
Iron Transport Tocopheryl Polyethylene Glycol Succinate in Animal Health and Diseases

1
Vets Plus, Inc., Menomonie, WI 54751, USA
2
Breathitt Veterinary Center, Toxicology Department, Murray State University, Hopkinsville, KY 42240, USA
*
Author to whom correspondence should be addressed.
Molecules 2019, 24(23), 4289; https://doi.org/10.3390/molecules24234289
Submission received: 20 June 2019
/
Revised: 18 November 2019
/
Accepted: 18 November 2019
/
Published: 25 November 2019
(This article belongs to the Special Issue Implication of Natural Compounds in Animal Wellbeing)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Gut health is the starting place for maintaining the overall health of an animal. Strategies to maintain gut health are, thus, an important part in achieving the goal of improving animal health. A new strategy to do this involves two molecules: the iron transport protein ovotransferrin (IT) and α-tocopheryl polyethylene glycol succinate (TPGS), which result in the novel formulation of ITPGS. These molecules help reduce gut pathogens, while enhancing the absorption and bioavailability of therapeutic drugs, phytomedicines, and nanomedicines. This, in turn, helps to maintain normal health in animals. Maintaining the gastrointestinal tract (GIT) in its normal condition is key for successful absorption and efficacy of any nutrient. A compromised GIT, due to an imbalance (dysbiosis) in the GIT microbiome, can lead to an impaired GI barrier system with impaired absorption and overall health of the animal. The molecules in ITPGS may address the issue of poor absorption by keeping the GI system healthy by maintaining the normal microbiome and improving the absorption of nutrients through multiple mechanisms involving antioxidative, anti-inflammatory, immunomodulatory, and antimicrobial activities. The ITPGS technology can allow the dose of active pharmaceutical or herbal medicine to be significantly reduced in order to attain equal or better efficacy. With complimentary actions between IT and TPGS, ITPGS presents a novel approach to increase the bioavailability of drugs, phytoconstituents, nutrients, and nanomedicines by enhanced transport to the tissues at the site of action, while reducing gut pathogen load. The ITPGS approach appears to be a novel strategy for maintaining the health of animals by manipulation of microbiota.
1. Introduction
Intestinal health is critically important for the welfare and performance of animals and humans. Enteric diseases that affect the structural integrity of the gastrointestinal tract (GIT) lead to high economic losses due to reduced weight gain, poor feed conversion efficiency, increased mortality rates, and greater medication costs. The overall health of an animal typically starts in the gut. This is due to two key aspects of the gut: it is the site in the body where an animal absorbs nutrients, but also where pathogens are usually encountered first. Researchers have, thus, focused efforts on identifying methods to maintain overall health in animals. The use of nutraceuticals is one such method. In in vivo studies, many nutraceuticals were reported to be poorly absorbed in the GIT, thus limiting bioavailability [1,2,3,4,5,6,7]. The dissolution and ability to permeate the GI wall are considered to be major barriers to drug/phytomedicine bioavailability [6,8]. As a result, the concentration of bioactive phytoconstituents(s) at the target site(s) is very low, thereby having a minimal effect. In addition to low bioavailability, natural products have poor solubility and stability. To overcome these obstacles, bioenhancers of herbal and non-herbal origins are indicated for drugs and phytochemicals [9,10,11]. The use of bioenhancers and potentiators dates back thousands of years [12,13,14]. Piperine from black pepper appears to be the first example of Yogvahi (herbs that are able to increase or potentiate the plasma concentration of drugs) mentioned in Ayurvedic medicine [15,16,17,18]. In other in vivo studies, peppermint oil has been reported to increase bioavailability of cyclosporine [19], and ginseng polysaccharides have been reported to enhance absorption of ginsenoside Rb1 [20].
In in vivo studies, Yurdakok-Dikmen et al. [11] reported that following oral administration, the biodegradation of herbal drugs occurs in the stomach or gut lumen, where membrane permeability and metabolism are key factors in absorption. Bioenhancers are from various classes and exert their effects via multiple mechanisms, such as enzymatic alterations, phytosome formulations, transporter protein alterations, cholagogue/cholerectic effect, and thermogenesis. Absorption of drugs/phytoconstituents can be enhanced by employing various strategies, such as manipulating formulation (micronization, nanocrystals/nanosuspensions, solid solutions, self-dispersing solid solutions with surfactants, lipid solutions, self-emulsifying drug delivery systems, and surfactant-cosolvent systems) [11,21].
In a number of in vivo studies, properties of TPGS (vitamin E TPGS) have been described to enhance the delivery of drugs, nutraceuticals, nanoparticles, and dietary nutrients [6,22]. The poor water solubility of vitamin E limits its absorption in the GIT, but esterification of vitamin E succinate with polyethylene glycol (PEG) 1000 makes it completely water soluble [6,23,24,25]. TPGS has an amphiphilic structure comprising of a hydrophilic polar head portion and lipophilic alkyl tail. It can be functionalized as an excellent solubilizer, emulsifier, and permeation and bioavailability enhancer of hydrophobic drugs and nutraceuticals [6,26,27,28,29]. TPGS can also be used in the development of nanomedicines, focusing on simple structures with multifunctional properties [6,29,30]. Of course, the preparation of nanomedicines is still at the laboratory scale and progress on developing novel nanomedicines is relatively slow. In both in vitro and in vivo studies, TPGS has also been shown to exhibit additional properties, such as a surfactant, apoptogenic, anticancer, immunomodulator, neuroprotection, and reversal of multidrug resistance (MDR) by inhibiting P-glycoprotein (P-gp) activity [6,29,31,32,33,34,35,36]. TPGS 1000 (1000 denoting the PEG chain molecular weight) has been approved by the FDA as a GRAS supplement to solubilize drugs in oral, parenteral, topical, nasal, and rectal/vaginal therapies [31,37,38,39]. TPGS has also been approved by the European Food Safety Authority (EFSA) as a safe pharmaceutical adjuvant used in drug formulation and for research purposes [39,40,41].
Vitamin E TPGS is a powerful tool in the formulation of poorly-soluble lipophilic compounds, as it enhances absorption and bioavailability via efficient micellar uptake into enterocytes. TPGS has a proven record of safety and efficacy in pharmaceutical, dietary supplement, food and beverage, personal care, animal nutrition, and cannabinoid products. It has the added benefit of being a clean label ingredient. In order to achieve greater stability, bioavailability, additional biological/pharmacological effects, and better therapeutic efficacy, TPGS technology has been further improvised by combining the iron transport (IT) protein of ovotransferrin with TPGS. The formulation of Iron Transport Polyethylene Glycol Succinate (ITPGS) is soluble in both fat and water and exerts multiple biological and pharmacological effects, such as antioxidative, anti-inflammatory, antimicrobial, immunomodulatory, healthy gut, and overall health improvement. This review describes various facets of ITPGS, including formulation, biological properties, pharmacological mechanisms, and therapeutic efficacy.
2. Source of Iron Transport Polyethylene Glycol Succinate (ITPGS) and its Formulations
d-α-Tocopheryl polyethylene glycol succinate (TPGS) is prepared from natural lipid-soluble α-tocopherol (vitamin E) by grafting to a polyethylene glycol (PEG) oligomer through a succinate diester linker [30]. The iron transport polyethylene glycol succinate (ITPGS) molecule, in addition to natural α-tocopherol, incorporates the iron transport aspect from ovotransferrin (formerly known as conalbumin) obtained from egg-white or its hydrolysates [42,43,44,45]. The structural formula of ITPGS (IT.TPGS) is shown in Figure 1.
3. Uses of ITPGS
Tocopheryl polyethylene glycol succinate (TPGS) has been used for many purposes, including as an efficient source of natural vitamin E, both for therapeutic and nutrition purposes. Vitamin E TPGS (a water-soluble derivative of vitamin E) is a non-ionic surfactant used as a solubilizer, absorption enhancer, emulsifier, vehicle for lipid-based drug/herbal formulation, and antioxidant. ITPGS, by containing iron from ovotransferrin, offers the possibility of vastly improved effects for maintaining normal health, especially in exerting antimicrobial effects in the gut, where pathogens are typically encountered first in an animal. In in vitro and in vivo studies, ovotransferrin has been reported to be involved in iron transport and iron delivery, and has antibacterial [46,47,48,49], immunomodulatory [44,50,51], and ROS scavenging activities [44,50,51]. By having α-tocopherol and iron transport, ITPGS appears to be a unique and novel composition of two moieties with a wide range of biological and pharmacological activities.
4. ITPGS as a Bioenhancer
In the past, a number of drug delivery systems, including liposomal formulations, nanoparticles (nanocrystals and nanosuspensions), and micellar formulations, have been developed and investigated for enhanced absorption and bioavailability. ITPGS/TPGS may not only increase the bioavailability of pharmaceuticals, herbal products, and nanomedicines, but also of nutrients, which are otherwise poorly soluble and less bioavailable. ITPGS has the potential to maintain a healthy gut environment for normal enterocytes, by improving uptake, transport, and bioavailability of nutrients. The mechanism by which water-soluble TPGS enters the enterocytes and becomes absorbed has been described by Neophytou and Constantinou [25]. The TPGS portion of ITPGS is hydrolyzed to free α-tocopherol in the stomach by nonenzymatic hydrolysis, in the proximity of the brush border epithelium by esterase hydrolysis, and on the surface of enterocytes via a lipase. As such, micelles of ITPGS with other substances can potentially pass through cell membranes, thereby enabling the absorption of the ITPGS, as observed with TPGS [6,37,52].
In vivo and in vitro studies suggest that the absorption, bioavailability, intracellular uptake, and localization of α-tocopherol is greater than any other natural isoforms of vitamin E [53,54,55,56,57,58,59,60]. Following oral administration of α-tocopherol and α-tocotrienol in mice, Khanna et al. [61] found detectable levels of both isoforms in the skin, heart, lungs, brain, liver, bone marrow, and blood, suggesting that these isoforms of vitamin E can be effectively transported to various organs and that they display nonspecific distribution to tissues. In in vivo studies, uptake and distribution of vitamin E have been shown to be increased by making it water soluble, such as in the TPGS or ITPGS form.
In in vivo and in vitro studies, water-soluble vitamin E TPGS has been investigated for various properties, such as solubilizer, micellar property as a surfactant, additive or emulsifier in drug formulations and in drug delivery systems, P-gp inhibitor, oral absorption enhancer, permeation enhancer, and bioavailability enhancer [6,27,30,32,62,63]. The ATP-binding cassette superfamily contains membrane transporter proteins that transport a wide range of diverse compounds across cellular membranes. P-gp is an important member of this family and a multi-specific drug efflux transporter that plays a significant role in governing the bioavailability of many clinically active drugs [64]. P-gp is primarily thought to serve as a natural protective function, by transporting substances, including toxins, from bacteria [65] out of cells. P-gp of epithelial cells is a membrane protein located in the apical membrane and functions as a multidrug efflux pump [66]. TPGS acts as a reversal agent for P-gp mediated MDR and inhibits P-gp mediated drug transport. These findings suggest that enhanced oral bioavailability of drugs co-administered with TPGS may, in part, be due to inhibition of P-gp in the intestine. As observed with TPGS, co-administration of ITPGS may enhance drug solubility, enhance P-gp mediated MDR, and increase the oral bioavailability and cellular uptake of drugs, nanoparticles, and herbal medicines. Collnot et al. [32] investigated TPGS analogs that varied in their PEG chain length, and/or possessed a modified hydrophobic core. Transport studies revealed that modulation of ATPase activity correlated with inhibitory potential for P-gp mediated efflux. These investigators suggested that ATPase inhibition is an essential factor in the inhibitory mechanism of TPGS on cellular efflux pumps [32]. A mechanistic understanding of the inhibition of P-gp by TPGS gives new insights into the structure activity relationship and improves rational design of more potent P-gp inhibitors.
