
Mentha pulegium L.: A Plant Underestimated for Its Toxicity to Be Recovered from the Perspective of the Circular Economy

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results and Discussion
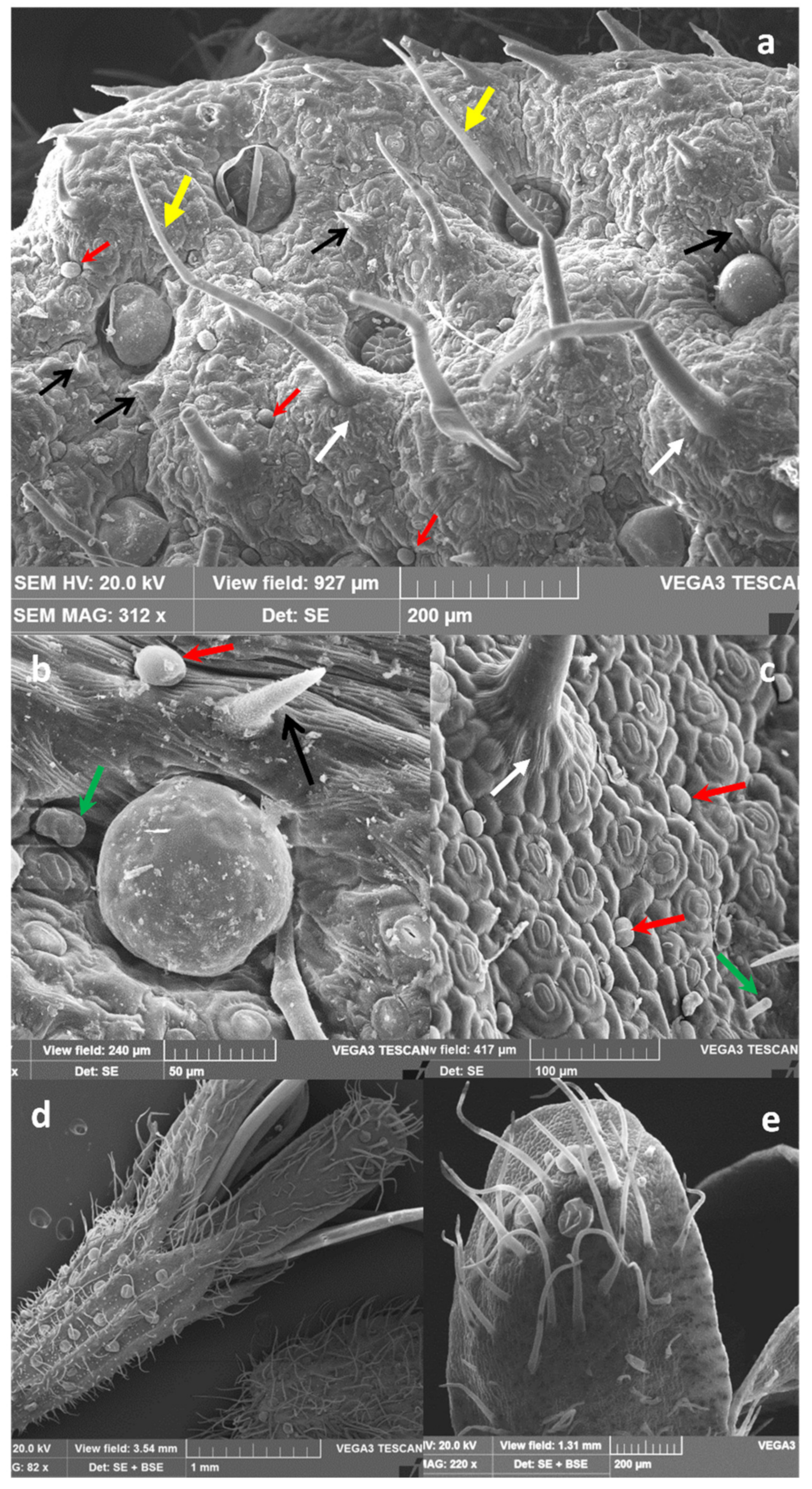
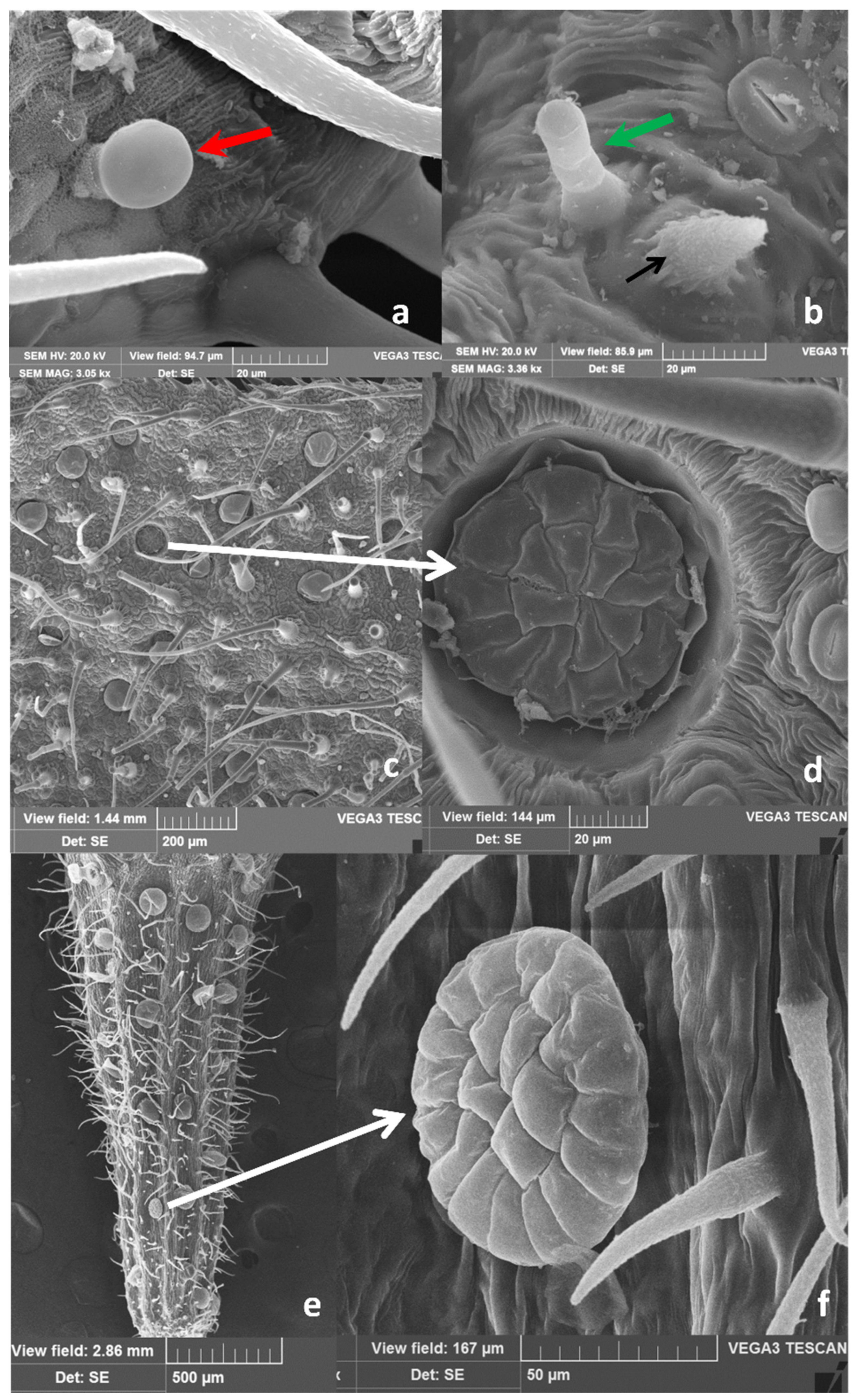
2.1. Micromorphological Studies and Essential Oil Yield
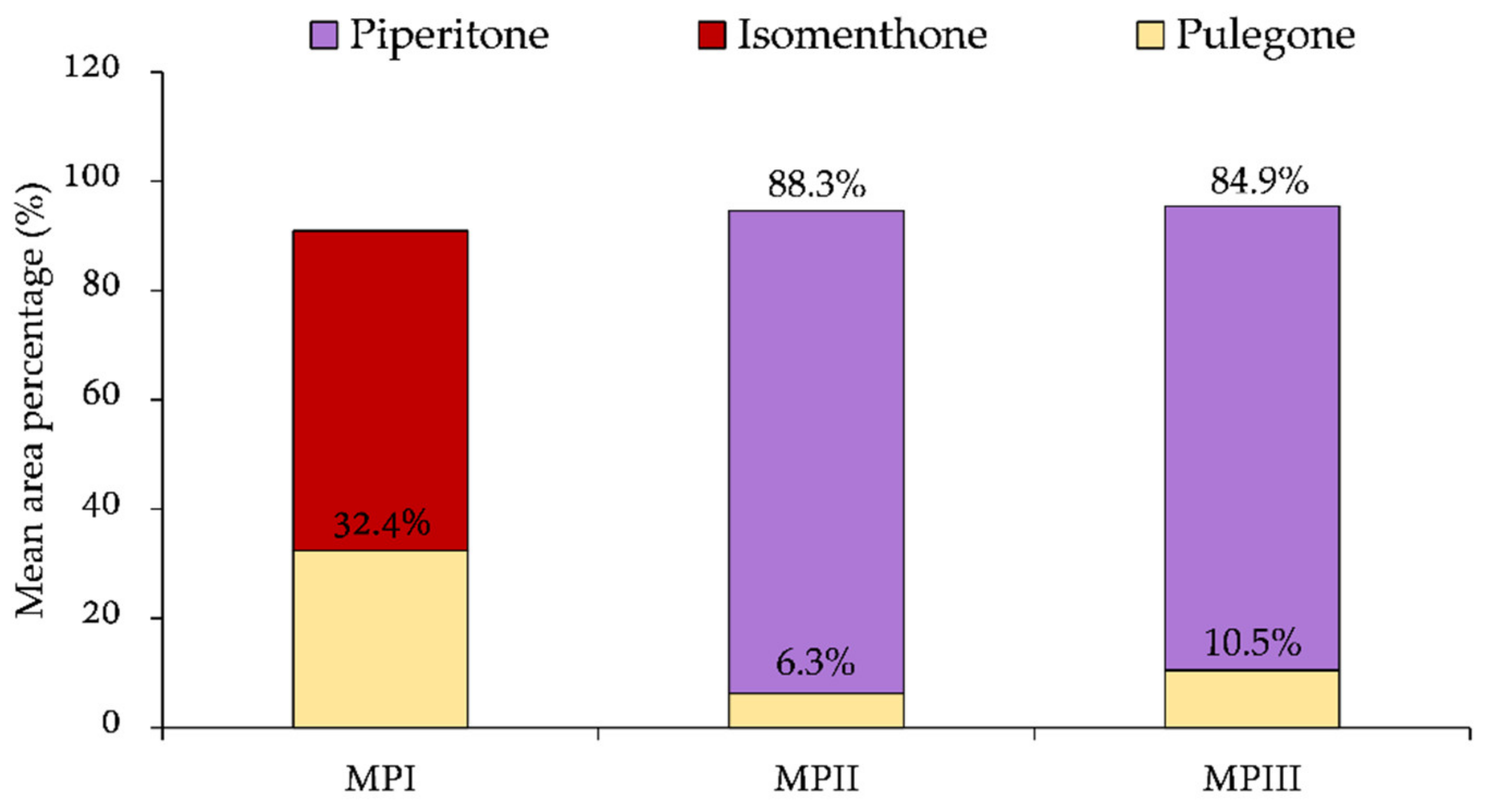
2.2. Essential Oil Composition
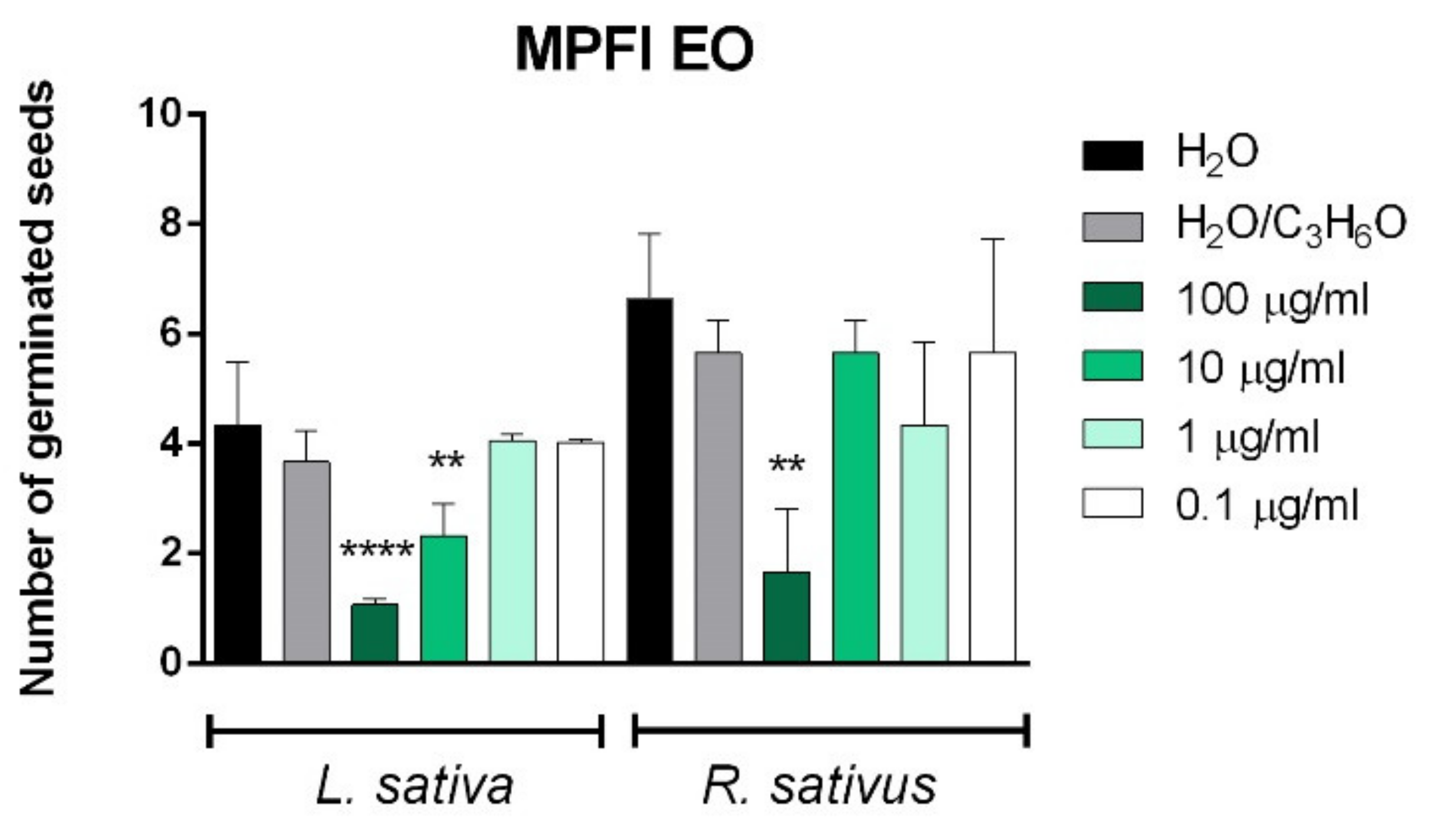
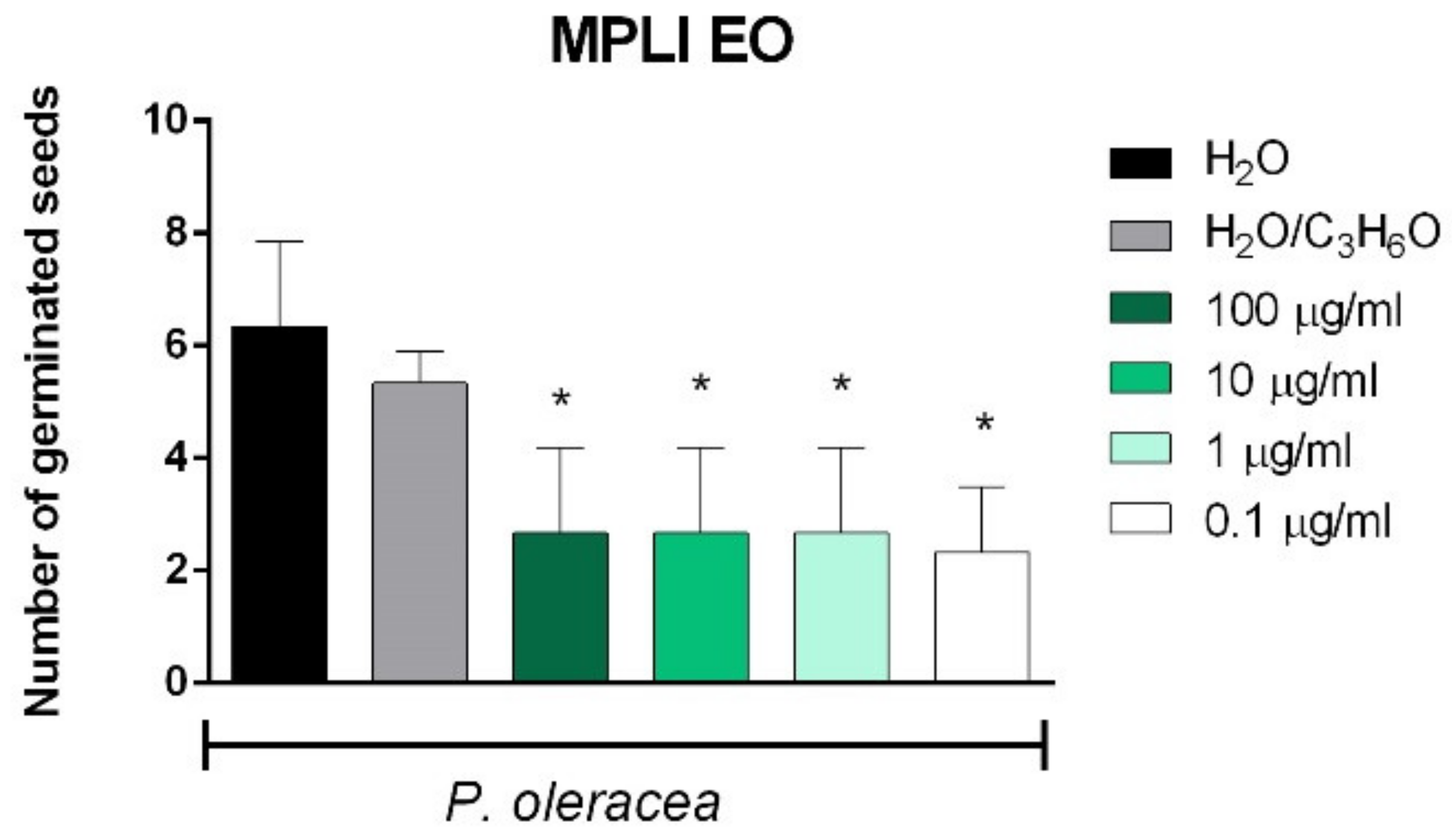
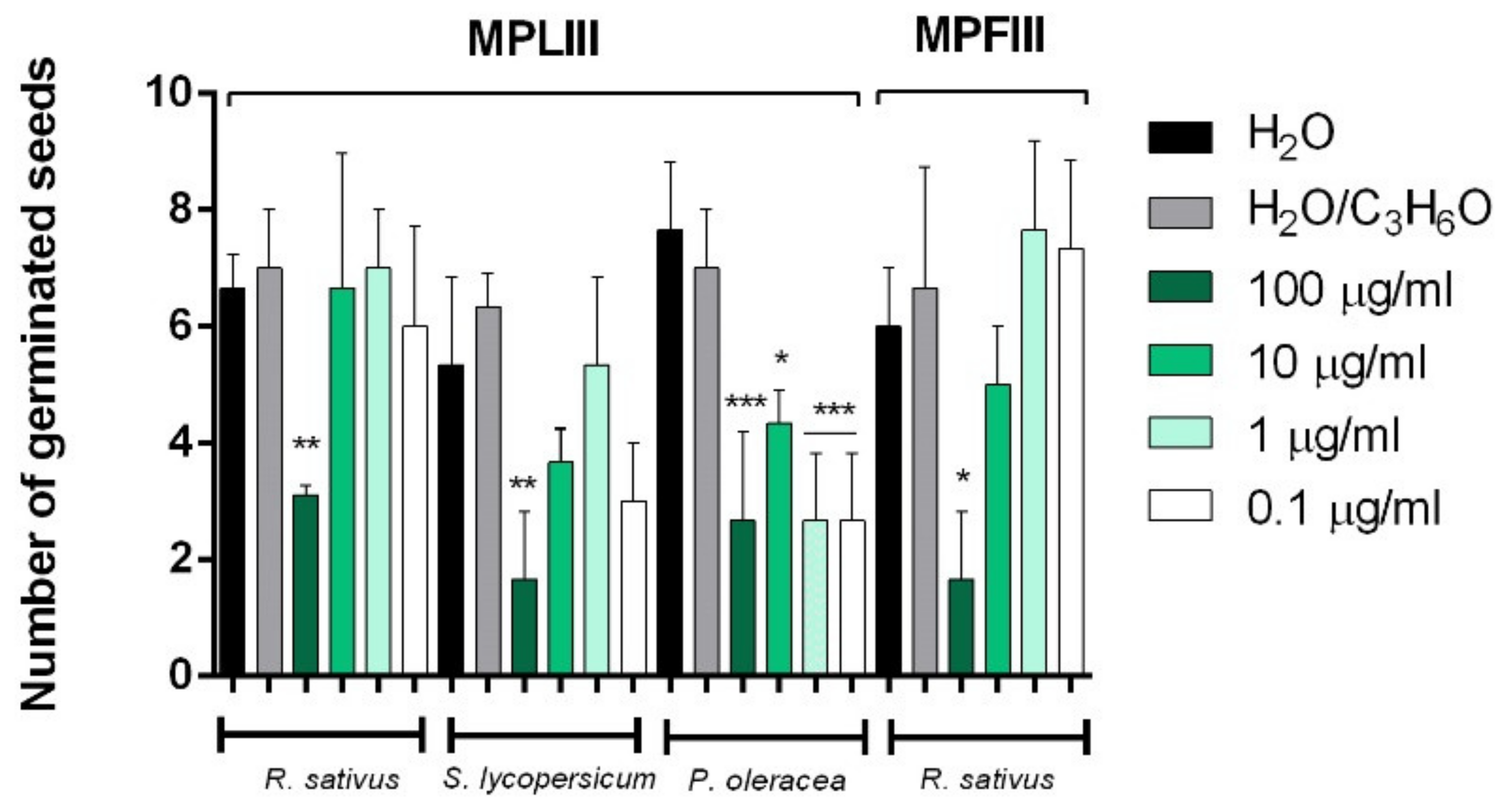
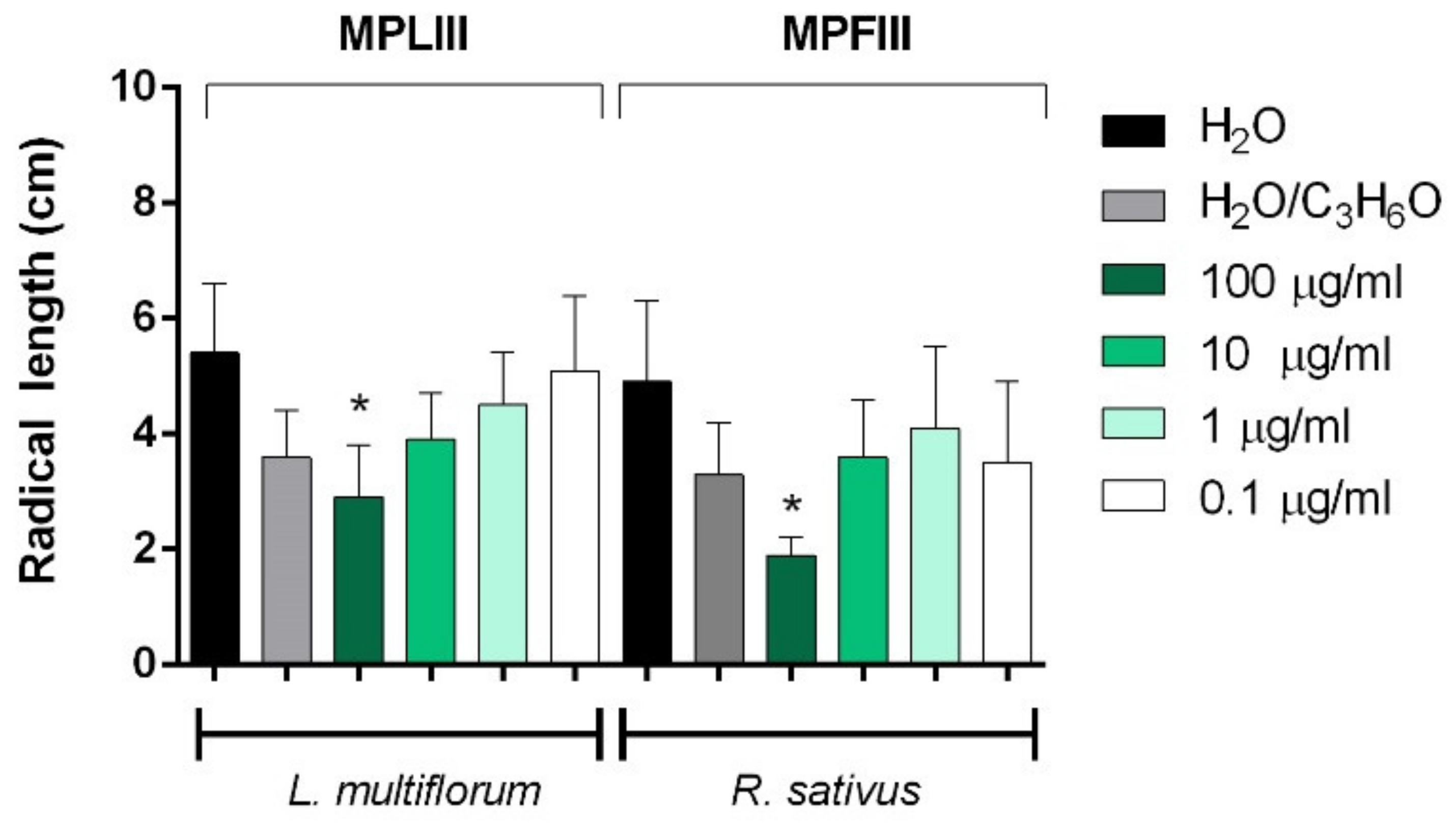
2.3. Phytotoxic and Anti-α-Amylase Activity
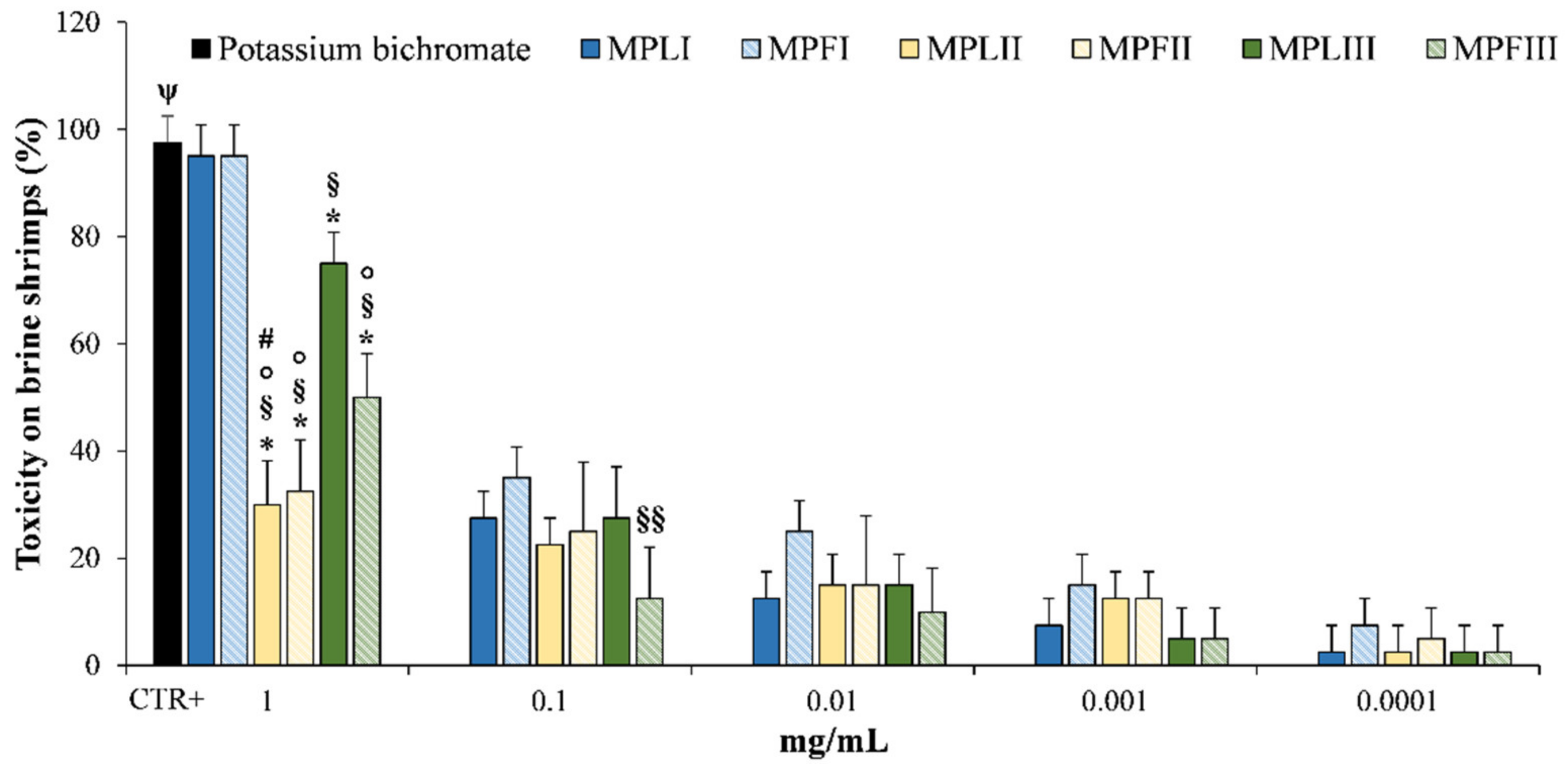
2.4. Toxicity
3. Materials and Methods
3.1. Standards and Reagents
3.2. Plant Material and Sample Preparation
3.3. Micromorphological Analyses
3.4. Phytochemical Characterization
3.5. Phytotoxic Activity
3.6. Alpha-Amylase Inhibitory Assay
3.7. Brine Shrimp Lethality Assay
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Ash, M.; Ash, I. Industrial Chemical Thesaurus, 5th ed.; Synapse Information Resources: Endicott, NY, USA, 2009. [Google Scholar]
- Chalchat, J.-C.; Gorunovic, M.S.; Maksimovic, Z.A.; Petrovic, S.D. Essential Oil of Wild Growing Mentha pulegium L. from Yugoslavia. J. Essent. Oil Res. 2000, 12, 598–600. [Google Scholar] [CrossRef]
- Harley, R.M.; Atkins, S.; Budantsev, A.L.; Cantino, P.D.; Conn, B.J.; Grayer, R.; De Kok, R.; Krestovskaja, T.; Morales, R.; Paton, A.J.; et al. Labiatae. In Flowering Plants Dicotyledons; Springer Science and Business Media LLC, 2004; Vol. 7, pp. 167–275. [Google Scholar]
- Van De Walle, E.; Riddle, J.M. Contraception and Abortion from the Ancient World to the Renaissance. Popul. Dev. Rev. 1994, 20, 221. [Google Scholar] [CrossRef]
