
Colletotrichum spp. from Soybean Cause Disease on Lupin and Can Induce Plant Growth-Promoting Effects

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Fungal Isolates and Culture Conditions
2.3. Hypocotyl Assay
2.4. Seed Inoculation Assay
2.5. Statistical Evaluation
3. Results
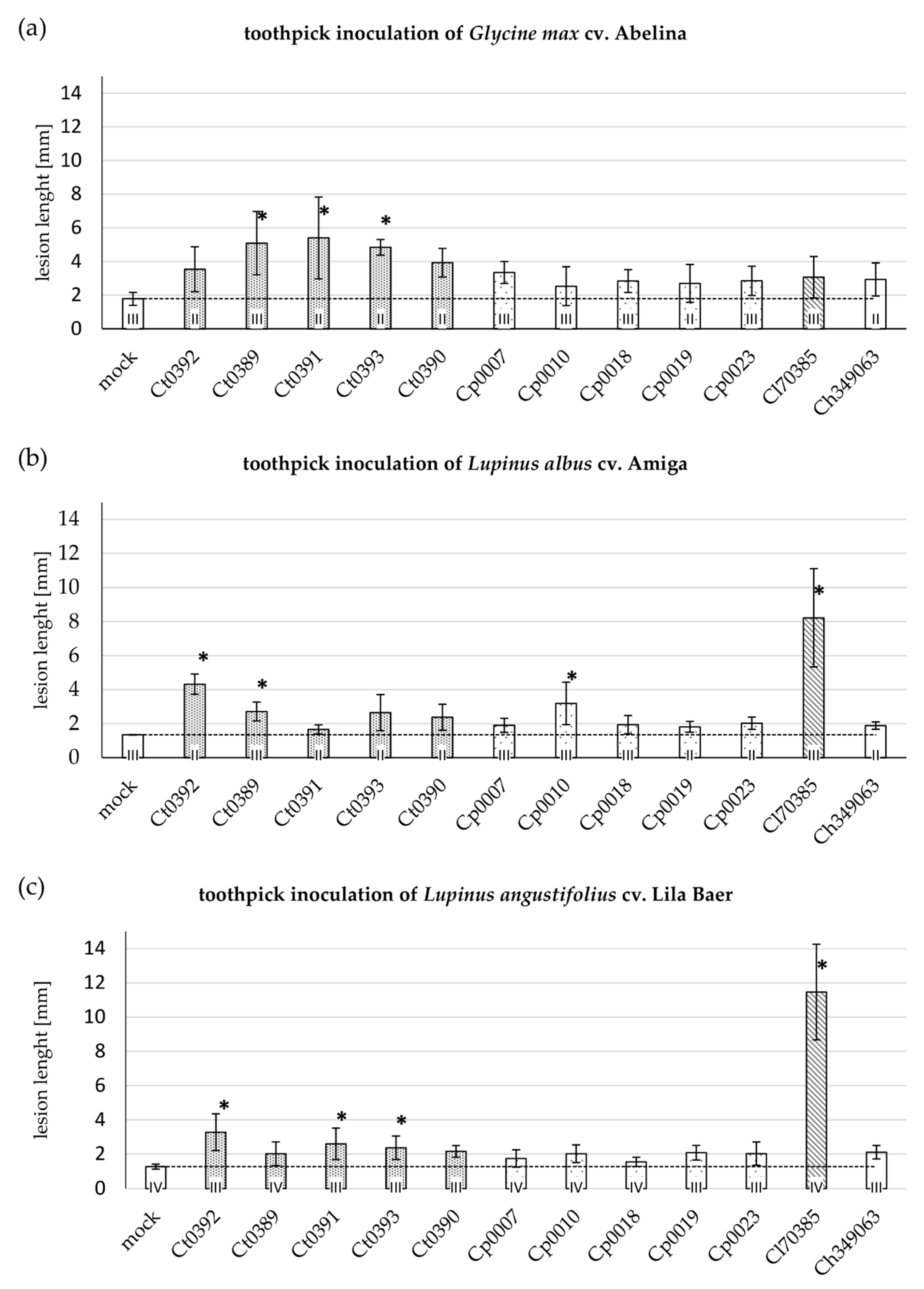
3.1. Cross-Infection Assays on Soybean and Lupin Hypocotyl
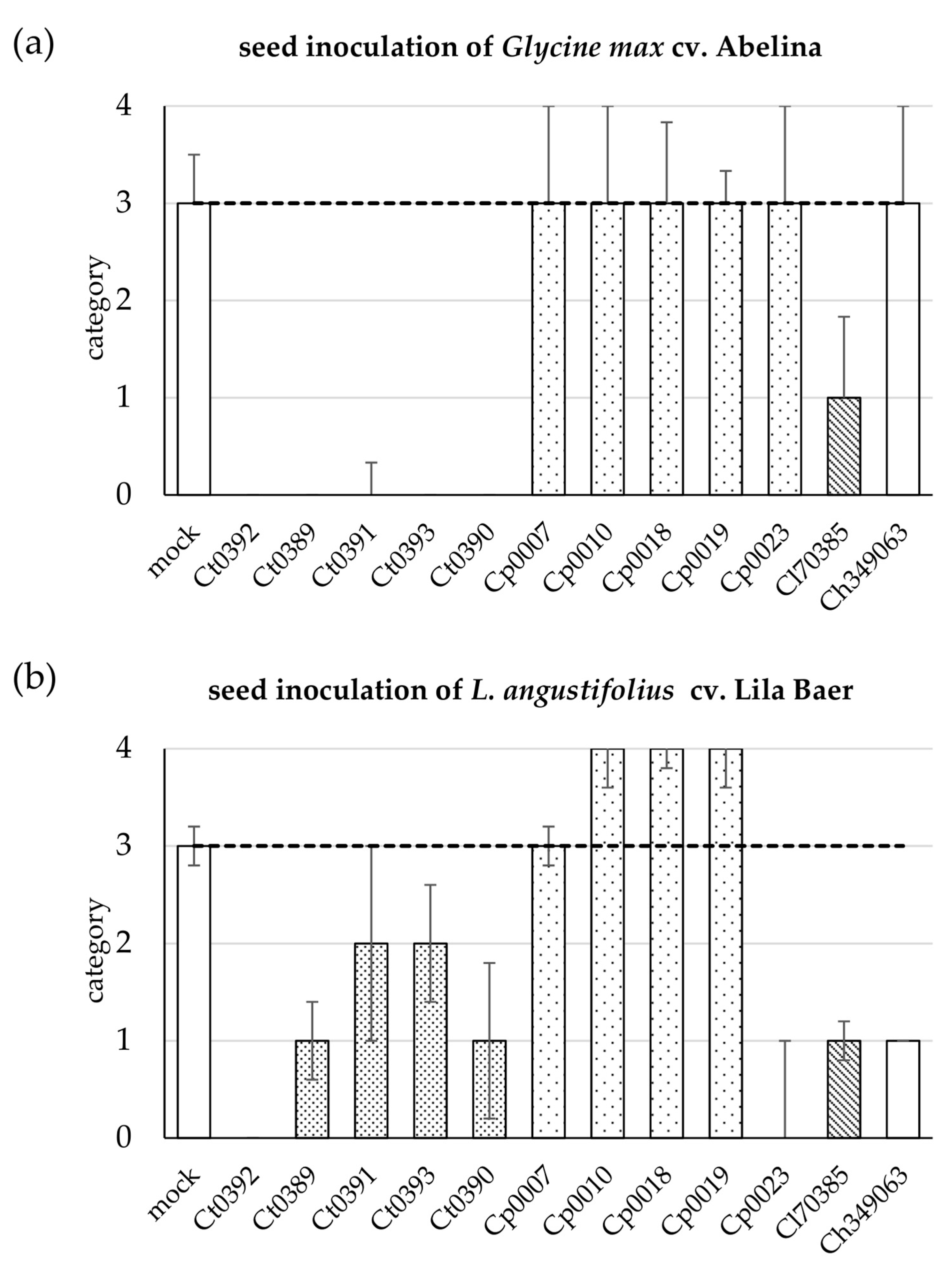
3.2. Cross-Infection Assays Using Inoculated Seeds of Soybean and Lupin
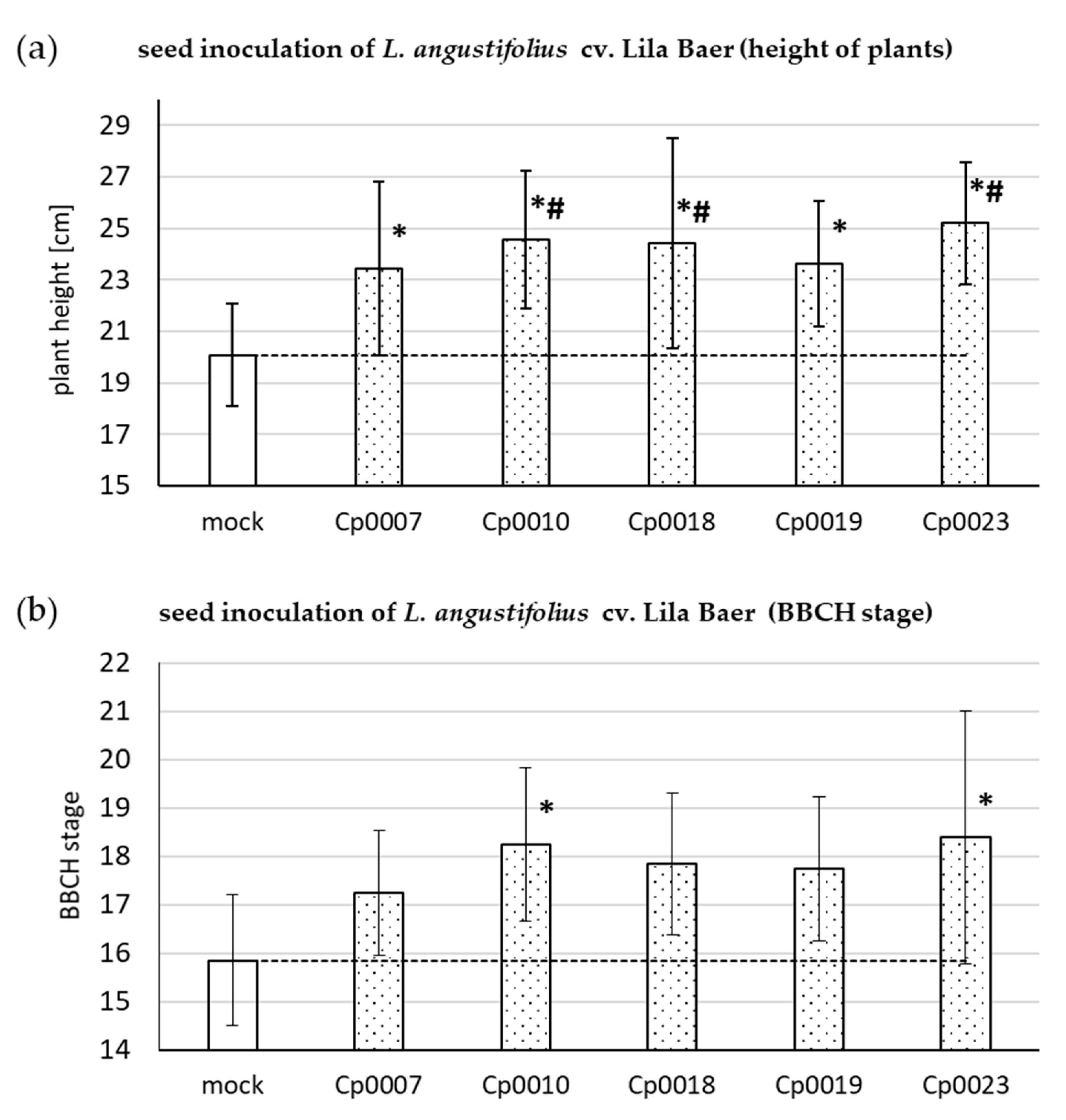
3.3. Plant Growth-Promoting Effect on Lupin
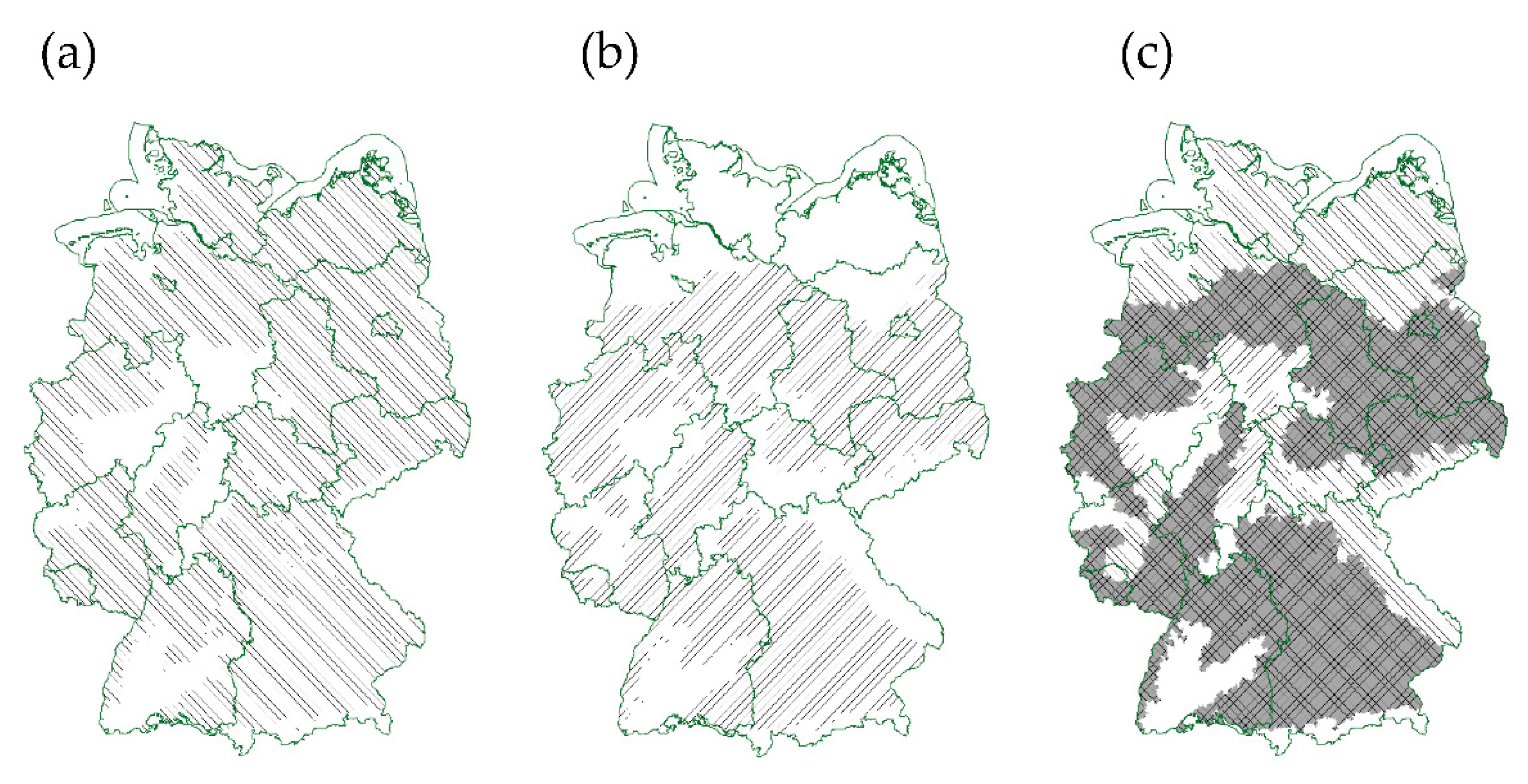
3.4. Comparison of Potential Soybean and Lupin Cultivation Areas in Germany
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lucas, M.M.; Stoddard, F.L.; Annicchiarico, P.; Frías, J.; Martínez-Villaluenga, C.; Sussmann, D.; Duranti, M.; Seger, A.; Zander, P.M.; Pueyo, J.J. The future of lupin as a protein crop in Europe. Front. Plant Sci. 2015, 6, 1–6. [Google Scholar] [CrossRef]
- Talhinhas, P.; Baroncelli, R.; Le Floch, G. Anthracnose of lupins caused by Colletotrichum lupini: A recent disease and a successful worldwide pathogen. J. Plant Pathol. 2016, 98, 5–14. [Google Scholar] [CrossRef]
- Tian, Q.; Lu, C.; Wang, S.; Xiong, Q.; Zhang, H.; Wang, Y.; Zheng, X. Rapid diagnosis of soybean anthracnose caused by Colletotrichum truncatum using a loop-mediated isothermal amplification (LAMP) assay. Eur. J. Plant Pathol. 2017, 148, 785–793. [Google Scholar] [CrossRef]
