Acyclic Triterpenoid Isolated from Alpinia katsumadai Alleviates Formalin-Induced Chronic Mouse Paw Inflammation by Inhibiting the Phosphorylation of ERK and NF-κB

 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. PHT Decreases NO and PGE2 Production in LPS-Induced J774 Cells
2.2. PHT Suppresses Inducible NO Synthase (iNOS) and Cyclooxygenase 2 (COX2) Expression in LPS-Induced J774 Cells
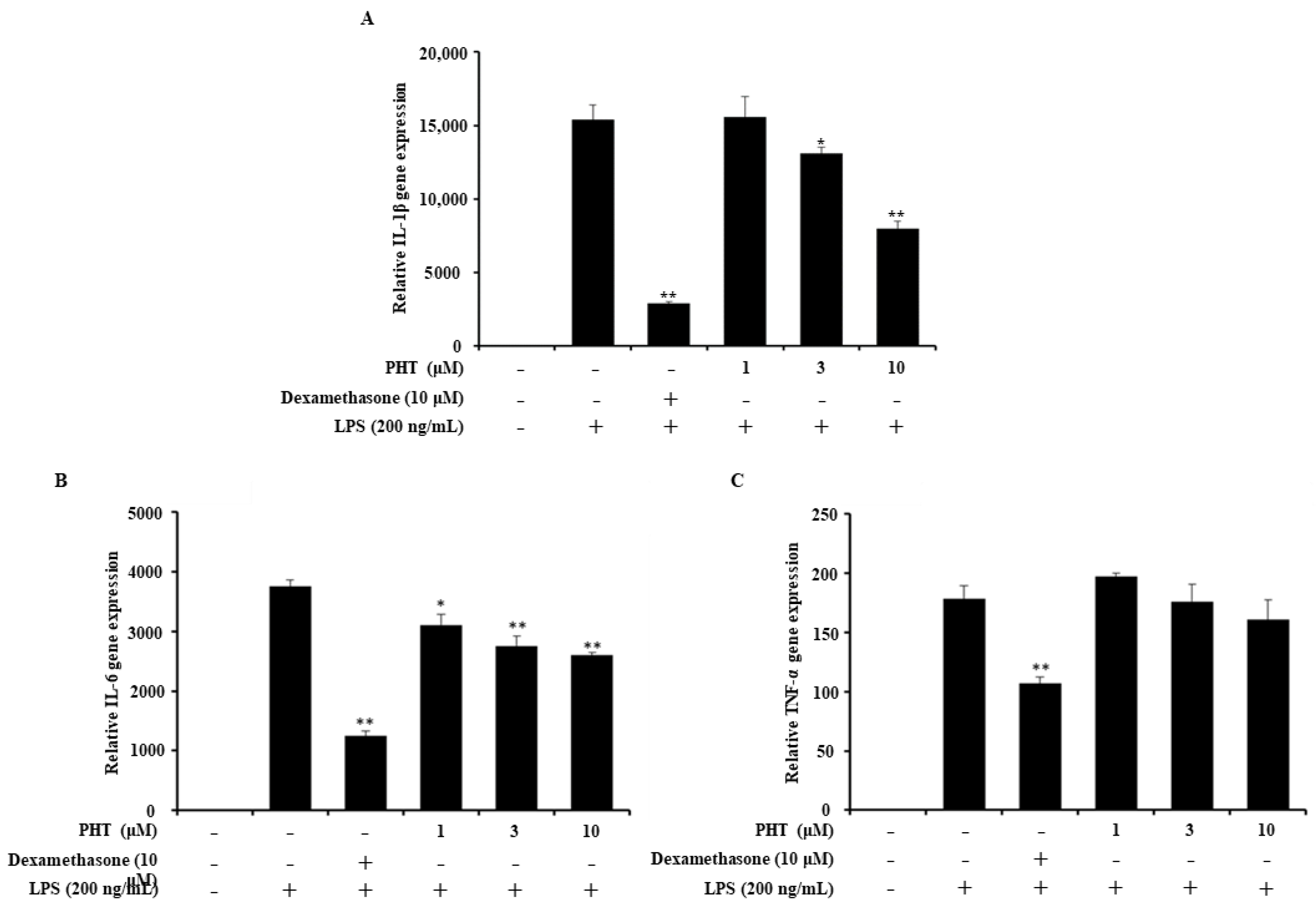
2.3. PHT Downregulates IL-1β and IL-6 But Not TNF-α Gene Expression in LPS-Induced J774 Cells
2.4. PHT Inhibits the Phosphorylation of ERK and NF-κB p65 But Not JNK or p38 in LPS-Induced J774 Cells
2.5. PHT Alleviates Formalin-Induced Chronic Mouse Paw Inflammation
3. Materials and Methods
3.1. Materials and Reagents
3.2. Isolation of PHT
3.2.1. General Experimental Procedure
3.2.2. Extraction and Isolation of PHT
3.2.3. 2,3,5,22,23-Pentahydroxy-2,6,10,15,19,23-Hexamethyl-Tetracosa-6,10,14,18-Tetraene (PHT)
3.3. NO Assay and MTT Assay
3.4. ELISA
3.5. Quantitative Real-Time PCR
3.6. Immunoblot Analysis
3.7. Animals and Induction of Formalin-Induced Chronic Mouse Paw Inflammation Model
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, S.; Niu, J.; Schmidt, C.; Sclabas, G.M.; Peng, B.; Uwagawa, T.; Li, Z.; Evans, D.B.; Abbruzzese, J.L.; Chiao, P.J. NF-κB and AP-1 connection: Mechanism of NF-κB-dependent regulation of AP-1 activity. Mol. Cell. Biol. 2004, 24, 7806–7819. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. Signaling to NF-κB by Toll-like receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef]
- Schaible, H.G.; von Banchet, G.S.; Boettger, M.K.; Bräuer, R.; Gajda, M.; Richter, F.; Hensellek, S.; Brenn, D.; Natura, G. The role of proinflammatory cytokines in the generation and maintenance of joint pain. Ann. N. Y. Acad. Sci. 2010, 1193, 60–69. [Google Scholar] [CrossRef]
- Feghali, C.A.; Wright, T.M. Cytokines in acute and chronic inflammation. Front. Biosci. 1997, 2, d12–d26. [Google Scholar]
- Nikolaus, S.; Bauditz, J.; Gionchetti, P.; Witt, C.; Lochs, H.; Schreiber, S. Increased secretion of pro-inflammatory cytokines by circulating polymorphonuclear neutrophils and regulation by interleukin 10 during intestinal inflammation. Gut 1998, 42, 470–476. [Google Scholar] [CrossRef] [Green Version]
- Martel-Pelletier, J.; Alaaeddine, N.; Pelletier, J.P. Cytokines and their role in the pathophysiology of osteoarthritis. Front. Biosci. 1999, 4, d694–d703. [Google Scholar] [CrossRef] [Green Version]
- Chung, K.F.; Barnes, P.J. Cytokines in asthma. Thorax 1999, 54, 825–857. [Google Scholar] [CrossRef] [Green Version]