- Gordon, P.; Khojasteh, S.C. A decades-long investigation of acute metabolism-based hepatotoxicity by herbal constituents: a case study of pennyroyal oil. Drug Metab. Rev. 2014, 47, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Nickavar, B.; Jabbareh, F. Analysis of the essential oil from Mentha pulegium and identification of its antioxidant constituents. J. Essent. Oil Bear. Plant 2018, 21, 223–229. [Google Scholar] [CrossRef]
- Teixeira, B.; Marques, A.; Ramos, C.; Batista, I.; Serrano, C.; Matos, O.; Neng, N.R.; Nogueira, J.M.; Saraiva, J.A.; Nunes, M.L. European pennyroyal (Mentha pulegium) from Portugal: Chemical composition of essential oil and antioxidant and antimicrobial properties of extracts and essential oil. Ind. Crop. Prod. 2012, 36, 81–87. [Google Scholar] [CrossRef]
- Carmichael, P.G. Pennyroyal Metabolites in Human Poisoning. Ann. Intern. Med. 1997, 126, 250. [Google Scholar] [CrossRef]
- Sullivan, J.B.; Rumack, B.H.; Thomas, H.; Peterson, R.G.; Bryson, P. Pennyroyal Oil Poisoning and Hepatotoxicity. JAMA 1979, 242, 2873–2874. [Google Scholar] [CrossRef]
- Anderson, I.B.; Mullen, W.H.; Meeker, J.E.; Khojasteh-Bakht, S.C.; Oishi, S.; Nelson, S.D.; Blanc, P.D. Pennyroyal Toxicity: Measurement of Toxic Metabolite Levels in Two Cases and Review of the Literature. Ann. Intern. Med. 1996, 124, 726. [Google Scholar] [CrossRef]
- Bakerink, J.A.; Gospe, S.M.; Dimand, R.J.; Eldridge, M.W. Multiple organ failure after ingestion of pennyroyal oil from herbal tea in two infants. Pediatrics 1996, 98, 944–947. [Google Scholar]
- Da Rocha, M.S.; Dodmane, P.R.; Arnold, L.L.; Pennington, K.L.; Anwar, M.M.; Adams, B.R.; Taylor, S.V.; Wermes, C.; Cohen, S.M. Mode of Action of Pulegone on the Urinary Bladder of F344 Rats. Toxicol. Sci. 2012, 128, 1–8. [Google Scholar] [CrossRef] [Green Version]
- National Toxicology Program. Toxicology and carcinogenesis studies of pulegone (CAS No. 89-82-7) in F344/N rats and B6C3F1 mice (gavage studies). Natl. Toxicol. Program. Tech. Rep. Ser. 2011, 563, 1–201. [Google Scholar]
