
In Silico Analysis of P450s and Their Role in Secondary Metabolism in the Bacterial Class Gammaproteobacteria

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Only Few Gammaproteobacterial Species Have P450s
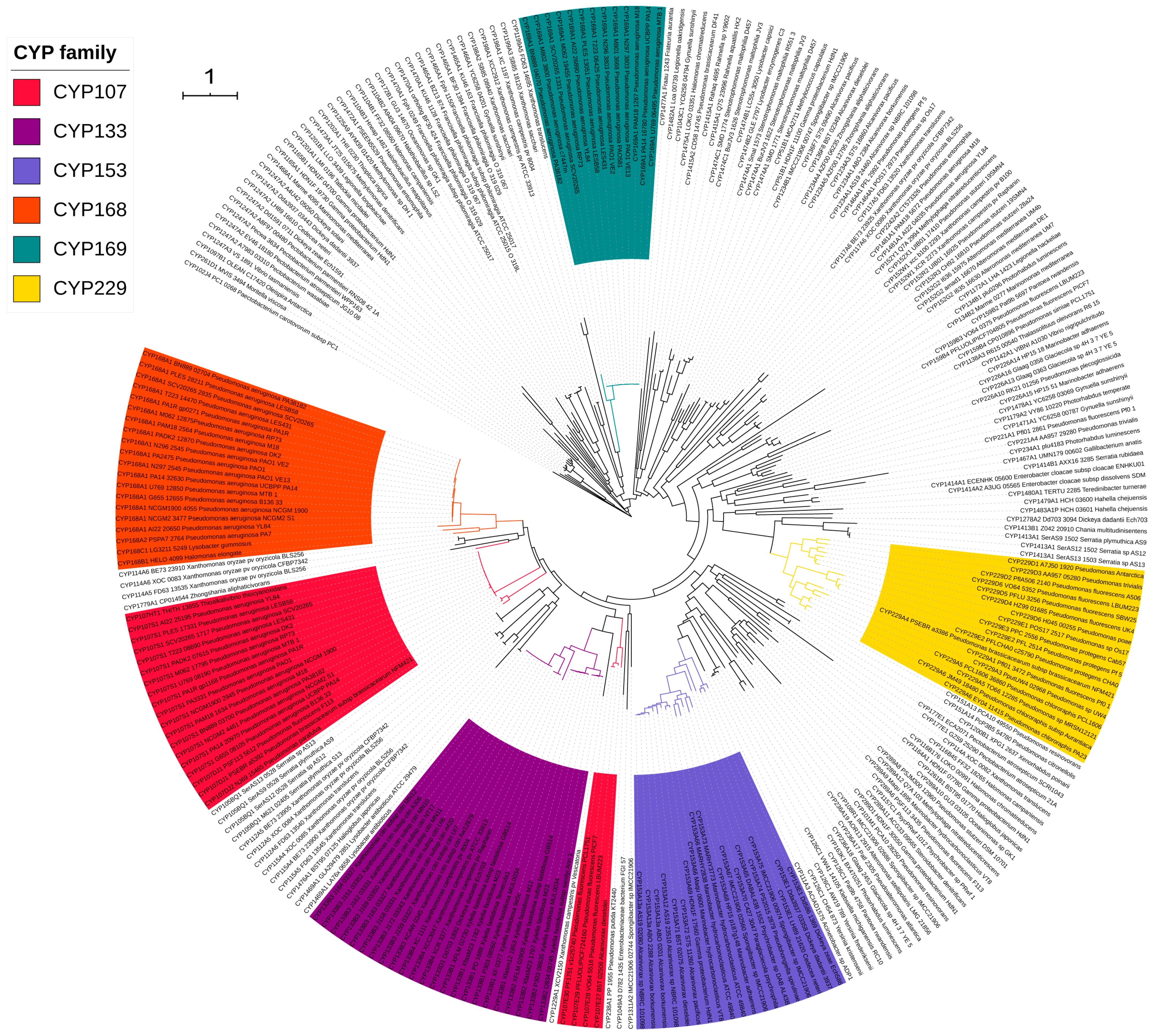
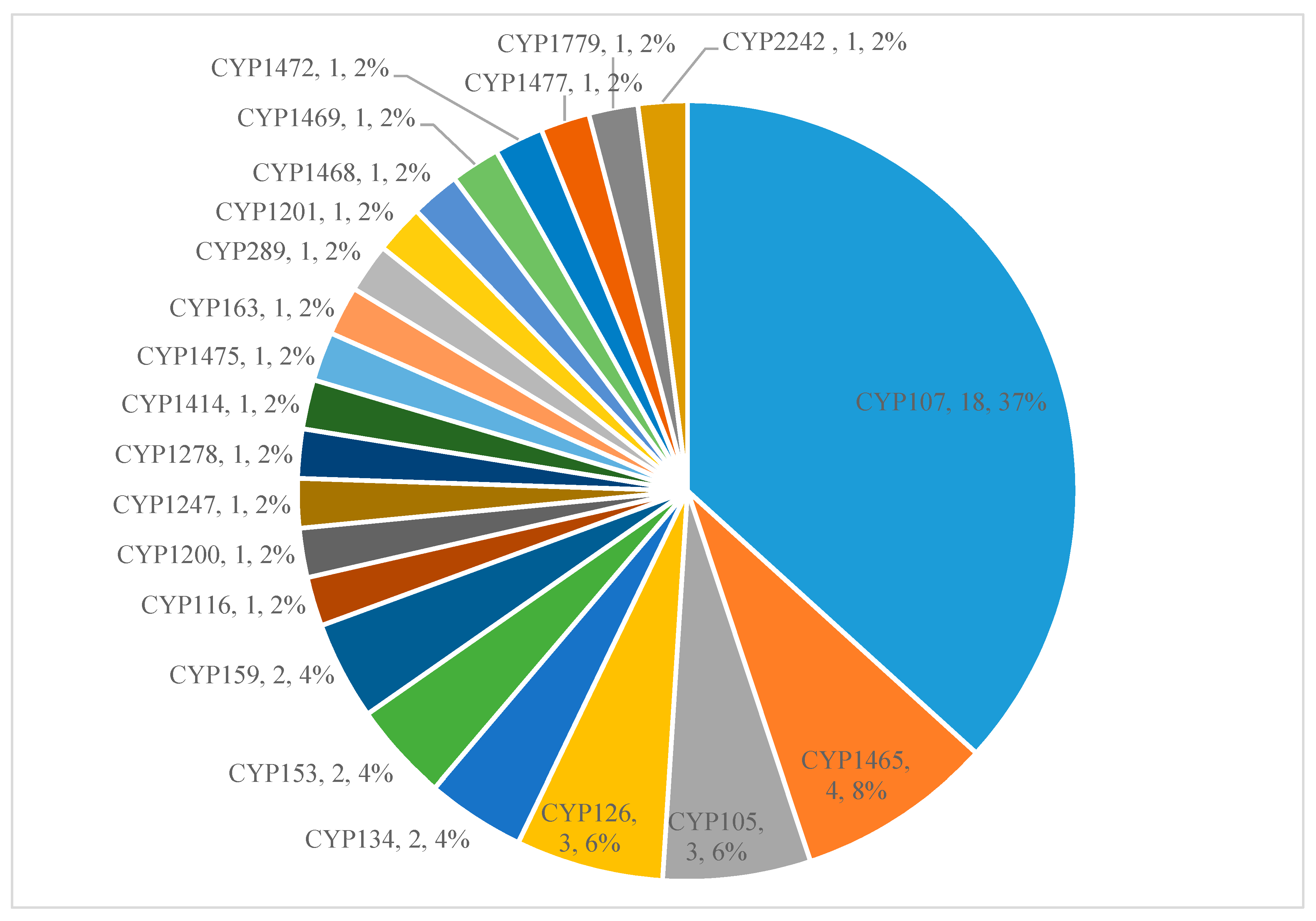
2.2. A Few P450 Families Are Expanded in Gammaproteobacterial Species
2.3. A Few P450s Are Involved in Secondary Metabolism in Gammaproteobacterial Species
2.4. Functional Prediction of Gammaproteobacterial P450s
2.5. Impact of Lifestyle on P450 Repertoire Is Also Evident in Gammaproteobacteria
3. Materials and Methods
3.1. Species and Their Genome Database Information
3.2. Genome Data Mining and Annotation of P450s
3.3. Phylogenetic Analysis of P450s
3.4. Generation of P450 Profile Heat Maps
3.5. Identification of P450 Part of Secondary Metabolite BGCs
3.6. Data Analysis
3.7. Comparative Analysis of P450s and Gene Cluster Data
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Berman, J.J. Taxonomic Guide to Infectious Diseases: Understanding the Biologic Classes of Pathogenic Organisms; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Kersters, K.; De Vos, P.; Gillis, M.; Swings, J.; Vandamme, P.; Stackebrandt, E. Introduction to the Proteobacteria. In The Prokaryotes: A Handbook on the Biology of Bacteria; Springer: New York, NY, USA, 2006; Volume 5, pp. 3–37. [Google Scholar]
- Williams, K.P.; Kelly, D.P. Proposal for a new class within the phylum Proteobacteria, Acidithiobacillia classis nov., with the type order Acidithiobacillales, and emended description of the class Gammaproteobacteria. Int. J. Syst. Evol. Microbiol. 2013, 63, 2901–2906. [Google Scholar] [CrossRef]
- Liu, L.; Chen, D.; Liu, L.; Lan, R.; Hao, S.; Jin, W.; Sun, H.; Wang, Y.; Liang, Y.; Xu, J. Genetic diversity, multidrug resistance, and virulence of Citrobacter freundii from diarrheal patients and healthy individuals. Front. Cell. Infect. Microbiol. 2018, 8, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, Z.E.; Shaker, M.; Baxter, J.D. Urinary tract infection caused by Citrobacter koseri in a patient with spina bifida, an ileal conduit and renal caluli progressing to peri-nephric abscess and empyema. Urol. Case Rep. 2017, 11, 22–24. [Google Scholar] [CrossRef]
- Annavajhala, M.K.; Gomez-Simmonds, A.; Uhlemann, A.-C. Multidrug-resistant Enterobacter cloacae complex emerging as a global, diversifying threat. Front. Microbiol. 2019, 10, 44. [Google Scholar] [CrossRef] [Green Version]
- Davin-Regli, A. Enterobacter aerogenes and Enterobacter cloacae, versatile bacterial pathogens confronting antibiotic treatment. Front. Microbiol. 2015, 6, 392. [Google Scholar] [CrossRef] [Green Version]
- Abdesselam, K.; Pagotto, F. Bacteria: Cronobacter (Enterobacter) sakazakii and Other Cronobacter spp. Encycl. Food Saf. 2014, 1, 424–432. [Google Scholar] [CrossRef]
- Rojas-Lopez, M.; Monterio, R.; Pizza, M.; Desvaux, M.; Rosini, R. Intestinal pathogenic Escherichia coli: Insights for vaccine development. Front. Microbiol. 2018, 9, 440. [Google Scholar] [CrossRef]
- Oaks, J.L.; Besser, T.E.; Walk, S.T.; Gordon, D.M.; Beckmen, K.B.; Burek, K.A.; Haldorson, G.J.; Bradway, D.S.; Ouellette, L.; Rurangirwa, F.R. Escherichia albertii in wild and domestic birds. Emerg. Infect. Dis. 2010, 16, 638. [Google Scholar] [CrossRef] [PubMed]
- Ooka, T.; Tokuoka, E.; Furukawa, M.; Nagamura, T.; Ogura, Y.; Arisawa, K.; Harada, S.; Hayashi, T. Human gastroenteritis outbreak associated with Escherichia albertii, Japan. Emerg. Infect. Dis. 2013, 19, 144–146. [Google Scholar] [CrossRef]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumoniae: Going on the offense with a strong defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef] [Green Version]