- Al-Janadi, M.; Al-Balla, S.; Al-Dalaan, A.; Raziuddin, S. Cytokine profile in systemic lupus erythematosus, rheumatoid arthritis, and other rheumatic diseases. J. Clin. Immunol. 1993, 13, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Shin, H.T.; Hwang, H.J.; Kim, J.H. Antioxidant activity of extracts from Alpinia katsumadai seed. Phytotherapy Res. 2003, 17, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Koyama, K.; Takahashi, K.; Tai, T.; Nunoura, Y.; Watanabe, K. Two novel anti-emetic principles of Alpinia katsumadai. J. Nat. Prod. 1999, 62, 1672–1674. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Song, Z.M.; He, C.L.; Zhang, L.Y. A constituent of Alpinia katsumadai suppresses allergic airway inflammation. Phytochem. Lett. 2017, 22, 149–153. [Google Scholar] [CrossRef]
- Yang, J.; Dai, Y.; Xia, Y.F.; Huang, W.Z.; Wang, Z.T. Alpinia katsumadai hayata prevents mouse sepsis induced by cecal ligation and puncture through promoting bacterial clearance and downregulating systemic inflammation. Phytother. Res. 2009, 23, 267–273. [Google Scholar] [CrossRef]
- Jang, H.J.; Lee, S.J.; Lee, S.; Jung, K.; Lee, S.W.; Rho, M.C. Acyclic Triterpenoids from Alpinia katsumadai inhibit IL-6-induced STAT3 activation. Molecules 2017, 22, 1611. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.F. Bacterial triterpenoids. Microbiol. Rev. 1984, 48, 181. [Google Scholar] [CrossRef]
- Ishimatsu, A.; Matsuura, H.; Sano, T.; Kaya, K.; Watanabe, M.M. Biosynthesis of isoprene units in the C34 botryococcene molecule produced by Botryococcus braunii strain Bot-22. Procedia Environ. Sci. 2012, 15, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.Y.; Lee, M.H.; Choi, J.H.; Kim, Y.K. 2, 3, 22, 23-Tetrahydroxyl-2, 6, 10, 15, 19, 23-hexamethyl-6, 10, 14, 18-tetracosatetraene, an acyclic triterpenoid isolated from the seeds of Alpinia katsumadai, Inhibits Acyl-CoA: Cholesterol Acyltransferase Activity. Biol. Pharm. Bull. 2012, 35, 2092–2096. [Google Scholar] [CrossRef] [Green Version]
- Murata, T.; Miyase, T.; Muregi, F.W.; Naoshima-Ishibashi, Y.; Umehara, K.; Warashina, T.; Kanou, S.; Mkoji, G.M.; Terada, M.; Ishih, A. Antiplasmodial triterpenoids from Ekebergia capensis. J. Nat. Prod. 2008, 71, 167–174. [Google Scholar] [CrossRef]
- Teixeira, E.W.; Negri, G.; Salatino, A.; Stringheta, P.C. Seasonal variation, chemical composition and antioxidant activity of Brazilian propolis samples. Evid. -Based Complementary Altern. Med. 2010, 7. [Google Scholar] [CrossRef] [PubMed]
- Rho, M.C.; Kim, Y.K.; Lee, H.S.; Jun, C.D.; Kim, K.; Lee, S.W.; Choi, J.H.; Song, G.Y. New acyclic triterpenoids compound, and pharmaceutical composition comprising Alpinia katsumadai extract or acyclic triterpenoids compounds isolated from the same. International Patent Application No. WO/2008/133387, 6 November 2008. [Google Scholar]
