Phytostabilization of Pb-Zn Mine Tailings with Amorpha fruticosa Aided by Organic Amendments and Triple Superphosphate


 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Amendments on pH and EC of Tailings
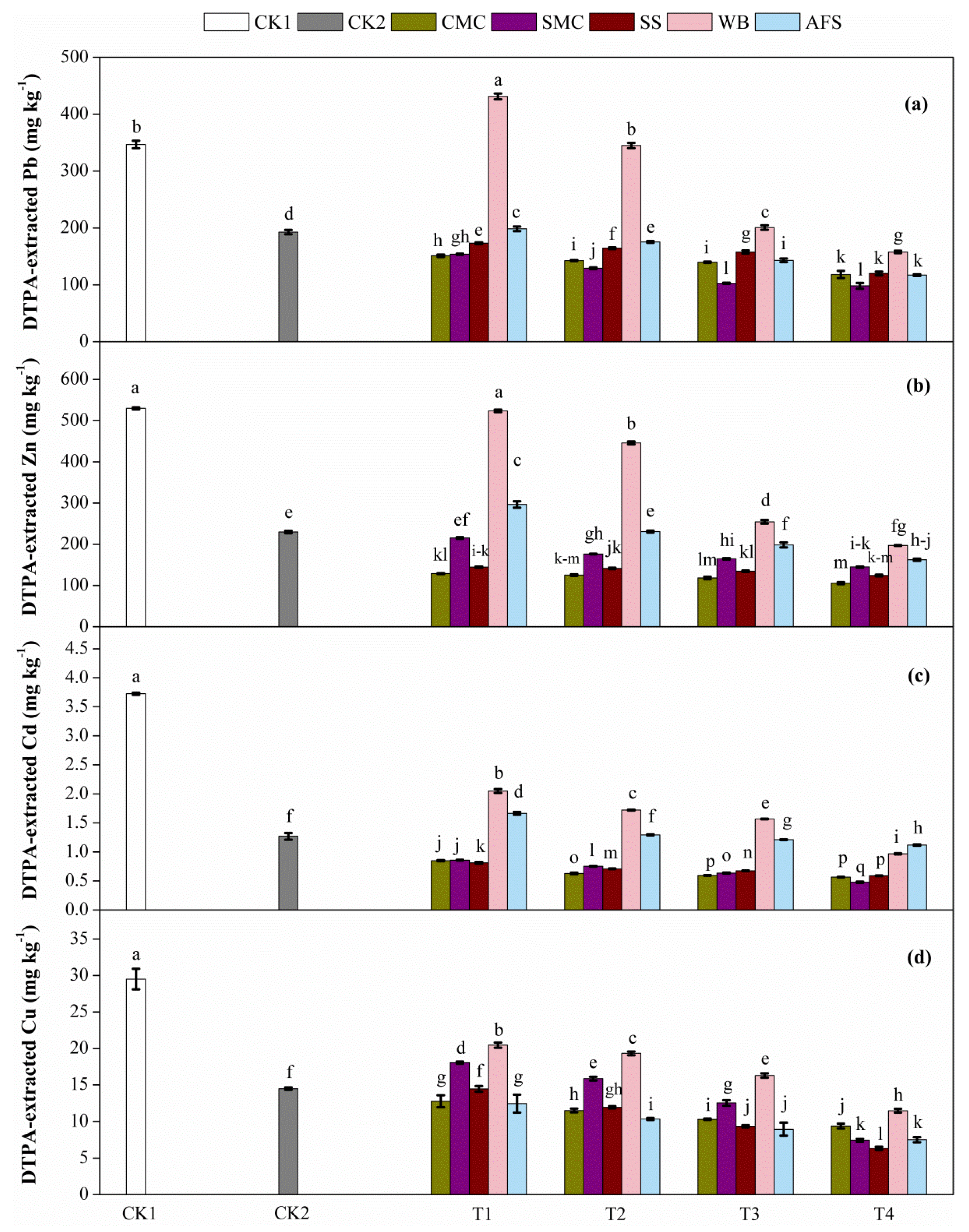
2.2. DTPA-Extracted Pb, Zn, Cd and Cu Concentrations in Treated Tailings
2.3. Impact of Amendments on Dry Biomass Yield and Chlorophyll Content
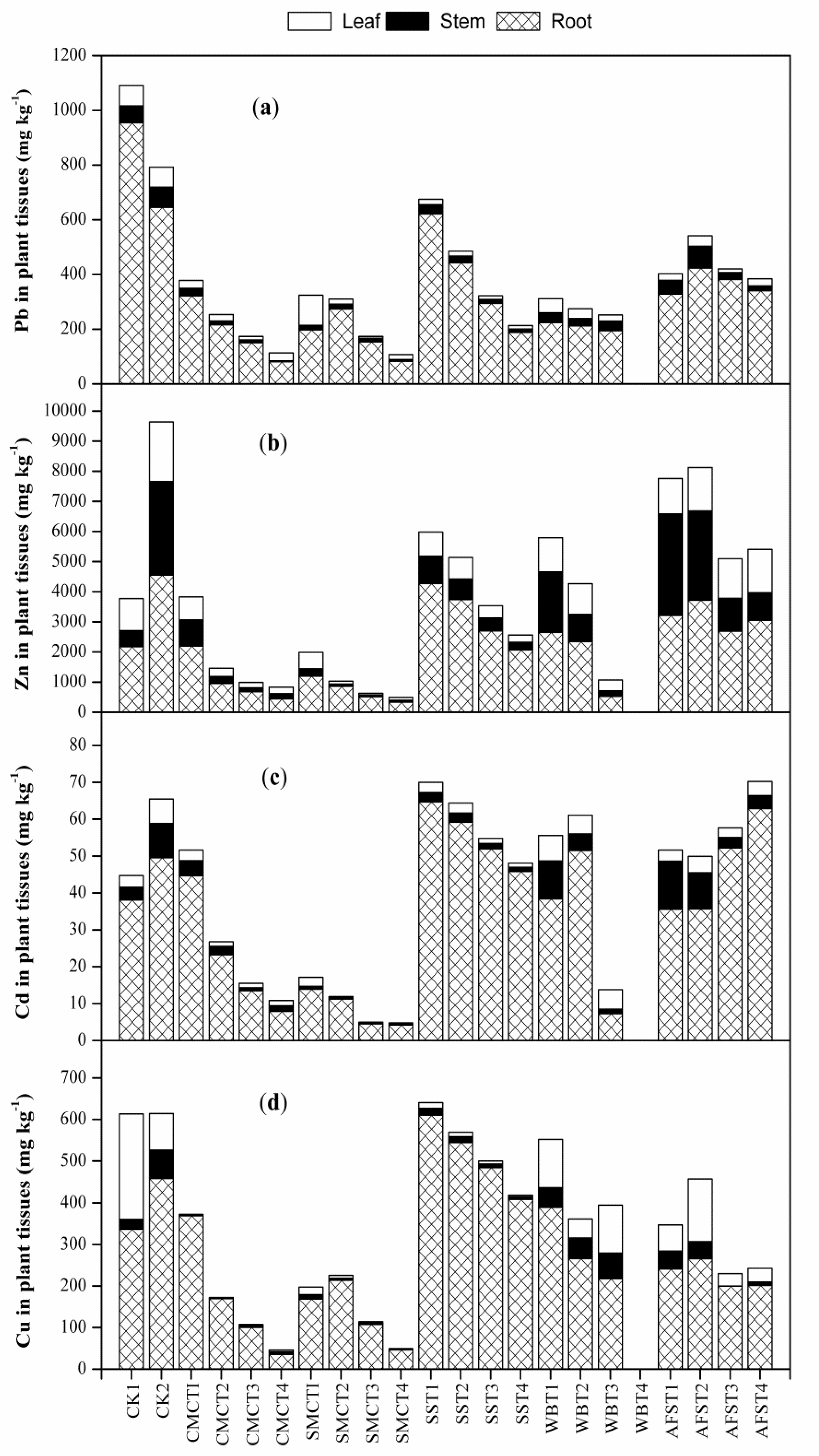
2.4. Effect of Amendments on Pb, Zn, Cd and Cu Uptake by A. Fruticosa
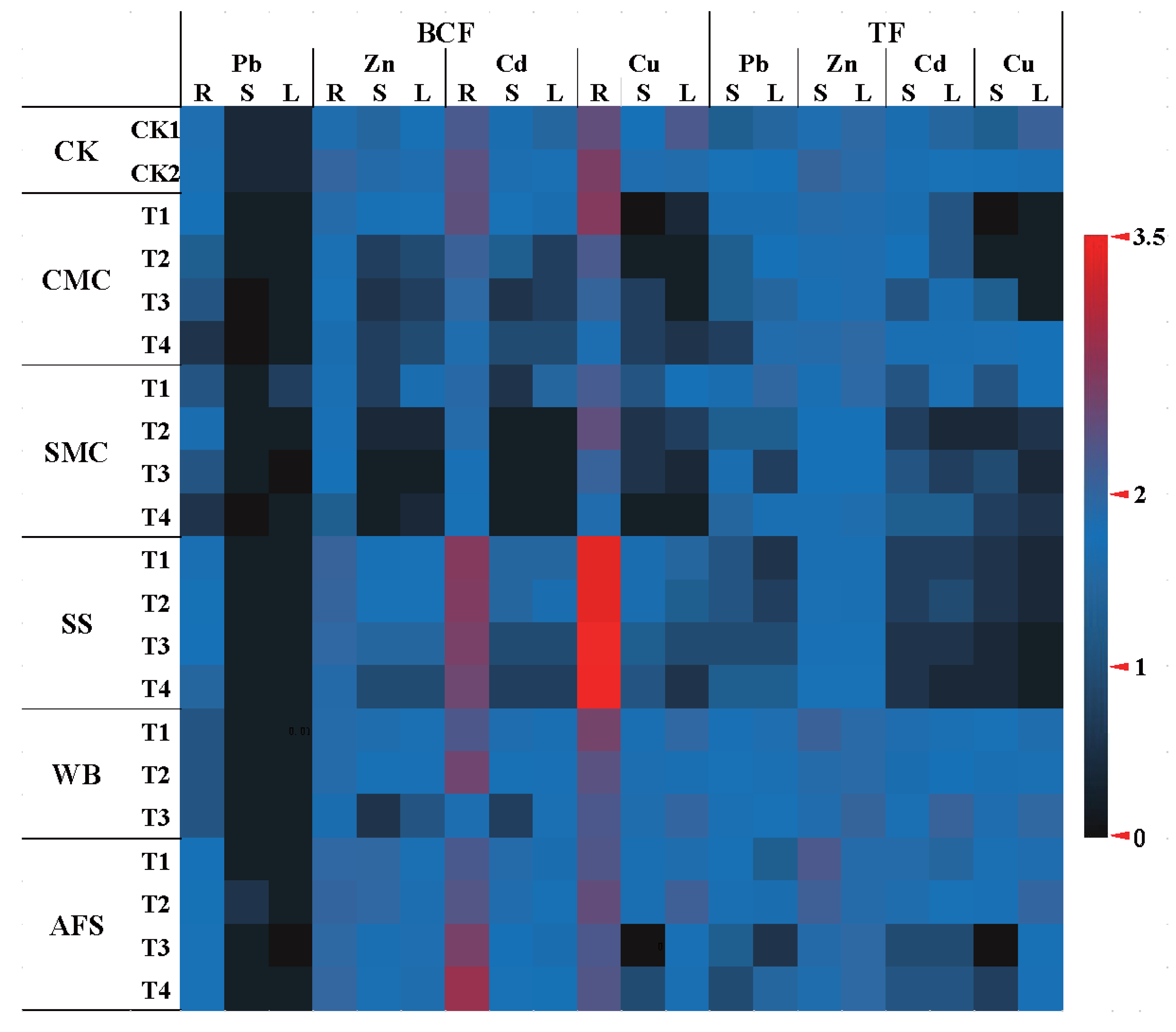
2.5. Phytoremediation Efficiency of A. fruticosa
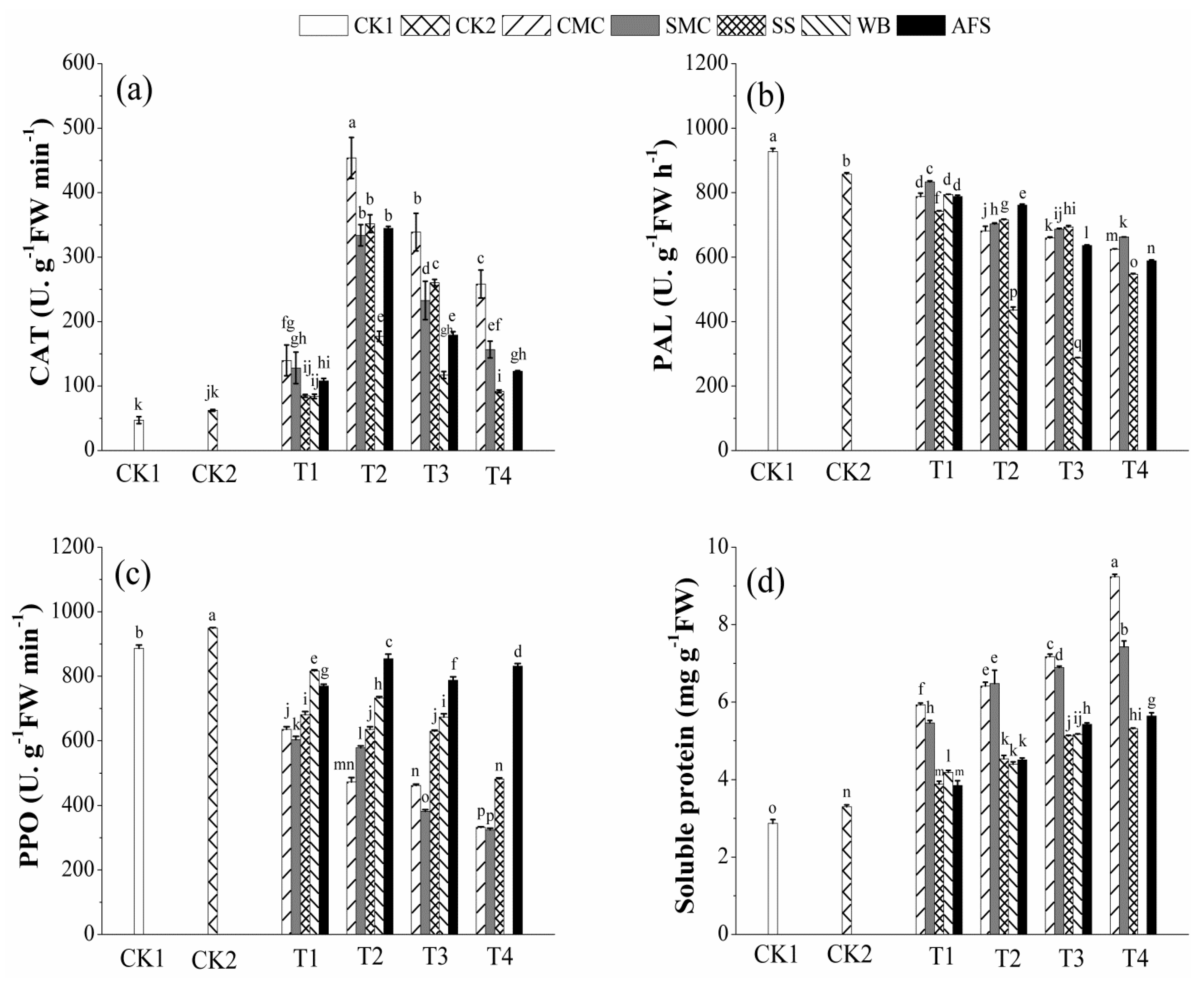
2.6. Effect of Amendments on Antioxidant Enzyme Activities and Soluble Protein Content of A. fruticosa Leaf
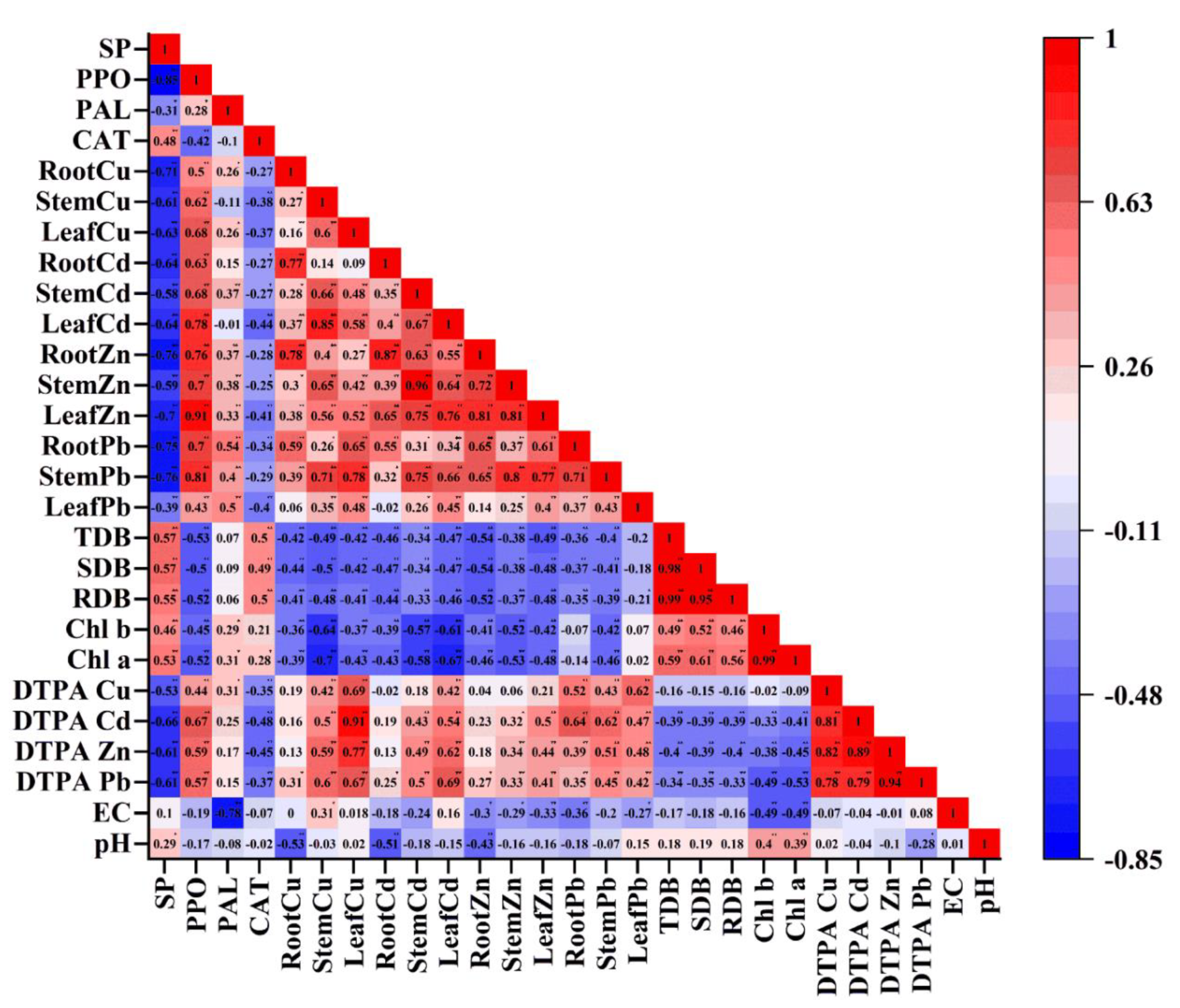
2.7. Relationship among Tailings Properties, Plant Growth Parameters, Metal Concentrations in Plant Tissues, Plant Antioxidant Enzymes and Soluble Protein Content
3. Materials and Methods
3.1. Sampling and Characterization of the Studied Pb-Zn Mine Tailings and Amendments
3.2. Pot Experiment
3.3. Sample Analyses
3.4. Bioconcentration Factor and Translocation Factor
3.5. Quality Control and Statistical Analyses
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ciarkowska, K.; Gargiulo, L.; Mele, G. Natural restoration of soils on mine heaps with similar technogenic parent material: A case study of long-term soil evolution in Silesian-Krakow Upland Poland. Geoderma 2016, 261, 141–150. [Google Scholar] [CrossRef]
- Pająk, M.; Gąsiorek, M.; Cygan, A.; Wanic, T. Concentrations of Cd, Pb and Zn in the top layer of soil and needles of scots pine (Pinus sylvestris L.): A case study of two extremely different conditions of the forest environment in Poland. Fresenius Environ. Bull. 2015, 24, 71–76. [Google Scholar]
- Zhuang, P.; Lu, H.; Li, Z.; Zou, B.; McBride, M.B. Multiple exposure and effects assessment of heavy metals in the population near mining area in South China. PLoS ONE 2014, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, M.H. Ecological restoration of mine degraded soils, with emphasis on metal contaminated soils. Chemosphere 2003, 50, 775–780. [Google Scholar] [CrossRef]
- Chen, T.; Lei, C.; Yan, B.; Xiao, X. Metal recovery from the copper sulfide tailing with leaching and fractional precipitation technology. Hydrometallurgy 2014, 147, 178–182. [Google Scholar] [CrossRef]
- Mendez, M.O.; Maier, R.M. Phytostabilization of mine tailings in arid and semiarid environments - An emerging remediation technology. Environ. Health Persp. 2008, 116, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-H.; Ji, W.; Lee, W.-S.; Koo, N.; Koh, I.H.; Kim, M.-S.; Park, J.-S. Influence of amendments and aided phytostabilization on metal availability and mobility in Pb/Zn mine tailings. J. Environ. Manag. 2014, 139, 15–21. [Google Scholar] [CrossRef]
- Yang, S.-X.; Liao, B.; Yang, Z.-H.; Chai, L.-Y.; Li, J.-T. Revegetation of extremely acid mine soils based on aided phytostabilization: A case study from southern China. Sci. Total Environ. 2016, 562, 427–434. [Google Scholar] [CrossRef]
- Barbosa, B.; Fernando, A.L. Aided phytostabilization of mine waste. In Bio-Geotechnologies for Mine Site Rehabilitation; Elsevier: Amsterdam, The Netherlands, 2018; pp. 147–157. [Google Scholar]
- Branzini, A.; Zubillaga, M. Comparative use of soil organic and inorganic amendments in heavy metals stabilization. Appl. Environ. Soil Sci. 2012, 2012. [Google Scholar] [CrossRef] [Green Version]
- Solís-Dominguez, F.A.; White, S.A.; Hutter, T.B.; Amistadi, M.K.; Root, R.A.; Chorover, J.; Maier, R.M. Response of key soil parameters during compost-assisted phytostabilization in extremely acidic tailings: Effect of plant species. Environ. Sci. Technol. 2012, 46, 1019–1027. [Google Scholar] [CrossRef] [Green Version]
- Mingorance, M.D.; Franco, I.; Rossini-Oliva, S. Application of different soil conditioners to restorate mine tailings with native (Cistus ladanifer L.) and non-native species (Medicago sativa L.). J. Geochem. Explor. 2017, 174, 35–45. [Google Scholar] [CrossRef]
- Qu, J.; Zang, T.; Gu, H.; Li, K.; Hu, Y.; Ren, G.; Xu, X.; Jin, Y. Biosorption of copper ions from aqueous solution by Flammulina velutipes spent substrate. BioResources 2015, 10, 8058–8075. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Jin, Y.; Song, T.; Liang, J.; Bai, X.; Yu, S.; Teng, C.; Wang, X.; Qu, J.; Huang, X. Removal of Cr(VI) by surfactant modified Auricularia auricula spent substrate: Biosorption condition and mechanism. Environ. Sci. Pollut. Res. 2017, 24, 17626–17641. [Google Scholar] [CrossRef] [PubMed]