In an in vivo study, Chen et al. [2] demonstrated that TPGS significantly enhanced oral bioavailability of the phytomedicine berberine chloride (BBR): peak concentration (Cmax) increased by 2.9 times and area under the curve (AUC0–36) increased by 1.9 times in rats. It has been reported that BBR is a substrate of P-gp, and that P-gp plays an important role in the absorption of BBR [2,67]. The absorption enhancing property of TPGS may be due to its ability to affect the biological activity of P-gp and thereby reduce the excretion of absorbed BBR into the intestinal lumen. Currently, TPGS is considered a proven bioenhancer of BBR. TPGS has also been reported to enhance absorption and bioavailability of several drugs of different classes [23,39,68,69,70,71,72,73]. In a number of in vivo studies, TPGS has also been shown to enhance absorption and bioavailability of nutraceuticals [2,6,74,75,76]. It is hypothesized that ITPGS is expected to be a better bioenhancer than TPGS because with potentially less pathogenic bacteria present, less energy is required by enterocytes and the gut to fight pathogens.
5. Free Radical Scavenging and Antioxidative Properties
The antioxidative properties of ITPGS are based on their hydrolysis by cytoplasmic esterases that liberate free α-tocopherol, which then localizes in the cell membranes and quenches free radicals. This protects the membrane from lipid peroxidation and damage [30,77,78]. ITPGS works as a free radical scavenger and antioxidant in stressed cells and thereby promotes a healthy environment. Both TPGS and ovotransferrin molecules have shown antioxidant properties via different mechanisms. In both in vitro and in vivo studies, isoforms of natural vitamin E have been reported to exert various biological and pharmacological properties, such as antioxidative, anti-inflammatory, neuroprotective, antithrombotic, and anticancer [25,77,78,79,80].
Previous in vitro and in vivo studies have shown that mitochondria contain the highest concentration of vitamin E and its analogs and that vitamin E accelerates ATP resynthesis [81,82,83]. Vitamin E mainly acts as a chain-breaking antioxidant and radical scavenger, protecting cell membranes against oxidative damage [81,84,85,86]. In addition, vitamin E regulates ROS production [86], maintains oxidative phosphorylation in mitochondria, and accelerates restitution of high-energy phosphates [87,88]. In in vitro studies, mitochondrial targeting α-tocopherol and α-tocopheryl succinate have been shown to exert apoptotic and anticancer properties [83,89,90,91].
From in vitro and in vivo studies, it is well established that α-tocopherol and other isoforms of vitamin E, in addition to antioxidant properties, can interact with enzymes, proteins, lipids, and transcription factors [25,92]. α-Tocopherol can inhibit the function of PKC, 5-lipoxygenase, and phospholipase at the posttranscriptional level and can activate phosphatase 2A. α-Tocopherol is also reported to inhibit cellular proliferation, platelet aggregation, and monocyte attachment [93]. An antioxidative effect can be attributed to ovotransferrin [44] as well as α-tocopherol. Both in vitro and in vivo studies have also shown that ovotransferrin can enhance the antioxidative effect of other nutraceutical ingredients. For example, conjugation of ovotransferrin with catechin showed improved antioxidant activity [94].
6. Anti-Inflammatory Properties
Gastrointestinal inflammation can be induced by a large number of microorganisms [95,96,97,98,99]. The microbial imbalance (dysbiosis) can be caused by several factors, such as diet, disease, infection, medications, toxicants, aging, etc. A dysbiotic microbiota may be sufficient to provoke intestinal inflammation, such as that seen in inflammatory bowel disease, and there is much current interest in quantitative and/or qualitative abnormalities of the microbiota that may be associated with systemic immune, allergic, obesity, metabolic, and infectious disorders, periodontal and respiratory diseases, and cancer [99,100,101,102,103,104,105]. In a number of in vivo and in vitro studies, the interaction between the gut microbiota and the immune system has been investigated [99,106,107,108]. Nieuwenhuizen et al. [109] hypothesized that failure of the gut is an important pathophysiological phenomenon of the generalized inflammatory response that leads to multiple organ dysfunction syndrome (MODS). Abnormal colonization, infection of gut origin, and bacterial translocation are all signs of gut failure that have been implicated in the pathogenesis of MODS.
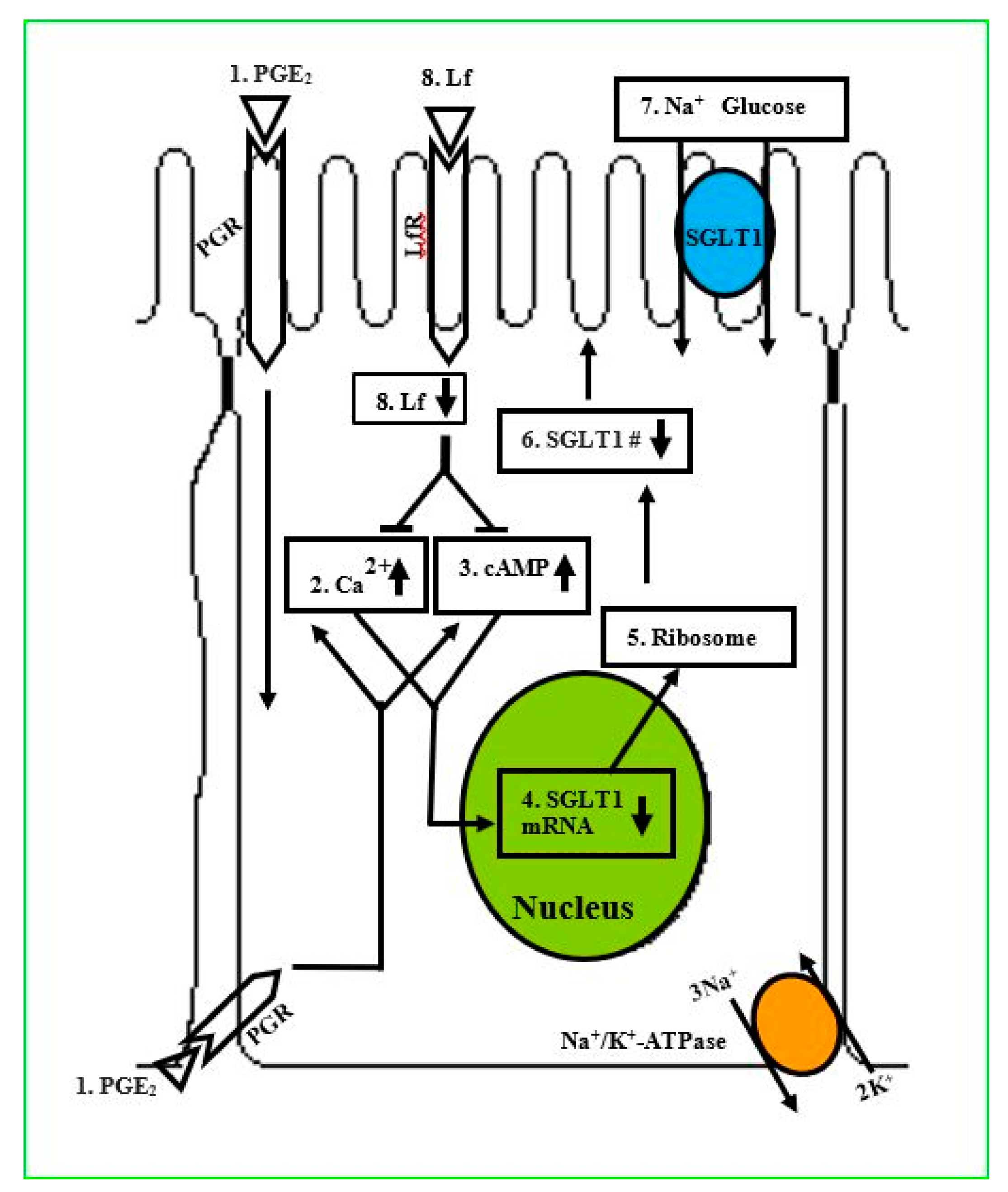
In an in vitro study, Talukder et al. [110] examined the actions of lactoferrin (Lf) to prevent the prostaglandin E2 (PGE2) effect on Na+-dependent glucose cotransport (SGLT1) for glucose absorption in enterocytes. These investigators grew intestinal epithelial cells (IEC-6) on transwell plates, treated with phlorizin, PGE2, AH6809, and Lf, and performed 3-O-methyl d-glucopyranose (OMG) uptake within 10 days postconfluent. Na+-dependent OMG uptake, phlorizin, and immunoblotting studies showed the activity and apical membrane localization of SGLT1 in IEC-6 cells. Findings revealed that PGE2 inhibits SGLT1 in a concentration- and time-dependent manner with an inhibitory constant (Ki) of 50.0 nmol/L and was antagonized by prostanoid receptor inhibitor, AH6809. It was also observed that PGE2 did not alter Na+/K+-ATPase activity. In contrast, quantitative real-time polymerase chain reaction (qRT-PCR) and Western blot analysis revealed that SGLT1-specific transcripts and protein expression level were decreased 3-fold by PGE2. Furthermore, PGE2 treatment increased intracellular cAMP and Ca2+ concentrations and decreased SGLT1 expression on the apical membrane, and these effects were ameliorated by Lf. It was concluded that Lf ameliorates the PGE2 inhibition of SGLT1 via the Ca2+- and cAMP-signaling pathways (Figure 2). Taken together, iron binding protein molecules such as Lf or ovotransferrin appear to play an important role in significantly reducing inflammation of the intestine and restoring the absorption of nutrients.
Figure 2 shows a schematic pathway for glucose absorption in cells of a PGE2-mediated inhibition of SGLT1 that is intercepted by Lactoferrin (Lf). (1) PGE2: extracellular PGE2. PGR: Prostanoid receptors localized on the plasma membrane. (2) Ca2+1: increased intracellular Ca2+ concentration level ([Ca2+]i) because of PGE2. (3) cAMP: increased intracellular cyclic adenosine Monophosphate ([cAMP]i) level because of PGE2. (4) SGLT1 mRNA: SGLT1 mRNA transcription and abundance are decreased because of PGE2. (5) Ribosome: synthesis of SGLT1 protein numbers depend on the abundance of SGLT1 mRNA transcripts. (6) SGLT1 protein no.: quantity of SGLT1 protein on the apical membrane is reduced. (7) Na+-Glucose: Na+-dependent glucose uptake is reduced because of PGE2. LfR: Lf receptors. 8) Lf: Lactoferrin endocytosized by receptor-mediated transcytosis and inhibits the PGE2-mediated upregulation of [Ca2+]i and [cAMP]i levels, and increased Na+-glucose uptake in dependent IEC-6 cells by increasing the Vmax (courtesy of Talukder et al. [110]). It is reasonable to believe that ovotransferrin may also work in the same manner and have a similar mechanism to increase glucose uptake.
ITPGS works as an inflammatory regulator or balancer to maintain normal gut health. Iron is greatly required by pathogenic bacteria in metabolic processes for their growth and replication, in contrast to probiotics (non-pathogenic bacteria) that do not need much iron. The IT molecule sequesters iron and deprives pathogenic bacteria from iron. In addition, IT molecules cause osmolysis. Thus, IT molecules can act as a bacteriostatic or bactericide for pathogenic bacteria, while they do not kill probiotics in the gut. Furthermore, ITPGS appears to maintain a healthy inflammatory response in the body by regulating biomarkers such as interleukins and TNF-alpha. The anti-inflammatory property of ITPGS can also be attributed to vitamin E [78].
