Natural Occurrence, Bioactivity and Biosynthesis of Elaiophylin Analogues


1
State Key Laboratory of Dairy Biotechnology, Technology Center and Dairy Research Institute of Bright Dairy & Food Co. Ltd., 1518 West Jiangchang Road, Shanghai 200436, China
2
School of Pharmacy, Second Military Medical University, 325 Guohe Road, Shanghai 200433, China
3
College of Food Science and Technology, Shanghai Ocean University, 999 Huchenghuan Road, Shanghai 201306, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Molecules 2019, 24(21), 3840; https://doi.org/10.3390/molecules24213840
Submission received: 30 September 2019
/
Revised: 21 October 2019
/
Accepted: 22 October 2019
/
Published: 25 October 2019
(This article belongs to the Section Natural Products Chemistry)
Abstract
:Elaiophylins belong to a special family of 16-membered macrodiolides with C2-symmetry. They have exhibited remarkable biological activities, such as antimicrobial, anthelmintic, anticancer, immunosuppressive, anti-inflammatory, antiviral, and α-glucosidase inhibitory activities. A member of elaiophylins, efomycin M, is as a specific small molecule inhibitor of selectin in preclinical trial for the treatment of psoriasis, ischemia-reperfusion, and allergy. The biosynthetic investigation of elaiophylins has uncovered a unique thioesterase, which is responsible for the formation of C2-symmetric diolide. We herein summarize the natural occurrence, bioactivity, and biosynthesis of elaiophylins covering the literatures from 1959 to 2019. Hopefully, this review will inspire further research interests of these compounds and encourage the discovery of new analogues by metabolic engineering or genome mining.
Keywords:
elaiophylin; efomycin; macrodiolide; C2-symmetry; antimicrobial; anticancer; immunosuppressive; biosynthesis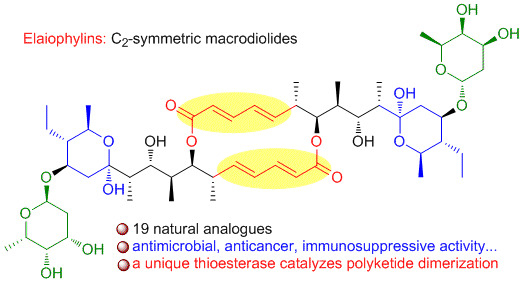
1. Introduction
The macrolides are a group of antibiotics whose activity stems from the presence of a large macrocyclic lactone ring to which one or more deoxy sugars may be attached. Normally, macrolides are assembled by the polyketide synthases (PKSs), which are large multifunctional enzymes that are composed of multiple discrete domains, each being responsible for catalyzing a single reaction of the polyketide biosynthetic process [1]. Macrodiolides, often classified into a small group of macrolides, represent a heterogenous group of microbial metabolites whose common feature is the presence of a cyclic diester moiety in their structure [2]. To date, only several families of macrodiolide natural products have been discovered, including elaiophylins [3], conglobatins [4], samroiyotmycins [5], bispolides [6], vermiculin [7], and pyrenophorin [8], from actinomycetes or fungi. Among them, elaiophylins are 16-membered glycosylated macrodiolides featuring with unusual C2-symmetry and hemiketal moiety. Elaiophylins have displayed a wide range of biological properties, such as antimicrobial, anthelmintic, anticancer, immunosuppressive, anti-inflammatory, antiviral, and α-glucosidase inhibitory activity. The anticancer and immunosuppressive activities have attracted extensive interests for further mechanism investigation. Efomycin M, as a new type of selective inhibitor of selectin functions, is currently in preclinical trial for the treatment of psoriasis, ischemia-reperfusion, and allergy. In this review, we summarize the natural occurrence, biological activity, the structure-activity relationship, and biosynthesis of elaiophylin family with structures that were derived from different Streptomyces species covering literatures from 1959 to 2019. The chemical synthesis of elaiophylins is not included.
2. Natural Occurrence
The first member, elaiophylin (1, also known as azalomycin B, salbomycin, efomycin E, or gopalamycin), was isolated from a culture of Streptomyces melanosporus in 1959 [3,9,10,11] and obtained as azalomycin B in 1960 [12]. The structure was assigned by chemical degradation, analyses of NMR and MS spectroscopic data [9,13,14]. Its absolute configuration was determined by a X-ray crystallographic analyses [15]. Later on, efomycin E derived from Streptomyces sp. BS 1261 and gopalamycin isolated from Streptomyces hygroscopicus MSU-625 and MSU-616 were found to be identical to elaiophylin [16,17]. Gopalamycin was originally assumed not to be elaiophylin, because it showed significant antifungal activity that elaiophylin did not have. However, their structures are the same according to the X-ray crystal analysis [17]. Subsequently, elaiophylin was found to be produced from various Streptomyces species [18]. Sporulation conditions of Streptomyces melanosporofaciens were optimized to maximize the concentration of elaiophylin at 1.2 g/kg fermentation medium and a described procedure resulted in the preparation of elaiophylin of pharmaceutical grade [19].
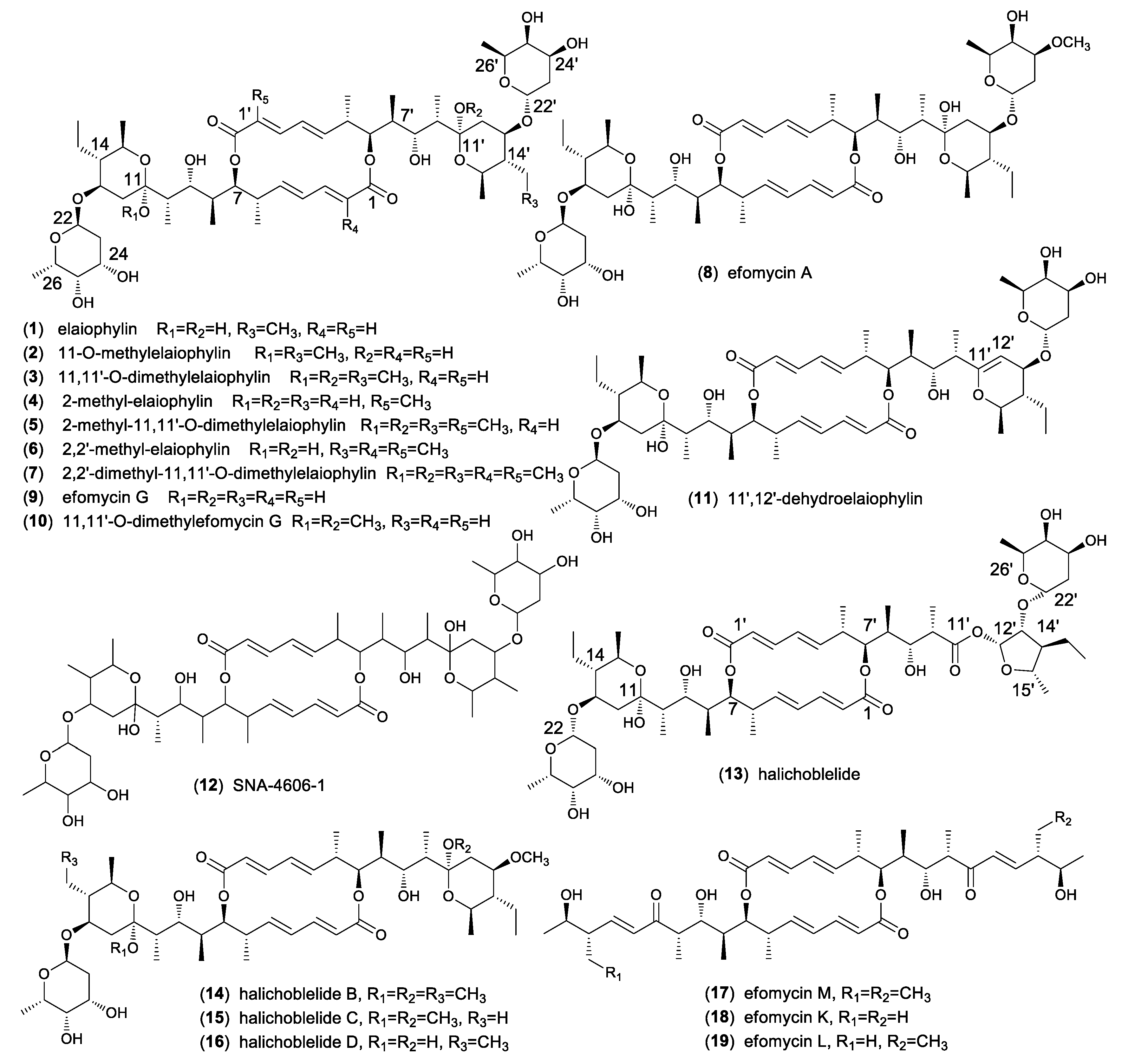
Along with the discovery of elaiophylin, eighteen analogues were obtained from different Streptomyces spp as shown in Figure 1. 11-O-methylelaiophylin (2) and 11,11’-O-dimethylelaiophylin (3) were isolated from the mycelium cake of Streptomyces strains HKI-0113 and HKI-0114. The structures were determined by MS and NMR spectrometric investigations [20]. From cultures of the Indonesian soil Streptomyces sp. ICBB 9297 four new elaiophylin macrolides, 2-methyl-elaiophylin (4), 2-methyl-11,11’-O-dimethylelaiophylin (5), 2,2’-dimethyl-elaiophylin (6), and 2,2’-dimethyl-11,11’-O-dimethylelaiophylin (7), along with 1–3 were isolated. Their structures contain an additional methyl groups at C-2 or C-2’, which are different from other elaiophylin analogues [21]. The Bayer company researchers reported the purification of efomycin(e)s A-F from the culture of a soil-derived Streptomyces sp. BS1261 and their mixtures showed potential as performance promoters in farm animals and exhibited antibacterial and antiviral activities [22]. The chemical and physical properties of efomycins A-F and their NMR data were reported. Among them, efomycin E were identical to elaiophylin and efomycin A (8) differed from elaiophylin by the methoxy substitution at C-24′. However, the structures of efomycin