- Houben, D.; Evrard, L.; Sonnet, P. Mobility, bioavailability and pH-dependent leaching of cadmium, zinc and lead in a contaminated soil amended with biochar. Chemosphere 2013, 92, 1450–1457. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, Y.; Zhang, Y.; Liu, Y.; Liu, S.; Guo, J.; Li, R.; Wu, S.; Chen, B. Growth and metal uptake of energy sugarcane (Saccharum spp.) in different metal mine tailings with soil amendments. J. Environ. Sci. 2014, 26, 1080–1089. [Google Scholar] [CrossRef]
- Cao, X.; Wahbi, A.; Ma, L.; Li, B.; Yang, Y. Immobilization of Zn, Cu, and Pb in contaminated soils using phosphate rock and phosphoric acid. J. Hazard. Mater. 2009, 164, 555–564. [Google Scholar] [CrossRef]
- Hamon, R.; McLaughlin, M.; Cozens, G. Mechanisms of attenuation of metal availability in in situ remediation treatments. Environ. Sci. Technol. 2002, 36, 3991–3996. [Google Scholar] [CrossRef]
- Melamed, R.; Cao, X.; Chen, M.; Ma, L.Q. Field assessment of lead immobilization in a contaminated soil after phosphate application. Sci. Total Environ. 2003, 305, 117–127. [Google Scholar] [CrossRef]
- Weber, J.S.; Goyne, K.W.; Luxton, T.P.; Thompson, A.L. Phosphate treatment of lead-contaminated soil: Effects on water quality, plant uptake, and lead speciation. J. Environ. Qual. 2015, 44, 1127–1136. [Google Scholar] [CrossRef]
- Pulford, I.; Watson, C. Phytoremediation of heavy metal-contaminated land by trees-A review. Environ. Int. 2003, 29, 529–540. [Google Scholar] [CrossRef]
- Seo, K.W.; Son, Y.; Rhoades, C.C.; Noh, N.J.; Koo, J.W.; Kim, J.G. Seedling growth and heavy metal accumulation of candidate woody species for revegetating Korean mine spoils. Restor. Ecol. 2008, 16, 702–712. [Google Scholar] [CrossRef]
- Li, M.S. Ecological restoration of mineland with particular reference to the metalliferous mine wasteland in China: A review of research and practice. Sci. Total Environ. 2006, 357, 38–53. [Google Scholar] [CrossRef] [PubMed]
- Murtaza, G.; Javed, W.; Hussain, A.; Qadir, M.; Aslam, M. Soil-applied zinc and copper suppress cadmium uptake and improve the performance of cereals and legumes. Int. J. Phytoremediat. 2017, 19, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Gu, X.-Q.; Zhang, T.-J.; Liu, H.-H.; Ou, Q.-J.; Peng, C.-L. Phytotoxic effects of Cu, Cd and Zn on the seagrass Thalassia hemprichii and metal accumulation in plants growing in Xincun Bay, Hainan, China. Ecotoxicology 2018, 27, 517–526. [Google Scholar] [CrossRef]
- Srivastava, S.; Dubey, R. Manganese-excess induces oxidative stress, lowers the pool of antioxidants and elevates activities of key antioxidative enzymes in rice seedlings. Plant Growth Regul. 2011, 64, 1–16. [Google Scholar] [CrossRef]
- Mishra, J.; Singh, R.; Arora, N.K. Alleviation of heavy metal stress in plants and remediation of soil by rhizosphere microorganisms. Front. Microbiol. 2017, 8, 1706. [Google Scholar] [CrossRef]
- Etesami, H. Bacterial mediated alleviation of heavy metal stress and decreased accumulation of metals in plant tissues: Mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 147, 175–191. [Google Scholar] [CrossRef]
- Ahmad, P.; Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; John, R.; Egamberdieva, D.; Gucel, S. Role of Trichoderma harzianum in mitigating NaCl stress in Indian mustard (Brassica juncea L) through antioxidative defense system. Front. Plant Sci. 2015, 6, 868. [Google Scholar] [CrossRef] [Green Version]
- Al Mahmud, J.; Hasanuzzaman, M.; Nahar, K.; Bhuyan, M.H.M.B.; Fujita, M. Insights into citric acid-induced cadmium tolerance and phytoremediation in Brassica juncea L.: Coordinated functions of metal chelation, antioxidant defense and glyoxalase systems. Ecotoxicol. Environ. Saf. 2018, 147, 990–1001. [Google Scholar] [CrossRef]
- Salas-Moreno, M.; Contreras-Puentes, N.; Rodríguez-Cavallo, E.; Jorrín-Novo, J.; Marrugo-Negrete, J.; Méndez-Cuadro, D. Protein Carbonylation As a Biomarker of Heavy Metal, Cd and Pb, Damage in Paspalum fasciculatum Willd. ex Flüggé. Plants 2019, 8, 513. [Google Scholar] [CrossRef] [Green Version]
- Jordan, S.N.; Mullen, G.J.; Courtney, R.G. Utilization of spent mushroom compost for the revegetation of lead–zinc tailings: Effects on physico-chemical properties of tailings and growth of Lolium perenne. Biores. Technol. 2008, 99, 8125–8129. [Google Scholar] [CrossRef] [PubMed]
