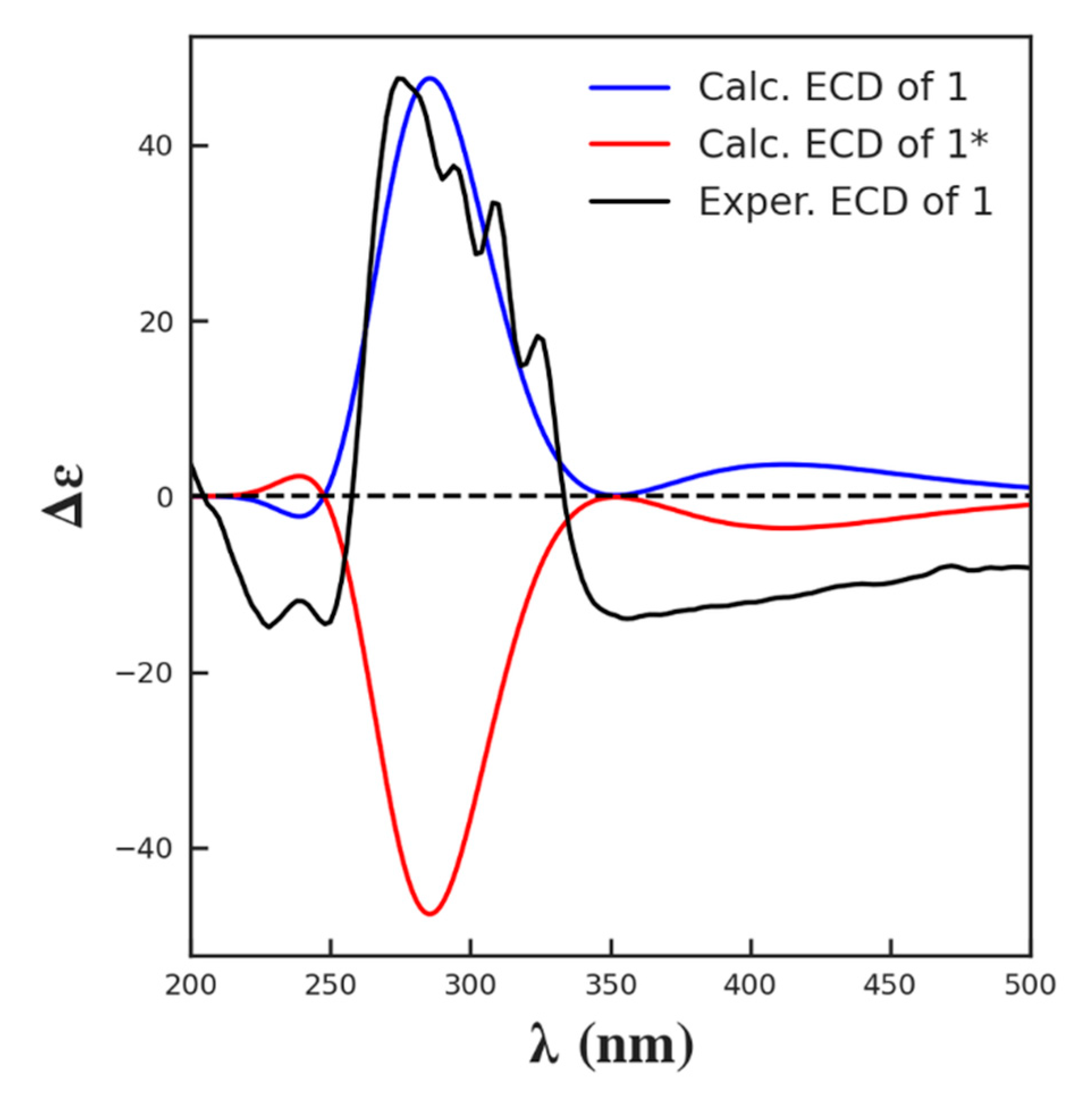
2.4. In Silico Prediction of Stereochemistry of Actinospene Core Structure
2.5. Bioactivity Analysis of the Actinospene
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Fermentation, Extraction, and Isolation
3.3. ECD Calculations
3.4. Antifungal Activity Assays
3.5. Microorganisms and Culture Conditions
3.6. Bioinformatics
4. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Almeida, F.; Rodrigues, M.L.; Coelho, C. The still underestimated problem of fungal diseases worldwide. Front. Microbiol. 2019, 10, 214. [Google Scholar] [CrossRef] [Green Version]
- Ons, L.; Bylemans, D.; Thevissen, K.; Cammue, B.P.A. Combining biocontrol agents with chemical fungicides for integrated plant fungal disease control. Microorganisms 2020, 8, 1930. [Google Scholar] [CrossRef]
- De Coninck, B.; Timmermans, P.; Vos, C.; Cammue, B.P.; Kazan, K. What lies beneath: Belowground defense strategies in plants. Trends Plant Sci. 2015, 20, 91–101. [Google Scholar] [CrossRef]
- Yao, X.; Zhang, Z.; Huang, J.; Wei, S.; Sun, X.; Chen, Y.; Liu, H.; Li, S. Candicidin isomer production is essential for biocontrol of cucumber rhizoctonia rot by Streptomyces albidoflavus W68. Appl. Environ. Microbiol. 2021, 87, e03078-20. [Google Scholar] [CrossRef]
- Han, X.; Wang, J.; Liu, L.; Shen, F.; Meng, Q.; Li, X.; Li, Y.; Liu, D. Identification and predictions regarding the biosynthesis pathway of polyene macrolides produced by Streptomyces roseoflavus Men-myco-93-63. Appl. Environ. Microbiol. 2021, 87, e03157-20. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; de Rond, T.; Moore, B.S. Mining genomes to illuminate the specialized chemistry of life. Nat. Rev. Genet. 2021, 22, 553–571. [Google Scholar] [CrossRef] [PubMed]
- Zotchev, S.B. Polyene macrolide antibiotics and their applications in human therapy. Curr. Med. Chem. 2003, 10, 211–223. [Google Scholar] [CrossRef]
- Hazen, E.L.; Brown, R. Two antifungal agents produced by a soil actinomycete. Science 1950, 112, 423. [Google Scholar] [PubMed]
- Stiller, E.T.; Vandeputte, J.; Wachtel, J.L. Amphotericins A and B, antifungal antibiotics produced by a streptomycete. II. The isolation and properties of the crystalline amphotericins. Antibiot. Annu. 1955, 3, 587–591. [Google Scholar]
- Caffrey, P.; De Poire, E.; Sheehan, J.; Sweeney, P. Polyene macrolide biosynthesis in streptomycetes and related bacteria: Recent advances from genome sequencing and experimental studies. Appl. Microbiol. Biotechnol. 2016, 100, 3893–3908. [Google Scholar] [CrossRef] [Green Version]
- Intra, B.; Greule, A.; Bechthold, A.; Euanorasetr, J.; Paululat, T.; Panbangred, W. Thailandins A and B, New polyene macrolactone compounds isolated from Actinokineospora bangkokensis Strain 44EHW(T), possessing antifungal activity against anthracnose fungi and pathogenic yeasts. J. Agric. Food Chem. 2016, 64, 5171–5179. [Google Scholar] [CrossRef]