- Effah, C.Y.; Sun, T.; Liu, S.; Wu, Y. Klebsiella pneumoniae: An increasing threat to public health. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nizet, V.; Klein, J. Bacterial sepsis and meningitis. Infect. Dis. Fetus Newborn Infant 2011, 8, 217–271. [Google Scholar]
- Schaffer, J.N.; Pearson, M.M. Proteus mirabilis and urinary tract infections. In Urinary Tract Infections: Molecular Pathogenesis and Clinical Management; John Wiley & Sons: New York, NY, USA, 2017; pp. 383–433. [Google Scholar]
- Matthews, S.J.; Lancaster, J.W. Urinary tract infections in the elderly population. Am. J. Geriatr. Pharmacother. 2011, 9, 286–309. [Google Scholar] [CrossRef]
- Papazafiropoulou, A.; Daniil, I.; Sotiropoulos, A.; Balampani, E.; Kokolaki, A.; Bousboulas, S.; Konstantopoulou, S.; Skliros, E.; Petropoulou, D.; Pappas, S. Prevalence of asymptomatic bacteriuria in type 2 diabetic subjects with and without microalbu-minuria. BMC Res. Notes 2010, 3, 169. [Google Scholar] [CrossRef] [Green Version]
- Drzewiecka, D. Significance and roles of Proteus spp. bacteria in natural environments. Microb. Ecol. 2016, 72, 741–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andino, A.; Hanning, I. Salmonella enterica: Survival, colonization, and virulence differences among serovars. Sci. World J. 2015, 2015, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanna, A.; KhAnnA, M.; AggArWAl, A. Serratia marcescens—A rare opportunistic nosocomial pathogen and measures to limit its spread in hospitalized patients. J. Clin. Diagn. Res. JCDR 2013, 7, 243. [Google Scholar]
- Drancourt, M. Acute diarrhea. Infect. Dis. 2017, 1, 335–340.e2. [Google Scholar]
- Baker, S.; The, H.C. Recent insights into Shigella: A major contributor to the global diarrhoeal disease burden. Curr. Opin. Infect. Dis. 2018, 31, 449. [Google Scholar] [CrossRef]
- Ari, T.B.; Neerinckx, S.; Gage, K.L.; Kreppel, K.; Laudisoit, A.; Leirs, H.; Stenseth, N.C. Plague and climate: Scales matter. PLoS Pathog. 2011, 7, e1002160. [Google Scholar]
- Sabina, Y.; Rahman, A.; Ray, R.C.; Montet, D. Yersinia enterocolitica: Mode of transmission, molecular insights of virulence, and pathogenesis of infection. J. Pathog. 2011, 2011, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, H.; Shimizu, S.; Inatsu, Y. Yersinia enterocolitica and Yersinia pseudotuberculosis detection in foods. J. Pathog. 2011, 2011, 735308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein an-notation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, I.-M.A.; Chu, K.; Palaniappan, K.; Pillay, M.; Ratner, A.; Huang, J.; Huntemann, M.; Varghese, N.; White, J.R.; Seshadri, R.; et al. IMG/M v.5.0: An integrated data management and comparative analysis system for microbial genomes and microbiomes. Nucleic Acids Res. 2019, 47, D666–D677. [Google Scholar] [CrossRef]
- Akapo, O.O.; Padayachee, T.; Chen, W.; Kappo, A.P.; Yu, J.H.; Nelson, D.R.; Syed, K. Distribution and diversity of cytochrome P450 monooxygenases in the cungal class Tremellomycetes. Int. J. Mol. Sci. 2019, 20, 2889. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.R. Cytochrome P450 nomenclature. Methods Mol. Biol. 1998, 107, 15–24. [Google Scholar] [PubMed]
- Nelson, D.R. Cytochrome P450 Nomenclature, 2004. Methods Mol. Biol. 2006, 320, 1–10. [Google Scholar] [PubMed]