- Gil-Loaiza, J.; White, S.A.; Root, R.A.; Solís-Dominguez, F.A.; Hammond, C.M.; Chorover, J.; Maier, R.M. Phytostabilization of mine tailings using compost-assisted direct planting: Translating greenhouse results to the field. Sci. Total Environ. 2016, 565, 451–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elekes, C.C. Eco-technological solutions for the remediation of polluted soil and heavy metal recovery. In Environmental Risk Assessment of Soil Contamination; IHernández-Soriano, M.C., Ed.; TechOpen: London, UK, 2014; pp. 309–335. [Google Scholar]
- Lahori, A.H.; Zhang, Z.; Shaheen, S.M.; Rinklebe, J.; Guo, Z.; Li, R.; Mahar, A.; Wang, Z.; Ren, C.; Mi, S.; et al. Mono-and co-applications of Ca-bentonite with zeolite, Ca-hydroxide, and tobacco biochar affect phytoavailability and uptake of copper and lead in a gold mine-polluted soil. J. Hazard. Mater. 2019, 374, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Sial, T.A.; Khan, M.N.; Lan, Z.; Kumbhar, F.; Ying, Z.; Zhang, J.; Sun, D.; Li, X. Contrasting effects of banana peels waste and its biochar on greenhouse gas emissions and soil biochemical properties. Process Saf. Environ. Protect. 2019, 122, 366–377. [Google Scholar] [CrossRef]
- Sial, T.A.; Lan, Z.; Khan, M.N.; Zhao, Y.; Kumbhar, F.; Liu, J.; Zhang, A.; Hill, R.L.; Lahori, A.H.; Memon, M. Evaluation of orange peel waste and its biochar on greenhouse gas emissions and soil biochemical properties within a loess soil. Waste Manag. 2019, 87, 125–134. [Google Scholar] [CrossRef]
- Sial, T.A.; Liu, J.; Zhao, Y.; Khan, M.N.; Lan, Z.; Zhang, J.; Kumbhar, F.; Akhtar, K.; Rajpar, I. Co-application of milk tea waste and NPK fertilizers to improve sandy soil biochemical properties and wheat growth. Molecules 2019, 24, 423. [Google Scholar] [CrossRef] [Green Version]
- Arif, M.S.; Riaz, M.; Shahzad, S.M.; Yasmeen, T.; Ashraf, M.; Siddique, M.; Mubarik, M.S.; Bragazza, L.; Buttler, A. Fresh and composted industrial sludge restore soil functions in surface soil of degraded agricultural land. Sci. Total Environ. 2018, 619, 517–527. [Google Scholar] [CrossRef]
- Chiu, K.K.; Ye, Z.H.; Wong, M.H. Growth of Vetiveria zizanioides and Phragmities australis on Pb/Zn and Cu mine tailings amended with manure compost and sewage sludge: A greenhouse study. Bioresour. Technol. 2006, 97, 158–170. [Google Scholar] [CrossRef]
- Foster, E.J.; Hansen, N.; Wallenstein, M.; Cotrufo, M.F. Biochar and manure amendments impact soil nutrients and microbial enzymatic activities in a semi-arid irrigated maize cropping system. Agric. Ecosyst. Environ. 2016, 233, 404–414. [Google Scholar] [CrossRef] [Green Version]
- Lomaglio, T.; Hattab-Hambli, N.; Miard, F.; Lebrun, M.; Nandillon, R.; Trupiano, D.; Scippa, G.S.; Gauthier, A.; Motelica-Heino, M.; Bourgerie, S.; et al. Cd, Pb, and Zn mobility and (bio)availability in contaminated soils from a former smelting site amended with biochar. Environ. Sci. Pollut. Res. 2018, 25, 25744–25756. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Ryan, J.A.; Yang, J. In vitro soil Pb solubility in the presence of hydroxyapatite. Environ. Sci. Technol. 1998, 32, 2763–2768. [Google Scholar] [CrossRef]
- Mahar, A.; Wang, P.; Ali, A.; Guo, Z.; Awasthi, M.K.; Lahori, A.H.; Wang, Q.; Shen, F.; Li, R.; Zhang, Z. Impact of CaO, fly ash, sulfur and Na2S on the (im)mobilization and phytoavailability of Cd, Cu and Pb in contaminated soil. Ecotoxicol. Environ. Saf. 2016, 134, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Gucwa-Przepióra, E.; Małkowski, E.; Sas-Nowosielska, A.; Kucharski, R.; Krzyżak, J.; Kita, A.; Römkens, P.F.A.M. Effect of chemophytostabilization practices on arbuscular mycorrhiza colonization of Deschampsia cespitosa ecotype Waryński at different soil depths. Environ. Pollut. 2007, 150, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-H.; Lai, Y.-J.; Hseu, Z.-Y. Efficacy of cheap amendments for stabilizing trace elements in contaminated paddy fields. Chemosphere 2018, 198, 130–138. [Google Scholar] [CrossRef]
- Clemente, R.; Escolar, A.; Bernal, M.P. Heavy metals fractionation and organic matter mineralisation in contaminated calcareous soil amended with organic materials. Bioresour. Technol. 2006, 97, 1894–1901. [Google Scholar] [CrossRef]
- Chen, J.J.; Zhou, J.M.; Goldsbrough, P.B. Characterization of phytochelatin synthase from tomato. Physiol. Plant. 1997, 101, 165–172. [Google Scholar] [CrossRef]
- Valipour, M.; Shahbazi, K.; Khanmirzaei, A. Chemical immobilization of lead, cadmium, copper, and nickel in contaminated soils by phosphate amendments. CLEAN–Soil Air Water 2016, 44, 572–578. [Google Scholar] [CrossRef]