- Cohen, S.M.; Eisenbrand, G.; Fukushima, S.; Gooderham, N.J.; Guengerich, F.P.; Hecht, S.S.; Rietjens, I.; Rosol, T.; Harman, C.; Taylor, S.V. GRAS 29 flavoring substances. Food Technol. 2020, 74, 44–65. [Google Scholar]
- Maia, M.F.; Moore, S.J. Plant-based insect repellents: a review of their efficacy, development and testing. Malar. J. 2011, 10, S11–S14. [Google Scholar] [CrossRef] [Green Version]
- Werker, E. Function of essential oil-secreting glandular hairs in aromatic plans of Lamiacea—a review. Flavour Fragr. J. 1993, 8, 249–255. [Google Scholar] [CrossRef]
- Martins, M.B.G. Optical microscopy studies and scanning electron microscopy in leaf the Mentha spicata and Mentha spicata X suaveolens (Lamiaceae). Bragantia 2002, 61, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, L.; Póvoa, O.; Teixeira, G.; Figueiredo, A.C.; Moldão, M.; Monteiro, A. Trichomes micromorphology and essential oil variation at different developmental stages of cultivated and wild growing Mentha pulegium L. populations from Portugal. Ind. Crop. Prod. 2013, 43, 692–700. [Google Scholar] [CrossRef]
- Yu, X.; Liang, C.; Fang, H.; Qi, X.; Li, W.; Shang, Q. Variation of trichome morphology and essential oil composition of seven Mentha species. Biochem. Syst. Ecol. 2018, 79, 30–36. [Google Scholar] [CrossRef]
- Karray-Bouraoui, N.; Rabhi, M.; Neffati, M.; Baldan, B.; Ranieri, A.; Marzouk, B.; Lachaâl, M.; Smaoui, A. Salt effect on yield and composition of shoot essential oil and trichome morphology and density on leaves of Mentha pulegium. Ind. Crop. Prod. 2009, 30, 338–343. [Google Scholar] [CrossRef]
- Ascensão, L.; Pais, M.S. The leaf capitate trichomes of Leonotis leonurus: Histochemistry, ultrastructure and secretion. Ann. Bot. 1998, 81, 263–271. [Google Scholar] [CrossRef]