- Farrell, A.J.; Blake, D.R.; Palmer, R.M.; Moncada, S. Increased concentrations of nitrite in synovial fluid and serum samples suggest increased nitric oxide synthesis in rheumatic diseases. Ann. Rheum. Dis. 1992, 51, 1219–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, H.; Hla, T.; Maier, J.A.; Crofford, L.J.; Case, J.P.; Maciag, T.; Wilder, R.L. In vivo cyclooxygenase expression in synovial tissues of patients with rheumatoid arthritis and osteoarthritis and rats with adjuvant and streptococcal cell wall arthritis. J. Clin. Investig. 1992, 89, 97–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, P.; Tripathi, P.; Kashyap, L.; Singh, V. The role of nitric oxide in inflammatory reactions. Fems Immunol. Med. Microbiol. 2007, 51, 443–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mardini, I.A.; FitzGerald, G.A. Selective inhibitors of cyclooxygenase-2. Mol. Interv. 2001, 1, 30. [Google Scholar]
- Menaka, K.B.; Ramesh, A.; Thomas, B.; Kumari, N.S. Estimation of nitric oxide as an inflammatory marker in periodontitis. J. Indian Soc. Periodontol. 2009, 13, 75. [Google Scholar] [CrossRef]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef]
- Aikawa, N. Cytokine storm in the pathogenesis of multiple organ dysfunction syndrome associated with surgical insults. Nihon Geka Gakkai Zasshi 1996, 97, 771–777. [Google Scholar] [PubMed]
- Peresi, E.; Silva, S.M.; Calvi, S.A.; Marcondes-Machado, J. Cytokines and acute phase serum proteins as markers of inflammatory regression during the treatment of pulmonary tuberculosis. J. Bras. Pneumol. 2008, 34, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Kellum, J.A.; Kong, L.; Fink, M.P.; Weissfeld, L.A.; Yealy, D.M.; Pinsky, M.R.; Fine, J.; Krichevsky, A.; Delude, R.L.; Angus, D.C. Understanding the inflammatory cytokine response in pneumonia and sepsis: Results of the Genetic and Inflammatory Markers of Sepsis (GenIMS) Study. Arch. Intern. Med. 2007, 167, 1655–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.N.; Shin, S.A.; Choo, G.S.; Kim, H.J.; Park, Y.S.; Kim, B.S.; Kim, S.K.; Cho, S.D.; Nam, J.S.; Choi, C.S.; et al. Anti-inflammatory effect of quercetin and galangin in LPS-stimulated RAW264. 7 macrophages and DNCB-induced atopic dermatitis animal models. Int. J. Mol. Med. 2018, 41, 888–898. [Google Scholar] [PubMed]