- Spring, S.; Scheuner, C.; Goker, M.; Klenk, H.P. A taxonomic framework for emerging groups of ecologically important marine Gammaproteobacteria based on the reconstruction of evolutionary relationships using genome-scale data. Front. Microbiol. 2015, 6, 281. [Google Scholar] [CrossRef] [Green Version]
- Korzhenkov, A.A.; Toshchakov, S.V.; Golyshina, O.V.; Ferrer, M.; Chernikova, T.N.; Jaeger, K.-E.; Yakimov, M.M.; Golyshin, P.N. Aerobic hydrocarbon-degrading Gammaproteobacteria: Oleiphilaceae and relatives. Taxon. Genom. Ecophysiol. Hydrocarb. Degrad. Microbes 2019, 153–166. [Google Scholar] [CrossRef]
- Nelson, D.R. Cytochrome P450 diversity in the tree of life. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2018, 1866, 141–154. [Google Scholar] [CrossRef]
- Lamb, D.C.; Follmer, A.H.; Goldstone, J.V.; Nelson, D.R.; Warrilow, A.G.; Price, C.L.; True, M.Y.; Kelly, S.L.; Poulos, T.L.; Stegeman, J.J. On the occurrence of cytochrome P450 in viruses. Proc. Natl. Acad. Sci. USA 2019, 116, 12343–12352. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, R. Cytochromes P450 as versatile biocatalysts. J. Biotechnol. 2006, 124, 128–145. [Google Scholar] [CrossRef]
- Sono, M.; Roach, M.P.; Coulter, E.D.; Dawson, J.H. Heme-containing oxygenases. Chem. Rev. 1996, 96, 2841–2888. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H. Fifty Years of Cytochrome P450 Research; Springer: Tokyo, Japan, 2014; pp. IX, 409. [Google Scholar]
- Guengerich, F.P. A history of the roles of cytochrome P450 enzymes in the toxicity of drugs. Toxicol. Res. 2021, 37, 1–23. [Google Scholar] [CrossRef]
- Lepesheva, G.I.; Friggeri, L.; Waterman, M.R. CYP51 as drug targets for fungi and protozoan parasites: Past, present and future. Parasitology 2018, 145, 1820–1836. [Google Scholar] [CrossRef]
- Jawallapersand, P.; Mashele, S.S.; Kovačič, L.; Stojan, J.; Komel, R.; Pakala, S.B.; Kraševec, N.; Syed, K. Cytochrome P450 monooxygenase CYP53 family in fungi: Comparative structural and evolutionary analysis and its role as a common alternative anti-fungal drug target. PLoS ONE 2014, 9, e107209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz de Montellano, P.R. Potential drug targets in the Mycobacterium tuberculosis cytochrome P450 system. J. Inorg. Biochem. 2018, 180, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Podust, L.M.; Sherman, D.H. Diversity of P450 enzymes in the biosynthesis of natural products. Nat. Prod. Rep. 2012, 29, 1251–1266. [Google Scholar] [CrossRef] [Green Version]
- Greule, A.; Stok, J.E.; De Voss, J.J.; Cryle, M.J. Unrivalled diversity: The many roles and reactions of bacterial cytochromes P450 in secondary metabolism. Nat. Prod. Rep. 2018, 35, 757–791. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.F.; Tatsuta, K.; Gunji, H.; Ishiyama, T.; Hutchinson, C.R. Substrate specificity of 6-deoxyerythronolide B hydroxylase, a bacterial cytochrome P450 of erythromycin A biosynthesis. Biochemistry 1993, 32, 1905–1913. [Google Scholar] [CrossRef]
- Bischoff, D.; Bister, B.; Bertazzo, M.; Pfeifer, V.; Stegmann, E.; Nicholson, G.J.; Keller, S.; Pelzer, S.; Wohlleben, W.; Süssmuth, R.D. The biosynthesis of vancomycin-type glycopeptide antibiotics—A model for oxidative side-chain cross-linking by oxygenases coupled to the action of peptide synthetases. ChemBioChem 2005, 6, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Jennewein, S.; Park, H.; DeJong, J.M.; Long, R.M.; Bollon, A.P.; Croteau, R.B. Coexpression in yeast of Taxus cytochrome P450 reductase with cytochrome P450 oxygenases involved in Taxol biosynthesis. Biotechnol. Bioeng. 2005, 89, 588–598. [Google Scholar] [CrossRef] [PubMed]