7. Antimicrobial Properties
In vitro and in vivo studies suggest that the iron (Fe2+ or Fe3+)-binding protein ‘transferrin’ in ovotransferrin (from egg-white) or lactoferrin (from colostrum and milk) exerts antimicrobial [111,112,113,114,115,116,117,118], immunomodulatory [117], anti-inflammatory [119] properties, and anti-cancer [120] effects. In an in vitro study, Valenti et al. [111] reported that Pseudomonas spp., Escherichia coli, and Streptococcus mutans were the most sensitive and Proteus spp. and Klebsiella spp. were the most resistant to the antibacterial action of ovotransferrin.
IT molecules starve pathogenic bacteria (Gram positive and Gram negative) of iron, and the same IT molecules greatly enhance the probiotic bacteria (Lactobacillus and Bifidobacteria) [118]. In addition, endometritis, usually caused by an infection in 7–20% of dairy cows within 10 days postpartum, deteriorates health, decreases milk production, and causes a huge economic loss to farms. Unpublished in vivo studies with IT molecules showed that rectal temperature returned to normal after one week of intrauterine ovotransferrin treatment. Mean pH of uterine discharge from metritis cows was 8.8 (control 6.8) and changed to 7.0 after one week of ovotransferrin treatment. Mucus score was 6 times higher, and total number of polymorphs was more than 70 times higher in cows with metritis and significantly (p < 0.01) reduced (2 and 4 times, respectively, compared with control) after ovotransferrin treatment. Bacterial culture of uterine swabs demonstrated the presence of different types of bacteria, including Gram positive, Gram negative, α- and β-hemolytic, rods, and cocci. Zone of inhibition studies with different concentrations of ovotransferrin demonstrated that 5% would be more than enough to kill all types of bacteria found in these studies. Thus, it was concluded that IT molecules are a potent antimicrobial protein and have the ability to kill different types of bacteria in the gut and especially pathogenic bacteria that require iron in in vitro and clinical applications. While numerous pathogenic bacteria need iron to survive, many probiotic spp. (such as Lactobacillus spp.) need less iron [121] or no iron [122,123] for their survival. Indeed, orally administered iron has a direct impact on microbial composition in the gut. It can result in reduction in the beneficial microbiota and the expansion of pathobionts, and this can also provide an opportunity for the expansion of enteric pathogens [124]. As such, the concept of limiting or having just enough iron in the gut may have some beneficial aspects for general health of an animal. ITPGS promotes a healthy gut environment for normal microbial growth. Thus, nutrients can be more readily absorbed due to a reduction of pathogenic bacteria that may be competing for nutrients.
In the gut, ITPGS molecules are absorbed, and then exert their biological and pharmacological actions. Ovotransferrin deprives pathogenic bacteria of iron and other nutrients needed for survival. Keep in mind that TPGS inhibits P-gp, and it is necessary to reduce pathogens that may be producing toxins that would normally be removed from enterocytes by P-gp. The combination of TPGS with ovotransferrin allows this to occur. Less pathogenic bacteria present in the GIT may promote the growth of probiotic bacteria, thereby creating a healthy microbial balance. In addition, IT molecules improve the health of intestinal villi by scavenging free radicals and reducing oxidative stress, thus improving the healthy inflammatory response.
8. Effects on Gut Microbiota and Enterocytes
The relevance of gut microbiome and microbiota is well established in health and diseases of humans and animals [99,125,126]. In healthy gut, a microbial ecosystem exists in eubiosis state characterized by a dominance of potentially beneficial microbial species. However, dysbiosis is an imbalance in composition and number of gut commensal microorganisms characterized by a heavy load of pathogenic bacteria [127] and altered gut microbiome [128]. Emerging evidence suggests that dysbiosis may adversely influence the intestinal microbiome and eventually GI structure and function. Indiscriminate use of antibiotics, chemotherapy, and stress have been implicated in the development of dysbiosis. The host’s iron and dietary iron are reported to alter the microbiota composition leading to dysbiosis [128]. In a recent study, the effect of ovotransferrin was investigated on cyclophosphamide-induced intestinal microbiota and dysbiosis [129]. The study suggested that ovotransferrin improved the diversity and richness of the intestinal microbiome, and therefore ITPGS is likely to reverse dysbiosis to eubiosis.
Currently, a large number of in vivo studies suggest that nutraceuticals (including prebiotics, probiotics, and synbiotics) are used to maintain the normal gut biota and to prevent or treat GIT-related diseases [117,118,130,131,132,133,134,135,136,137,138]. The GIT contains a complex polymicrobial community that consists of bacteria, viruses, archaea, fungi, and parasites [99,139,140,141]. The bacterial community is the most prominent and important factor in maintaining homeostasis of this complex gut environment. The human gut microbiota is composed of more than one hundred trillion microbial cells, including aerobic and anaerobic species as well as Gram-positive and Gram-negative species [142]. The term ‘microbiota’ is defined as a set of commensal, autochthonic microorganisms, co-existing with a host without causing any harm [143,144]. Freter [145] defined intestinal microbiota as “the usually complex mixture of bacterial populations that colonize a given area of the GIT in individual human or animal hosts that have not been affected by medical or experimental intervention or disease”. Gut microbiota differ substantially among animal strains and species, gender, and between humans and animals [99,135,140,141,146,147,148].
It is well established that microbiota plays a great role in the health of the GIT as well as of other vital organs [149,150,151,152,153]. The GIT is the largest reservoir of commensal bacteria in the human or animal body, providing nutrients and space for the survival of microbes while concurrently operating mucosal barriers to confine the microbial population [154,155]. Commensal organisms are known to promote gut health through the induction of mucus production and enterocyte turnover [156]. As such, the gut microbiota is a key factor in shaping the biochemical profile of the diet and, therefore, its impact on host health and disease [141,157,158,159,160]. In an in vitro study, Yu et al. [155] also described that the epithelial cells linked by tight junctions not only physically separate the microbiota from the lamina propria, but also secrete proinflammatory cytokines and reactive oxygen species (ROS) in response to pathogen invasion and metabolic stress and serve as a sentinel to the underlying immune cells. Commensal bacteria are involved in various physiological functions in the gut, and inflammation and microbial imbalance (dysbiosis) may cause bacterial invasion and pathology [140,155,161,162,163,164,165].
In in vitro and in vivo studies, Rowland et al. [160] described that gut microbiota-derived products can be absorbed from the gut and enter the host’s endogenous and exogenous pathways to influence the overall metabolic phenotype of the host. In addition, metabolites generated by the host can be secreted into the gut via the enterohepatic circulation, and serve as substrates for the resident microbes. It is evident from in vivo and in vitro studies that gut microbiota not only influence the metabolism, health, and diseases of GIT, but also of other vital organs, including liver, kidney, nervous system, and immune system [99,140,142,153,155,158,166,167,168,169,170,171,172]. Recent in vivo studies investigating interactions between microbial communities and the host have shown that gut bacteria can cause or contribute to the occurrence of conditions such as allergies, inflammatory bowel diseases, rectal cancer, diabetes, obesity, and behavioral alterations [101,103,105,108,141,153,173,174]. In fact, failure of the gut can lead to MODS, as in most cases, gut dysfunction precedes the development of MODS [109,172,175,176]. It is plausible that gut bacteria may be initiating and perpetuating the production of local inflammatory mediators, which may produce distant organ injury [105,109,152,153,172,177,178].
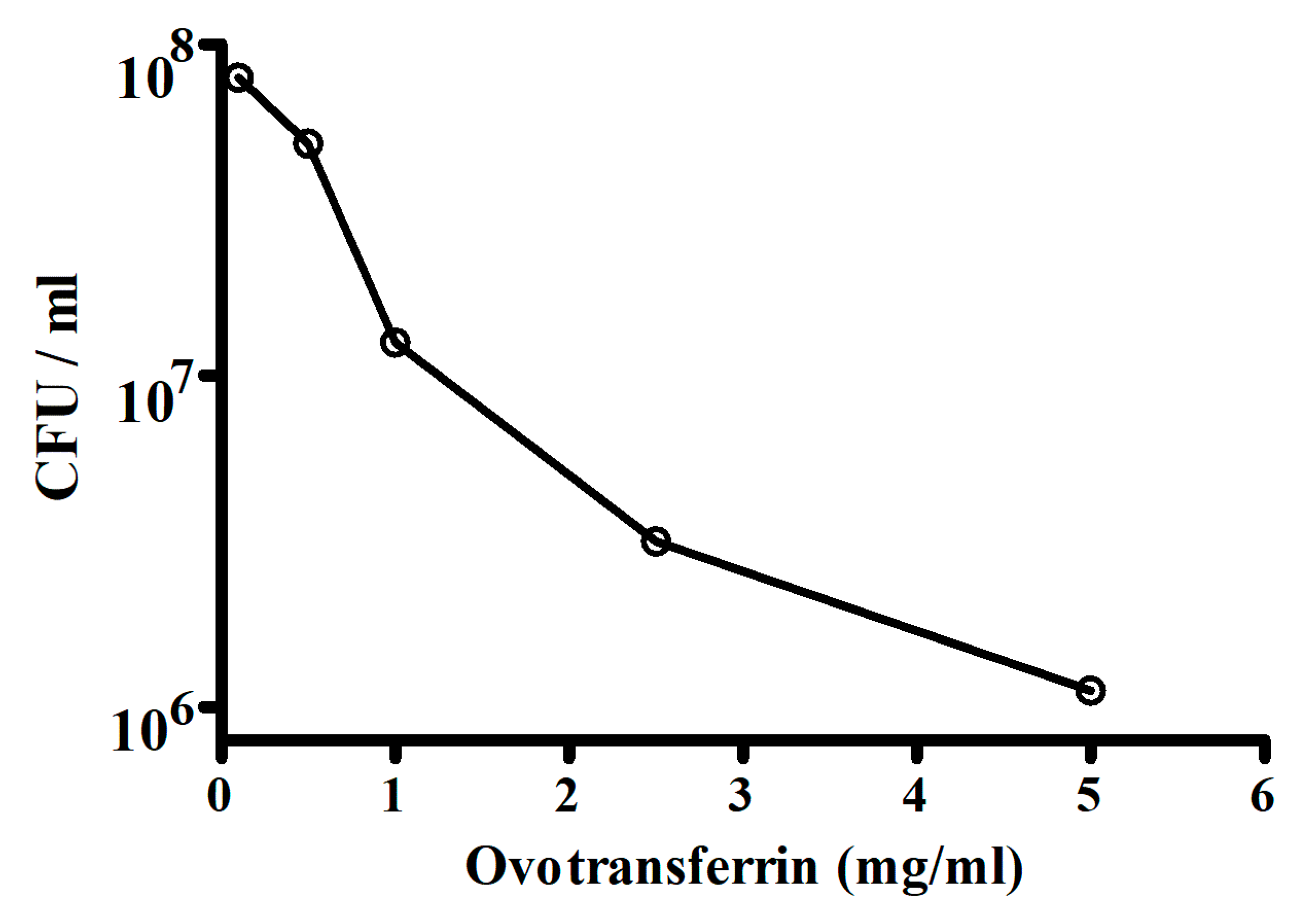
Escherichia coli is one of the important causes of diarrhea in humans and animals. The antibacterial activity of ovotransferrin towards Escherichia coli strain W1485 was tested by Valenti et al. [111]. The bacterial growth was measured in colony-forming units (CFU) and the results of the antibacterial activity of ovotransferrin are shown in Figure 3. The antibacterial activity increased with the concentrations of ovotransferrin. They indicated that in the presence of ovotransferrin, slow changes that progressively limit their ability to grow may occur in the microorganisms. It could be argued therefore that these changes reflect a direct interaction of the protein with the bacteria or a deprivation of an internal iron pool in slow equilibrium with the medium or both.