B, C, D, and F remained unassigned. From the same strain, efomycin G (9) was later isolated which possesses a methyl group at C-14’ in contrast to an ethyl group in elaiophylin [23] 11,11’-O-dimethylefomycin G (10) and 11′,12′-dehydroelaiophylin (11) were purified from culture extracts of a marine sediment derived strain Streptomyces sp. 7-145 by a PCR-based genetic screening experiment targeting the dTDP-glucose-4,6-dehydratase gene [24]. SNA-4606-1 (12) was isolated as an enzyme inhibitor from the cultured broth of Streptomyces sp. SNA-4606 along with 1 and 9. The structure was determined by NMR spectroscopic data, which showed the presence of methyl groups at both C-14 and C-14’. However, its relative configuration was not determined [25]. Halichoblelide (13) had been isolated from Streptomyces hygroscopicus originally separated from the marine fish Halichoeres bleekeri. Its structure and absolute configuration was elucidated by application of NMR spectroscopic analyses, the modified Mosher’s method, and CD spectra of the other products [26]. Two analogs, halichoblelides B (14) and C (15), were isolated from the same strain with one of the 6-deoxyfucose units being replaced by a methoxy group. Their absolute configurations were elucidated on the basis of spectroscopic analyses using one-dimensional (1D) and two-dimensional (2D) NMR techniques and chemical transformations [27]. Halichoblelide D (16) was separated and identified from the culture broth of mangrove strain Streptomyces sp. 219807. Its absolute configuration was determined by comparing the CD spectrum with those of the reported analogues [28]. Efomycin M (17) was originally prepared by base-catalyzed β-elimination of the deoxyfucose side chains of the mixture of efomycins [29]. Later, it was isolated from a rice endophytic actinomycete strain Streptomyces sp. BCC72023 along with efomycin G (9) [30]. Efomycin M differs from other elaiophylins by the absence of glycosylated dihydroxypyrane moiety and the presence of an unsaturated enone. As part of the explorations for antifungal activity of Streptomyces sp. M56 against both the termite mutualistic cultivar (Termitomyces spp.) and competitors/antagonists of this cultivar (Pseudoxylaria spp.), two new elaiophylin analogues, efomycins K (18) and L (19), and five known derivatives (1–3, 9) were isolated. Similar to efomycin M, 18 and 19 carried an unsaturated enone moiety and the structures were determined by analyzing NMR and HR-ESIMS data and by comparative CD spectroscopy [31].
The elaiophylin family shows high structural similarity in the diolide backbone, the hemiketal and 6-deoxyfucose moieties. Most structural variations of elaiophylin exist in different substitution patterns, such as proton or methyl groups at C-2 and/or C-2′, hydroxy, or methoxy groups at C-11 and/or C-11′, as well as methyl or ethyl groups at C-14 and/or C-14′. In contrast to elaiophylin (1) halichoblelide B-D (14–16) have commonness where one 6-deoxyfucose unit at C-13’ is replaced by a methoxy group. Efomycins K-M (17–19) are unique by the absence of glycosylated hemiketal moiety and the presence of unsaturated enone. The elaiophylin analogues that have methoxy substitutions at C-11 and/or C-11′ might be artifacts when MeOH was used during isolation and purification processes [32]. For example, when exposed to MeOH, the natural product elaiophylin (1) undergoes stereospecific acetal alkylation to 11-O-methylelaiophylin (2) and 11,11’-O-dimethylelaiophylin (3), while the co-metabolite efomycin G (9) undergoes transformation to 11,11’-O-dimethylefomycin G (10) [21].
3. Biological Activities
Elaiophylin and its derivatives have exhibited a myriad of activities, including antimicrobial, anthelmintic, anticancer, immunosuppressive, anti-inflammatory, antiviral, and α-glucosidase inhibitory activities. The mechanisms of elaiophylins on anticancer and immunosuppressive activities have been extensively explored. The relationship between the structure and biological activity of elaiophylin derivatives has also been studied, as outlined below.
3.1. Antimicrobial and Anthelmintic Activity
Elaiophylins exhibited antimicrobial activities against Gram-positive bacteria, but it was not active against Gram-negative bacteria, yeast, or fungi. Lee et al. reported an antimicrobial spectrum for elaiophylin with a concentration up to 100 μg/mL [33]. Although elaiophylin (1) has no activity against Candida albicans, it was able to markedly enhance rapamycin’s antifungal activity [34]. The mechanism of antibacterial activity of elaiophylin is not clear. Probably, it is related to the ability to form stable, long-lasting cation selective ion channels in microbial bilayer membranes [35].
The antimicrobial activities of 11-O-methylelaiophylin (2) and 11,11′-O-dimethylelaiophylin (3) were evaluated in an agar plate diffusion assay. They both showed antimicrobial activity against Gram-positive bacteria Bacillus subtilis, Staphylococcus aureus, Mycobacterium sp. SG, and Enterococcus faecium at the concentration of 50 μg/mL, but were not active against C. albicans [20]. In another biological test, 3 displayed lower antimicrobial activity than 2, which suggested the importance of free hemiketal hydroxyl in the antimicrobial activity [36]. Sheng et al. reported that compounds 4–7 displayed comparable antibacterial activity to 1 against S. aureus, with minimum inhibitory concentration (MIC) values in the range of 0.78–3.13 μg/mL, but did not inhibit the growth of B. subtilis, Pseudomonas aeruginosa, or Escherichia coli, with a concentration up to 100 μg/mL. Compounds 4 and 5, which have only one pendant methyl group at C-2, showed good activities against M. smegmatis with an MIC value of 6.25 μg/mL, whereas compound 7, which contains two methyl groups, and compounds 1–3, which do not have the C-2 and/or C-2′ methyl groups, showed no growth inhibition [21]. Efomycin G (9) exhibited antibacterial activity against Plasmodium falciparum, M. tuberculosis H37Ra, B. cereus, with MIC values of 2.37, 12.0, and 3.13 μg/mL, respectively. Efomycin M (17) showed antimalarial activity against Plasmodium falciparum with a MIC value of 5.23 μg/mL [30].