- van Beilen, J.B.; Holtackers, R.; Lüscher, D.; Bauer, U.; Witholt, B.; Duetz, W.A. Biocatalytic production of perillyl alcohol from limonene by using a novel Mycobacterium sp. cytochrome P450 alkane hydroxylase expressed in Pseudomonas putida. Appl. Environ. Microbiol. 2005, 71, 1737–1744. [Google Scholar] [CrossRef] [Green Version]
- Timmermans, M.L.; Paudel, Y.P.; Ross, A.C. Investigating the biosynthesis of natural products from marine proteobacteria: A survey of molecules and strategies. Mar. Drugs 2017, 15, 235. [Google Scholar] [CrossRef] [Green Version]
- Senate, L.M.; Tjatji, M.P.; Pillay, K.; Chen, W.; Zondo, N.M.; Syed, P.R.; Mnguni, F.C.; Chiliza, Z.E.; Bamal, H.D.; Karpoormath, R.; et al. Similarities, variations, and evolution of cytochrome P450s in Streptomyces versus Mycobacterium. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mnguni, F.C.; Padayachee, T.; Chen, W.; Gront, D.; Yu, J.-H.; Nelson, D.R.; Syed, K. More P450s are involved in secondary metabolite biosynthesis in Streptomyces compared to Bacillus, Cyanobacteria and Mycobacterium. Int. J. Mol. Sci. 2020, 21, 4814. [Google Scholar] [CrossRef]
- Parvez, M.; Qhanya, L.B.; Mthakathi, N.T.; Kgosiemang, I.K.R.; Bamal, H.D.; Pagadala, N.S.; Xie, T.; Yang, H.; Chen, H.; Theron, C.W.; et al. Molecular evolutionary dynamics of cytochrome P450 monooxygenases across kingdoms: Special focus on mycobacterial P450s. Sci. Rep. 2016, 6, 33099. [Google Scholar] [CrossRef] [Green Version]
- Syed, P.R.; Chen, W.; Nelson, D.R.; Kappo, A.P.; Yu, J.-H.; Karpoormath, R.; Syed, K. Cytochrome P450 monooxygenase CYP139 family involved in the synthesis of secondary metabolites in 824 Mycobacterial Species. Int. J. Mol. Sci. 2019, 20, 2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngcobo, N.S.; Chiliza, Z.E.; Chen, W.; Yu, J.-H.; Nelson, D.R.; Tuszynski, J.A.; Preto, J.; Syed, K. Comparative analysis, structural insights, and substrate/drug interaction of CYP128A1 in Mycobacterium tuberculosis. Int. J. Mol. Sci. 2020, 21, 4816. [Google Scholar] [CrossRef]
- Khumalo, M.J.; Nzuza, N.; Padayachee, T.; Chen, W.; Yu, J.H.; Nelson, D.R.; Syed, K. Comprehensive analyses of cytochrome P450 monooxygenases and secondary metabolite biosynthetic gene clusters in Cyanobacteria. Int. J. Mol. Sci. 2020, 21, 656. [Google Scholar] [CrossRef] [Green Version]
- Padayachee, T.; Nzuza, N.; Chen, W.; Nelson, D.R.; Syed, K. Impact of lifestyle on cytochrome P450 monooxygenase repertoire is clearly evident in the bacterial phylum Firmicutes. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Mthethwa, B.C.; Chen, W.; Ngwenya, M.L.; Kappo, A.P.; Syed, P.R.; Karpoormath, R.; Yu, J.-H.; Nelson, D.R.; Syed, K. Comparative analyses of cytochrome P450s and those associated with secondary metabolism in Bacillus Species. Int. J. Mol. Sci. 2018, 19, 3623. [Google Scholar] [CrossRef] [Green Version]
- Sello, M.M.; Jafta, N.; Nelson, D.R.; Chen, W.; Yu, J.-H.; Parvez, M.; Kgosiemang, I.K.R.; Monyaki, R.; Raselemane, S.C.; Qhanya, L.B.; et al. Diversity and evolution of cytochrome P450 monooxygenases in Oomycetes. Sci. Rep. 2015, 5, 11572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munro, A.W.; Leys, D.G.; McLean, K.J.; Marshall, K.R.; Ost, T.W.; Daff, S.; Miles, C.S.; Chapman, S.K.; Lysek, D.A.; Moser, C.C.; et al. P450 BM3: The very model of a modern flavocytochrome. Trends Biochem. Sci. 2002, 27, 250–257. [Google Scholar] [CrossRef]
- Li, H.; Poulos, T.L. The structure of the cytochrome p450BM-3 haem domain complexed with the fatty acid substrate, palmitoleic acid. Nat. Struct. Biol. 1997, 4, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Noble, M.; Miles, C.; Reid, G.; Chapman, S.; Munro, A. Catalytic properties of key active site mutants of flavocytochrome P-450 BM. Biochem. Soc. Trans. 1999, 27, A44. [Google Scholar] [CrossRef]