- Ploper, L.D.; Backman, P.A. Nature and Management of Fungal Diseases Affecting Soybean Stems, Pods, and Seeds. In Pest Management in Soybean; Springer: Dordrecht, The Netherlands, 1992; pp. 174–184. [Google Scholar]
- Murphy-Bokern, D.; Stoddard, F.L.; Watson, C.A. Legumes in cropping systems. Legum. Crop. Syst. 2017, 1–256. [Google Scholar]
- Rogério, F.; Gladieux, P.; Massola, N.S.; Ciampi-Guillardi, M. Multiple Introductions Without Admixture of Colletotrichum truncatum Associated with Soybean Anthracnose in Brazil. Phytopathology 2019, 109, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Barbieri, M.C.G.; Ciampi-Guillardi, M.; Moraes, S.R.G.; Bonaldo, S.M.; Rogério, F.; Linhares, R.R.; Massola, N.S. First Report of Colletotrichum cliviae Causing Anthracnose on Soybean in Brazil. Plant Dis. 2017, 101, 1677. [Google Scholar] [CrossRef]
- Rogério, F.; Ciampi-Guillardi, M.; Barbieri, M.C.G.; Bragança, C.A.D.; Seixas, C.D.S.; Almeida, A.M.R.; Massola, N.S. Phylogeny and variability of Colletotrichum truncatum associated with soybean anthracnose in Brazil. J. Appl. Microbiol. 2017, 122, 402–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, M.D.; Fonseca, M.E.N.; Dias-Neto, J.J.; Santos, M.D.M.; Pandolfo, G.M.; Boiteux, L.S.; Café-Filho, A.C. Biology, pathogenicity, and haplotype analyses of Colletotrichum cliviae: A novel soybean anthracnose agent in warm tropical areas. Trop. Plant Pathol. 2018, 43, 439–451. [Google Scholar] [CrossRef]