- Chen, H.-S.; Huang, Q.-Y.; Liu, L.-N.; Cai, P.; Liang, W.; Li, M. Poultry manure compost alleviates the phytotoxicity of soil cadmium: Influence on growth of pakchoi (Brassica chinensis L.). Pedosphere 2010, 20, 63–70. [Google Scholar] [CrossRef]
- Egene, C.E.; Van Poucke, R.; Ok, Y.S.; Meers, E.; Tack, F.M.G. Impact of organic amendments (biochar, compost and peat) on Cd and Zn mobility and solubility in contaminated soil of the Campine region after three years. Sci. Total Environ. 2018, 626, 195–202. [Google Scholar] [CrossRef]
- Lombi, E.; Hamon, R.E.; Wieshammer, G.; McLaughlin, M.J.; McGrath, S.P. Assessment of the use of industrial by-products to remediate a copper- and arsenic-contaminated soil. J. Environ. Qual. 2004, 33, 902–910. [Google Scholar] [CrossRef]
- Mehlhorn, J.; Besold, J.; Lezama Pacheco, J.S.; Gustafsson, J.P.; Kretzschmar, R.; Planer-Friedrich, B. Copper mobilization and immobilization along an organic matter and redox gradient-Insights from a mofette site. Environ. Sci. Technol. 2018, 52, 13698–13707. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.; Curaqueo, G.; Khan, N.; Bolan, N.; Cea, M.; Eugenia, G.M.; Cornejo, P.; Ok, Y.S.; Borie, F. Chicken-manure-derived biochar reduced bioavailability of copper in a contaminated soil. J. Soils Sediments 2017, 17, 741–750. [Google Scholar] [CrossRef]
- Wang, Q.; Awasthi, M.K.; Ren, X.; Zhao, J.; Li, R.; Shen, F.; Zhang, Z. Effect of calcium bentonite on Zn and Cu mobility and their accumulation in vegetable growth in soil amended with compost during consecutive planting. Environ. Sci. Pollut. Res. 2017, 24, 15645–15654. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Shu, W.; Zhang, Z.; Lan, C.; Wong, M.H. Evaluation of major constraints to revegetation of lead/zinc mine tailings using bioassay techniques. Chemosphere 2002, 47, 1103–1111. [Google Scholar] [CrossRef] [Green Version]
- Lucchini, P.; Quilliam, R.S.; DeLuca, T.H.; Vamerali, T.; Jones, D.L. Does biochar application alter heavy metal dynamics in agricultural soil? Agric. Ecosys. Environ. 2014, 184, 149–157. [Google Scholar] [CrossRef]
- Xian, Y.; Wang, M.; Chen, W. Quantitative assessment on soil enzyme activities of heavy metal contaminated soils with various soil properties. Chemosphere 2015, 139, 604–608. [Google Scholar] [CrossRef]
- Küpper, H.; Andresen, E. Mechanisms of metal toxicity in plants. Metallomics 2016, 8, 269–285. [Google Scholar] [CrossRef]
- Zouari, M.; Elloumi, N.; Ahmed, C.B.; Delmail, D.; Rouina, B.B.; Abdallah, F.B.; Labrousse, P. Exogenous proline enhances growth, mineral uptake, antioxidant defense, and reduces cadmium-induced oxidative damage in young date palm (Phoenix dactylifera L.). Ecol. Eng. 2016, 86, 202–209. [Google Scholar] [CrossRef]
- Hosseinzadeh, S.R.; Amiri, H.; Ismaili, A. Effect of vermicompost fertilizer on photosynthetic characteristics of chickpea (Cicer arietinum L.) under drought stress. Photosynthetica 2016, 54, 87–92. [Google Scholar] [CrossRef]
- Chai, M.; Shi, F.; Li, R.; Qiu, G.; Liu, F.; Liu, L. Growth and physiological responses to copper stress in a halophyte Spartina alterniflora (Poaceae). Acta Physiol. Plant. 2014, 36, 745–754. [Google Scholar] [CrossRef]
- Bonanno, G. Trace element accumulation and distribution in the organs of Phragmites australis (common reed) and biomonitoring applications. Ecotoxicol. Environ. Saf. 2011, 74, 1057–1064. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.D.; Ma, L.Q.; Chen, M.; Singh, S.P.; Harris, W.G. Impacts of phosphate amendments on lead biogeochemistry at a contaminated site. Environ. Sci. Technol. 2002, 36, 5296–5304. [Google Scholar] [CrossRef] [PubMed]
- Badrloo, T.A.; Nobariyan, M.R.S.; Bostani, A.; Ghorbani, H.; Abbaspour, A. Solubility and phytoavailability of phosphorus and lead in a contaminated soil amended with two phosphorus fertilizers. Commun. Soil Sci. Plant Anal. 2016, 47, 1967–1974. [Google Scholar] [CrossRef]
- Kushwaha, A.; Hans, N.; Kumar, S.; Rani, R. A critical review on speciation, mobilization and toxicity of lead in soil-microbe-plant system and bioremediation strategies. Ecotoxicol. Environ. Saf. 2018, 147, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Rodriguez, E.; da Conceição Santos, M.; Azevedo, R.; Correia, C.; Moutinho-Pereira, J.; de Oliveira, J.M.P.F.; Dias, M.C. Photosynthesis light-independent reactions are sensitive biomarkers to monitor lead phytotoxicity in a Pb-tolerant Pisum sativum cultivar. Environ. Sci. Pollut. Res. 2015, 22, 574–585. [Google Scholar] [CrossRef]