Lactoferrin is an iron-binding glycoprotein, belonging to the transferrin protein family that ovotransferrin also belongs to. Lactoferrin has antioxidant and antimicrobial properties and has been used for treating diarrhea [179]. Other uses include stimulating the immune system, preventing tissue damage related to aging, promoting healthy intestinal bacteria, preventing cancer, and regulating the way the body processes iron. The iron binding actions of ovotransferrin and lactoferrin make it useful to research for functional similarity and functional substitutions in formulations. For example, in vitro studies Valenti et al. [180] showed that ovotransferrin and lactoferrin have nearly identical abilities for antimicrobial activity against E. coli and Streptococcus mutans. This suggests functional equivalency between ovotransferrin and lactoferrin for at least their bacteriostatic actions against pathogenic bacteria and provides further rationale for ovotransferrin in formulations that may have relied on lactoferrin.
An example of where ovotransferrin may have equivalency with lactoferrin is in an in vivo study of Talukder and Harada [181], who investigated the effects of the iron-binding multifunctional glycoprotein, bovine lactoferrin (BLf), on lipopolysaccharide (LPS)-induced diarrheagenic activity, GI transit, and intestinal fluid content in mice. These investigators found that LPS accumulated abundant fluid in the small intestine in a dose-dependent manner, induced diarrhea, but decreased GI transit. It was observed that pretreatment with BLf significantly attenuated the effects of LPS on diarrheagenic activity and intestinal content, but reversed GI transit time, when compared with NG-nitro-l-arginine-methyl ester (L-NAME, a non-selective NOS inhibitor) or indomethacin (an inhibitor of prostaglandin synthesis). In addition, both plasma NO and PGE2 in enterocytes were found to increase in LPS-treated mice and were reversed by BLf. It was concluded that the action of BLf against LPS was specific and that it exerts antidiarrheal and anti-inflammatory activities by modulating the cyclooxygenase pathway (NO and PGE2) in the gut [181]. As ovotransferrin and lactoferrin have similar antimicrobial actions, it is possible that ovotransferrin may have similar modes of action as lactoferrin for potential antidiarrheal and anti-inflammatory activities.
In an in vitro study, the turnover rates of intestinal epithelial cells (enterocytes) are governed by the pace of crypt cell proliferation and villus/surface cell shedding [155]. In vitro studies have shown that tight junctional proteins are present at the base of basolateral membranes between two neighboring enterocytes flanking the extruding cells, and thus barrier functions are sustained at the villus tips [182]. Excessive epithelial cell death by pathogenic microbes [183,184], metabolic stress [185], nonsteroidal anti-inflammatory drugs (NSAIDS), and acidic and enzymatic agents [186,187] may lead to villous surface denaturation and gut leakiness.
In essence, the use of microbiota manipulation to improve animal health is a promising and ongoing field of research [141,188]. Nutraceuticals are commonly used to improve gut dysbiosis, but their bioavailability is often very poor. ITPGS has been shown to facilitate the maintenance of a healthy microbiota and microbiome in the GIT in rodents, and therefore may enhance the absorption of nutraceuticals [189].
9. Immunomodulatory Properties
Intestinal epithelium maintains a physical barrier to prevent microbial invasion while concomitantly modulating an immune component, which activates host defenses to eliminate invading pathogens [108,176]. These barrier and immune functions of the intestinal epithelium coexist with their equally important mission of maintaining an optimal environment for absorption of nutrients and maintenance of electrolyte balance.
In vitro and in vivo studies demonstrate that normal microbiota exerts profound effects on the mucosal immune system in the gut and regulates immune responses outside the gut [99,100,150,190,191,192]. Mazmanian et al. [193] provided the molecular basis for host-bacterial symbiosis and revealed the archetypal molecule of commensal bacteria (for example, polysaccharide from Bacteroides fragilis) that mediates development of the host immune system. The microbiota plays a pivotal role in regulating immunological tolerance to allergen exposure outside the GIT. IT molecules, of which ovotransferrin is the donor of iron for the ITPGS, have a wide range of biofunctions, and are a major component of the mammalian innate immune system.
10. ITPGS in Enteric Diseases
Porcine epidemic diarrhea virus (PEDV) is a corona virus, which causes an enteric disease only in pigs. It produces acute, severe diarrhea that transmits rapidly to pigs of all ages. Our unpublished data from an in vivo study show that ovotransferrin (IT) is also effective in controlling diarrhea caused by PEDV. Different groups of piglets were treated with electrolytes and IT. There was complete protection against mortality in IT-treated groups. Data showed that electrolytes and IT, in combination, were potent enough to control severe diarrhea.
Avian coccidiosis is an infectious protozoan disease caused by Eimeria. It is one of the most important diseases of poultry, which causes tremendous loss to the industry throughout the world. These protozoa can infect and multiply within the mucosal epithelia in different parts of the gut via oral ingestion. As a result, they cause inflammation, hemorrhage, and diarrhea, and may lead to death. Thus, morbidity and mortality in poultry cost the poultry industry a global loss of about 2.4 billion US dollars annually. Seventy percent of the estimated cost is due to subclinical coccidiosis, as it impacts weight gain and feed conversion ratio (FCR). Recent unpublished data of an in vivo study show that IT can prevent E. tenella infestation in 7–21 days old chicks. In addition, weight gain was significantly increased in IT-treated groups. Thus, IT can be used to treat and control subclinical coccidiosis to prevent morbidity and mortality in the poultry industry.
11. Concluding Remarks and Future Perspective
Improving gut health is a sound strategy for improving the overall health of an animal. Any new method to do this will be an improvement in the art; this novel ITPGS strategy is one such development. Iron transport polyethylene glycol succinate (ITPGS) is a unique formulation consisting of ovotransferrin from a natural source (eggs) and an α-tocopherol format. The ability of the iron binding and tocopheryl polyethylene glycol succinate appear to exert complimentary actions in the body. ITPGS may serve as a multifunction molecule that can be a solubilizer, stabilizer, surfactant, additive, emulsifier, absorption enhancer, and permeation enhancer. It can offer sustained and targeted delivery of pharmaceuticals, nutraceuticals, and nanomedicines. ITPGS exerts antioxidative, anti-inflammatory, immunomodulatory, antimicrobial and many other biological and pharmacological actions. The ITPGS formulation may prove to be a superior biomaterial for enhancing the efficacy of micelles, nanoparticles, liposomes, or other drug delivery systems and this may have hope for the future. Studies have already shown that ITPGS formulation is efficacious and safe in clinical settings for treating enteric diseases via antimicrobial and antioxidant activities. As microbial imbalances can be caused by several factors resulting in comorbidity situations in the gut, the use of ITPGS may be of great use, especially if polypharmacy approaches are used to treat a patient. This would potentially be due to less total drugs being used in a patient and the chance of more cross drug interactions or toxicities could be minimized with the IPTGS method. The influence of commensal microbiota and its associated medical comorbidities are only beginning to be uncovered. However, it is clear that multi-morbidity and the associated use of multiple medicines is common in older populations [194]. The use of ITPGS methodology when polypharmacy [195,196,197,198] approaches are used may have real value in aging populations of both humans and animals. While further research needs to be conducted for its application in these comorbidity and polypharmacy conditions, the possibility that this strategy can help maintain or improve gut health, and thus improve the overall health of animals, seems very hopeful.
Funding
This research received no external funding.
Acknowledgments
The authors would like to thank Denise M. Gupta and Robin B. Doss for their technical assistance in preparation of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kang, M.J.; Cho, J.Y.; Shim, B.H.; Kim, D.K.; Lee, J. Bioavailability enhancing activities of natural compounds from medicinal plants. J. Med. Plant Res. 2009, 3, 1204–1211. [Google Scholar]
- Chen, W.; Miao, Y.-Q.; Fan, D.-J.; Yang, S.-S.; Lin, X.; Meng, L.-K.; Tang, X. Bioavailability study of erberine and the enhancing effects of TPGS on intestinal absorption in rats. AAPS Pharmscitech 2011, 12, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Jhanwar, B.; Gupta, S. Biopotentiation using herbs: Novel technique for poor bioavailable drugs. Int. J. PharmTech Res. 2014, 6, 443–454. [Google Scholar]
- Ramalingam, P.; Ko, Y.T. Improved oral delivery of resveratrol from N-trimethyl chitosan-g-palmitic acid surface-modified solid lipid nanoparticles. Colloids Surf. B Biointerfaces 2016, 139, 52–61. [Google Scholar] [CrossRef]
- Gupta, R.C.; Lall, R.; Srivastava, A.; Sinha, A. Hyaluronic acid: Molecular mechanisms and therapeutic trajectory. Front. Vet. Sci. 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yan, L.; Tang, E.K.Y.; Zhang, Z.; Chen, W.; Liu, G. Synthesis of TPGS/curcumin nanoparticles by thin-film hydration and evaluation of their anti-colon cancer efficacy in in vitro and in vivo. Front. Pharmacol. 2019, 10, 769. [Google Scholar] [CrossRef]
- Pandit, A.P.; Joshi, S.R.; Dalal, P.S.; Patole, V.C. Curcumin as a permeability enhancer enhanced the antihyperlipidemic activity of dietary green tea extract. BMC Complement. Altern. Med. 2019, 19. [Google Scholar] [CrossRef]
- Ning, Z.; Wang, C.; Liu, Y.; Song, Z.; Ma, X.; Liang, D.; Liu, Z.; Lu, A. Integrating strategies of herbal metabolomics, network pharmacology, and experiment validation to investigate Frankincense processing effects. Front. Pharmacol. 2018. [Google Scholar] [CrossRef]
- Kesarwani, K.; Gupta, R. Bioavailability enhancers of herbal origin: An overview. Asian Pac. J. Trop. Biomed. 2013, 3, 253–266. [Google Scholar] [CrossRef]
- Ajazuddin, A.A.; Alexander, A.; Qureshi, A.; Kumari, L.; Vaishnav, P.; Sharma, M.; Saraf, S.; Saraf, S. Role of herbal bioactives as a potential bioavailabilty enhancer for active pharmaceutical ingredients. Fitoterapia 2014, 97, 1–14. [Google Scholar] [CrossRef]
- Yurdakok-Dikmen, B.; Turgut, Y.; Filazi, A. Herbal bioenhancers in veterinary phytomedicine. Front. Vet. Sci. 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Devi, S.; Patel, J.H.; Patel, U.D.; Bhavsar, S.K.; Thaker, A.M. Indian herbal bioenhancers: A review. Pharmacogn. Rev. 2009, 3, 90–91. [Google Scholar]