- Damm, U.; Sato, T.; Alizadeh, A.; Groenewald, J.Z.; Crous, P.W. The Colletotrichum dracaenophilum, C. magnum and C. orchidearum species complexes. Stud. Mycol. 2019, 92, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Pecchia, S.; Caggiano, B.; Da Lio, D.; Cafà, G.; Le Floch, G.; Baroncelli, R. Molecular Detection of the Seed-Borne Pathogen Colletotrichum lupini Targeting the Hyper-Variable IGS Region of the Ribosomal Cluster. Plants 2019, 8, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nirenberg, H.I.; Feiler, U.; Hagedorn, G. Description of Colletotrichum lupini comb. nov. in modern terms. Mycologia 2002, 94, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Berger, J.D.; Buirchell, B.J.; Luckett, D.J.; Nelson, M.N. Domestication bottlenecks limit genetic diversity and constrain adaptation in narrow-leafed lupin (Lupinus angustifolius L.). Theor. Appl. Genet. 2012, 124, 637–652. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.; Dieterich, R.; Nelson, M.N.; Kamphuis, L.G.; Singh, K.B.; Rotter, B.; Krezdorn, N.; Winter, P.; Wehling, P.; Ruge-Wehling, B. Characterization and mapping of LanrBo: A locus conferring anthracnose resistance in narrow-leafed lupin (Lupinus angustifolius L.). Theor. Appl. Genet. 2015, 128, 2121–2130. [Google Scholar] [CrossRef] [PubMed]