- Aparicio, J.F.; Barreales, E.G.; Payero, T.D.; Vicente, C.M.; de Pedro, A.; Santos-Aberturas, J. Biotechnological production and application of the antibiotic pimaricin: Biosynthesis and its regulation. Appl. Microbiol. Biotechnol. 2016, 100, 61–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, D.; Yang, X.; Liu, J.; Caiyin, Q.; Zhao, G.; Li, L.; Qiao, J. Global evolution of glycosylated polyene macrolide antibiotic biosynthesis. Mol. Phylogenetics Evol. 2018, 127, 239–247. [Google Scholar] [CrossRef]
- Aparicio, J.F.; Caffrey, P.; Gil, J.A.; Zotchev, S.B. Polyene antibiotic biosynthesis gene clusters. Appl. Microbiol. Biotechnol. 2003, 61, 179–188. [Google Scholar] [CrossRef]
- Kämpfer, P.; Glaeser, S.P.; Busse, H.J.; Abdelmohsen, U.R.; Ahmed, S.; Hentschel, U. Actinokineospora spheciospongiae sp. nov., isolated from the marine sponge Spheciospongia vagabunda. Int. J. Syst. Evol. Microbiol. 2015, 65, 879–884. [Google Scholar] [CrossRef]
- Harjes, J.; Ryu, T.; Abdelmohsen, U.R.; Moitinho-Silva, L.; Horn, H.; Ravasi, T.; Hentschel, U. Draft genome sequence of the antitrypanosomally active sponge-associated bacterium Actinokineospora sp. Strain EG49. Genome Announc. 2014, 2, e00160-14. [Google Scholar] [CrossRef] [Green Version]
- Abdelmohsen, U.R.; Pimentel-Elardo, S.M.; Hanora, A.; Radwan, M.; Abou-El-Ela, S.H.; Ahmed, S.; Hentschel, U. Isolation, phylogenetic analysis and anti-infective activity screening of marine sponge-associated actinomycetes. Mar. Drugs 2010, 8, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreales, E.G.; Rumbero, Á.; Payero, T.D.; de Pedro, A.; Jambrina, E.; Aparicio, J.F. Structural and bioactivity characterization of filipin derivatives from engineered Streptomyces filipinensis strains reveals clues for reduced haemolytic action. Antibiotics 2020, 9, 413. [Google Scholar] [CrossRef]
- Silverstein, R.M.; Bassler, G.C. Spectrometric identification of organic compounds. J. Chem. Educ. 1962, 39, 218–250. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. AntiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Brautaset, T.; Sekurova, O.N.; Sletta, H.; Ellingsen, T.E.; StrŁm, A.R.; Valla, S.; Zotchev, S.B. Biosynthesis of the polyene antifungal antibiotic nystatin in Streptomyces noursei ATCC 11455: Analysis of the gene cluster and deduction of the biosynthetic pathway. Chem. Biol. 2000, 7, 395–403. [Google Scholar] [CrossRef] [Green Version]
- Cao, B.; Yao, F.; Zheng, X.; Cui, D.; Shao, Y.; Zhu, C.; Deng, Z.; You, D. Genome mining of the biosynthetic gene cluster of the polyene macrolide antibiotic tetramycin and characterization of a P450 monooxygenase involved in the hydroxylation of the tetramycin B polyol segment. Chembiochem: A Eur. J. Chem. Biol. 2012, 13, 2234–2242. [Google Scholar] [CrossRef] [PubMed]