The elaiophylin (1) and its analogues (2, 3, 9, 10, and 11) were also evaluated for antibacterial activities against drug-resistant pathogens, including methicillin-resistant S. aureus (MRSA) and vancomycin-resistant enterococci (VRE) strains (Table 1). Particularly, compounds 1, 2, 9, and 11 showed potent activity with MIC values of 1–4 μg/mL against a number of MRSA and VRE strains, most of which were highly resistant (MIC > 256 μg/mL) to the controls. Furthermore, compounds 1, 2, 3, 9 inhibited methicillin resistant S. epidermidis (MRSE), with MIC values in the range of 2–16 μg/mL. The lack of cross-resistance between these compounds and erythromycin and azithromycin indicated that elaiophylins are probably not affected by the common macrolide resistance mechanism in Gram-positive bacteria. In comparison to 1, the 11-methoxylated derivative (2) suffered from a roughly two-fold decrease in activity against most strains (MIC values 1–2 vs. 2–4 μg/mL). Furthermore, the 11,11′-dimethoxylated derivatives 3 and 10 showed dramatic 8- to 32- fold reductions as compared to 1 and 9 with OH-11 and OH-11′ (MIC values 8–64 vs. 1–2 μg/mL), respectively. These results highlight the importance of the hemiketal moiety with respect to antibacterial activity. In addition, the replacement of ethyl substituent at C-14′ by a methyl group resulted in a 2- to 4- fold decrease in potency (1 vs. 9; 3 vs. 10), which indicates that the C-14/C-14′ alkyl group might also be crucial for antibacterial activity [24].
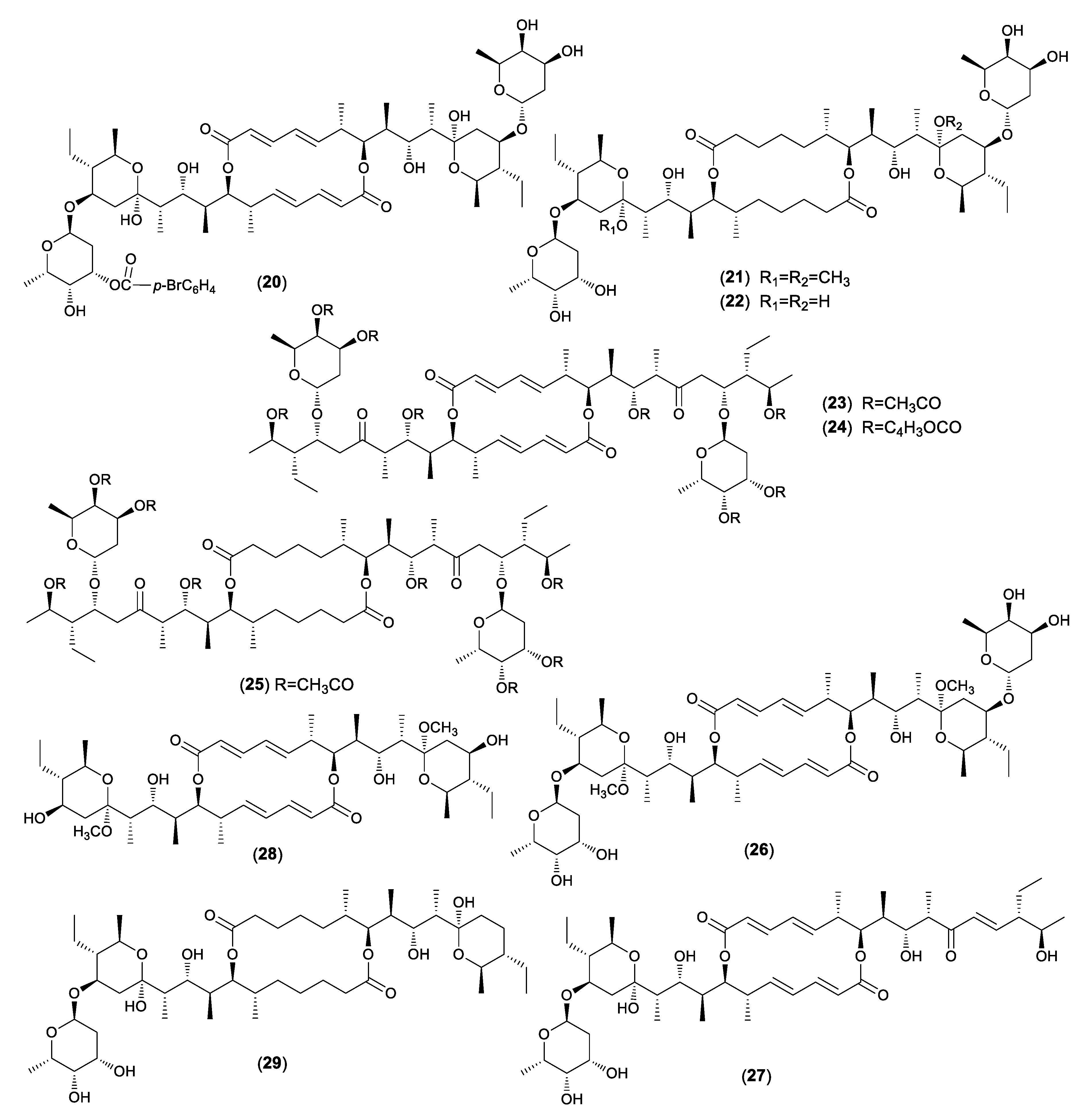
Hammann et al. described the chemical transformation of 34 acyl derivatives and six de-glycosidation products of elaiophylin (1) and antibacterial evaluation against Gram-positive bacteria (Figure 2) [37]. The acetalization of 1 to 2 led to a 50% reduction of antibacterial activity. All of the tetra-acyl derivatives were mainly inactive. The bromobenzoylation of 1 showed that the introduction of one acyl group (20) remained some activity against bacteria. However, in the higher acylated products, the activity was totally lost. The dimethyloctahydroelaiophylin (21) exhibited antibacterial activity, but, surprisingly, the octahydroelaiophylin (22) was inactive. The products with the open hemiketal ring (23–25) exhibited no activity. While the unsymmetric de-glycosidation products (26, 27) had small antibacterial activity, the symmetric de-glycosidation compounds (17, 28) were inactive. The remained activity of some derivatives could be explained by the fact that one part of the molecule still had an intact hemiketal moiety. Furthermore, compound 21 and the de-glycosidation products (25, 27, 29) expressed strong anthelmintic activity against the nematode Caenorhabditis elegans at a concentration of 100 ppm.
3.2. Anticancer Effect
3.2.1. Cytotoxicity.
Elaiophylin (1) showed moderate cytotoxicities against mouse embryo fibroblast (NIH-3T3), ras transformed-NIH3T3 (F25), human gastric cancer (SNU-1), human hepatocellular carcinoma (SNU-354), viblastine sensitive human epidermoid carcinoma (KB-3-1), and resistant cervical cancer (KB-V1) cell lines, with IC50 values in the range of 0.39–4.40 μM. However, the compound did not show any differential effect between these cells [33]. 11-O-methylelaiophylin (2) and 11,11′-O-dimethylelaiophylin (3) displayed cytotoxicity against L929 mouse fibroblast cells, K562 human leukemia cells, and HeLa cell cultures with IC50 values in the range of 0.7–2.4 μg/mL. In comparison to 1, 2 and 3 displayed somewhat lower cytotoxicity [20]. In another cytotoxic bioassay, 1 and 2 showed comparable cytotoxicities against human lung adenocarcinoma (A549), human prostatic carcinoma (PC-3), human breast adenocarcinoma (MCF-7 and MCF-7/ADR), mouse fibroblast (L929), NIH-3T3, F25, SNU-1, SNU-354, KB-3-1, and KB-V1 cell lines with adriamycin, but 3 was less active than 1 and 2 [36]. These results indicated that the OH-11 or OH-11′ of hemiketal was important for cytotoxic activity. Halichoblelide (12) showed potent cytotoxicity against the murine P388 lymphocytic leukemia cell line (ED50 = 0.63) and 39 human cancer cell lines. The tested mean value of log GI50 over all cell lines appeared to be −5.25 [26]. Halichoblelides B (13) and C (14) also exhibited significant cytotoxic activity against the P388 cell and, in addition, appreciable cytotoxicity against a disease-oriented panel of 39 human cancer cell lines [27]. Han et al. had evaluated the cytotoxic activities of halichoblelide D (15) as well as 1–5, which exhibited IC50 values ranging from 0.19 to 2.12 μM [28]. Efomycin G (9) and M (17) were evaluated for cytotoxicity against MCF-7, KB, and human small-cell lung cancer (NCI-H187) and non-cancerous (Vero) cells, with IC50 values of 10.34, 23.50, 8.03, 5.37 and 5.16, 7.86, 1.56, 3.68 μg/mL [30].