- Jacob, I.; Feuerstein, U.; Heinz, M.; Schott, M.; Urbatzka, P. Evaluation of new breeding lines of white lupin with improved resistance to anthracnose. Euphytica 2017, 213, 1–10. [Google Scholar] [CrossRef]
- Douanla-Meli, C.; Unger, J.-G.; Langer, E. Multi-approach analysis of the diversity in Colletotrichum cliviae sensu lato. Antonie Van Leeuwenhoek 2018, 111, 423–435. [Google Scholar] [CrossRef]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef]
- Scandiani, M.M.; Ruberti, D.S.; Giorda, L.M.; Pioli, R.N.; Luque, A.G.; Bottai, H.; Ivancovich, J.J.; Aoki, T.; O’Donnell, K. Comparison of inoculation methods for characterizing relative aggressiveness of two soybean sudden-death syndrome pathogens, Fusarium virguliforme and F. tucumaniae. Trop. Plant Pathol. 2011, 36, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants. BBCH Monograph; Julius Kühn-Institut (JKI): Quedlinburg, Germany, 2018; ISBN 978-3-95547-071-5. [Google Scholar]
- Dracup, M.; Kirby, E.J.M. Lupin Development Guide; University of Western Australia Press: Perth, Australia, 1996; ISBN 978-1875560660. [Google Scholar]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Taylor, P.W.J. Life styles of Colletotrichum species and implications for plant biosecurity. Fungal Biol. Rev. 2017, 31, 155–168. [Google Scholar] [CrossRef]