- Drabu, S.; Khatri, S.; Babu, S.; Lohani, P. Use of herbal bioenhancers to increase the bioavailability of drugs. Res. J. Pharm. Biol. Chem. Sci. 2011, 2, 107–119. [Google Scholar]
- Jain, G.; Patil, U.K. Strategies for enhancement of bioavailability of medicinal agents with natural products. Int. J. Pharm. Sci. Res. 2015, 6, 5315–5324. [Google Scholar]
- Atal, C.K.; Dubey, R.K.; Singh, J. Biochemical basis of enhanced drug bioavailability by piperine: Evidence that piperine is a potent inhibitor of drug metabolism. J. Pharmacol. Exp. Ther. 1985, 232, 258–262. [Google Scholar]
- Annamalai, A.R.; Manavalan, R. Effects of “Trikatu” and its individual components and piperine on gastrointestinal tracts: Trikatu: A bioavailable enhancer. Indian Drugs 1990, 27, 595–604. [Google Scholar]
- Johri, R.K.; Zutshi, U. An Ayurvedic formulation ‘Trikatu’ and its constituents. J. Ethnopharmacol. 1992, 37, 85–91. [Google Scholar] [CrossRef]
- Patil, U.K.; Singh, A.; Chakraborty, A.K. Role of piperine as a bioavailability enhancer. Int. J. Recent Adv. Pharm. Res. 2011, 4, 16–23. [Google Scholar]
- Wacher, V.J.; Wong, S.; Wong, H.T. Peppermint oil enhances cyclosporine oral bioavailability in rats: Comparison with D-alpha-tocopheryl poly(ethylene glycol 1000) succinate (TPGS) and ketoconazole. J. Pharm. Sci. 2002, 91, 77–90. [Google Scholar] [CrossRef]
- Shen, H.; Gao, X.-J.; Li, T.; Jing, W.H.; Han, B.L.; Jia, Y.M.; Yan, Z.X.; Li, S.L.; Yan, R. Ginseng polysaccharides enhanced ginsenoside Rb1 and microbial metabolites exposure through enhancing intestinal absorption and affecting gut microbial metabolism. J. Ethnopharmacol. 2018, 216, 47–56. [Google Scholar] [CrossRef]
- Gupta, S.; Kesarla, R.; Omri, A. Formulation strategies to improve the bioavailability of poorly absorbed drugs with special emphasis on self-emulsifying systems. ISRN Pharm. 2013, 2013, 848043. [Google Scholar] [CrossRef] [PubMed]
- Zou, T.; Gu, L. TPGS emulsified zein nanoparticles enhanced oral bioavailability of daidzin: In vitro characteristics and in vivo performance. Mol. Pharm. 2013, 10, 2062. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H.; Hopkins, W.K. Characteristics of D-α-tocopheryl PEG 1000 succinate for applications as an absorption enhancer in drug delivery systems. Pharmacol. Technol. 1999, 23, 52–68. [Google Scholar]
- Zhang, Z.; Tan, S.; Feng, S.S. Vitamin E TPGS as a molecular biomaterial for drug delivery. Biomaterials 2012, 33, 4889–4906. [Google Scholar] [CrossRef] [PubMed]
- Neophytou, C.M.; Constantinou, A.I. Drug delivery innovations for enhancing the anticancer potential of vitamin E isoforms and their derivatives. BioMed Res Int. 2015, 2015, 584862. [Google Scholar] [CrossRef]
- Mu, L.; Feng, S.S. Vitamin E TPGS used as emulsifier in the solvent evaporation/extraction technique for fabrication of polymeric nanospheres for controlled release of paclitaxel (Taxol®). J. Control. Release 2002, 80, 129. [Google Scholar] [CrossRef]
- Guo, Y.; Luo, J.; Tan, S.; Otieno, B.O.; Zhang, Z. The application of vitamin E TPGS in drug delivery. Eur. J. Pharm. Sci. 2013, 49, 175–186. [Google Scholar] [CrossRef]
- Guan, X.; Gao, M.; Xu, H.; Zhang, C.; Liu, H.; Lv, L.; Deng, S.; Gao, D.; Tian, Y. Quercetin-loaded poly(lactic-co-glycolic acid)-D-alpha-tocopheryl polyethylene glycol 1000 succinate nanoparticles for the targeted treatment of liver cancer. Drug Deliv. 2016, 23, 3307–3318. [Google Scholar] [CrossRef]
- Yang, C.; Wu, T.; Qi, Y.; Zhang, Z. Recent advances in the application of vitamin E TPGS for drug delivery. Theranostics 2018, 8, 464–485. [Google Scholar] [CrossRef]
- Yan, A.; Bussche, A.V.D.; Kane, A.B.; Hurt, R.H. Tocopheryl polyethylene glycol succinate as a safe, antioxidant surfactant for processing carbon nanotubes and fullerenes. Carbon 2007, 24, 2463–2470. [Google Scholar] [CrossRef]
- Youk, H.J.; Lee, E.; Choi, M.K.; Lee, Y.-J.; Chung, J.H.; Kim, S.-H.; Lee, S.-H.; Lim, S.-J. Enhanced anticancer efficacy of alpha-tocopheryl succinate by conjugation with polyethylene glycol. J. Control. Release 2005, 107, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Collnot, E.M.; Baldes, C.; Wempe, M.F.; Kappl, R.; Hüttermann, J.; Hyatt, J.A.; Edgar, K.J.; Schaefer, U.F.; Lehr, C.-M. Mechanism of inhibition of P-glycoprotein mediated efflux by vitamin E TPGS: Influence of ATPase activity and membrane fluidity. Mol. Pharm. 2007, 4, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Collnot, E.M.; Baldes, C.; Schaefer, U.F.; Edgar, K.J.; Wempe, M.F.; Lehr, C.M. Vitamin E TPGS P-glycoprotein inhibition mechanism: Influence on conformational flexibility, intracellular ATP levels, and role of time and site access. Mol. Pharm. 2010, 7, 642–651. [Google Scholar] [CrossRef] [PubMed]
- Mi, Y.; Zhao, J.; Feng, S.S. Vitamin E TPGS prodrug micelles for hydrophilic drug delivery with neuroprotective effects. Int. J. Pharm. 2012, 438, 98–106. [Google Scholar] [CrossRef]
- Fan, Z.; Wu, J.; Fang, X.; Sha, X. A new function of vitamin E-TPGS in the intestinal lymphatic transport of lipophilic drugs: Enhancing the secretion of chylomicrons. Int. J. Pharm. 2013, 445, 141–147. [Google Scholar] [CrossRef]
- Zhu, H.; Chen, H.; Zeng, X.; Wang, Z.; Zhang, X.; Wu, Y.; Gao, Y.; Zhang, J.; Liu, K.; Liu, R.; et al. Co-delivery of chemotherapeutic drugs with vitamin E TPGS by porous PLGA nanoparticles for enhanced chemotherapy against multi-drug resistance. Biomaterials 2014, 35, 2391–2400. [Google Scholar] [CrossRef]
- Traber, M.G.; Thellman, C.A.; Rindler, M.J.; Rindler, M.J.; Kayden, H.J. Uptake of intact TPGS (D-α-tocopheryl polyethylene glycol 1000 succinate) a water-miscible form of vitamin E by human cells in vitro. Am. J. Clin Nutr. 1998, 48, 605–611. [Google Scholar] [CrossRef]
- Hidiroglou, N.; McDowell, L.R.; Batra, T.R. Tissue alpha-tocopherol concentrations following supplementation with various forms of vitamin E in sheep. Reprod. Nutr. Dev. 1994, 34, 273–278. [Google Scholar] [CrossRef]
- Varma, M.V.; Panchagnula, R. Enhanced oral paclitaxel absorption with vitamin E-TPGS: Effect on solubility and permeability in vitro, in situ and in vivo. Eur. J. Pharm. Sci. 2005, 25, 445–453. [Google Scholar] [CrossRef]
- Constantinides, P.P.; Han, J.; Davis, S.S. Advances in the use of tocols as drug delivery vehicles. Pharm. Res. 2006, 23, 243–255. [Google Scholar] [CrossRef]
- European Food Safety Authority. Opinion of the scientific panel on food additives, flavorings, processing aids and materials in contact with food on a request from the commission related to D-alpha-tocopheryl polyethylene glycol 1000 succinate (TPGS) in use for food for particular nutritional purposes. EFSA J. 2007, 5, 490. [Google Scholar]
- Ko, K.Y.; Ahn, D.U. An economic and simple purification procedure for the large scale production of ovotransferrin from egg white. Poult. Sci. 2008, 87, 1441–1450. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Chahal, B.; Majumder, K.; You, S.-J.; Wu, J. Identification of novel antioxidative peptides derived from a thermolytic hydrolysate of ovotransferrin by LC-MS/MS. J. Agric. Food Chem. 2010, 58, 7664–7672. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Moon, S.H.; Ahn, D.U. Antioxidant effects of ovotransferrin and its hydrolysates. Poultr. Sci. 2012, 91, 2747–2754. [Google Scholar] [CrossRef] [PubMed]
- Abeyrathne, E.D.N.S.; Lee, H.Y.; Ahn, D.U. Separation of ovotransferrin and ovomucoid from chicken egg white. Poutr. Sci. 2014, 93, 1010–1017. [Google Scholar] [CrossRef]
- Ibrahim, H.R.; Sugimoto, Y.; Aoki, T. Ovotransferrin antimicrobial peptide kills bacteria through a membrane damage mechanism. Biochim. Biophys. Acta 2000, 1523, 196–205. [Google Scholar] [CrossRef]
- Aguilera, O.; Quiros, L.M.; Fierro, J.F. Transferrins selectively cause ion eflux through bacterial and artificial membranes. FEBS Lett. 2003, 548, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Hirota, Y.; Yang, M.P.; Araki, S.; Yoshihara, K. Enhancing effects of chicken egg white derivatives on the phagocytic response in the dog. J. Vet. Med. Sci. 1995, 57, 825–829. [Google Scholar] [CrossRef] [Green Version]
- Otani, H.; Odashima, M. Inhibition of proliferative responses of mouse spleen lymphocytes by lacto- and ovotransferrins. Food Agric. Immunol. 1997, 9, 193–202. [Google Scholar] [CrossRef]
- Ibrahim, H.R.; Hoq, M.I.; Aoki, T. Ovotransferrin possess SOD-like superoxide anion scavenging activity that is promoted by copper and manganese binding. Int. J. Biol. Macromol. 2007, 41, 631–640. [Google Scholar] [CrossRef]
- Huang, W.Y.; Majumder, K.; Wu, J. Oxygen radical absorbance capacity of peptides from egg white protein ovotransferrin and their interaction with phytochemicals. Food Chem. 2010, 123, 635–641. [Google Scholar] [CrossRef]
- Meng, X.; Liu, J.; Yu, X.; Li, J.; Lu, X.; Shen, T. Pluronic F127 and D-α-tocopheryl polyethylene glycol succinate (TPGS) mixed micelles for targeting drug delivery across the blood-brain barrier. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Massey, J.B. Kinetics of transfer of α-tocopherol between model and native plasma lipoproteins. Biochim. Biophys. Acta 1984, 793, 387–392. [Google Scholar] [CrossRef]
- Drevon, C.A. Absorption, transport and metabolism of vitamin E. Free Radic. Res. Commun. 1991, 14, 229–246. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, I.; Imasato, Y.; Sasaki, E. Lymphatic transport of α, γ, and δ-tocotrienols and α-tocopherol in rats. Int. J. Vit. Nutr. Res. 1996, 66, 217–221. [Google Scholar]
- Yap, S.P.; Yuen, K.H.; Wong, J.W. Pharmacokinetics and bioavailability of alpha-, gamma-, and delta-tocotrienols under different food status. J. Pharm. Pharmacol. 2001, 53, 67–71. [Google Scholar] [CrossRef]
- Yap, S.P.; Yuen, K.H.; Lim, A.B. Influence of route of administration on the absorption and disposition of alpha-, gamma-, and delta-tocotrienols in rats. J. Pharm. Pharmacol. 2003, 55, 53–58. [Google Scholar] [CrossRef]
- Qian, J.; Wilson, K.; Nava, P.; Nava, P.; Morley, S.; Atkinson, J.; Manor, D. Intracellular localization of α-tocopherol transfer protein and α-tocopherol. Ann. N. Y. Acad. Sci. 2004, 1031, 330–331. [Google Scholar] [CrossRef]
- Qian, J.; Morley, S.; Wilson, K.; Nava, P. Intracellular trafficking of vitamin E in hepatocytes: The role of tocopherol transfer protein. J. Lipid Res. 2005, 46, 2072–2082. [Google Scholar] [CrossRef] [Green Version]
- Abuasal, B.S.; Qosa, H.; Sylvester, P.W.; Kaddoumi, A. Comparison of the intestinal absorption and bioavailability of gamma-tocotrienol and alpha-tocopherol: In vitro, in situ and in vivo studies. Biopharm. Drug Dispos. 2012, 33, 246–256. [Google Scholar] [CrossRef]
- Khanna, S.; Patel, V.; Rink, C.; Roy, S.; Sen, C.K. Delivery of orally supplemented α-tocotrienol to vital organs of rats and tocopherol-transport protein deficient mice. Free Radic. Biol. Med. 2005, 39, 1310–1319. [Google Scholar] [CrossRef] [Green Version]
- Dintaman, J.M.; Silverman, J.A. Inhibition of P-glycoprotein by D-α-tocopheryl polyethylene glycol 1000 succinate (TPGS). Pharm. Res. 1999, 16, 1550–1556. [Google Scholar] [CrossRef] [PubMed]
- Robin, Y. Using tocophersolan for drug delivery. Pharm. Technol. 2015, 39. Available online: https://www.pixelmags.com/awrv1/#magazines/1367/issues/110846/pages/51 (accessed on 25 November 2019).