- Poulos, T.L.; Finzel, B.C.; Howard, A.J. Crystal structure of substrate-free Pseudomonas putida cytochrome P-450. Biochemistry 1986, 25, 5314–5322. [Google Scholar] [CrossRef] [PubMed]
- McLean, K.J.; Leys, D.; Munro, A.W. Microbial cytochrome P450s. In Cytochrome P450: Structure, Mechanism, and Biochemistry, 4th ed.; Oritz de Montellano, P.R., Ed.; Springer International Publishing: New York, NY, USA, 2015; Chapter 6; pp. 261–407. [Google Scholar]
- Moody, S.C.; Loveridge, E.J. CYP105-diverse structures, functions and roles in an intriguing family of enzymes in Streptomyces. J. Appl. Microbiol. 2014, 117, 1549–1563. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.-Z.; Li, X.-F.; Yang, W.; Dong, X.; Yu, J.; Zhu, S.-L.; Li, M.; Xie, L.; Tong, W.-Y. Identification and functional analysis of cytochrome P450 complement in Streptomyces virginiae IBL14. BMC Genom. 2013, 14, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.-S.; Yamada, A.; Matsunaga, I.; Ichihara, K.; Adachi, S.-I.; Park, S.-Y.; Shiro, Y. Crystallization and preliminary X-ray diffraction analysis of fatty-acid hydroxylase cytochrome P450BSβ from Bacillus subtilis. Acta Crystallogr. Sect. D Biol. Crystallogr. 2002, 58, 687–689. [Google Scholar] [CrossRef]
- Lee, D.-S.; Yamada, A.; Sugimoto, H.; Matsunaga, I.; Ogura, H.; Ichihara, K.; Adachi, S.-I.; Park, S.-Y.; Shiro, Y. Substrate recognition and molecular mechanism of fatty acid hydroxylation by cytochrome P450 from Bacillus subtilis crystallographic, spectroscopic, and mutational studies. J. Biol. Chem. 2003, 278, 9761–9767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girvan, H.M.; Poddar, H.; McLean, K.J.; Nelson, D.R.; Hollywood, K.A.; Levy, C.W.; Leys, D.; Munro, A.W. Structural and catalytic properties of the peroxygenase P450 enzyme CYP152K6 from Bacillus methanolicus. J. Inorg. Biochem. 2018, 188, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Funhoff, E.G.; Bauer, U.; García-Rubio, I.; Witholt, B.; van Beilen, J.B. CYP153A6, a soluble P450 oxygenase catalyzing terminal-alkane hydroxylation. J. Bacteriol. 2006, 188, 5220–5227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.J.; Patrauchan, M.A.; Florizone, C.; Eltis, L.D.; Mohn, W.W. Distinct roles for two CYP226 family cytochromes P450 in abietane diterpenoid catabolism by Burkholderia xenovorans LB400. J. Bacteriol. 2007, 190, 1575–1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, C.; Wyndham, R. Characterization of tdt genes for the degradation of tricyclic diterpenes by Pseudomonas diterpeniphila A19-6a. Can. J. Microbiol. 2002, 48, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Davydov, D.R.; Sineva, E.V.; Davydova, N.Y.; Bartlett, U.H.; Halpert, J.R. CYP261 enzymes from deep sea bacteria: A clue to conformational heterogeneity in cytochromes P450. Biotechnol. Appl. Biochem. 2013, 60, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R. Insect CYP genes and P450 enzymes. In Insect Molecular Biology and Biochemistry; Academic Press: Cambridge, MA, USA, 2012; pp. 236–316. [Google Scholar]
- Hamberger, B.; Bak, S. Plant P450s as versatile drivers for evolution of species-specific chemical diversity. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120426. [Google Scholar] [CrossRef] [Green Version]