- Maffei, M.; Chialva, F.; Sacco, T. Glandular trichomes and essential oils in developing peppermint leaves. I. Variation of peltate trichome number and terpene distribution within leaves. New Phytol. 1989, 111, 707–716. [Google Scholar] [CrossRef]
- Turner, G.W.; Gershenzon, J.; Croteau, R.B. Distribution of Peltate Glandular Trichomes on Developing Leaves of Peppermint. Plant Physiol. 2000, 124, 655–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, A.I.; Anwar, F.; Nigam, P.S.; Ashraf, M.; Gilani, A.H. Seasonal variation in content, chemical composition and antimicrobial and cytotoxic activities of essential oils from four Mentha species. J. Sci. Food Agric. 2010, 90, 1827–1836. [Google Scholar] [CrossRef] [PubMed]
- Kokkini, S.; Hanlidou, E.; Karousou, R.; Lanaras, T. Clinal Variation of Mentha pulegium Essential Oils Along the Climatic Gradient of Greece. J. Essent. Oil Res. 2004, 16, 588–593. [Google Scholar] [CrossRef]
- Benayad, N.; Ebrahim, W.; Hakiki, A.; Mosaddak, M. Chemical characterization and insecticidal evaluation of the essential oil of Mentha suaveolens L. and Mentha pulegium L. growing in Morocco. Sci. Study Res. Chem. Chem. Eng. Biotech. Food Ind. 2012, 13, 27–32. [Google Scholar]
- Silva, L.F.; Cardoso, M.D.G.; Batista, L.R.; Gomes, M.D.S.; Rodrigues, L.M.A.; Rezende, D.A.D.C.S.; Teixeira, M.L.; Carvalho, M.S.S.; Santiago, J.D.A.; Nelson, D.L. Chemical Characterization, Antibacterial and Antioxidant Activities of Essential Oils of Mentha viridis L. and Mentha pulegium L. (L). Am. J. Plant Sci. 2015, 6, 666–675. [Google Scholar] [CrossRef] [Green Version]
- Fancello, F.; Zara, S.; Petretto, G.L.; Chessa, M.; Addis, R.; Rourke, J.P.; Pintore, G. Essential oils from three species of Mentha harvested in Sardinia: chemical characterization and evaluation of their biological activity. Int. J. Food Prop. 2017, 20, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mahboubi, M.; Haghi, G. Antimicrobial activity and chemical composition of Mentha pulegium L. essential oil. J. Ethnopharmacol. 