- Akhtar, N.; Ahad, A.; Khar, R.K.; Jaggi, M.; Aqil, M.; Iqbal, Z. The emerging role of P-glycoprotein inhibitors in drug delivery: A patent review. Expert Opin. Ther. Pat. 2011, 21, 561–576. [Google Scholar] [CrossRef] [PubMed]
- Loeb, J.M.; Faith, H.G.; Czuprynski, C.J. Intestinal P glycoprotein acts as a natural defense mechanism against Listeria monocytogenes. Infect. Immun. 2004, 72, 3849–3854. [Google Scholar]
- Endicott, J.A.; Ling, V. The biochemistry of P-glycoprotein-mediated multidrug resistance. Annu. Rev. Biochem. 1989, 58, 137–171. [Google Scholar] [CrossRef]
- Pan, G.Y.; Wang, G.J.; Liu, X.D.; Fawcett, J.P.; Xie, Y.-Y. The involvement of P-glycoprotein in berberine absorption. Pharmacol. Toxicol. 2002, 91, 193–197. [Google Scholar] [CrossRef]
- Chang, T.; Benet, L.Z.; Hebert, M.F. The effect of water-soluble vitamin E on cyclosporine pharmacokinetics in healthy volunteers. Clin. Pharmacol. Ther. 1996, 59, 297–303. [Google Scholar] [CrossRef]
- Yu, L.; Bridgers, A.; Polli, J.; Vickers, A. Vitamin E-TPGS increases absorption flux of an HIV protease inhibitor by enhancing its solubility and permeability. Pharm. Res. 1999, 16, 1812–1817. [Google Scholar] [CrossRef]
- Leu, B.L.; Huang, J.D. Inhibition of intestinal P-glycoprotein and effects on etoposide absorption. Cancer Chemother. Pharmacol. 1995, 35, 432–436. [Google Scholar] [CrossRef]
- Fischer, J.R.; Harkin, K.R.; Freeman, L.C. Concurrent administration of water-soluble vitamin E can increase the oral bioavailability of cyclosporine A in healthy dogs. Vet. Ther. Res. Appl. Vet. Med. 2002, 3, 465–473. [Google Scholar]
- Prasad, Y.V.; Puthli, S.P.; Eaimtrakarn, S.; Ishida, M.; Yoshikawa, Y.; Shibata, N.; Takada, K. Enhanced intestinal absorption of vancomycin with Labrasol and D-alpha-tocopheryl PEG 1000 succinate in rats. Int. J. Pharm. 2003, 250, 181–190. [Google Scholar] [CrossRef]
- Ke, W.T.; Lin, S.Y.; Ho, H.-O.; Sheu, M.-T. Physical characterization of microemulsion systems using tocopheryl polyethylene glycol 1000 succinate (TPGS) as a surfactant for the oral delivery of protein drugs. J. Control. Release 2005, 102, 489–507. [Google Scholar] [CrossRef] [PubMed]
- Argao, E.A.; Heubi, J.E.; Hollis, B.W.; Tsang, R.C. D-alpha-tocopheryl glycol-1000 succinate enhances the absorption of vitamin D in chronic cholestatic liver disease of infancy and childhood. Pediatr. Res. 1992, 31, 146–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bittner, B.; Guenzi, A.; Fullhardt, P.; Zuercher, G. Improvement of the bioavailability of colchicine in rats by co-administration of D-alpha-tocopherol polyethylene glycol 1000 succinate and a polyethoxylated derivative of 12-hydroxy-stearic acid. Arzneimittel-Forschung 2002, 52, 684–688. [Google Scholar] [PubMed]
- Gao, Y.; Li, Z.; Sun, M.; Li, H. Preparation, characterization, pharmacokinetics, and tissue distribution of curcumin nanosuspesion with TPGS as stabilizer. Drug Dev. Ind. Pharm. 2010, 36, 1225–1234. [Google Scholar] [CrossRef]
- Traber, M.G.; Atkinson, J. Vitamin E, antioxidant and nothing more. Free Radic. Biol. Med. 2007, 43, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q. Natural forms of vitamin E: Metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy. Free Radic. Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef] [Green Version]
- Khanna, S.; Roy, S.; Slivka, A.; Craft, T.K.S.; Chaki, S.; Rink, C.; Notestine, M.A.; DeVries, A.C.; Parinandi, N.L.; Sen, C.K. Neuroprotective properties of the natural vitamin E α-tocotrienol. Stroke 2005, 36, 2258–2264. [Google Scholar] [CrossRef]
- Constantinou, C.; Papas, A.; Constantinou, A.I. Vitamin E and cancer: An insight into the anticancer activities of vitamin E isomers and analogs. Int. J. Cancer 2008, 123, 739–752. [Google Scholar] [CrossRef]
- Marubayashi, S.; Dohi, K.; Ochi, K.; Kawasaki, T. Role of free radicals in ischemic rat liver cell injury: Prevention of damage by α-tocopherol administration. Surgery 1986, 99, 184–191. [Google Scholar]
- Bjorneboe, A.; Bjorneboe, G.E.A.; Nenseter, M.S.; Hagen, B.F. Effect of dietary deficiency and supplement with all-rac-alpha-tocopherol on hepatic content in rats. J. Nutr. 1991, 121, 1208–1213. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.F.; Jameson, J.A.; Tilly, D.; Prochazka, L.; Rohlena, J.; Valis, K.; Truska, J.; Zobalova, R.; Mahadavian, E.; Kluchova, K.; et al. Mitochondrial targeting of alpha-tocopheryl succinate enhances its pro-apoptotic efficacy: A new paradigm for effective cancer therapy. Free Radic. Biol. Med. 2011, 50, 1546–1555. [Google Scholar] [CrossRef] [PubMed]
- Chow, C.K. Vitamin E and oxidative stress. Free Radic. Biol. Med. 1991, 11, 232–251. [Google Scholar]
- Van Acker, S.A.B.E.; Koymans, L.M.H.; Bast, A. Molecular pharmacology of vitamin E: Structural aspects of antioxidant activity. Free Radic. Biol. Med. 1993, 15, 311–328. [Google Scholar] [CrossRef]
- Chow, C.K.; Ibrahim, W.; Wei, Z.; Chan, A.C. Vitamin E regulates mitochondrial hydrogen peroxide generation. Free Radic. Biol. Med. 1999, 27, 580–587. [Google Scholar] [CrossRef]
- Kotegawa, M.; Sugiyama, M.; Shoji, T. Effect of α-tocopherol on high energy phosphate metabolite levels in rat heart by 31P-NMR using a Langendorff perfusion technique. J. Mol. Cell. Cardiol. 1993, 25, 1067–1074. [Google Scholar] [CrossRef]
- Punz, A.; Nanobashvili, J.; Fuegl, A.; Roth, E. Effect of α-tocopherol pretreatment on high energy metabolites in rabbit skeletal muscle after ischemia-reperfusion. Clin. Nutr. 1998, 17, 85–87. [Google Scholar] [CrossRef]
- Hama, S.; Utsumi, S.; Fukuda, Y.; Nakayama, K.; Okamura, Y.; Tsuchiya, H.; Fukuzawa, K.; Harashima, H.; Kogure, K. Development of a novel drug delivery system consisting of an antitumor agent tocopheryl succinate. J. Control. Release 2012, 161, 843–851. [Google Scholar] [CrossRef]
- Kovarova, J.; Bajkikova, M.; Vondrusova, M.; Stursa, J. Mitochondrial targeting of α-tocopheryl succinate enhances its anti-mesothelioma efficacy. Redox Rep. 2014, 19, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Truska, J.; Dong, L.F.; Rohlena, J.; Stursa, J.; Vondrusova, M.; Goodwin, J.; Nguyen, M.; Kluckova, K.; Rychtarcikova, K.; Lettlova, S.; et al. Mitochondrially targeted vitamin E succinate modulates expression of mitochondrial DNA transcripts and mitochondrial biogenesis. Antioxid. Redox Signal. 2015, 22, 883–900. [Google Scholar]
- Hensley, K.; Benaksas, E.; Bolli, R.; Comp, P.; Grammas, P.; Hamdheydari, L.; Mou, S.; Pye, Q.N.; Stoddard, M.F.; Wallis, G.; et al. New perspectives on vitamin E: γ-tocopherol and carboxyethylhydroxychroman metabolites in biology and medicine. Free Radic. Biol. Med. 2004, 36, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zingg, J.-M.; Azzi, A. Non-antioxidant activities of vitamin E. Curr. Med. Chem. 2004, 11, 1113–1133. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Luo, Y.; Wu, J. Conjugation of ovotransferrin with catechin shows improved antioxidant activity. J. Agric. Food Chem. 2014, 62, 2581–2587. [Google Scholar] [CrossRef] [PubMed]
- Sancak, A.A.; Rutgers, H.C.; Hart, C.A.; Batt, R.M. Prevalence of enteropathic Escherichia coli in dogs with acute and chronic diarrhea. Vet. Rec. 2004, 154, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Seepersadsingh, N.; Adesiyun, A.A.; Seebaransingh, R. Prevalence and antimicrobial resistance of Salmonella Spp. in non-diarrheic dogs in Trinidad. J. Vet. Med. B Infect. Dis. Vet. Publ. Health. 2004, 51, 337–342. [Google Scholar] [CrossRef]
- Honneffer, J.B.; Minamoto, Y.; Suchodolski, J.S. Microbiota alterations in acute and chronic gastrointestinal inflammation of cats and dogs. World J. Gastroenterol. 2014, 20, 16489–16497. [Google Scholar] [CrossRef]
- Halfvarson, J. Dynamics of the human gut microbiome in inflammatory bowel disease. Nat. Microbiol. 2017, 13. [Google Scholar] [CrossRef] [Green Version]
- Eissa, N.; Kittana, H.; Gomes-Neto, J.C.; Hussein, H. Mucosal immunity and gut microbiota in dogs with chronic enteropathy. Res. Vet. Sci. 2019, 122, 156–164. [Google Scholar] [CrossRef]
- Noverr, R.P.; Huffnagle, G.B. Does the microbiota regulate immune responses outside the gut? Trends Microbiol. 2004, 12, 562–568. [Google Scholar] [CrossRef]
- Boulangé, C.L.; Neves, A.L.; Chilloux, J.; Nicholson, J.K.; Dumas, M.-E. Impact of gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. 2016, 8, 42. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Pitmon, E.; Wang, K. Microbiome, inflammation, and colorectal cancer. Semin. Immunol. 2017, 32, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari, Z.; Gibson, D.; Hekmatdoost, A. Nonalcoholic fatty liver disease, the gut microbiome, and diet. Adv. Nutr. 2017, 8, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.P.; Chang, S.H.; Tang, C.Y.; Liou, M.-L.; Tsai, S.-J.J.; Lin, Y.-L. Composition analysis and feature selection of the oral microbiota associated with periodontal disease. BioMed Res. Int. 2018, 2018, 3130607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, P.; Messina, N.; Mohn, W.W.; Finlay, B.B.; Curtis, N. Association between the intestinal microbiota and allergic sensitization, eczema, and asthma: A systematic review. J. Allergy Clin. Immunol. 2018, 143, 467–485. [Google Scholar] [CrossRef] [PubMed]
- Neish, A.S.; Denning, T.L. Advances in understanding the interaction between the gut microbiota and adaptive mucosal immune responses. F1000 Biol. Rep. 2010, 2, 27. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Littman, D.R.; Macpherson, A.J. Interactions between the microbiota and the immune system. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprouse, M.L.; Bates, N.A.; Felix, K.M.; Wu, H.J. Impact of gut microbiota on gut-distal autoimmunity: A focus on T cells. Immunology 2018, 156, 305–318. [Google Scholar] [CrossRef] [Green Version]
- Nieuwenhuizen, G.A.P.; Goris, R.J.A. The gut: The ‘motor’ of multiple organ dysfunction syndrome? Curr. Opin. Clin. Nutr. Metab. Care 1999, 2, 399–404. [Google Scholar] [CrossRef]
- Talukder, J.R.; Griffin, A.; Jaima, A. Lactoferrin ameliorates prostaglandin E2-mediated inhibition of Na+ -glucose cotransport in enterocytes. Can. J. Physiol. Pharmacol. 2014, 92, 9–20. [Google Scholar] [CrossRef]
- Valenti, P.; Antonini, G.; Fanelli, M.R.; Orsi, N.; Antonini, E. Antibacterial activity of matrix-bound ovotransferrin. Antimicrob. Agents Chemother. 1982, 21, 840–841. [Google Scholar] [CrossRef] [Green Version]
- Farnaud, S.; Evans, R.W. Lactoferrin-A multifunctional protein with antimicrobial properties. Mol. Immunol. 2003, 40, 395–405. [Google Scholar] [CrossRef]
- Valenti, P.; Antonini, G. Lactoferrin: An important host defense against microbial and viral attack. Cell. Mol. Life Sci. 2005, 62, 2576–2587. [Google Scholar] [CrossRef] [PubMed]
- Ammendolia, M.G.; Pietrantoni, A.; Tinari, P.; Valenti, P.; Superti, F. Bovine lactoferrin inhibits echovirus endocytic pathway by interacting with viral structural polypeptides. Antivir. Res. 2007, 73, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Awano, N.; Fukui, T.; Sasaki, H.; Kyuwa, S. The protective effects of lactoferrin against murine norovirus infection through inhibition of both viral attachment and replication. Biochem. Biophys. Res. Commun. 2013, 434, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Talukder, J.; Srivastava, A.; Ray, A.; Lall, R. Treatment of Infectious Endometritis with a Novel Protein; VPI-O22, in Cows; Available online: https://www.fasebj.org/doi/10.1096/fasebj.2018.32.1_supplement.882.12 (accessed on 25 November 2019).