- Ngwenya, M.L.; Chen, W.; Basson, A.K.; Shandu, J.S.; Yu, J.-H.; Nelson, D.R.; Syed, K. Blooming of unusual cytochrome P450s by tandem duplication in the pathogenic fungus Conidiobolus coronatus. Int. J. Mol. Sci. 2018, 19, 1711. [Google Scholar] [CrossRef] [Green Version]
- Matowane, R.G.; Wieteska, L.; Bamal, H.D.; Kgosiemang, I.K.R.; Van Wyk, M.; Manume, N.A.; Abdalla, S.M.H.; Mashele, S.S.; Gront, D.; Syed, K. In silico analysis of cytochrome P450 monooxygenases in chronic granulomatous infectious fungus Sporothrix schenckii: Special focus on CYP51. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2018, 1866, 166–177. [Google Scholar] [CrossRef]
- Qhanya, L.B.; Matowane, G.; Chen, W.; Sun, Y.; Letsimo, E.M.; Parvez, M.; Yu, J.-H.; Mashele, S.S.; Syed, K. Genome-wide annotation and comparative analysis of cytochrome P450 monooxygenases in basidiomycete biotrophic plant pathogens. PLoS ONE 2015, 10, e0142100. [Google Scholar] [CrossRef] [Green Version]
- Kgosiemang, I.K.R.; Syed, K.; Mashele, S.S. Comparative genomics and evolutionary analysis of cytochrome P450 monooxygenases in fungal subphylum Saccharomycotina. J. Pure Appl. Microbiol. 2014, 8, 12. [Google Scholar]
- Syed, K.; Shale, K.; Pagadala, N.S.; Tuszynski, J. Systematic identification and evolutionary analysis of catalytically versatile cytochrome P450 monooxygenase families enriched in model basidiomycete fungi. PLoS ONE 2014, 9, e86683. [Google Scholar] [CrossRef]
- Suzuki, H.; Macdonald, J.; Syed, K.; Salamov, A.; Hori, C.; Aerts, A.; Henrissat, B.; Wiebenga, A.; A Vankuyk, P.; Barry, K.; et al. Comparative genomics of the white-rot fungi, Phanerochaete carnosa and P. chrysosporium, to elucidate the genetic basis of the distinct wood types they colonize. BMC Genom. 2012, 13, 444. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Sato, Y.; Furumichi, M.; Morishima, K.; Tanabe, M. New approach for understanding genome variations in KEGG. Nucleic Acids Res. 2019, 47, D590–D595. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; I Hurwitz, D.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Graham, S.E.; Peterson, J.A. How similar are P450s and what can their differences Teach Us? Arch. Biochem. Biophys. 1999, 369, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Syed, K.; Mashele, S.S. Comparative analysis of P450 signature motifs EXXR and CXG in the large and diverse kingdom of fungi: Identification of evolutionarily conserved amino acid patterns characteristic of P450 Family. PLoS ONE 2014, 9, e95616. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Kuma, K.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef]
- Boc, A.; Diallo, A.B.; Makarenkov, V. T-REX: A web server for inferring, validating and visualizing phylogenetic trees and networks. Nucleic Acids Res. 2012, 40, W573–W579. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, A.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. TM4: A Free, Open-Source System for Microarray Data Management and Analysis. Biotechniques 2003, 34, 374–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blin, K.; Andreu, V.P.; Santos, E.L.C.D.L.; Del Carratore, F.; Lee, S.Y.; Medema, M.H.; Weber, T. The antiSMASH database version 2: A comprehensive resource on secondary metabolite biosynthetic gene clusters. Nucleic Acids Res. 2019, 47, D625–D630. [Google Scholar] [CrossRef] [PubMed] [Green Version]





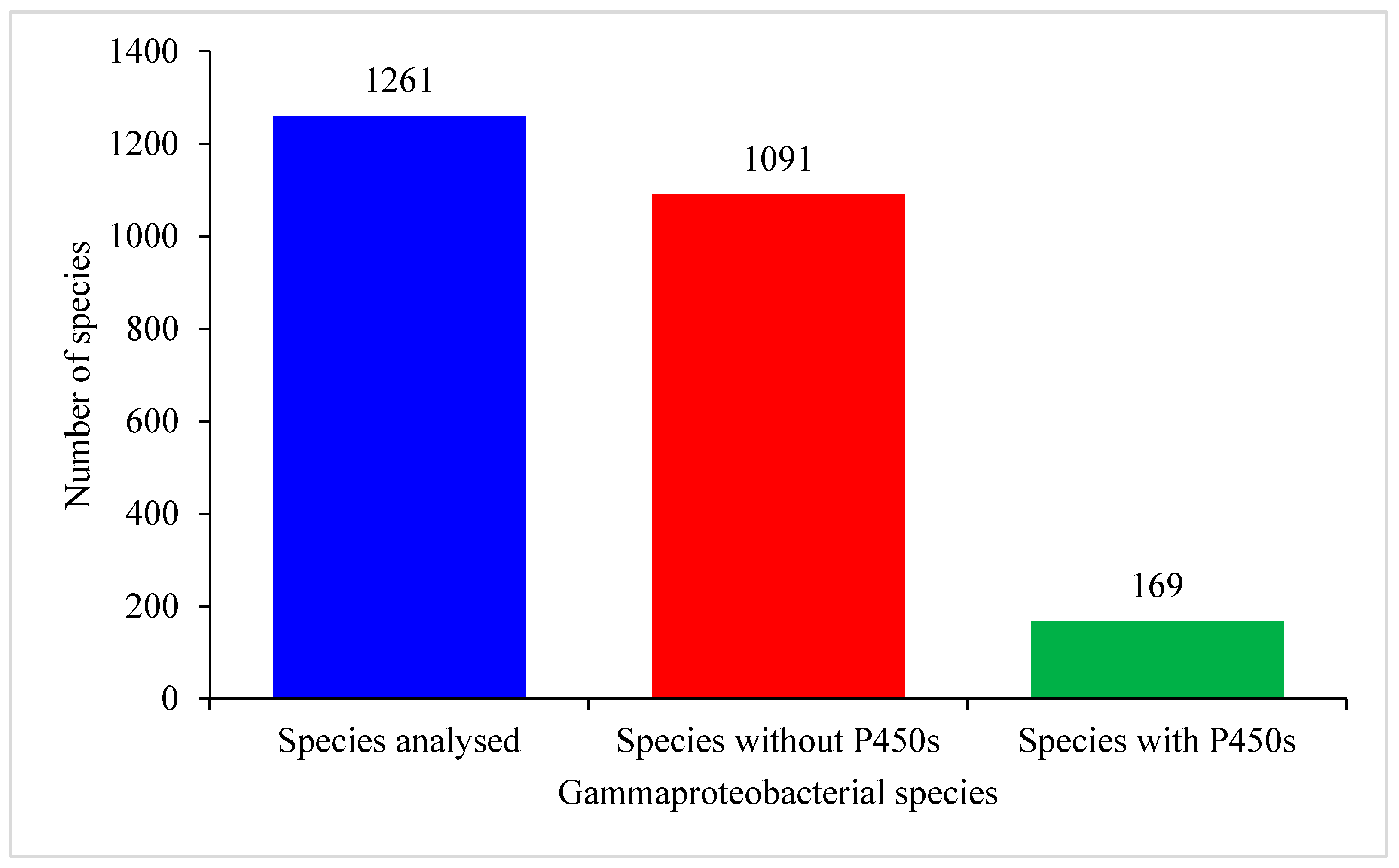{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | General Information | Reference(s) |
|---|---|---|
| Citrobacter freundii | It is a commensal resident of the intestinal tracts of both humans and animals. It causes diarrhea and other infections in humans. | [4] |