2008, 119, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Khadraoui, A.; Khelifa, A.; Hadjmeliani, M.; Mehdaoui, R.; Hachama, K.; Tidu, A.; Azari, Z.; Obot, I.; Zarrouk, A. Extraction, characterization and anti-corrosion activity of Mentha pulegium oil: Weight loss, electrochemical, thermodynamic and surface studies. J. Mol. Liq. 2016, 216, 724–731. [Google Scholar] [CrossRef]
- El-Ghorab, A.H. The Chemical Composition of the Mentha pulegium L. Essential Oil from Egypt and its Antioxidant Activity. J. Essent. Oil Bear. Plants 2006, 9, 183–195. [Google Scholar] [CrossRef]
- Kasrati, A.; Alaoui Jamali, C.; Bekkouche, K.; Spooner-Hart, R.; Leach, D.; Abbad, A. Chemical characterization and insecti-cidal properties of essential oils from different wild populations of Mentha suaveolens subsp. Timija (Briq.) Harley from Morocco. Chem. Biodivers. 2015, 12, 823–831. [Google Scholar] [CrossRef]
- Ketoh, G.K.; Koumaglo, H.K.; Glitho, I.A.; Huignard, J. Comparative effects of Cymbopogon schoenanthus essential oil and piperitone on Callosobruchus maculatus development. Fitoterapia 2006, 77, 506–510. [Google Scholar] [CrossRef]
- Saleh, M.A.; Belal, M.H.; El-Baroty, G. Fungicidal Activity of Artemisia herba alba Asso (Asteraceae). J. Environ. Sci. Heal. Part B 2006, 41, 237–244. [Google Scholar] [CrossRef]
- Kimbaris, A.C.; González-Coloma, A.; Andrés, M.F.; Vidali, V.P.; Polissiou, M.G.; Santana-Méridas, O. Biocidal compounds from Mentha sp. essential oils and their structure–activity relationships. Chem. Biodivers. 2017, 14, e1600270. [Google Scholar] [CrossRef] [PubMed]
- Santana, O.; Andrés, M.F.; Sanz, J.; Errahmani, N.; Abdeslam, L.; González-Coloma, A. Valorization of Essential Oils from Moroccan Aromatic Plants. Nat. Prod. Commun. 2014, 9, 1109–1114. [Google Scholar] [CrossRef] [Green Version]