3.2.2. Autophagy Inhibitory Activity.
Elaiophylin (1) was identified as a novel autophagy inhibitor, with a significant antitumor effect as a single agent or in combination in human ovarian cancer cells. It can promote autophagosome accumulation, but block autophagic flux by attenuating lysosomal cathepsin activity, resulting in the accumulation of SQSTM1/p62 in various cell lines. Moreover, 1 can destabilize lysosomes, as indicated by CTSB/cathepsin B and CTSD/ cathepsin D release from lysosomes into the cytoplasm. Elaiophylin decreases cell viability, especially when with cisplatin or under hypoxic conditions. Furthermore, the administration of a lower dose (2 mg/kg) of 1 achieved a significant antitumor effect without toxicity in an orthotopic ovarian cancer model with metastasis. However, high doses (8 mg/kg) of 1 lead to dysfunction of Paneth cells, which resembled the intestinal phenotype of ATG16L1-deficient mice [38].
3.2.3. Inducing Apoptosis and Proliferation Activity.
Elaiophylin (1) has been assessed on multiple myeloma (MM) cells with mutant TP53. The results indicated that 1 exerted anti-myeloma activity by inducing apoptosis and proliferation arrest. Elaiophylin blocked autophagy flux in MM cells and subsequently induced the persistent activation of endoplasmic reticulum stress. In vivo studies revealed that 1 could suppress MM cell growth without obvious side effects in zebra fish embryo and mouse xenograft models. The exposure of human MM cells with mutant TP53 to 1 blocked autophagy flux and induced cell death, which partially involved ER stress-associated apoptosis. Therefore, it is a promising therapeutic strategy that 1 can targeted disrupt the cellular protein handling system for overcoming incurable MM, even with TP53 mutations [39].
3.2.4. Antiangiogenic Activity.
Elaiophylin (1) was demonstrated to have potent antiangiogenic activity from both in vitro and in vivo angiogenesis assays. Elaiophylin dramatically suppressed in vitro angiogenic characteristics, such as proliferation, migration, adhesion, invasion, and tube formation of human umbilical vein endothelial cells (HUVECs) stimulated by vascular endothelial growth factor (VEGF) at non-toxic concentrations. In addition, 1 immensely inhibited in vivo angiogenesis of the chorioallantoic membrane from growing chick embryos without cytotoxicity. The activation of VEGF receptor 2 in HUVECs by VEGF was inhibited by 1, which resulted in the suppression of VEGF-induced activation of downstream signaling molecules, Akt, extracellular signal-regulated kinase 1/2, c-Jun N-terminal kinase, p38, nuclear factor-κB, and matrix metalloproteinase -2 and -9. Elaiophylin down-regulated the expression of VEGF by inhibiting hypoxia inducible factor-1α accumulation in tumor cells. The results demonstrate that 1 blocks tumor cell-induced angiogenesis and may be utilized as a new type of antiangiogenic agent for cancer therapy [40].
3.3. Immunosuppressive and Anti-Inflammatory Activity
Elaiophylin (1) and 11-O-methylelaiophylin (2) showed a potent inhibitory effect (97.5% and 97.6%, respectively) on the activation of B cells by lipopolysaccharide as well as the proliferation of mouse splenic lymphocytes stimulated by mitogens at 1 μg/mL, but 11,11′-O-dimethylelaiophylin (3) did not. This result indicated that 1 and 2 would be effective immunosuppressants and 11-OH /11′-OH might be important in immunosuppressive activity [36].
Elaiophylin (1), efomycin A (8) and G (9) were observed to dose-dependently reduce the adhesion of human polymorphonuclear granulocytes to the endothelium. The inhibition rates of adhesion were 93%, 60%, and 95% by 1, 8, and 9, respectively, in a concentration of 5 μM. The results indicated the potential use of 1, 8, and 9 for the treatment of acute and chronic inflammations, especially to the use as medicaments in the therapy of myocardial infarct [16].
Efomycins were demonstrated as a new class of selective small-molecule inhibitors of selectin functions. Efomycin M (17), which was nontoxic and showed selective inhibitory effects on selectin-mediated leukocyte endothelial adhesion in vitro, significantly diminished rolling in mouse ear venules in vivo. In addition, 17 alleviated cutaneous inflammation in mouse models of psoriasis, one of the most common chronic inflammatory skin disorders. Molecular modeling demonstrated a spatial conformation of 17, mimicking naturally occurring selectin ligands. Efomycin M might be efficacious in the treatment of human inflammatory disorders [29,41]. Efomycin M could diminish thrombus formation and alleviated myocardial infarction and reperfusion injury by the inhibition of P-selectin–dependent platelet functions [42]. In addition, 17 therapy diminished L-selectin-mediated lymphocyte rolling and T cell mediated-allergic reactions [43]. Efomycin M is currently in preclinical study for the treatment of psoriasis, ischemia-reperfusion, and allergy [44].
3.4. Antiviral Activity and α-glucosidase Inhibitory Activity
Lee et al. discovered the antiviral effect of 11-O-methylelaiophylin (2) while screening for α-glucosidase inhibition. Compound 2 was found to be an α-glucosidase inhibitor with an IC50 value of 10 μM. It showed the mixed type inhibition of α-glucosidase with a Ki value of 5.94 μM. In addition, 2 inhibited the intracellular trafficking of hemagglutinin neuramidase (HN), a glycoprotein of Newcastle disease virus (NDV) in baby hamster kidney (BHK) cells. 11-O-methylelaiophylin inhibited the cell surface expression of NDV-HN glycoprotein without significantly affecting HN glycoprotein synthesis in NDV-infected BHK cells [45]. The connection of antiviral and α-glucosidase inhibitory activity might be explained for the reason that glucosidase inhibitions in the cell bring about an alteration on cell to cell signaling and virus recognition to the cell.
3.5. Other Activity
Elaiophylin (1), efomycin (9), and SNA-4606-1 (12) have shown inhibitory activity against testosterone 5a-reductase derived from rat prostate with IC50 values of 5.8, 8.7, and 6.6 μM, respectively [25]. Elaiophylin has been found to exert more inhibitory effect than dexamethasone on the synthesis of nitric oxide at non-toxic concentration to macrophage Raw 264.7 cells stimulated with lipopolysaccharide. This indicated that 1 had considerable potential as an inhibitor of NO production, which might have good correlation with the cytotoxic effect and anti- inflammatory activity [33].