- Bechinger, C. Optical Measurements of Invasive Forces Exerted by Appressoria of a Plant Pathogenic Fungus. Science 1999, 285, 1896–1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Küster, S.; Ludwig, N.; Willers, G.; Hoffmann, J.; Deising, H.B.; Kiesow, A. Thin PTFE-like membranes allow characterizing germination and mechanical penetration competence of pathogenic fungi. Acta Biomater. 2008, 4, 1809–1818. [Google Scholar] [CrossRef]
- Ludwig, N.; Loehrer, M.; Hempel, M.; Mathea, S.; Schliebner, I.; Menzel, M.; Kiesow, A.; Schaffrath, U.; Deising, H.B.; Horbach, R. Melanin Is Not Required for Turgor Generation but Enhances Cell-Wall Rigidity in Appressoria of the Corn Pathogen Colletotrichum graminicola. Mol. Plant-Microbe Interact. 2014, 27, 315–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loehrer, M.; Botterweck, J.; Jahnke, J.; Mahlmann, D.M.; Gaetgens, J.; Oldiges, M.; Horbach, R.; Deising, H.; Schaffrath, U. In vivo assessment by Mach-Zehnder double-beam interferometry of the invasive force exerted by the Asian soybean rust fungus (Phakopsora pachyrhizi). New Phytol. 2014, 203, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Hutcheson, S.W. CURRENT CONCEPTS OF ACTIVE DEFENSE IN PLANTS. Annu. Rev. Phytopathol. 1998, 36, 59–90. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.F.; Crous, P.W. Colletotrichum: Complex Species or Species Complexes? CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2012; Volume 73, ISBN 9789070351922. [Google Scholar]