- Arteca, R.N. Plant Growth Substances: Principles and Applications; Springer: New York, NY, USA, 2013; pp. 332–347. [Google Scholar]
- Murtaza, G.; Asghar, R. α-Amylase activities during seed development and germination in pea (Pisum sativum L.) treated with salicylic acid. Pak. J. Bot. 2012, 44, 1823–1829. [Google Scholar]
- Figueroa-Pérez, M.G.; Rocha-Guzmán, N.E.; Pérez-Ramírez, I.F.; Mercado-Silva, E.; Reynoso-Camacho, R. Metabolite Profile, Antioxidant Capacity, and Inhibition of Digestive Enzymes in Infusions of Peppermint (Mentha piperita) Grown under Drought Stress. J. Agric. Food Chem. 2014, 62, 12027–12033. [Google Scholar] [CrossRef]
- De Sousa, D.P.; Nóbrega, F.F.; De Lima, M.R.; De Almeida, R.N. Pharmacological activity of (R)-(+)-pulegone, a chemical constituent of essential oils. Z. Nat. C J. Biosci. 2011, 66, 353–359. [Google Scholar]
- Souldouzi, R.; Razi, M.; Shalizar Jalali, A.; Jalilzadeh-Amin, G.; Amani, S. Effect of (R)-(+) Pulegone on ovarian tissue; corre-lation with expression of aromatase Cyp19 and ovarian selected genes in mice. Cell J. 2018, 20, 231–243. [Google Scholar]
- Pharmacopée Européenne 1; Conseil de l’Europe: Sainte Rune, France, 1996.
- Chieco, C.; Rotondi, A.; Morrone, L.; Rapparini, F.; Baraldi, R. An ethanol-based fixation method for anatomical and mi-cro-morphological characterization of leaves of various tree species. Biotech. Histochem. 2013, 88, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; Trombetta, D.; Cornara, L.; Valussi, M.; De Feo, V.; Caputo, L. Characterization and Phytotoxicity Assessment of Essential Oils from Plant Byproducts. Molecules 2019, 24, 2941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NIST 08 Mass Spectral Library (NIST/EPA/NIH). 2008. Available online: www.nist.gov (accessed on 1 February 2021).