- Bonanno, G.; Lo Giudice, R. Heavy metal bioaccumulation by the organs of Phragmites australis (common reed) and their potential use as contamination indicators. Ecol. Indic. 2010, 10, 639–645. [Google Scholar] [CrossRef]
- Yang, Y.; Liang, Y.; Ghosh, A.; Song, Y.; Chen, H.; Tang, M. Assessment of arbuscular mycorrhizal fungi status and heavy metal accumulation characteristics of tree species in a lead-Zinc mine area: Potential applications for phytoremediation. Environ. Sci. Pollut. Res. 2015, 22, 13179–13193. [Google Scholar] [CrossRef]
- Pandey, V.C. Phytoremediation of heavy metals from fly ash pond by Azolla caroliniana. Ecotoxicol. Environ. Saf. 2012, 82, 8–12. [Google Scholar] [CrossRef]
- Hart, J.J.; Welch, R.M.; Norvell, W.A.; Clarke, J.M.; Kochian, L.V. Zinc effects on cadmium accumulation and partitioning in near-isogenic lines of durum wheat that differ in grain cadmium concentration. New Phytol. 2005, 167, 391–401. [Google Scholar] [CrossRef]
- Mendez, M.O.; Maier, R.M. Phytoremediation of mine tailings in temperate and arid environments. Rev. Environ. Sci. Biotechnol. 2008, 7, 47–59. [Google Scholar] [CrossRef]
- Abbaspour, A.; Kalbasi, M.; Hajrasuliha, S.; Fotovat, A. Effect of organic matter and salinity on ethylenediaminetetraacetic acid-Extractable and solution species of cadmium and lead in three agricultural soils. Commun. Soil Sci. Plant Anal. 2008, 39, 983–1005. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef] [Green Version]
- Liao, M.T.; Hedley, M.J.; Woolley, D.J.; Brooks, R.R.; Nichols, M.A. Copper uptake and translocation in chicory (Cichorium intybus L. cv. Grasslands Puna) and tomato (Lycopersicon esculentum Mill. cv. Rondy) plants grown in NFT system. I. Copper uptake and distribution in plants. Plant Soil 2000, 221, 135–142. [Google Scholar] [CrossRef]
- Cheng, J.; Qiu, H.; Chang, Z.; Jiang, Z.; Yin, W. The effect of cadmium on the growth and antioxidant response for freshwater algae Chlorella Vulgaris. SpringerPlus 2016, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Feng, M.; Yin, H.; Peng, H.; Liu, Z.; Lu, G.; Dang, Z. Hexavalent chromium induced oxidative stress and apoptosis in Pycnoporus sanguineus. Environ. Pollut. 2017, 228, 128–139. [Google Scholar] [CrossRef]
- Mehmood, S.; Saeed, D.A.; Rizwan, M.; Khan, M.N.; Aziz, O.; Bashir, S.; Ibrahim, M.; Ditta, A.; Akmal, M.; Mumtaz, M.A.; et al. Impact of different amendments on biochemical responses of sesame (Sesamum indicum L.) plants grown in lead-cadmium contaminated soil. Plant Physiol. Biochem. 2018, 132, 345–355. [Google Scholar] [CrossRef]
- Guo, D.; Ali, A.; Ren, C.; Du, J.; Li, R.; Lahori, A.H.; Xiao, R.; Zhang, Z.; Zhang, Z. EDTA and organic acids assisted phytoextraction of Cd and Zn from a smelter contaminated soil by potherb mustard (Brassica juncea, Coss) and evaluation of its bioindicators. Ecotoxicol. Environ. Saf. 2019, 167, 396–403. [Google Scholar] [CrossRef]
- Jiang, K.; Wu, B.; Wang, C.; Ran, Q. Ecotoxicological effects of metals with different concentrations and types on the morphological and physiological performance of wheat. Ecotoxicol. Environ. Saf. 2019, 167, 345–353. [Google Scholar] [CrossRef]
- Duan, C.; Fang, L.; Yang, C.; Chen, W.; Cui, Y.; Li, S. Reveal the response of enzyme activities to heavy metals through in situ zymography. Ecotoxicol. Environ. Saf. 2018, 156, 106–115. [Google Scholar] [CrossRef]
- Gong, Q.; Wang, L.; Dai, T.; Zhou, J.; Kang, Q.; Chen, H.; Li, K.; Li, Z. Effects of copper on the growth, antioxidant enzymes and photosynthesis of spinach seedlings. Ecotoxicol. Environ. Saf. 2019, 171, 771–780. [Google Scholar] [CrossRef]
- Zhang, Y.; Ji, H. Physiological responses and accumulation characteristics of turfgrasses exposed to potentially toxic elements. J. Environ. Manag. 2019, 246, 796–807. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Chan, G.; Zhang, J.; Liang, J.; Wong, M.H. Physiological aspects of vetiver grass for rehabilitation in abandoned metalliferous mine wastes. Chemosphere 2003, 52, 1559–1570. [Google Scholar] [CrossRef]
- Kaya, C.; Akram, N.A.; Sürücü, A.; Ashraf, M. Alleviating effect of nitric oxide on oxidative stress and antioxidant defence system in pepper (Capsicum annuum L.) plants exposed to cadmium and lead toxicity applied separately or in combination. Sci. Hortic. 2019, 255, 52–60. [Google Scholar] [CrossRef]