4. Biosynthesis
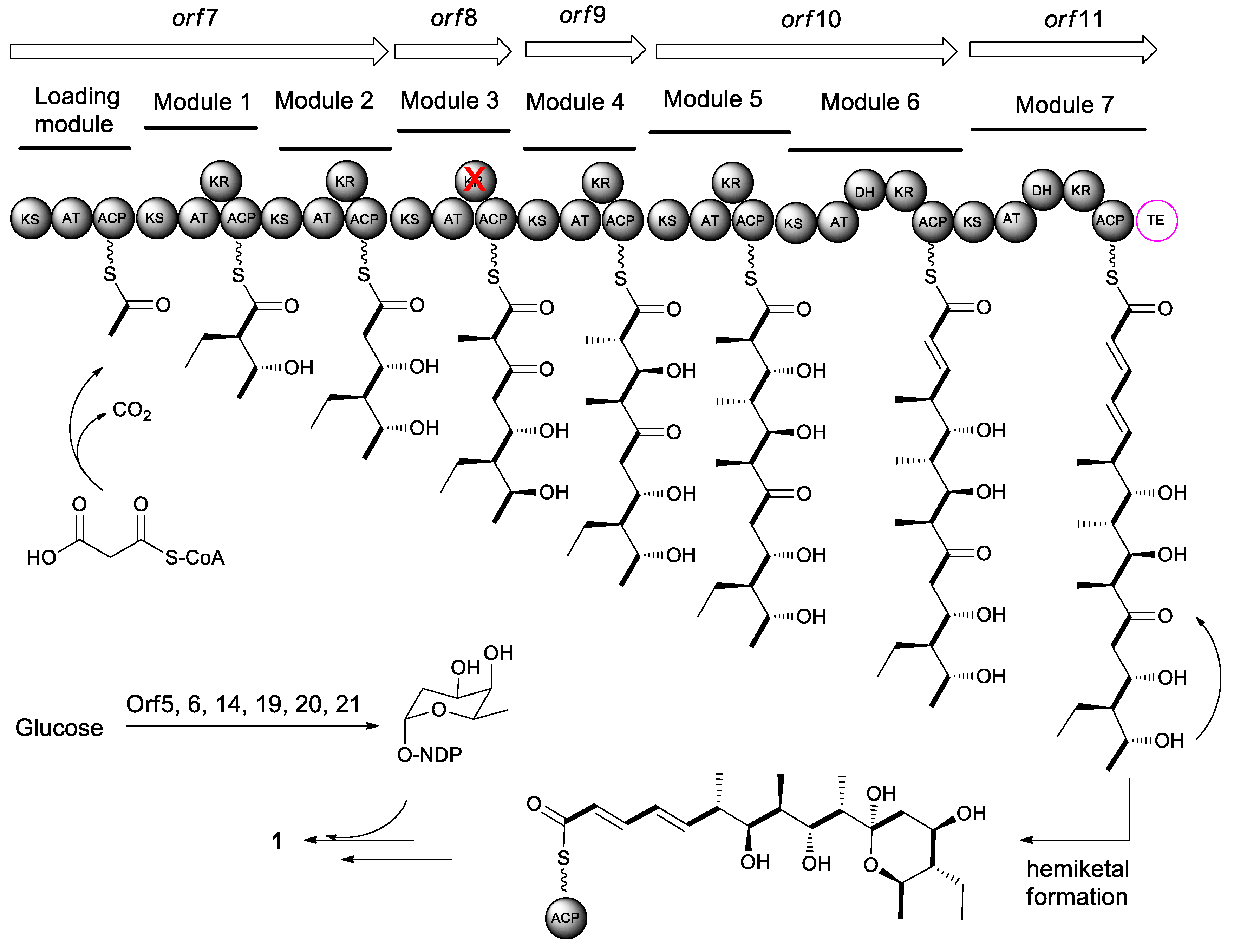
The feeding experiments with [1-13C, 18O2]-labeled precursors have revealed that all carbon and oxygen atoms of macrolactone core of elaiophylin (1) were derived from PKS precursors [46]. Elaiophylin is assembled by a type I polyketide synthases following the PKS pattern collinearity [1]. Generally, the individual domains in PKSs are grouped into functional modules containing basic modules, such as an acyl-carrier protein transferase domain (AT), an acyl-carrier protein domain (ACP), and a ketosynthase domain (KS), and modification modules, such as a β-ketoreductase domain (KR), a dehydratase domain (DH), and an enoyl reductase domain (ER). The growing polyketide chain transfers from the ACP of the first extender module to the KS of the next module. Once the carbon chain reaches the final extension, the linear polyketide is released by a thioesterase (TE).
The elaiophylin biosynthetic gene clusters (ela) have been identified in Streptomyces sp. DSM4137 [47], Streptomyces sp. NRRL 30748 [48], Streptomyces autolyticus CGMCC 0516 [49], and Streptomyces sp. M56. [31] The ela cluster in Streptomyces sp. NRRL 30748 comprises 24 individual open reading frames (orf) spanning approximately 63.2 kb DNA. The core cluster consists of five PKSs genes (orf 7–11) that include eight PKS modules in total as well as a free-standing TE domain (Figure 3). Orfs 5, 6, 14, and 19–21 are involved in the biosynthesis and loading of 6-deoxyfucose sugar unit [48]. The alignment of conserved motifs of active site of AT domain in the polyketide backbones is consistent and collinear with the elaiophylin structure [21]. The structural variations, such as the pendant methyl group at C-2/C-2′ and the methyl or ethyl groups at C-14/C-14′ in some elaiophylin analogues, might be due to relaxed substrate specificity of the PKSs that are responsible for elaiophylin biosynthesis in the individual producing strains. A TetR family regulator GdmRIII was found to affect the biosynthetic pathways of both geldanamycin and elaiophylin in the strain of Streptomyces autolyticus CGMCC0516. The GdmRIII plays a positive regulatory role in the biosynthesis of geldanamycin, but a negative role in elaiophylin [50].
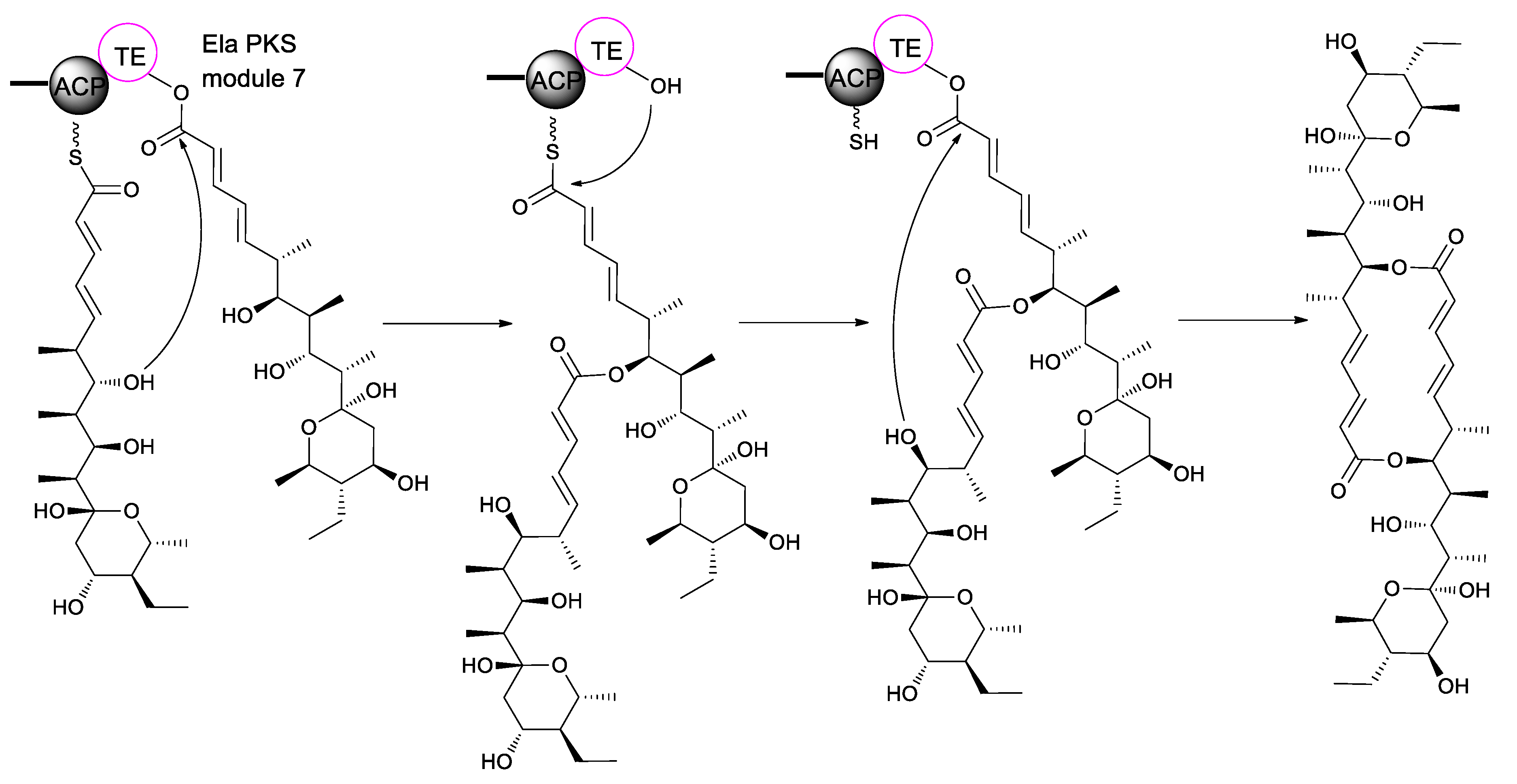
Biosynthetically, elaiophylin is quite interesting, as it is formed from two linear polyketide precursors, which, when dimerized, produce a cyclic polyketide that possesses a unique C2-symmetry that is only found in a small group of polyketides. To probe the mechanism and selectivity of diolide formation, Zhou et al. had constructed ring formation in vitro by using a non-natural substrate [51]. The incubation of recombinant elaiophylin thioesterase with a synthetic analogue of the presumed monomeric polyketide precursor of elaiophylin, specifically its N-acetylcysteamine thioester, produced a novel 16-membered C2-symmetric macrodiolide. A linear dimeric thioester is an intermediate in ring formation, which indicates the iterative use of the thioesterase active site in ligation and subsequent cyclization. The iterative ela-TE must catalyze a total of two acylation and two deacylation reactions to form the diolide (Figure 4). Furthermore, the ela thioesterase acts on a mixture of pentaketide and tetraketide thioesters to give both the symmetric decaketide diolides and the asymmetric hybrid nonaketide diolides. Such thioesterases have potential as tools for in vitro construction of novel diolides.