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; pp. 1–809. [Google Scholar]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dineshkumar, B.; Mitra, A.; Manjunatha, M. A comparative study of alpha amylase inhibitory activities of common an-ti-diabetic plants at Kharagpur 1 block. IJGP 2010, 4, 115–121. [Google Scholar]
- Caputo, L.; Cornara, L.; Bazzicalupo, M.; De Francesco, C.; De Feo, V.; Trombetta, D.; Smeriglio, A. Chemical Composition and Biological Activities of Essential Oils from Peels of Three Citrus Species. Molecules 2020, 25, 1890. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MP-I | MP-II | MP-III | ||
|---|---|---|---|---|
| Density (n/mm2) | leaf ad. s. | 5.0 ± 1.6# (3) | 2.6 ± 1.0# (3) | 3.6 ± 0.4# (3) |
| leaf ab. s. | 21.6 ± 1.8# (3) | 18.1 ± 3.7# (3) | 15.9 ± 4.3# (3) | |
| fl. calyx | 13.7 ± 2.8* (3) | 22.1 ± 1.6* (3) | 19.0 ± 1.6 (3) | |
| Diameter (μÌm) | leaf | 87.2 ± 3.4*# (16) | 86.4 ± 4.5# (15) | 83.0 ± 3.2*# (15) |
| fl. calyx | 100.1 ± 3.7# (10) | 101.7 ± 4.0# (10) | 104.5 ± 10.2# (10) | |
| EO yield (w/w %) | leaf | 3.2 | 2.7 | 2.9 |
| flower | 2.8 | 2.7 | 2.6 |
| KIa | Identificationb | MPLI | MPFI | MPLII | MPFII | MPLIII | MPFIII | |
|---|---|---|---|---|---|---|---|---|
| 2.5-diethyl tetrahydrofuran | 884 | 1,2 | - | t | - | - | t | - |
| Santolina triene | 908 | 1,2 | - | - | - | - | t | - |
| α-Thujene | 924 | 1,2,3 | - | t | - | t | t | - |
| α-Pinene | 939 | 1,2,3 | 0.1 ± 0.0* | 0.3 ± 0.0* | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | t |
| Camphene | 954 | 1,2,3 | - | t | - | t | t | - |
| Thuja-2,4(10)-diene | 960 | 1,2 | - | - | - | - | t | |
| β-Thujene | 968 | 1,2,3 | t | - | 0.3 ± 0.0* | 0.1 ± 0.0* | t | - |
| Sabinene | 975 | 1,2,3 | - | t | 0.2 ± 0.0§ | 0.2 ± 0.0§ | 0.2 ± 0.0§ | t |
| β-Pinene | 979 | 1,2,3 | 0.1 ± 0.0* | 0.3 ± 0.0* | - | - | - | - |
| 3-Octanone | 983 | 1,2 | 0.1 ± 0.0* | t | - | t | - | - |
| Myrcene | 990 | 1,2,3 | 0.1 ± 0.0 | 0.3 ± 0.0* | 0.1 ± 0.0 | 0.2 ± 0.0* | 0.1 ± 0.0 | - |
| 3-Octanol | 995 | 1,2 | 1.1 ± 0.1* | 0.9 ± 0.0* | 0.8 ± 0.0 | 0.8 ± 0.1 | 0.2 ± 0.0* | - |
| α-Phellandrene | 1002 | 1,2 | - | - | - | t | t | - |
| p-Mentha-1(7),8-diene | 1004 | 1,2 | t | - | - | - | - | - |
| t-Butyl Benzene | 1008 | 1,2 | - | t | - | - | - | - |
| δ-Carene | 1011 | 1,2,3 | - | t | - | - | - | - |
| α-Terpinene | 1017 | 1,2,3 | - | - | - | t | - | - |
| 1,2,3,4-Tetramethylbenzene | 1027 | 1,2 | t | - | - | - | - | |
| Limonene | 1029 | 1,2,3 | 1.7 ± 0.1 | 2.3 ± 0.1* | 1.8 ± 0.1 | 3.6 ± 0.2* | 0.3 ± 0.0* | t |
| 1.8-Cineole | 1031 | 1,2,3 | 0.1 ± 0.0* | t | 0.1 ± 0.0 | 0.1 ± 0.0 | t | - |
| (Z)-β-Ocimene | 1037 | 1,2 | t | t | - | t | - | - |
| γ-Terpinene | 1059 | 1,2,3 | t | - | - | t | - | |
| Terpinolene | 1088 | 1,2,3 | t | t | - | t | t | - |
| trans-Sabinene hydrate | 1098 | 1,2 | t | t | - | t | t | - |
| cis-Thujone | 1102 | 1,2 | t | |||||
| 3-Octanol acetate | 1123 | 1,2 | 0.2 ± 0.0* | - | - | - | - | - |
| Menthone | 1152 | 1,2,3 | 0.8 ± 0.0° | 0.8 ± 0.0° | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.8 ± 0.0° | 0.3 ± 0.0* |
| Isomenthone | 1162 | 1,2,3 | 34.0 ± 1.1* | 30.9 ± 1.4* | 7.2 ± 0.3 | 5.3 ± 0.3* | 13.0 ± 0.9* | 8.0 ± 0.3 |
| Menthol | 1171 | 1,2,3 | 1.0 ± 0.1* | 1.2 ± 0.0* | - | t | 0.5 ± 0.0* | - |
| iso-Menthol | 1182 | 1,2 | 1.3 ± 0.1* | 0.4 ± 0.0* | - | t | t | - |
| α-Terpineol | 1188 | 1,2,3 | - | - | - | t | 0.2 ± 0.0 | - |