- Michelet, L.; Roach, T.; Fischer, B.B.; Bedhomme, M.; Lemaire, S.D.; Krieger-Liszkay, A. Down-regulation of catalase activity allows transient accumulation of a hydrogen peroxide signal in Chlamydomonas reinhardtii. Plant Cell Environ. 2013, 36, 1204–1213. [Google Scholar] [CrossRef]
- Sidhu, G.P.S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Tolerance and hyperaccumulation of cadmium by a wild, unpalatable herb Coronopus didymus (L.) Sm.(Brassicaceae). Ecotoxicol. Environ. Saf. 2017, 135, 209–215. [Google Scholar] [CrossRef]
- Chen, J.; Shafi, M.; Li, S.; Wang, Y.; Wu, J.; Ye, Z.; Peng, D.; Yan, W.; Liu, D. Copper induced oxidative stresses, antioxidant responses and phytoremediation potential of Moso bamboo (Phyllostachys pubescens). Sci. Rep. 2015, 5, 13554. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Guo, D.; Mahar, A.; Wang, P.; Ma, F.; Shen, F.; Li, R.; Zhang, Z. Phytoextraction of toxic trace elements by Sorghum bicolor inoculated with Streptomyces pactum (Act12) in contaminated soils. Ecotoxicol. Environ. Saf. 2017, 139, 202–209. [Google Scholar] [CrossRef]
- Ali, A.; Guo, D.; Mahar, A.; Wang, Z.; Muhammad, D.; Li, R.; Wang, P.; Shen, F.; Xue, Q.; Zhang, Z. Role of Streptomyces pactum in phytoremediation of trace elements by Brassica juncea in mine polluted soils. Ecotoxicol. Environ. Saf. 2017, 144, 387–395. [Google Scholar] [CrossRef]
- Ali, A.; Guo, D.; Mahar, A.; Ma, F.; Li, R.; Shen, F.; Wang, P.; Zhang, Z. Streptomyces pactum assisted phytoremediation in Zn/Pb smelter contaminated soil of Feng County and its impact on enzymatic activities. Sci. Rep. 2017, 7, 46087. [Google Scholar] [CrossRef] [Green Version]
- Weisany, W.; Sohrabi, Y.; Heidari, G.; Siosemardeh, A.; Ghassemi-Golezani, K. Changes in antioxidant enzymes activity and plant performance by salinity stress and zinc application in soybean (Glycine max L.). Plant Omics 2012, 5, 60. [Google Scholar]
- Michael, P.I.; Krishnaswamy, M. The effect of zinc stress combined with high irradiance stress on membrane damage and antioxidative response in bean seedlings. Environ. Exp. Bot. 2011, 74, 171–177. [Google Scholar] [CrossRef]
- Younis, U.; Qayyum, M.F.; Shah, M.H.R.; Danish, S.; Shahzad, A.N.; Malik, S.A.; Mahmood, S. Growth, survival, and heavy metal (Cd and Ni) uptake of spinach (Spinacia oleracea) and fenugreek (Trigonella corniculata) in a biochar-amended sewage-irrigated contaminated soil. J. Plant Nutr. Soil Sci. 2015, 178, 209–217. [Google Scholar] [CrossRef]
- Ismaiel, M.M.S.; Said, A.A. Tolerance of Pseudochlorella pringsheimii to Cd and Pb stress: Role of antioxidants and biochemical contents in metal detoxification. Ecotoxicol. Environ. Saf. 2018, 164, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Amm, I.; Sommer, T.; Wolf, D.H. Protein quality control and elimination of protein waste: The role of the ubiquitin-proteasome system. Biochim. Biophys. ActaMolecul. Cell Res. 2014, 1843, 182–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Okekeogbu, I.; Sangireddy, S.; Ye, Z.; Li, H.; Bhatti, S.; Hui, D.; McDonald, D.W.; Yang, Y.; Giri, S. Proteome modification in tomato plants upon long-term aluminum treatment. J. Proteome Res. 2016, 15, 1670–1684. [Google Scholar] [CrossRef] [PubMed]
- Bouker, H.; Yssaad, H.A.R.; Arbaoui, M.; Amaria-Belarbi, A. Effect of zinc on parameters (protein, soluble sugar and proline) in bean (Viciafaba L.). Plant Arch. 2019, 19, 2920–2924. [Google Scholar]
- Nayek, S.; Gupta, S.; Saha, R.N. Metal accumulation and its effects in relation to biochemical response of vegetables irrigated with metal contaminated water and wastewater. J. Hazard. Mater. 2010, 178, 588–595. [Google Scholar] [CrossRef]
- Balk, J.; Schaedler, T.A. Iron cofactor assembly in plants. Annu. Rev. Plant Biol. 2014, 65, 125–153. [Google Scholar] [CrossRef]
- Yao, Y.; Tian, M.; Wu, S. Mineral resources exploitation and sustainable development of Fengxian County in Shanxi province. Miner. Resour. Geol. 2004, 18, 470–475. [Google Scholar]
- USEPA Method 9045D. Soil and Waste pH; United States Environmental Protection Agency: Washington, DC, USA, 2002.
- ASTM D1125-95. Standard Test Methods for Electrical Conductivity and Resistivity of Water; ASTM International: West Conshohocken, PA, USA, 2005. [Google Scholar]
- Sochan, A.; Bieganowski, A.; Ryżak, M.; Dobrowolski, R.; Bartmiński, P. Comparison of soil texture determined by two dispersion units of Mastersizer 2000. Int. Agrophys. 2012, 26, 99–102. [Google Scholar] [CrossRef]
- USEPA Method 9080. Cation-Exchange Capacity of Soils (Ammonium Acetate); United States Environmental Protection Agency: Washington, DC, USA, 1986.