- Talukder, J. Egg derived ovotransferrin and lactoferrin. In Nutraceuticals in Veterinary Medicine; Gupta, R.C., Srivastava, A., Lall, R., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 235–243. [Google Scholar]
- Talukder, J. Nutraceuticals in gastrointestinal conditions. In Nutraceuticals in Veterinary Medicine; Gupta, R.C., Srivastava, A., Lall, R., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 467–479. [Google Scholar]
- Connely, O.M. Anti-inflammatory activities of lactoferrin. J. Am. Coll. Nutr. 2001, 438, 389S–395S. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.R.; Kiyono, T. Novel anticancer activity of the autocleaved ovotransferrin against human colon and breast cancer cells. J. Agric. Food Chem. 2009, 57, 11383–11390. [Google Scholar] [CrossRef]
- Elli, M.; Zink, R.; Rytz, A.; Reniero, R. Iron requirement of Lactobacillus spp. in completely chemically defined growth media. J. Appl. Microbiol. 2000, 88, 695–703. [Google Scholar] [CrossRef]
- Imbert, M.; Blondeau, R. On the iron requirement of Lactobacilli grown in chemically defined medium. Curr. Microbiol. 1998, 37, 64–66. [Google Scholar] [CrossRef]
- Archibald, F. Lactobacillus plantarum, an organism not requiring iron. FEMS Microbiol. Lett. 1983, 19, 29–32. [Google Scholar] [CrossRef]
- Yilmaz, B.; Li, H. Gut microbiota and iron: The crucial actors in health and disease. Pharmaceuticals 2018, 11, 98. [Google Scholar] [CrossRef] [Green Version]
- Scott, K.P.; Antoine, J.-M.; Midtvedt, T.; Hemert, S.V. Manipulating the gut microbiota to maintain health and treat disease. Microb. Ecol. Health Dis. 2015, 26, 25877. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Hofstaedter, C.E.; Zhao, C.; Mattei, L.; Tanes, C.; Clarke, E.; Lauder, A.; Sherrill-Mix, S.; Chehoud, C.; Kelsen, C.; et al. Optimizing methods and dodging pitfalls in microbiome research. Microbiome 2017, 5, 52. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-J.; Li, S.; Gan, R.Y.; Zhou, T.; Xu, D.-P.; Li, H.-B. Impacts of gut bacteria on human health and diseases. Int. J. Mol. Sci. 2015, 16, 7493–7519. [Google Scholar] [CrossRef] [PubMed]
- Kortman, G.A.M.; Raffatellu, M.; Swinkels, D.W.; Tjalsma, H. Nutritional iron turned inside out: Intestinal stress from a gut microbial perspective. FEMS Microbiol. Rev. 2014, 38, 1202–1234. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Jiang, Y.; Yao, Y.; Wu, N.; Luo, J.; Hu, M.; Tu, Y.; Xu, M. Ovotransferrin ameliorates the dysbiosis of immunomodulatory-function and intestinal microbiota induced by cyclophosphamide. Food Funct. 2019, 10, 1109–1122. [Google Scholar] [CrossRef]
- Anadon, A.; Martinez-Larranaga, M.R.; Ares, I.; Martinez, M.A. Prebiotics: Safety and toxicity considerations. In Nutraceuticals: Efficacy, Safety and Toxicity; Gupta, R.C., Ed.; Academic Press/Elsevier: Amsterdam, The Netherlands, 2016; pp. 757–775. [Google Scholar]
- Anadon, A.; Martinez-Larranaga, M.R.; Ares, I.; Martinez, M.A. Probiotics: Safety and toxicity considerations. In Nutraceuticals: Efficacy, Safety and Toxicity; Gupta, R.C., Ed.; Academic Press/Elsevier: Amsterdam, The Netherlands, 2016; pp. 777–798. [Google Scholar]
- Anadon, A.; Ares, I.; Martinez-Larranaga, M.R.; Martinez, M.A. Prebiotics and probiotics in feed and animal health. In Nutraceuticals in Veterinary Medicine; Gupta, R.C., Srivastava, A., Lall, R., Eds.; Springer: Cham, Switzerland, 2019; pp. 261–286. [Google Scholar]
- Banerjee, A.; Giri, R. Nutraceuticals in gastrointestinal disorders. In Nutraceuticals: Efficacy, Safety and Toxicity; Gupta, R.C., Ed.; Academic Press/Elsevier: Amsterdam, The Netherlands, 2016; pp. 109–122. [Google Scholar]
- Malik, J.K.; Ahmad, A.H.; Kalpana, S.; Prakash, A.; Gupta, R.C. Synbiotics: Safety and toxicity considerations. In Nutraceuticals: Efficacy, Safety and Toxicity; Gupta, R.C., Ed.; Academic Press/Elsevier: Amsterdam, The Netherlands, 2016; pp. 811–822. [Google Scholar]
- Most, J.; Penders, J.; Lucchesi, M.; Goossens, G.H.; Blaak, E.E. Gut microbiota composition in relation to the metabolic response to 12-week combined polyphenol supplementation in overnight men and women. Eur. J. Clin. Nutr. 2017, 71, 1040–1045. [Google Scholar] [CrossRef]
- Feng, W.; Ao, H.; Peng, C. Gut microbiota, short-chain fatty acids, and herbal medicine. Front. Pharmacol. 2018, 9, 1354. [Google Scholar] [CrossRef]
- Guo, Y.; Xie, J.; Li, X.; Yuan, Y.; Zhang, L.; Hu, W.; Luo, H.; Yu, H.; Zhang, R. Antidepressant effects of Rosemary extracts associated with anti-inflammatory effect and rebalance of gut microbiota. Front. Pharmacol. 2018, 9, 1126. [Google Scholar] [CrossRef] [Green Version]
- Santangelo, R.; Silvestrini, A.; Mancuso, C. Ginsenosides, catechins, quercetin and gut microbiota: Current evidence of challenging interactions. Food Chem. Toxicol. 2019, 123, 42–49. [Google Scholar] [CrossRef]
- Lederberg, J. Infectious history. Science 2000, 288, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Redfern, A.; Suchodolski, J.; Jergens, A. Role of the gastrointestinal microbiota in small animal health and disease. Vet. Rec. 2017, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.; Weese, J.S. Understanding the intestinal microbiome in health and disease. Vet. Clin. N. Am. Equine Pract. 2018, 34, 1012. [Google Scholar] [CrossRef] [PubMed]
- Antza, C.; Stabouli, S.; Kotsis, V. Gut microbiota in kidney disease and hypertension. Pharmacol. Res. 2018, 130, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2000, 90, 859–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cisek, A.A. Chicken intestinal microbiota function with a special emphasis on the role of probiotic bacteria. Pol. J. Vet. Sci. 2014, 17, 385–394. [Google Scholar] [CrossRef] [Green Version]
- Freter, R. Factors affecting the microecology of the gut. In Probiotics; Springer: Dordrecht, The Netherlands, 1992; pp. 111–144. [Google Scholar] [CrossRef]
- Hildebrand, F.; Nguyen, T.L.; Brinkman, B.; Yunta, R.G.; Cauwe, B.; Vandenabeele, P.; Liston, A.; Raes, J. Inflammation-associated enterotypes, host genotype, cage and inter-individual effects drive gut microbiota variation in common laboratory mice. Genome Biol. 2013, 14, R4. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.C.; Silva, G.; Ramos, R.V.; Staempfli, H.R.; Arroyo, L.G.; Kim, P.; Weese, J.S. Characterization and comparison of the bacterial microbiota in different gastrointestinal tract compartments in horses. Vet. J. 2015, 205, 74–80. [Google Scholar] [CrossRef]
- Franklin, C.L.; Ericsson, A.C. Microbiota and reproducibility of rodent models. Lab. Anim. 2017, 46, 114–122. [Google Scholar] [CrossRef]
- Tappenden, K.A.; Deutsch, A.S. The physiological relevance of the intestinal microbiota-contributions to human health. J. Am. Coll. Nutr. 2007, 26, 679S–683S. [Google Scholar] [CrossRef]
- Neish, A.S. Microbes in gastrointestinal health and disease. Gastroenterology 2009, 136, 65–80. [Google Scholar] [CrossRef] [Green Version]
- O’Keefe, S.J. Nutrition and colonic health: The critical role of the microbiota. Curr. Opin. Gastroenterol. 2008, 24, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Galland, L. The gut microbiome and the brain. J. Med. Food. 2014, 17, 1261–1272. [Google Scholar] [CrossRef] [PubMed]
- Tremlett, H.; Bauer, K.C.; Appel-Cresswell, S.; Finlay, B.B.; Waubant, E. The gut microbiome in human neurological disease: A review. Ann. Neurol. 2017, 82, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Manson, J.M.; Rauch, M.; Gilmore, M.S. The commensal microbiology of the gastrointestinal tract. Adv. Exp. Med. Biol. 2008, 635, 15–28. [Google Scholar]
- Yu, L.C.H.; Wang, J.-T.; Wei, S.-C.; Ni, Y.-H. Host-microbial interactions and regulation of intestinal epithelial barrier function: From physiology to pathology. World J. Gastrointest. Pathophysiol. 2012, 3, 27–43. [Google Scholar] [CrossRef]
- Kamadan, N.; Seo, S.U.; Chen, G.Y.; Nunez, G. Role of the gut microbiota in immunity and inflammatory disease. Nat. Rev. Immunol. 2013, 13, 321–335. [Google Scholar] [CrossRef]
- Holmes, E.; Li, J.V.; Athanasiou, T.; Ashrafian, H.; Nicholson, J.K. Understanding the role of gut microbiome-host metabolic signal disruption in health and disease. Trends Microbiol. 2011, 19, 349–359. [Google Scholar] [CrossRef]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.A.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; et al. The gut microbiota and host health: A new clinical frontier. Gut 2016, 65, 330–339. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Verbrugghe, A.; Lourenco, M.; Janssens, G.P.J.; Liu, D.J.K.; de Wiele, T.V.; Eeckhaut, V.; Immerseel, F.V.; de Maele, I.V.; Niu, Y.; et al. Does canine inflammatory bowel disease influence gut microbial profile and host metabolism. BMC Vet. Res. 2016, 12, 114. [Google Scholar] [CrossRef] [Green Version]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Craven, M.; Egan, C.E.; Dowd, S.E.; McDonough, S.P.; Dogan, B.; Denkers, E.Y.; Bowman, D. Inflammation drives dysbiosis and bacterial invasion in murine models of ileal Crohn’s disease. PLoS ONE 2012, 7, e41594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guard, B.C.; Barr, J.W.; Reddivari, L.; Klemashevich, C.; Jayaraman, A.; Steiner, J.M.; Vanamala, J.; Suchodolski, J.S. Characterization of microbial dysbiosis and metabolomic changes in dogs with acute diarrhea. PLoS ONE 2015, 10, e0127259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez-Baeza, Y.H.; Suchodolski, E.R.; Knight, R.J.S. Dogs and human inflammatory bowel disease rely on overlapping yet distinct dysbiosis networks. Nat. Microbiol. 2016, 1, 16177. [Google Scholar] [CrossRef] [PubMed]