| Isopulegone | 1229 | 1,2 | 0.1 ± 0.0¥ | t | - | - | 0.1 ± 0.0¥ | - |
| Pulegone | 1237 | 1,2,3 | 56.8 ± 2.2Φ | 60.2 ± 1.9Φ | - | - | 0.2 ± 0.0* | - |
| (2Z)-hexenyl isovalerate | 1244 | 1,2 | - | - | - | t | - | - |
| (2E)-hexenyl valerate | 1247 | 1,2 | - | - | t | t | - | - |
| Piperitone | 1252 | 1,2,3 | 0.5 ± 0.0* | 0.7 ± 0.0* | 88.5 ± 2.3 | 88.0 ± 2.9 | 80.3 ± 1.7* | 89.4 ± 1.6 |
| cis-Carvone oxide | 1263 | 1,2 | t | - | - | - | - | - |
| neo-Menthyl acetate | 1272 | 1,2 | 0.1 ± 0.0* | - | - | - | - | - |
| trans-Carvone oxide | 1276 | 1,2 | t | - | - | - | 0.2 ± 0.0 | - |
| neo-Menthyl acetate | 1278 | 1,2 | 0.4 ± 0.0* | t | - | - | 0.1 ± 0.0* | - |
| iso-Menthyl acetate | 1305 | 1,2 | 0.6 ± 0.0* | 0.8 ± 0.0* | - | 0.3 ± 0.0* | 2.9 ± 0.1* | 1.9 ± 0.1* |
| α-Copaene | 1376 | 1,2,3 | t | t | - | t | t | - |
| β-bourbonene | 1385 | 1,2 | t | - | t | t | t | - |
| β-Cubebene | 1388 | 1,2 | t | t | - | - | - | - |
| (E)-Caryophyllene | 1419 | 1,2,3 | 0.1 ± 0.0* | t | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.1 ± 0.0* |
| β-Copaene | 1432 | 1,2,3 | t | t | - | - | - | |
| α-Humulene | 1454 | 1,2,3 | 0.1 ± 0.0* | t | 0.2 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | t |
| Germacrene D | 1481 | 1,2 | 0.4 ± 0.0* | 0.3 ± 0.0* | t | t | t | - |
| α-selinene | 1498 | 1,2 | - | t | - | - | - | - |
| δ-Cadinene | 1523 | 1,2,3 | t | t | - | - | - | - |
| α-Cadinol | 1654 | 1,2,3 | t | - | - | - | - | - |
| (Z.Z)-Farnesyl acetone | 1860 | 1,2 | - | t | - | - | - | - |
| Monoterpene hydrocarbons Monoterpenes oxigenated Sesquiterpene hydrocarbons Others | 2.2 | 3.4 | 2.6 | 4.4 | 0.9 | 0.2 | ||
| 92.2 | 92.8 | 96.0 | 93.7 | 94.5 | 97.7 | |||
| 0.8 | 0.4 | 0.5 | 0.4 | 0.4 | 0.2 | |||
| 4.8 | 3.5 | 0.9 | 1.5 | 4.2 | 1.9 | |||
| Essential Oils | IC50 Values (µg/mL) |
|---|---|
| MPLI | 662.8 ± 5.8 |
| MPFI | 785.3 ± 7.1 |
| MPLII | N.A. |
| MPFII | 1076.7 ± 8.9 |
| MPLIII | 830.9 ± 6.3 |
| MPFIII | 638.9 ± 9.1 |
| Acarbose (positive control) | 109.3 ± 5.6 |
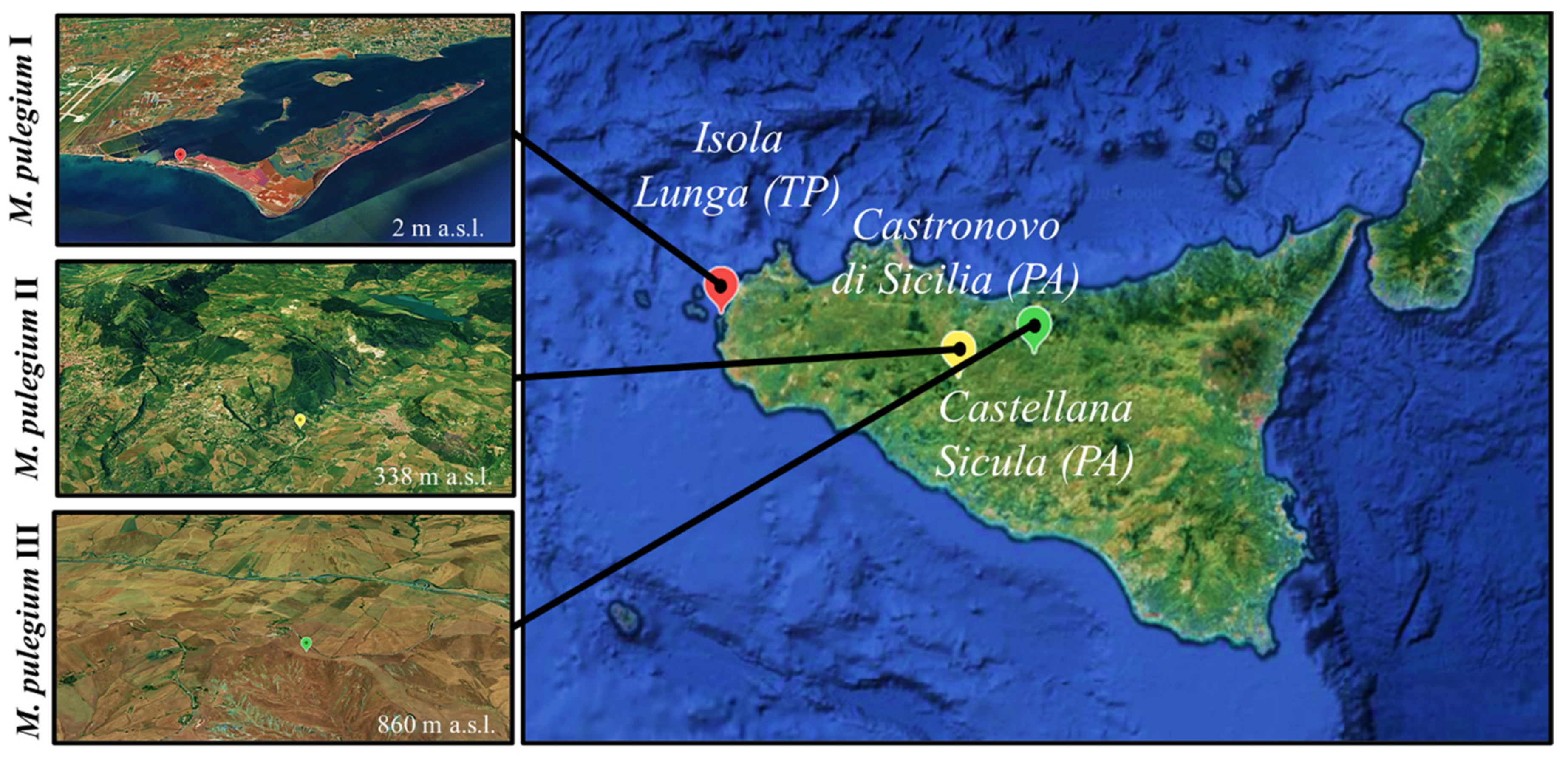
| Collection Site | GPS Coordinates | Site Features | Voucher Specimens |
|---|---|---|---|
| I: Isola Lunga (TP) | 37°54′06″ N, 12°27′07″ E | Clayey-saline depressions | R&Sp * 05/19 |
| II: Castronovo di Sicilia (PA) | 37°40′09″ N, 13°38′54″ E | Clayey-moist soil | R&Sc § 06/19 |
| III: Castellana sicula (PA) | 37°45′35″ N, 13°59′35″ E | Clayey-moist soil | R&Sc § 07/19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caputo, L.; Cornara, L.; Raimondo, F.M.; De Feo, V.; Vanin, S.; Denaro, M.; Trombetta, D.; Smeriglio, A. Mentha pulegium L.: A Plant Underestimated for Its Toxicity to Be Recovered from the Perspective of the Circular Economy. Molecules 2021, 26, 2154. https://doi.org/10.3390/molecules26082154
Caputo L, Cornara L, Raimondo FM, De Feo V, Vanin S, Denaro M, Trombetta D, Smeriglio A. Mentha pulegium L.: A Plant Underestimated for Its Toxicity to Be Recovered from the Perspective of the Circular Economy. Molecules. 2021; 26(8):2154. https://doi.org/10.3390/molecules26082154
Chicago/Turabian StyleCaputo, Lucia, Laura Cornara, Francesco Maria Raimondo, Vincenzo De Feo, Stefano Vanin, Marcella Denaro, Domenico Trombetta, and Antonella Smeriglio. 2021. "Mentha pulegium L.: A Plant Underestimated for Its Toxicity to Be Recovered from the Perspective of the Circular Economy" Molecules 26, no. 8: 2154. https://doi.org/10.3390/molecules26082154