- Qin, Z.; Xie, J.-F.; Quan, G.-M.; Zhang, J.-E.; Mao, D.-J.; DiTommaso, A. Impacts of the invasive annual herb Ambrosia artemisiifolia L. on soil microbial carbon source utilization and enzymatic activities. Eur. J. Soil Biol. 2014, 60, 58–66. [Google Scholar] [CrossRef]
- Alvarenga, P.; Mourinha, C.; Farto, M.; Santos, T.; Palma, P.; Sengo, J.; Morais, M.-C.; Cunha-Queda, C. Sewage sludge, compost and other representative organic wastes as agricultural soil amendments: Benefits versus limiting factors. Waste Manag. 2015, 40, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Knudsen, D.; Peterson, G.; Pratt, P. Lithium, sodium, and potassium. In Methods of Soil Analysis, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1983; pp. 225–246. [Google Scholar]
- USEPA Method 3051A. Microwave Assisted Acid Digestion of Sediments, Sludges, Soils, and Oils; United States Environmental Protection Agency: Washington, DC, USA, 2007.
- Lindsay, W.L.; Norvell, W. Development of a DTPA soil test for zinc, iron, manganese, and copper. Soil Sci. Soc. Am. J. 1978, 42, 421–428. [Google Scholar] [CrossRef]
- Álvarez-Rogel, J.; Tercero Gómez, M.d.C.; Conesa, H.M.; Párraga-Aguado, I.; González-Alcaraz, M.N. Biochar from sewage sludge and pruning trees reduced porewater Cd, Pb and Zn concentrations in acidic, but not basic, mine soils under hydric conditions. J. Environ. Manag. 2018, 223, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Kříbek, B.; Nyambe, I.; Majer, V.; Knésl, I.; Mihaljevič, M.; Ettler, V.; Vaněk, A.; Penížek, V.; Sracek, O. Soil contamination near the Kabwe Pb-Zn smelter in Zambia: Environmental impacts and remediation measures proposal. J. Geochem. Explor. 2019, 197, 159–173. [Google Scholar] [CrossRef]
- Zhu, G.; Xiao, H.; Guo, Q.; Song, B.; Zheng, G.; Zhang, Z.; Zhao, J.; Okoli, C.P. Heavy metal contents and enrichment characteristics of dominant plants in wasteland of the downstream of a lead-zinc mining area in Guangxi, Southwest China. Ecotoxicol. Environ. Saf. 2018, 151, 266–271. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Choudhary, S.P.; Chen, S.; Xia, X.; Shi, K.; Zhou, Y.; Yu, J. Role of brassinosteroids in alleviation of phenanthrenecadmium co-contamination-induced photosynthetic inhibition and oxidative stress in tomato. J. Exp. Bot. 2013, 64, 199–213. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cao, Y.; Ma, C.; Chen, G.; Zhang, J.; Xing, B. Physiological and biochemical responses of Salix integraThunb. under copper stress as affected by soil flooding. Environ. Pollut. 2017, 225, 644–653. [Google Scholar] [CrossRef]
- Anning, A.K.; Akoto, R. Assisted phytoremediation of heavy metal contaminated soil from a mined site with Typha latifolia and Chrysopogon zizanioides. Ecotoxicol. Environ. Saf. 2018, 148, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Salas-Moreno, M.; Marrugo-Negrete, J. Phytoremediation potential of Cd and Pb-contaminated soils by Paspalum fasciculatum Willd. ex Flüggé. Int. J. Phytoremediat. 2020, 22, 87–97. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Upon request the experimental samples will be available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | pH | EC (dS m−1) | Chl a Content (mg g−1FW) | Chl b Content (mg g−1FW) | Shoot Dry Biomass (g pot−1) | Root Dry Biomass (g pot−1) | Total Dry Biomass (g pot−1) |
|---|---|---|---|---|---|---|---|
| CK1 | 6.64 ± 0.14 d–g | 1.08 ± 0.03 j | 1.70 ± 0.01 h | 0.68 ± 0.00 g | 1.80 ± 0.00 e | 0.72 ± 0.00 f | 2.52 ± 0.00 e |
| CK2 | 6.63 ± 0.25 d–h | 1.11 ± 0.04 ij | 0.98 ± 0.07 k | 0.37 ± 0.03 j | 1.78 ± 0.01 e | 0.71 ± 0.00 f | 2.49 ± 0.01 e |
| CMCT1 | 6.69 ± 0.09 c–f | 1.24 ± 0.07 h–j | 2.22 ± 0.03 f | 0.71 ± 0.00 g | 11.82 ± 0.15 a | 16.58 ± 3.44 a | 28.40 ± 3.59 a |
| CMCT2 | 6.56 ± 0.19 e–i | 1.37 ± 0.11 f–j | 2.36 ± 0.01 e | 0.76 ± 0.00 f | 12.21 ± 4.60 a | 19.32 ± 4.29 a | 31.53 ± 5.58 a |
| CMCT3 | 6.63 ± 0.05 d–h | 1.59 ± 0.17 e–g | 2.47 ± 0.01 d | 0.75 ± 0.00 f | 13.48 ± 1.43 a | 19.45 ± 1.88 a | 32.93 ± 1.45 a |
| CMCT4 | 6.62 ± 0.13 d–h | 1.79 ± 0.34 de | 1.71 ± 0.01 h | 0.57 ± 0.00 h | 5.15 ± 2.26 cd | 5.55 ± 3.99 cd | 10.70 ± 6.23 cd |
| SMCT1 | 6.85 ± 0.11 b–d | 1.08 ± 0.03 j | 3.08 ± 0.02 b | 1.13 ± 0.01 b | 5.25 ± 1.58 cd | 4.40 ± 2.25 de | 9.65 ± 3.83 d |
| SMCT2 | 6.84 ± 0.17 b–d | 1.12 ± 0.06 ij | 3.28 ± 0.03 a | 1.16 ± 0.02 a | 7.90 ± 1.88 bc | 8.43 ± 1.44 bc | 16.33 ± 3.21 bc |
| SMCT3 | 6.69 ± 0.15 c–f | 1.28 ± 0.06 g–j | 3.29 ± 0.00 a | 1.17 ± 0.00 a | 8.17 ± 3.90 b | 9.48 ± 5.38 b | 17.65 ± 9.27 b |
| SMCT4 | 7.06 ± 0.10 b | 1.41 ± 0.01 f–i | 2.91 ± 0.02 c | 0.99 ± 0.01 c | 5.22 ± 2.82 cd | 6.02 ± 4.51 b–d | 11.24 ± 7.32 cd |
| SST1 | 6.36 ± 0.15 h–j | 1.24 ± 0.02 h–j | 2.16 ± 0.02 g | 0.83 ± 0.01 e | 1.87 ± 0.00 e | 0.74 ± 0.00 ef | 2.61 ± 0.00 e |
| SST2 | 6.39 ± 0.21 g–j | 1.52 ± 0.07 e–h | 1.26 ± 0.02 i | 0.47 ± 0.00 i | 1.85 ± 0.00 e | 0.73 ± 0.00 f | 2.58 ± 0.00 e |
| SST3 | 6.44 ± 0.10 f–j | 1.98 ± 0.12 d | 1.03 ± 0.01 j | 0.30 ± 0.01 k | 1.80 ± 0.00 e | 0.72 ± 0.00 f | 2.52 ± 0.00 e |
| SST4 | 6.25 ± 0.13 j | 2.31 ± 0.05 c | 0.63 ± 0.02 m | 0.20 ± 0.06 l | 1.75 ± 0.00 e | 0.71 ± 0.00 f | 2.46 ± 0.00 e |