5. Conclusions
This review has highlighted various aspects of elaiophylins, including natural discovery, biological activities, relationship between structure and activity, and biosynthesis. Up to date, 19 natural elaiophylin derivatives have been discovered from different Streptomyces spp. Elaiophylins have displayed a wide range of biological properties, especially the remarkable antimicrobial, anticancer, and immunosuppressive activities. Elaiophylins have potent antimicrobial activities against Gram-positive bacteria, even the drug-resistant pathogens, including MRSA and VRE strains, which indicated that elaiophylins might not be affected by the common macrolide resistance mechanism of bacteria. The anticancer effect of elaiophylin (1) has been extensively studied that autophagy, inducing apoptosis and proliferation, and antiangiogenic activity were all probably involved. Structure-activity relationship results highlight the importance of the C2-symmetric diolide skeleton and the hemiketal moiety with respect to both antibacterial and anticancer activities. As for the aspect of immunosuppressive activity, efomycin M (17), without glycosylated hemiketal moiety and with unsaturated enone, is a promising immunosuppressant lead for the treatment of psoriasis, ischemia-reperfusion, and allergy. The ela thioesterase is unique in the TE family of polyketide. It has shown great potential as tools to the in vitro construction of novel diolides by acting on a mixture of various pentaketide or tetraketide thioesters. It is also promising to discover new elaiophylin analogues by metabolic engineering or genomic mining with the genomic information of ela thioesterase. Hopefully, this review will provide reference for further research interests of these compounds and lay the foundation for new drug discovery.
Author Contributions
M.G. conceptualized the review and drafted the initial version of the manuscript. M.-x.Z. and W.-h.W. contributed in undertaking the database search for the literature and discussion. P.S. supervised and revised the manuscript. All authors read and approved the final manuscript.
Funding
This work was funded by National Natural Science Foundation of China (81622044, 81573342, and 41876184), Science and Technology Commission of Shanghai Municipality (18ZR1449600), and Shanghai Engineering Center of Dairy Biotechnology (19DZ2281400).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hertweck, C. The biosynthetic logic of polyketide diversity. Angew. Chem. Int. Ed. Engl. 2009, 48, 4688–4716. [Google Scholar] [CrossRef] [PubMed]
- Ōmura, S. Macrolide Antibiotics: Chemistry, Biology, and Practice, 2nd ed.; Academic Press: Boston, MA, USA, 2003. [Google Scholar]
- Arcamone, F.; Bertazzoli, C.; Ghione, M.; Scotti, T. Melanosporin and elaiophylin, new antibiotics from Streptomyces melanosporus (sive melonsporofaciens) n. sp. G. Microbiol. 1959, 7, 207–216. [Google Scholar]
- Westley, J.W.; Liu, C.M.; Evans, R.H.; Blount, J.F. Conglobatin, a novel macrolide dilactone from Streptomyces conglobatus ATCC 31005. J. Antibiot. 1979, 32, 874–877. [Google Scholar] [CrossRef] [PubMed]
- Dramae, A.; Nithithanasilp, S.; Choowong, W.; Rachtawee, P.; Prabpai, S.; Kongsaeree, P.; Pittayakhajonwut, P. Antimalarial 20-membered macrolides from Streptomyces sp. BCC33756. Tetrahedron 2013, 69, 8205–8208. [Google Scholar] [CrossRef]
- Okujo, N.; Iinuma, H.; George, A.; Eim, K.S.; Li, T.L.; Ting, N.S.; Jye, T.C.; Hotta, K.; Hatsu, M.; Fukagawa, Y.; et al. Bispolides, novel 20-membered ring macrodiolide antibiotics from microbispora. J. Antibiot. 2007, 60, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Fuska, J.; Nemec, P.; Kuhr, I. Vermiculine, a new antiprotozoal antibiotic from Penicillium vermiculatum. J. Antibiot. 1972, 25, 208–211. [Google Scholar] [CrossRef]
- Seebach, D.; Seuring, B.; Kalinowski, H.-O.; Lubosch, W.; Renger, B. Synthesis and determination of the absolute configuration of pyrenophorin and vermiculin. Angew. Chem. Int. Ed. Engl. 1977, 16, 264–265. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, H.; Keller-Schierlein, W. Stoffwechselprodukte von mikroorganismen. 202. mitteilung. Strukturaufklärung von elaiophylin: Spektroskopische untersuchungen und abbau. Helv. Chim. Acta 1981, 64, 407–424. [Google Scholar] [CrossRef]
- Neupert-Laves, K.; Dobler, M. Stoffwechselprodukte von mikroorganismen. 211. milteilung. röntgenstrukturanalyse von elaiophylin. Helv. Chim. Acta 1982, 65, 262–267. [Google Scholar] [CrossRef]
- Paulus, E.F.; Vertesy, L.; Sheldrick, G. Structure of salbomycin, C54H88O18. 2H2O. Acta Crystallogr. C 1984, 40, 700–703. [Google Scholar] [CrossRef]
- Arai, M. Azalomycins B and F, two new antibiotics. II. Properties of azalomycins B and F. J Antibiot. 1960, 13, 51–56. [Google Scholar] [PubMed]
- Takahashi, S.; Arai, M.; Oki, E. Chemical studies on azalomycins. I. Preliminary study of azalomycin-B. Chem. Pharm. Bull. 1967, 15, 1651–1656. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Oki, E. Chemical studies on azalomycins. 3. Alkaline degradation of azalomycin-B. Chem. Pharm. Bull. 1967, 15, 1726–1732. [Google Scholar] [CrossRef] [PubMed]
- Ley, S.V.; Neuhaus, D.; Williams, D.J. A conformational study of elaiophylin by X-ray crystallography and difference 1H NMR methods; observation of a selective sign reversal of the nuclear overhauser effect. Tetrahedron Lett. 1982, 23, 1207–1210. [Google Scholar] [CrossRef]
- Muller, H.; Bischoff, E.; Fugmann, B.; Weber, K.; Frobel, K.; Rosen, B.; Grutzmann, R.; Karmann, G.; Kohlsdorfer, C. Efomycins a, e and g as Antiinflammatory Agents. U.S. Patent 5,185,326A, 9 February 1993. [Google Scholar]
- Nair, M.G.; Chandra, A.; Thorogood, D.L.; Ammermann, E.; Walker, N.; Kiehs, K. Gopalamicin, an antifungal macrodiolide produced by soil actinomycetes. J. Agric. Food Chem. 1994, 42, 2308–2310. [Google Scholar] [CrossRef]