| Citrobacter koseri | It is present as part of human flora. It causes meningitis in neonates, infants and immune-compromised people and in adults it causes urinary tract infections. | [5] |
| Enterobacter cloacae | It is a common nosocomial pathogen capable of producing a wide variety of infections, such as pneumonia, urinary tract infections, and septicemia. | [6] |
| Enterobacter aerogenes | It is a commensal of humans and an opportunistic pathogen. It causes respiratory, urinary, blood, or gastrointestinal tract infections. | [7] |
| Enterobacter sakasakii | It is a food-borne pathogen causing a severe form of sepsis, necrotizing enterocolitis and meningitis in neonatal infants. | [8] |
| Escherichia coli | It is commonly found in human flora. Pathogenic E. coli can be divided into two groups: extra-intestinal pathogens associated mainly with neonatal meningitis and urinary tract infections in adults and intestinal pathogens causes diarrhea. | [9] |
| Escherichia albertii | It is an emerging human enteropathogen and avian pathogen that causes diarrhea. | [10,11] |
| Klebsiella pneumoniae | It used to be an opportunistic pathogen causing a wide range of infections in immunocompromised patients, but has recently emerged as a pathogen of healthy and immunocompetent people. It causes pneumonias, urinary tract infections, bacteremia, and liver abscesses. | [12,13] |
| Klebsiella oxytoca | It is an emerging pathogen causing a wide range of infections similar to K. pneumoniae. K. oxytoca causes neonatal infections of the bloodstream, urinary tract, central nervous system, lung, skin, and soft tissues. | [14] |
| Proteus mirabilis | It is a human opportunistic pathogen. It causes infections of the urinary tract, including cystitis and pyelonephritis. It is also found in cases of asymptomatic bacteriuria, particularly in the elderly and patients with type 2 diabetes. | [15,16,17] |
| Proteus vulgaris | It is also an opportunistic pathogen causing infections such as P. mirabilis. | [18] |
| Salmonella enterica | This food-borne pathogen causes typhoid and paratyphoid fever in humans. | [19] |
| Serratia marcescens | It is an opportunistic pathogen causing bacteremia/sepsis. | [20] |
| Shigella dysenteriae, S. flexneri, S. sonnei and S. boydii | Shigella spp. are responsible for acute diarrhea and are major contributors to the global diarrheal disease burden. | [21,22] |
| Yersinia pestis | It causes bubonic plague and pneumonia. It is well-known to cause one of the most devastating diseases of human history, the black death. | [23] |
| Yersinia enterocolitica | It causes acute diarrhea, mesenteric adenitis, terminal ileitis, and pseudo-appendicitis. | [24] |
| Yersinia pseudotuberculosis | It causes mesenteric lymphadenitis, diarrhea, and septicemia in humans. | [25] |
| Gammaproteobacterial Species | Firmicutes Species | Streptomyces Species | Mycobacterial Species | Cyanobacterial Species | |
|---|---|---|---|---|---|
| Total no. of species analyzed | 1261 | 972 | 203 | 60 | 114 |
| No of Species have P450s | 169 | 229 | 203 | 60 | 114 |
| No. of P450s | 277 | 712 | 5460 | 1784 | 341 |
| No. of families | 84 | 14 | 253 | 77 | 36 |
| No. of subfamilies | 105 | 53 | 698 | 132 | 79 |
| Dominant P450 family | CYP133 | CYP107 | CYP107 | CYP125 | CYP110 |
| Average no. of P450s | 0.2 | 1 | 27 | 30 | 3 |
| P450 diversity percentage | 0.18 | 0.008 | 0.02 | 0.07 | 0.09 |
| No. of P450s part of BGCs | 49 | 126 | 1231 | 204 | 27 |
| Percentage of P450s part of BGCs | 18 | 18 | 23 | 11 | 8 |
| Reference(s) | This work | [55,56] | [49,50] | [49,51] | [54] |