- Hacquard, S.; Kracher, B.; Hiruma, K.; Münch, P.C.; Garrido-Oter, R.; Thon, M.R.; Weimann, A.; Damm, U.; Dallery, J.-F.; Hainaut, M.; et al. Survival trade-offs in plant roots during colonization by closely related beneficial and pathogenic fungi. Nat. Commun. 2016, 7, 11362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiruma, K.; Gerlach, N.; Sacristán, S.; Nakano, R.T.; Hacquard, S.; Kracher, B.; Neumann, U.; Ramírez, D.; Bucher, M.; O’Connell, R.J.; et al. Root Endophyte Colletotrichum tofieldiae Confers Plant Fitness Benefits that Are Phosphate Status Dependent. Cell 2016, 165, 464–474. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, S.; Huckelhoven, R.; Schafer, P.; Imani, J.; Sharma, M.; Weiss, M.; Waller, F.; Kogel, K.-H. The root endophytic fungus Piriformospora indica requires host cell death for proliferation during mutualistic symbiosis with barley. Proc. Natl. Acad. Sci. USA 2006, 103, 18450–18457. [Google Scholar] [CrossRef] [Green Version]
- Zuccaro, A.; Lahrmann, U.; Güldener, U.; Langen, G.; Pfiffi, S.; Biedenkopf, D.; Wong, P.; Samans, B.; Grimm, C.; Basiewicz, M.; et al. Endophytic life strategies decoded by genome and transcriptome analyses of the mutualistic root symbiont Piriformospora indica. PLoS Pathog. 2011, 7, e1002290. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | ID in Manuscript | Origin | Literature |
|---|---|---|---|---|
| Colletotrichum lupini | BBA70358 (= CBS109222) | Cl70358 | lupin | [12,14] |
| C. truncatum | LFN0389 | Ct0389 | soybean | [8] |
| C. truncatum | LFN0390 | Ct0390 | soybean | [8] |