- Caffrey, P.; Lynch, S.; Flood, E.; Finnan, S.; Oliynyk, M. Amphotericin biosynthesis in Streptomyces nodosus: Deductions from analysis of polyketide synthase and late genes. Chem. Biol. 2001, 8, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Del Vecchio, F.; Petkovic, H.; Kendrew, S.G.; Low, L.; Wilkinson, B.; Lill, R.; Cortés, J.; Rudd, B.A.; Staunton, J.; Leadlay, P.F. Active-site residue, domain and module swaps in modular polyketide synthases. J. Ind. Microbiol. Biotechnol. 2003, 30, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Gaitatzis, N.; Silakowski, B.; Kunze, B.; Nordsiek, G.; Blöcker, H.; Höfle, G.; Müller, R. The biosynthesis of the aromatic myxobacterial electron transport inhibitor stigmatellin is directed by a novel type of modular polyketide synthase. J. Biol. Chem. 2002, 277, 13082–13090. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Hertweck, C. Iteration as programmed event during polyketide assembly; molecular analysis of the aureothin biosynthesis gene cluster. Chem. Biol. 2003, 10, 1225–1232. [Google Scholar] [CrossRef] [Green Version]
- Um, S.; Guo, H.; Thiengmag, S.; Benndorf, R.; Murphy, R.; Rischer, M.; Braga, D.; Poulsen, M.; de Beer, Z.W.; Lackner, G.; et al. Comparative genomic and metabolic analysis of Streptomyces sp. RB110 morphotypes illuminates genomic rearrangements and formation of a new 46-membered antimicrobial macrolide. Acs Chem. Biol. 2021, 16, 1482–1492. [Google Scholar] [CrossRef]
- Simunovic, V.; Müller, R. 3-hydroxy-3-methylglutaryl-CoA-like synthases direct the formation of methyl and ethyl side groups in the biosynthesis of the antibiotic myxovirescin A. Chembiochem: A Eur. J. Chem. Biol. 2007, 8, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Hertweck, C. The biosynthetic logic of polyketide diversity. Angew. Chem. (Int. Ed. Engl.) 2009, 48, 4688–4716. [Google Scholar] [CrossRef]
- Keatinge-Clay, A.T. A tylosin ketoreductase reveals how chirality is determined in polyketides. Chem. Biol. 2007, 14, 898–908. [Google Scholar] [CrossRef] [Green Version]
- Essig, S.; Bretzke, S.; Müller, R.; Menche, D. Full stereochemical determination of ajudazols A and B by bioinformatics gene cluster analysis and total synthesis of ajudazol B by an asymmetric ortholithiation strategy. J. Am. Chem. Soc. 2012, 134, 19362–19365. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, M.; Wu, C.; Tan, Y.; Li, J.; Hao, X.; Duan, Y.; Guan, Y.; Shang, X.; Wang, Y.; et al. Identification and proposed relative and absolute configurations of niphimycins C-E from the marine-derived Streptomyces sp. IMB7-145 by Genomic Analysis. J. Nat. Prod. 2018, 81, 178–187. [Google Scholar] [CrossRef]
- Ishida, K.; Lincke, T.; Hertweck, C. Assembly and absolute configuration of short-lived polyketides from Burkholderia thailandensis. Angew. Chem. (Int. Ed. Engl.) 2012, 51, 5470–5474. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Li, Y.; Zhu, J.; Shen, Y.; Wang, H. Targeted discovery of the polyene macrolide hexacosalactone A from Streptomyces by reporter-guided selection of fermentation media. J. Nat. Prod. 2021, 84, 1924–1929. [Google Scholar] [CrossRef] [PubMed]