| Family | Subfamily | P450 Count | Percentage Contribution |
|---|---|---|---|
| CYP101 | M | 1 | 0.36 |
| CYP102 | J | 1 | 0.36 |
| CYP1043 | C | 1 | 0.36 |
| CYP1049 | A | 1 | 0.36 |
| CYP105 | BQ | 4 | 1.44 |
| CYP107 | DJ | 3 | 7.94 |
| E | 4 | ||
| HT | 1 | ||
| S | 14 | ||
| CYP108 | R | 1 | 0.36 |
| CYP1097 | B | 1 | 0.36 |
| CYP1104 | B | 3 | 1.08 |
| CYP111 | A | 1 | 0.36 |
| CYP112 | A | 3 | 1.08 |
| CYP1138 | A | 1 | 0.36 |
| CYP1142 | A | 1 | 0.36 |
| CYP114 | A | 4 | 1.44 |
| CYP1157 | C | 1 | 0.36 |
| CYP115 | A | 3 | 1.08 |
| CYP1164 | A | 1 | 0.36 |
| CYP1165 | A | 1 | 0.72 |
| B | 1 | ||
| CYP116 | B | 2 | 0.72 |
| CYP1172 | A | 1 | 0.36 |
| CYP1179 | A | 1 | 0.36 |
| CYP117 | A | 3 | 1.08 |
| CYP1199 | A | 2 | 0.72 |
| CYP1200 | B | 1 | 0.36 |
| CYP1201 | A | 1 | 0.72 |
| B | 1 | ||
| CYP1202 | A | 1 | 0.36 |
| CYP1225 | A | 1 | 0.36 |
| CYP1234 | A | 5 | 2.17 |
| B | 1 | ||
| CYP1247 | A | 9 | 3.25 |
| CYP1261 | B | 1 | 0.36 |
| CYP126 | C | 4 | 1.44 |
| CYP1278 | A | 1 | 0.36 |
| CYP1311 | A | 1 | 0.36 |
| CYP133 | B | 22 | 8.30 |
| D | 1 | ||
| CYP134 | B | 2 | 0.72 |
| CYP136 | F | 2 | 0.72 |
| CYP1413 | A | 3 | 1.44 |
| B | 1 | ||
| CYP1414 | A | 1 | 0.72 |
| B | 1 | ||
| CYP1415 | A | 3 | 1.08 |
| CYP1464 | A | 2 | 0.72 |
| CYP1465 | A | 4 | 1.44 |
| CYP1466 | A | 1 | 0.36 |
| CYP1467 | A | 1 | 0.36 |
| CYP1468 | A | 1 | 0.36 |
| CYP1469 | A | 2 | 0.72 |
| CYP1470 | A | 3 | 1.08 |
| CYP1471 | A | 1 | 0.36 |
| CYP1472 | A | 1 | 0.36 |
| CYP1473 | A | 1 | 0.36 |
| CYP1474 | A | 3 | 2.53 |
| B | 2 | ||
| C | 2 | ||
| CYP1475 | A | 1 | 0.36 |
| CYP1476 | A | 1 | 0.36 |
| CYP1477 | A | 1 | 0.36 |
| CYP1478 | A | 1 | 0.36 |
| CYP1479 | A | 1 | 0.36 |
| CYP1480 | A | 1 | 0.36 |
| CYP1481 | A | 2 | 0.72 |
| CYP1482 | A | 1 | 0.36 |
| CYP1483 | A | 1 | 0.36 |
| CYP151 | A | 2 | 0.72 |
| CYP152 | G | 3 | 3.25 |
| R | 2 | ||
| W | 2 | ||
| X | 1 | ||
| Y | 1 | ||
| CYP153 | A | 16 | 6.86 |
| E | 3 | ||
| CYP159 | B | 3 | 1.08 |
| CYP168 | A | 18 | 7.22 |
| B | 1 | ||
| C | 1 | ||
| CYP169 | A | 13 | 4.69 |
| CYP172 | B | 1 | 0.36 |
| CYP177 | E | 2 | 0.72 |
| CYP198 | A | 3 | 1.08 |
| CYP221 | A | 2 | 0.72 |
| CYP226 | A | 5 | 1.81 |
| CYP229 | A | 7 | 6.50 |
| D | 7 | ||
| E | 4 | ||
| CYP234 | A | 1 | 0.36 |
| CYP236 | A | 3 | 1.08 |
| CYP238 | A | 1 | 0.36 |
| CYP261 | D | 1 | 0.36 |
| CYP289 | A | 6 | 2.53 |
| D | 1 | ||
| CYP51 | B | 2 | 0.72 |
| CYP107 | S | 1 | 0.36 |
| CYP1229 | A | 1 | 0.36 |
| CYP1414 | A | 1 | 0.36 |
| CYP2242 | A | 1 | 0.36 |
| CYP159 | B | 1 | 0.36 |
| CYP163 | K | 1 | 0.36 |
| CYP1779 | A | 1 | 0.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Msomi, N.N.; Padayachee, T.; Nzuza, N.; Syed, P.R.; Kryś, J.D.; Chen, W.; Gront, D.; Nelson, D.R.; Syed, K. In Silico Analysis of P450s and Their Role in Secondary Metabolism in the Bacterial Class Gammaproteobacteria. Molecules 2021, 26, 1538. https://doi.org/10.3390/molecules26061538
Msomi NN, Padayachee T, Nzuza N, Syed PR, Kryś JD, Chen W, Gront D, Nelson DR, Syed K. In Silico Analysis of P450s and Their Role in Secondary Metabolism in the Bacterial Class Gammaproteobacteria. Molecules. 2021; 26(6):1538. https://doi.org/10.3390/molecules26061538
Chicago/Turabian StyleMsomi, Ntombizethu Nokuphiwa, Tiara Padayachee, Nomfundo Nzuza, Puleng Rosinah Syed, Justyna Dorota Kryś, Wanping Chen, Dominik Gront, David R. Nelson, and Khajamohiddin Syed. 2021. "In Silico Analysis of P450s and Their Role in Secondary Metabolism in the Bacterial Class Gammaproteobacteria" Molecules 26, no. 6: 1538. https://doi.org/10.3390/molecules26061538