- Lima, S.M.; Melo, J.G.; Militao, G.C.; Lima, G.M.; do Carmo, A.L.M.; Aguiar, J.S.; Araujo, R.M.; Braz-Filho, R.; Marchand, P.; Araujo, J.M.; et al. Characterization of the biochemical, physiological, and medicinal properties of Streptomyces hygroscopicus ACTMS-9H isolated from the Amazon (Brazil). Appl. Microbiol. Biotechnol. 2017, 101, 711–723. [Google Scholar] [CrossRef]
- Cikosova, M.; Blazsek, M.; Kubis, M.; Gajdosikova, J.; Borosova, G. Biotechnological preparation of the elaiophylin. Folia Microbiol. (Praha) 2004, 49, 731–736. [Google Scholar] [CrossRef]
- Ritzau, M.; Heinze, S.; Fleck, W.F.; Dahse, H.M.; Grafe, U. New macrodiolide antibiotics, 11-O-monomethyl- and 11, 11’-O-dimethylelaiophylins, from Streptomyces sp. HKI-0113 and HKI-0114. J. Nat. Prod. 1998, 61, 1337–1339. [Google Scholar] [CrossRef]
- Sheng, Y.; Lam, P.W.; Shahab, S.; Santosa, D.A.; Proteau, P.J.; Zabriskie, T.M.; Mahmud, T. Identification of elaiophylin skeletal variants from the Indonesian Streptomyces sp. ICBB 9297. J. Nat. Prod. 2015, 78, 2768–2775. [Google Scholar] [CrossRef]
- Frobel, K.; Bischoff, E.; Muller, H.; Salcher, O.; De Jong, A.; Berschauer, F.; Scheer, M. Efomycins as Performance Promoters in Animals. U.S. Patent 5,073,369, 17 December 1991. [Google Scholar]
- Frobel, K.; Muller, H.; Bischoff, E.; Salcher, O.; De Jong, A.; Berschauer, F.; Scheer, M. Efomycin G and It’s Use as Yield Promoter in Animals. U.S. Patent 4,927,810, 22 May 1990. [Google Scholar]
- Wu, C.; Tan, Y.; Gan, M.; Wang, Y.; Guan, Y.; Hu, X.; Zhou, H.; Shang, X.; You, X.; Yang, Z.; et al. Identification of elaiophylin derivatives from the marine-derived actinomycete Streptomyces sp. 7-145 using PCR-based screening. J. Nat. Prod. 2013, 76, 2153–2157. [Google Scholar] [CrossRef]
- Nakakoshi, M.; Kimura, K.; Nakajima, N.; Yoshihama, M.; Uramoto, M. SNA-4606-1, a new member of elaiophylins with enzyme inhibition activity against testosterone 5α-reductase. J. Antibiot. 1999, 52, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Minoura, K.; Numata, A. Halichoblelide, a potent cytotoxic macrolide from a Streptomyces species separated from a marine fish. Tetrahedron Lett. 2002, 43, 1721–1724. [Google Scholar] [CrossRef]
- Yamada, T.; Kikuchi, T.; Tanaka, R.; Numata, A. Halichoblelides B and C, potent cytotoxic macrolides from a Streptomyces species separated from a marine fish. Tetrahedron Lett. 2012, 53, 2842–2846. [Google Scholar] [CrossRef]
- Han, Y.; Tian, E.; Xu, D.; Ma, M.; Deng, Z.; Hong, K. Halichoblelide D, a new elaiophylin derivative with potent cytotoxic activity from mangrove-derived Streptomyces sp. 219807. Molecules 2016, 21, 970. [Google Scholar] [CrossRef]
- Schon, M.P.; Krahn, T.; Schon, M.; Rodriguez, M.L.; Antonicek, H.; Schultz, J.E.; Ludwig, R.J.; Zollner, T.M.; Bischoff, E.; Bremm, K.D.; et al. A new specific inhibitor of selectin, impairs leukocyte adhesion and alleviates cutaneous inflammation. Nat. Med. 2002, 8, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Supong, K.; Thawai, C.; Choowong, W.; Kittiwongwattana, C.; Thanaboripat, D.; Laosinwattana, C.; Koohakan, P.; Parinthawong, N.; Pittayakhajonwut, P. Antimicrobial compounds from endophytic Streptomyces sp. BCC72023 isolated from rice (Oryza sativa L.). Res. Microbiol. 2016, 167, 290–298. [Google Scholar] [CrossRef]
- Klassen, J.L.; Lee, S.R.; Poulsen, M.; Beemelmanns, C.; Kim, K.H. Efomycins K and L from a termite-associated Streptomyces sp. M56 and their putative biosynthetic origin. Front. Microbiol. 2019, 10, 1739. [Google Scholar] [CrossRef]
- Capon, R.J. Extracting value: Mechanistic insights into the formation of natural product artifacts–case studies in marine natural products. Nat. Prod. Rep. 2019, in press. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Kim, M.-S.; Kim, H.-S.; Kim, Y.-H.; Hong, S.-D.; Lee, J.-J. Structure determination and biological activities of elaiophylin produced by Streptomyces sp. MCY-846. J. Microbiol. Biotechnol. 1996, 6, 245–249. [Google Scholar]
- Fang, A.; Wong, G.K.; Demain, A.L. Enhancement of the antifungal activity of rapamycin by the coproduced elaiophylin and nigericin. J. Antibiot. 2000, 53, 158–162. [Google Scholar] [CrossRef]
- Grigoriev, P.A.; Schlegel, R.; Gräfe, U. Cation selective ion channels formed by macrodiolide antibiotic elaiophylin in lipid bilayer membranes. Bioelectrochemistry 2001, 54, 11–15. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Kim, H.-S.; Kim, Y.-H.; Han, S.-B.; Kim, H.-M.; Hong, S.-D.; Lee, J.-J. Immunosuppressive activity of elaiophylins. J. Microbiol. Biotechnol. 1997, 7, 272–277. [Google Scholar]
- Hammann, P.; Kretzschmar, G.; Seibert, G. Secondary metabolites by chemical screening. 7. I. Elaiophylin derivatives and their biological activities. J. Antibiot. 1990, 43, 1431–1440. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Fang, Y.; Yang, Y.; Qin, Y.; Wu, P.; Wang, T.; Lai, H.; Meng, L.; Wang, D.; Zheng, Z.; et al. Elaiophylin, a novel autophagy inhibitor, exerts antitumor activity as a single agent in ovarian cancer cells. Autophagy 2015, 11, 1849–1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Zhou, P.; Chen, X.; Zhao, L.; Tan, J.; Yang, Y.; Fang, Y.; Zhou, J. The novel autophagy inhibitor elaiophylin exerts antitumor activity against multiple myeloma with mutant TP53 in part through endoplasmic reticulum stress-induced apoptosis. Cancer Biol. Ther. 2017, 18, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.N.; Jang, J.P.; Han, J.M.; Jang, J.H.; Ahn, J.S.; Jung, H.J. Antiangiogenic potential of microbial metabolite elaiophylin for targeting tumor angiogenesis. Molecules 2018, 23, 563. [Google Scholar] [CrossRef]
- Antonicek, H.-P.; Bischoff, E.; Gondol, D.; Gutbrod, O.; Krahn, T.; Rodriguez, M.-L.; Schütz, H. Use of Efomycins. U.S. Patent 6,291515 B1, 18 September 2001. [Google Scholar]
- Oostingh, G.J.; Pozgajova, M.; Ludwig, R.J.; Krahn, T.; Boehncke, W.H.; Nieswandt, B.; Schon, M.P. Diminished thrombus formation and alleviation of myocardial infarction and reperfusion injury through antibody- or small-molecule-mediated inhibition of selectin-dependent platelet functions. Haematologica 2007, 92, 502–512. [Google Scholar] [CrossRef]