- de Sá, J.D.M.; Pereira, J.A.; Dethoup, T.; Cidade, H.; Sousa, M.E.; Rodrigues, I.C.; Costa, P.M.; Mistry, S.; Silva, A.M.S.; Kijjoa, A. Anthraquinones, diphenyl ethers, and their derivatives from the culture of the marine sponge-associated fungus Neosartorya spinosa KUFA 1047. Mar. Drugs 2021, 19, 457. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, Y.Y.; Peng, F.; Duan, W.T.; Wu, C.H.; Li, H.T.; Zhang, X.F.; Shi, Y.S. Neolignans and diarylheptanoids with anti-inflammatory activity from the rhizomes of Alpinia zerumbet. J. Agric. Food Chem. 2021, 69, 9229–9237. [Google Scholar] [CrossRef]
- Tawfike, A.; Attia, E.Z.; Desoukey, S.Y.; Hajjar, D.; Makki, A.A.; Schupp, P.J.; Edrada-Ebel, R.; Abdelmohsen, U.R. New bioactive metabolites from the elicited marine sponge-derived bacterium Actinokineospora spheciospongiae sp. nov. AMB Express 2019, 9, 12. [Google Scholar] [CrossRef]
- Takasaka, N.; Kaweewan, I.; Ohnishi-Kameyama, M.; Kodani, S. Isolation of a new antibacterial peptide actinokineosin from Actinokineospora spheciospongiae based on genome mining. Lett. Appl. Microbiol. 2017, 64, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Stephens, P.J.; Harada, N. ECD cotton effect approximated by the Gaussian curve and other methods. Chirality 2010, 22, 229–233. [Google Scholar] [CrossRef]
- Bonev, B.; Hooper, J.; Parisot, J. Principles of assessing bacterial susceptibility to antibiotics using the agar diffusion method. J. Antimicrob. Chemother. 2008, 61, 1295–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arendrup, M.; Meletiadis, J.; Mouton, J.; Lagrou, K.; Hamal, P. Guinea, Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for conidia forming moulds. Eucast Defin. Doc. Edef 9.3. 2015, 9, 1–23. [Google Scholar]
- 45. EUCAST method for susceptibility testing of moulds, version 9.3.2: Breakpoint tables for interpretation of MICs for antifungal agents, version 10.0. Eur. Comm. Antimicrob. Susceptibility Test. (Eucast) 2020. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Files/EUCAST_E_Def_9.3.2_Mould_testing_definitive_revised_2020.pdf (accessed on 19 November 2021).
- Arendrup, M.C.; Cuenca-Estrella, M.; Lass-Flörl, C.; Hope, W. EUCAST technical note on the EUCAST definitive document EDef 7.2: Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for yeasts EDef 7.2 (EUCAST-AFST). Clin. Microbiol. Infect. 2012, 18, E246–E247. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Higgins, D.G. Clustal Omega, accurate alignment of very large numbers of sequences. Methods Mol. Biol. 2014, 1079, 105–116. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carcon no. | Actinospene (1) | |||
|---|---|---|---|---|
| δC | δH (J in Hz) | COSY | HMBC | |
| 1 | 166.6, C | |||
| 2 | 50.5, CH | 3.64, da | 3 | 1, 3, 4 |
| 3 | 56.7, CH | 2.45, dda | 2, 4 | 4 |
| 4 | 54.4, CH | 2.33, d (7.6) | 3, 5 | 5 |
| 5 | 53.3, CH | 2.59, m | 4, 6 | 3, 6 |
| 6 | 39.9, CH2 | 1.95, ma | 5, 7 | 5, 7 |
| 0.94, m | 5, 7 | |||
| 7 | 66.0, CH | 3.98, m | 6, 8 | |