- Vo, V.A.; Lee, J.W.; Chang, J.E.; Kim, J.Y.; Kim, N.H.; Lee, H.J.; Kim, S.S.; Chun, W.; Kwon, Y.S. Avicularin inhibits lipopolysaccharide-induced inflammatory response by suppressing ERK phosphorylation in RAW 264.7 macrophages. Biomol. Ther. 2012, 20, 532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.Y.; Lim, M.S.; Kim, S.I.; Lee, H.J.; Kim, S.S.; Kwon, Y.S.; Chun, W. Quercetin-3-O-β-D-glucuronide suppresses lipopolysaccharide-induced JNK and ERK phosphorylation in LPS-challenged RAW264. 7 cells. Biomol. Ther. 2016, 24, 610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uesugi, M.; Nakajima, K.; Tohyama, Y.; Kohsaka, S.; Kurihara, T. Nonparticipation of nuclear factor kappa B (NFκB) in the signaling cascade of c-Jun N-terminal kinase (JNK)-and p38 mitogen-activated protein kinase (p38MAPK)-dependent tumor necrosis factor alpha (TNFα) induction in lipopolysaccharide (LPS)-stimulated microglia. Brain Res. 2006, 1073, 48–59. [Google Scholar]
- Tang, X.; Metzger, D.; Leeman, S.; Amar, S. LPS-induced TNF-α factor (LITAF)-deficient mice express reduced LPS-induced cytokine: Evidence for LITAF-dependent LPS signaling pathways. Proc. Natl. Acad. Sci. USA 2006, 103, 13777–13782. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Guo, P.; Lv, N.; Huang, D. Lipopolysaccharide-induced tumor necrosis factor-α factor enhances inflammation and is associated with cancer. Mol. Med. Rep. 2015, 12, 6399–6404. [Google Scholar] [CrossRef]
- Kim, H.D.; Cho, H.R.; Moon, S.B.; Shin, H.D.; Yang, K.J.; Park, B.R.; Jang, H.J.; Kim, L.S. l Lee, H.S.; Ku, S.K. Effect of exopolymers from Aureobasidium pullulans on formalin-induced chronic paw inflammation in mice. J. Microbiol. Biotechnol. 2006, 16, 1954–1960. [Google Scholar]
- Cho, Y.H.; Chung, I.K.; Cheon, W.H.; Lee, H.S.; Ku, S.K. Effect of DHU001, a polyherbal formula on formalin-induced paw chronic inflammation of mice. Toxicol. Res. 2011, 27, 95–102. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Not available. |





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, H.J.; Bak, S.G.; Lim, H.J.; Lee, S.W.; Lee, S.; Ku, S.-K.; Park, S.-I.; Lee, S.-J.; Rho, M.-C. Acyclic Triterpenoid Isolated from Alpinia katsumadai Alleviates Formalin-Induced Chronic Mouse Paw Inflammation by Inhibiting the Phosphorylation of ERK and NF-κB. Molecules 2020, 25, 3345. https://doi.org/10.3390/molecules25153345
Lim HJ, Bak SG, Lim HJ, Lee SW, Lee S, Ku S-K, Park S-I, Lee S-J, Rho M-C. Acyclic Triterpenoid Isolated from Alpinia katsumadai Alleviates Formalin-Induced Chronic Mouse Paw Inflammation by Inhibiting the Phosphorylation of ERK and NF-κB. Molecules. 2020; 25(15):3345. https://doi.org/10.3390/molecules25153345
Chicago/Turabian StyleLim, Hyung Jin, Seon Gyeong Bak, Hee Ju Lim, Seung Woong Lee, Soyoung Lee, Sae-Kwang Ku, Sang-Ik Park, Seung-Jae Lee, and Mun-Chual Rho. 2020. "Acyclic Triterpenoid Isolated from Alpinia katsumadai Alleviates Formalin-Induced Chronic Mouse Paw Inflammation by Inhibiting the Phosphorylation of ERK and NF-κB" Molecules 25, no. 15: 3345. https://doi.org/10.3390/molecules25153345