| WBT1 | 6.32 ± 0.20 ij | 1.61 ± 0.25 ef | 0.39 ± 0.00 n | 0.13 ± 0.00 no | 2.52 ± 0.00 de | 0.85 ± 0.00 ef | 3.37 ± 0.00 e |
| WBT2 | 6.49 ± 0.19 f-j | 1.92 ± 0.15 d | 0.17 ± 0.05 o | 0.17 ± 0.01 m | 1.80 ± 0.00 e | 0.78 ± 0.00 ef | 2.57 ± 0.00 e |
| WBT3 | 6.92 ± 0.14 bc | 3.56 ± 0.67 b | 0.10 ± 0.00 o | 0.10 ± 0.01 o | 1.76 ± 0.00 e | 0.75 ± 0.00 ef | 2.51 ± 0.00 e |
| WBT4 | 7.35 ± 0.11 a | 4.65 ± 0.26 a | - | - | - | - | - |
| AFST1 | 6.68 ± 0.21 c–f | 1.18 ± 0.06 ij | 0.38 ± 0.02 n | 0.14 ± 0.01 n | 1.87 ± 0.01 e | 0.80 ± 0.00 ef | 2.67 ± 0.01 e |
| AFST2 | 6.59 ± 0.16 d–i | 1.19 ± 0.01 ij | 0.74 ± 0.01 l | 0.28 ± 0.00 k | 2.22 ± 0.30 e | 0.86 ± 0.04 ef | 3.09 ± 0.34 e |
| AFST3 | 6.44 ± 0.00 f–j | 1.19 ± 0.01 ij | 1.02 ± 0.01 j | 0.39 ± 0.00 j | 2.54 ± 0.15 de | 0.89 ± 0.02 ef | 3.44 ± 0.15 e |
| AFST4 | 6.83 ± 0.06 b–e | 1.20 ± 0.01 ij | 2.23 ± 0.02 f | 0.86 ± 0.01 d | 2.58 ± 0.13 de | 0.91 ± 0.00 ef | 3.50 ± 0.13 e |
| Parameters | Tailings | AFS | CMC | SMC | SS | WB | TSP |
|---|---|---|---|---|---|---|---|
| pH (1:5 H2O) | 7.33 ± 0.07 | 7.85 ± 0.07 | 7.75 ± 0.04 | 7.36 ± 0.04 | 5.44 ± 0.02 | 10.50 ± 0.03 | 2.52 ± 0.01 |
| EC (1:5) (dS m−1) | 1.27 ± 0.01 | 0.15 ± 0.01 | 5.84 ± 0.01 | 5.87 ± 0.08 | 2.41 ± 0.02 | 26 ± 0.4 | 7.54 ± 0.1 |
| Clay% | 11.5 | 36.96 | - | - | - | - | - |
| Silt% | 21.7 | 53.45 | - | - | - | - | - |
| Sand% | 66.8 | 9.59 | - | - | - | - | - |
| Textural class | Sandy loam | Silty clay loam | - | - | - | - | - |
| CEC (cmol kg−1) | 10.53 ± 0.2 | 33.94 ± 0.9 | 62.41 ± 0.8 | 269 ± 0.7 | 72.60 ± 0.6 | 143 ± 0.3 | 127 ± 0.9 |
| Water holding capacity (%) | 15.36 ± 0.4 | 23.40 ± 0.2 | - | - | - | - | - |
| Moisture content (%) | 0.37 ± 0.01 | 0.92 ± 0.09 | 7.27 ± 0.2 | 8.6 ± 0.4 | 8.82 ± 0.1 | 12.46 ± 0.5 | - |
| C% | 4.40 ± 0.0 | 1.30 ± 0.0 | 30.49 ± 0.0 | 25.31 ± 0.0 | 32.28 ± 0.1 | 41.50 ± 0.1 | - |
| Total N (g kg−1) | 0.44 ± 0.0 | 0.33 ± 0.0 | 13.04 ± 0.1 | 11.44 ± 0.1 | 21.30 ± 0.1 | 9.50 ± 0.0 | - |
| Total P (g kg−1) | 0.20 ± 0.0 | 5.62 ± 0.0 | 8.75 ± 0.0 | 1.54 ± 0.0 | 2.19 ± 0.0 | 3.88 ± 0.0 | - |
| Total K (g kg−1) | 8.49 ± 0.0 | 10.87 ± 0.0 | 11.29 ± 0.0 | 8.58 ± 0.0 | 12.31 ± 0.0 | 82.50 ± 0.0 | - |
| Available N (mg kg−1) | 3.77 ± 0.1 | - | - | - | - | - | - |
| Available P (mg kg−1) | 6.69 ± 0.1 | 9.04 ± 0.1 | 351 ± 1.0 | 154 ± 0.1 | 529 ± 0.7 | 146 ± 0.8 | - |
| Available K (mg kg−1) | 3.98 ± 0.1 | 26.51 ± 0.1 | 197 ± 0.4 | 86.23 ± 0. | 498 ± 0.2 | 1051 ± 0.4 | - |
| Total Pb (mg kg−1) | 5614 ± 51 | 62.65 ± 5.9 | 21.42 ± 9.7 | 60.06 ± 3.6 | 38.34 ± 4.3 | 69.31 ± 2.9 | - |
| Total Zn (mg kg−1) | 15439 ± 114 | 109 ± 2.9 | 263 ± 2.4 | 90.84 ± 6.6 | 411 ± 7.1 | 64.35 ± 3.2 | - |
| Total Cd (mg kg−1) | 63.58 ± 1.1 | 2.73 ± 0.2 | 1.87 ± 0.2 | 4.00 ± 0.1 | 1.54 ± 0.1 | 4.57 ± 0.5 | - |
| Total Cu (mg kg−1) | 425 ± 2.9 | 19.67 ± 1.1 | 37.40 ± 0.3 | 14.93 ± 0.5 | 51.46 ± 0.7 | 25.49 ± 0.5 | - |
| Total Ni (mg kg−1) | 8.54 ± 1.2 | 22.49 ± 0.5 | 2.41 ± 0.1 | 1.14 ± 0.1 | 13.48 ± 1.9 | 3.82 ± 0.2 | - |
| Total Cr (mg kg−1) | 8.46 ± 0.4 | 20.35 ± 2.9 | 0.96 ± 0.1 | 7.58 ± 1.4 | 65.30 ± 4.8 | 39.88 ± 1.9 | - |
| Total Ca (mg kg−1) | 94,391 ± 2920 | 25,631 ± 773 | 19,146 ± 765 | 81,112 ± 1394 | 13,246 ± 680 | 54,197 ±1579 | - |
| Total Mg (mg kg−1) | 9051 ± 365 | 7354 ± 206 | 7286 ± 309 | 4761 ± 293 | 4127 ± 46.2 | 8340 ± 336 | - |
| Total Fe (mg kg−1) | 41,624 ± 387 | 25,628 ± 610 | 5488 ± 192 | 4920 ± 55.2 | 8935 ± 26.2 | 9544 ± 321 | - |
| Total Na (mg kg−1) | 305 ± 14.8 | 437 ± 41.6 | 2603 ± 122 | 617 ± 10.3 | 1411 ± 60.8 | 416 ± 18.6 | - |
| Total Mn (mg kg−1) | 1532 ± 30.5 | 630 ± 25.5 | 230 ± 11 | 216 ± 8.2 | 141.80 ± 3.8 | 414 ± 18.0 | - |
| DTPA Pb (mg kg−1) | 296 ± 4.5 | 9.14 ± 0.2 | 11.58 ± 0.4 | 6.29 ± 0.7 | 9.85 ± 0.2 | 13.39 ± 0.3 | - |
| DTPA Zn (mg kg−1) | 1325 ± 3.6 | 4.86 ± 0.2 | 95 ± 2.6 | 5.46 ± 0.3 | 24.12 ± 0.1 | 1.80 ± 0.2 | - |
| DTPA Cd (mg kg−1) | 9.17 ± 0.04 | 0.15 ± 0.0 | 0.17 ± 0.0 | 0.13 ± 0.01 | 0.11 ± 0.01 | 0.28 ± 0.01 | - |
| DTPA Cu (mg kg−1) | 126 ± 4.7 | 1.14 ± 0.03 | 5.82 ± 0.5 | 0.95 ± 0.02 | 3.81 ± 0.3 | 1.15 ± 0.1 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sikdar, A.; Wang, J.; Hasanuzzaman, M.; Liu, X.; Feng, S.; Roy, R.; Sial, T.A.; Lahori, A.H.; Arockiam Jeyasundar, P.G.S.; Wang, X. Phytostabilization of Pb-Zn Mine Tailings with Amorpha fruticosa Aided by Organic Amendments and Triple Superphosphate. Molecules 2020, 25, 1617. https://doi.org/10.3390/molecules25071617
Sikdar A, Wang J, Hasanuzzaman M, Liu X, Feng S, Roy R, Sial TA, Lahori AH, Arockiam Jeyasundar PGS, Wang X. Phytostabilization of Pb-Zn Mine Tailings with Amorpha fruticosa Aided by Organic Amendments and Triple Superphosphate. Molecules. 2020; 25(7):1617. https://doi.org/10.3390/molecules25071617
Chicago/Turabian StyleSikdar, Ashim, Jinxin Wang, Mirza Hasanuzzaman, Xiaoyang Liu, Shulin Feng, Rana Roy, Tanveer Ali Sial, Altaf Hussain Lahori, Parimala Gnana Soundari Arockiam Jeyasundar, and Xiuqing Wang. 2020. "Phytostabilization of Pb-Zn Mine Tailings with Amorpha fruticosa Aided by Organic Amendments and Triple Superphosphate" Molecules 25, no. 7: 1617. https://doi.org/10.3390/molecules25071617