| 8 | 42.7, CH2 | 1.84, ma | 7 | 7, 9 |
| 1.39, d (13.9) | 7 | 7, 9 | ||
| 9 | 98.5, C | |||
| 10 | 75.7, CH | 2.82, d (8.8) | 11 | 8, 11 |
| 11 | 69.8, CH | 3.73, dd (8.8, 10.8) | 10, 12 | 10, 12, 29 |
| 12 | 56.9, CH | 1.95, ma | 11, 13 | 11, 13, 14, 29 |
| 13 | 64.8, CH | 4.20, m | 12, 14 | 15 |
| 14 | 37.0, CH2 | 2.19, d (13.1) | 13, 15 | |
| 1.49, m | 13, 15 | |||
| 15 | 74.6, CH | 4.37, m | 14, 16 | |
| 16 | 136.4, CH | 5.87, dd (8.6, 14.9) | 15, 17 | 18 |
| 17 | 128.6, CH | 6.07, dd (11.0, 14.9) | 16, 18 | 15, 18, 19 |
| 18 | 129.3, CH | 6.47, dd (11.0, 15.0) | 17, 19 | 16, 17, 19, 20 |
| 19 | 135.6, CH | 6.23, d (15.0) | 18 | 17, 18, 20, 21, 30 |
| 20 | 134.6, C | |||
| 21 | 129.9, CH | 6.00, d (12.0) | 22 | 19, 22, 23, 30 |
| 22 | 128.8, CH | 6.33, dd (12.0, 13.7) | 21, 23 | 20, 21, 24 |
| 23 | 136.3, CH | 5.33, dd (9.5, 13.7) | 22, 24 | 21, 24, 31 |
| 24 | 39.5, CH | 2.54, ma | 23, 25, 31 | |
| 25 | 82.6, CH | 4.85, d (10.4) | 24 | 1, 23, 24, 26, 27, 31, 32 |
| 26 | 131.5, C | |||
| 27 | 126.3, CH | 5.57, q (6.5) | 28 | 25, 28, 32 |
| 28 | 13.0, CH3 | 1.59, d (6.5) | 27 | 26, 27 |
| 29 | 174.6, C | |||
| 30 | 12.7, CH3 | 1.84, s | 19, 20, 21 | |
| 31 | 16.3, CH3 | 0.81, d (6.2) | 24 | 23, 24, 25 |
| 32 | 10.6, CH3 | 1.55, s | 25, 26, 27 | |
| 1’ | 96.5, CH | 4.41, s | 2’ | 15, 2’ |
| 2’ | 69.8, CH | 3.64, ma | 1’, 3’ | 3’, 4’ |
| 3’ | 72.9, CH | 3.22, d (8.9) | 2’, 4’ | |
| 4’ | 54.6, CH | 2.46, ma | 3’, 5’ | 3’, 5’, 6’ |
| 5’ | 72.0, CH | 3.08, m | 4’, 6’ | |
| 6’ | 18.3, CH3 | 1.15, d (5.5) | 5’ | 4’, 5’ |
| Species | MIC (μg/mL) | ||
|---|---|---|---|
| Actinospene | Pimaricin | Amphotericin B | |
| Saccharomyces cerevisiae | 2 | 2 | 0.25 |
| Candida albicans | 8 | 2 | 0.25 |
| Cryptococcus neoformans | 2 | 2 | 0.25 |
| Fusariums oxysporum | 10 | 11 | 5 |
| Sclerotium rolfsi | 2 | 1.38 | 0.02 |
| Fusarium graminearum | 8 | 1.38 | 0.63 |
| Colletotrichum capsici | 50 | 11 | 2.5 |
| Alternaria alternate | 8 | 5.5 | 0.16 |
| Phytophthora capsici | 4 | 5.5 | 0.16 |
| Staphylococcus aureus | >64 | NT | NT |
| Escherichia coli | >64 | NT | NT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Zhang, C.; Cui, T.; Lei, P.; Guo, Z.; Wang, H.; Liu, Q. The Discovery of Actinospene, a New Polyene Macrolide with Broad Activity against Plant Fungal Pathogens and Pathogenic Yeasts. Molecules 2021, 26, 7020. https://doi.org/10.3390/molecules26227020
Tang Y, Zhang C, Cui T, Lei P, Guo Z, Wang H, Liu Q. The Discovery of Actinospene, a New Polyene Macrolide with Broad Activity against Plant Fungal Pathogens and Pathogenic Yeasts. Molecules. 2021; 26(22):7020. https://doi.org/10.3390/molecules26227020
Chicago/Turabian StyleTang, Ying, Cuiyang Zhang, Tianqi Cui, Ping Lei, Zhaohui Guo, Hailong Wang, and Qingshu Liu. 2021. "The Discovery of Actinospene, a New Polyene Macrolide with Broad Activity against Plant Fungal Pathogens and Pathogenic Yeasts" Molecules 26, no. 22: 7020. https://doi.org/10.3390/molecules26227020