- Oostingh, G.J.; Ludwig, R.J.; Enders, S.; Gruner, S.; Harms, G.; Boehncke, W.H.; Nieswandt, B.; Tauber, R.; Schon, M.P. Diminished lymphocyte adhesion and alleviation of allergic responses by small-molecule- or antibody-mediated inhibition of L-selectin functions. J. Investig. Dermatol. 2007, 127, 90–97. [Google Scholar] [CrossRef]
- Rossi, B.; Constantin, G. Anti-selectin therapy for the treatment of inflammatory diseases. Inflamm. Allergy Drug Targets 2008, 7, 85–93. [Google Scholar] [CrossRef]
- Lee, D.; Woo, J.-K.; Kim, D.; Kim, M.; Cho, S.K.; Kim, J.H.; Park, S.P.; Lee, H.Y.; Riu, K.Z.; Lee, D.-S. Antiviral activity of methylelaiophylin, an alpha-glucosidase inhibitor. J. Microbiol. Biotechnol. 2011, 21, 263–266. [Google Scholar]
- Gerlitz, M.; Hammann, P.; Thiericke, R.; Rohr, J. The biogenetic origin of the carbon skeleton and the oxygen atoms of elaiophylin, a symmetric macrodiolide antibiotic. J. Org. Chem. 1992, 57, 4030–4033. [Google Scholar] [CrossRef]
- Haydock, S.F.; Mironenko, T.; Ghoorahoo, H.I.; Leadlay, P.F. The putative elaiophylin biosynthetic gene cluster in Streptomyces sp. DSM4137 is adjacent to genes encoding adenosylcobalamin-dependent methylmalonyl CoA mutase and to genes for synthesis of cobalamin. J. Biotechnol. 2004, 113, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Haltli, B.A. Elaiophylin Biosynthetic Gene Cluster. U.S. Patent 7,595,187B2, 29 September 2009. [Google Scholar]
- Yin, M.; Jiang, M.; Ren, Z.; Dong, Y.; Lu, T. The complete genome sequence of Streptomyces autolyticus CGMCC 0516, the producer of geldanamycin, autolytimycin, reblastatin and elaiophylin. J. Biotechnol. 2017, 252, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Yin, M.; Wu, S.; Han, X.; Ji, K.; Wen, M.; Lu, T. GdmRIII, a TetR family transcriptional regulator, controls geldanamycin and elaiophylin biosynthesis in Streptomyces autolyticus CGMCC0516. Sci. Rep. 2017, 7, 4803. [Google Scholar] [CrossRef]
- Zhou, Y.; Prediger, P.; Dias, L.C.; Murphy, A.C.; Leadlay, P.F. Macrodiolide formation by the thioesterase of a modular polyketide synthase. Angew. Chem. Int. Ed. Engl. 2015, 54, 5232–5235. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structures of naturally occurring elaiophylins.

Figure 2.
Structures of chemical derivatives of elaiophylin.

Figure 3.
Proposed elaiophylin PKS assembly line.

Figure 4.
Biosynthetic mechanisms for the formation of elaiophylin diolide.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Antimicrobial bioassay results (MIC, μg/mL) for 1–3, 9, 10, and 11 a.
| Strains | Phenotype | 1 | 2 | 3 | 9 | 10 | 11 | Azithromycin | Erythromycin | Oxacillin | Vancomycin |
|---|---|---|---|---|---|---|---|---|---|---|---|
| S. aureus ATCC 29213 | MSSA | 1 | 2 | 16 | 2 | 32 | 2 | 2 | 0.25 | 0.5 | 0.5 |
| S. aureus 09-6 | MSSA | 1 | 2 | 16 | 2 | 32 | 2 | 2 | 0.25 | 0.5 | 0.5 |
| S. aureus ATCC 33591 | MRSA | 1 | 2 | 16 | 2 | 32 | 2 | >256 | >256 | >256 | 0.5 |
| S. aureus 09-13 | MRSA | 1 | 2 | 16 | 2 | 32 | 2 | >256 | >256 | >256 | 0.5 |
| S. aureus R6101 | MRSA | 2 | 1 | 4 | 2 | 16 | >64 | >256 | NT | >256 | NT |
| S. aureus ATCC 6538P | Thios-R | 0.5 | 2 | 16 | 2 | >128 | 4 | >256 | NT | NT | NT |
| Staphylococcus epidermidis ATCC 12228 | MSSE | 1 | 2 | 16 | 4 | 64 | >64 | >256 | 16 | >256 | 0.5 |
| S. epidermidis 09-9 | MSSE | 2 | 2 | 16 | NT | NT | NT | NT | NT | NT | 0.5 |
| S. epidermidis 09-3 | MRSE | 2 | 2 | 16 | 4 | 64 | >64 | >256 | >256 | >256 | 4 |
| Enterococcus faecalis ATCC 29212 | VSE | 1 | 32 | 16 | 2 | 64 | 2 | 8 | 16 | 4 | 1 |
| E. faecalis 09-8 | VSE | 1 | 2 | 16 | 16 | 64 | 2 | >256 | 32 | >256 | 1 |
| E. faecalis ATCC 51299 | VRE | 1 | 1 | 16 | 2 | 64 | 2 | >256 | 8 | >256 | 4 |
| E. faecalis W4138 | VRE | 1 | 2 | 16 | 4 | 64 | 4 | >256 | NT | NT | 4 |
| E. faecalis R6512 | VRE | 1 | 1 | 8 | 2 | >128 | 4 | >256 | NT | NT | >256 |
| Enterococcus faecium 09-10 | VSE | 2 | 2 | 32 | >64 | 64 | >64 | >256 | >256 | >256 | 1 |
| E. faecium ATCC 700221 | VRE | 1 | 2 | 16 | 2 | 64 | 4 | >256 | 64 | >256 | 128 |
| Micrococcus faecium ATCC 10240 | Apram-R | 0.5 | 2 | 16 | 2 | 128 | 2 | 2 | NT | NT | NT |
| Escherichia coli 09-1 | ESBL-prod | >256 | >256 | >256 | >64 | >128 | >64 | 64 | >256 | >256 | >256 |
| E. coli 09-20 | BL-prod | >256 | >256 | >256 | >64 | >128 | >64 | >256 | >256 | >256 | >128 |
| Klebsiella pneumoniae ATCC 700603 | ESBL-prod | >256 | >256 | >256 | >64 | >128 | >64 | 64 | >256 | >256 | >128 |
| K. pneumoniae ATCC BAA-2146 | NDM-1-prod | >256 | >256 | >256 | >64 | >128 | >64 | 64 | >256 | >256 | >128 |
| Pseudomonas aeruginosa ATCC 27853 | BL-prod | >256 | >256 | >256 | >64 | >128 | >64 | 256 | >256 | >256 | >128 |
| Morganella morganii ATCC 25830 | BL-prod | >256 | >256 | >256 | >64 | >128 | >64 | 64 | >256 | 256 | 128 |
a Abbreviations: Methicillin-susceptible S. aureus. (MSSA), Methicillin-resistant S. aureus (MRSA). Not tested (NT). Thiostrepton-resistant (Thios-R). Methicillin-susceptible S. epidermidis (MSSE). Methicillin-resistant S. epidermidis (MRSE). Vancomycin-susceptible Enterococcus (VSE). Vancomycin-resistant Enterococcus (MRE). Apramycin-resistant (Apram-R). Extended spectrum β-lactamase (ESBL). β-lactamase (BL). New Delhi metallo-β-lactamase 1 (NDM-1).
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gui, M.; Zhang, M.-x.; Wu, W.-h.; Sun, P. Natural Occurrence, Bioactivity and Biosynthesis of Elaiophylin Analogues. Molecules 2019, 24, 3840. https://doi.org/10.3390/molecules24213840
AMA Style
Gui M, Zhang M-x, Wu W-h, Sun P. Natural Occurrence, Bioactivity and Biosynthesis of Elaiophylin Analogues. Molecules. 2019; 24(21):3840. https://doi.org/10.3390/molecules24213840
Chicago/Turabian StyleGui, Min, Meng-xue Zhang, Wen-hui Wu, and Peng Sun. 2019. "Natural Occurrence, Bioactivity and Biosynthesis of Elaiophylin Analogues" Molecules 24, no. 21: 3840. https://doi.org/10.3390/molecules24213840