- Clarke, G.; Sandhu, K.V.; Griffin, B.T.; Dinan, T.G.; Cryan, J.F.; Hyland, N.P. Gut reactions: Breaking down xenobiotics-microbiome interactions. Pharmacol. Rev. 2019, 71, 198–224. [Google Scholar] [CrossRef]
- De Meyer, F.; Eeckhaut, V.; Ducatelle, R.; Dhaenens, M.; Daled, S.; Dedeurwaerder, A.; De Gussem, M.; Haesebrouck, F.; Deforce, D.; Immerseel, F.V. Host intestinal biomarker identification in a gut leakage model in broilers. Vet. Res. 2019, 50, 46. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, J.K.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; Pettersson, S. Host-gut microbiota metabolic interactions. Science 2012, 336, 1262–1267. [Google Scholar] [CrossRef] [Green Version]
- De Vadderr, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Backhed, F.; Mithieux, G. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.S.; Yu, J.S.; Yoo, H.H.; Kim, D.H. The role of gut microbiota in the pharmacokinetics of antihypertensive drugs. Pharmacol. Res. 2018, 130, 164–171. [Google Scholar] [CrossRef]
- De-Paula, V.D.R.; Forlenza, A.S.; Forlenza, O.V. Relevance of gut microbiota in cognition, behaviour and Alzheimer’s disease. Pharmacol. Res. 2018, 136, 29–34. [Google Scholar] [CrossRef]
- Gokulan, K.; Arnold, M.G.; Jensen, J.; Vanlandingham, M.; Twaddle, N.C.; Doerge, D.R. Exposure to arsenite in CD-1 mice during juvenile and adult stages: Effects on intestinal microbiota and gut-associated immune status. MBio 2018, 14, e01418-18. [Google Scholar] [CrossRef] [Green Version]
- Strandwitz, P. Neurotransmitter modulation by the gut microbiota. Brain Res. Part B 2018, 1693, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Sircana, A.; De Michieli, F.; Parente, R.; Framarin, L.; Leone, N.; Berrutti, M.; Paschetta, E.; Bongiovanni, D.; Musso, G. Gut microbiota, hypertension and chronic kidney disease: Recent advances. Pharmacol. Res. 2019, 144, 390–408. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.F. Microbial ecology: Human gut microbes associated with obesity. Nature 2016, 444, 1022–1023. [Google Scholar] [CrossRef]
- Goulet, O. Potential role of the intestinal microbiota in programming health and disease. Nutr. Rev. 2015, 73 (Suppl. 1), 32–40. [Google Scholar] [CrossRef]
- Nieuwenhuizen, G.A.; Deitch, E.A.; Goris, R.J. Infection, the gut and the development of the multiple organ dysfunction syndrome. Eur. J. Surg. 1996, 162, 259–273. [Google Scholar]
- Leaphart, C.L.; Tepas, J.J. The gut is a motor of organ system dysfunction. Surgery 2007, 141, 563–569. [Google Scholar] [CrossRef]
- Rakoff-Nahoum, S.; Paglino, J.; Eslami-Varzaneh, F.; Edberg, S.; Medzhitov, R. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell 2004, 118, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Shirkey, T.W.; Siggers, R.H.; Goldade, B.G.; Marshall, J. Effects of commensal bacteria on intestinal morphology and expression of proinflammatory cytokines in the gnotobiotic pigs. Exp. Biol. Med. 2006, 231, 1333–1345. [Google Scholar] [CrossRef]
- Giansanti, F.; Panella, G.; Leboffe, L.; Antonini, G. Lactoferrin from Milk: Nutraceutical and Pharmacological Properties. Pharmaceuticals 2016, 9, 61. [Google Scholar] [CrossRef] [Green Version]
- Valenti, P.; Antonini, G.; Von Hunostein, C.; Visca, P.; Antonini, E. Studies of the antimicrobial activity of ovotransferrin. Int. J. Tissue React. 1983, 1, 97–105. [Google Scholar]
- Talukder, M.J.R.; Harada, E. Bovine lactoferrin protects lipopolysaccharide-induced diarrhea modulating nitric oxide and prostaglandin E2 in mice. Can. J. Physiol. Pharmacol. 2007, 85, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Madara, J.L. Maintenance of the macromolecular barrier at cell extrusion sites in intestinal epithelium: Physiological rearrangement of tight junctions. J. Membr. Biol. 1990, 116, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Paesold, G.; Guiney, D.G.; Eckmann, L.; Kagnoff, M.F. Genes in the Salmonella virulence plasmid are essential for Salmonella-induced apoptosis in intestinal epithelial cells. Cell Microbiol. 2002, 4, 771–781. [Google Scholar] [CrossRef]
- Flynn, A.N.; Buret, A.G. Tight junctional disruption and apoptosis in an in vitro model of Citrobactor rodentium infection. Microb. Pathol. 2008, 45, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, J.K.; Huang, C.Y.; Lu, Y.Z.; Yang, C.Y. Magnetic resonance imaging detects intestinal barrier dysfunction in a rat model of acute mesenteric ischemia/reperfusion injury. Investig. Radiol. 2009, 44, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Redlak, M.J.; Power, J.J.; Miller, T.A. Prevention of deoxycholate-induced gastric apoptosis by aspirin: Roles of NF-kappaB and PKC signaling. J. Surg. Res. 2002, 145, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Omatsu, T.; Naito, Y.; Handa, O.; Hayashi, N.; Mizushima, K.; Qin, Y.; Hirata, I.; Adachi, S.; Okayama, T.; Kishimoto, E.; et al. Involvement of reactive oxygen species in indomethacin-induced apoptosis of small intestinal epithelial cells. J. Gastroenterol. 2009, 44 (Suppl. 19), 30–34. [Google Scholar] [CrossRef] [PubMed]
- Kamada, N.; Chen, G.Y.; Inohara, N.; Nunez, G. Control of pathogens and pathobionts by the gut microbiota. Nat. Immunol. 2013, 14, 685–690. [Google Scholar] [CrossRef]
- Ghattamaneni, N.K.R.; Panchal, S.K.; Brown, L. Nutraceuticals in rodent models as potential treatments for human inflammatory bowel disease. Pharmacol. Res. 2018, 132, 99–107. [Google Scholar] [CrossRef]
- Kelly, D.; Conway, S. Bacterial modulation of mucosal innate immunity. Mol. Immunol. 2005, 42, 895–901. [Google Scholar] [CrossRef]
- Neish, A.S. Mucosal immunity and the microbiome. Ann. Am. Thorac. Soc. 2014, 11, S28–S32. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.; Thaiss, C.A.; Elinav, E. Metabolites: Messengers between the microbiota and the immune system. Genes Dev. 2016, 30, 1589–1597. [Google Scholar] [CrossRef] [PubMed]
- Mazmanian, S.K.; Liu, C.H.; Tzianabos, A.O.; Kasper, D.L. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masnoon, N.; Shakib, S.; Kalisch-Ellett, L.; Caughey, G.E. What is polypharmacy? A systematic review of definitions. BMC Geriatr. 2017, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, A.S.; Zhang, S. Polypharmacology: Drug discovery for the future. Expert Rev. Clin. Pharmacol. 2013, 6, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dormán, G.; Flachner, B.; Hajdú, I.; András, C.D. Target identification and polypharmacology of nutraceuticals. In Nutraceuticals, Efficacy, Safety and Toxicity; Gupta, R.C., Ed.; Academic Press: Amsterdam, The Netherlands; Elsevier: Amsterdam, The Netherlands, 2016; pp. 263–286. [Google Scholar]
- Pitkälä, K.H.; Suominen, M.H.; Bell, J.S.; Strandberg, T.E. Herbal medications and other dietary supplements. A critical review for physicians caring for older people. Ann. Med. 2016, 48, 586–602. [Google Scholar] [CrossRef]
- Brown, A.C. An overview of herb and dietary supplement efficacy, safety and government regulations in the United States with suggested improvements. Part 1 of 5 series. Food Chem. Toxicol. 2017, 107, 449–471. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structural formula of ITPGS (IT.TPGS).

Figure 2.
Lactoferrin restoration of glucose absorption.

Figure 3.
Antibacterial activity of ovotransferrin (OvTf) against E. coli strain W1485 (5 × 105 CFU/mL) of different concentrations of OvTf. Ordinate, colony-forming units per milliliter; abscissa, concentrations of OvTf (courtesy of Valenti et al., 1982 [111]). Antibacterial activity increased with concentrations of OvTf.
Figure 3.
Antibacterial activity of ovotransferrin (OvTf) against E. coli strain W1485 (5 × 105 CFU/mL) of different concentrations of OvTf. Ordinate, colony-forming units per milliliter; abscissa, concentrations of OvTf (courtesy of Valenti et al., 1982 [111]). Antibacterial activity increased with concentrations of OvTf.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Srivastava, A.; Lall, R.; Talukder, J.; DuBourdieu, D.; Gupta, R.C. Iron Transport Tocopheryl Polyethylene Glycol Succinate in Animal Health and Diseases. Molecules 2019, 24, 4289. https://doi.org/10.3390/molecules24234289
AMA Style
Srivastava A, Lall R, Talukder J, DuBourdieu D, Gupta RC. Iron Transport Tocopheryl Polyethylene Glycol Succinate in Animal Health and Diseases. Molecules. 2019; 24(23):4289. https://doi.org/10.3390/molecules24234289
Chicago/Turabian StyleSrivastava, Ajay, Rajiv Lall, Jamil Talukder, Dan DuBourdieu, and Ramesh C. Gupta. 2019. "Iron Transport Tocopheryl Polyethylene Glycol Succinate in Animal Health and Diseases" Molecules 24, no. 23: 4289. https://doi.org/10.3390/molecules24234289