| C. truncatum | LFN0391 | Ct0391 | soybean | [8] |
| C. truncatum | LFN0392 | Ct0392 | soybean | [8] |
| C. truncatum | LFN0393 | Ct0393 | soybean | [8] |
| C. plurivorum | LFN0007 | Cp0007 | soybean | [8,10] |
| C. plurivorum | LFN0008 | Cp0008 | soybean | [8,9,10,16] |
| C. plurivorum | LFN0010 (= URM7540) | Cp0010 | soybean | [7,8,16] |
| C. plurivorum | LFN0018 (= URM7541) | Cp0018 | soybean | [7,8,16] |
| C. plurivorum | LFN0019 (= URM7542) | Cp0019 | soybean | [7,8,16] |
| C. plurivorum | LFN0023 | Cp0023 | soybean | [16] |
| C. higginsianum | IMI349063 | Ch349063 | Arabidopsis | [17] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wirtz, L.; Massola Júnior, N.S.; Castro, R.R.L.d.; Ruge-Wehling, B.; Schaffrath, U.; Loehrer, M. Colletotrichum spp. from Soybean Cause Disease on Lupin and Can Induce Plant Growth-Promoting Effects. Microorganisms 2021, 9, 1130. https://doi.org/10.3390/microorganisms9061130
Wirtz L, Massola Júnior NS, Castro RRLd, Ruge-Wehling B, Schaffrath U, Loehrer M. Colletotrichum spp. from Soybean Cause Disease on Lupin and Can Induce Plant Growth-Promoting Effects. Microorganisms. 2021; 9(6):1130. https://doi.org/10.3390/microorganisms9061130
Chicago/Turabian StyleWirtz, Louisa, Nelson Sidnei Massola Júnior, Renata Rebellato Linhares de Castro, Brigitte Ruge-Wehling, Ulrich Schaffrath, and Marco Loehrer. 2021. "Colletotrichum spp. from Soybean Cause Disease on Lupin and Can Induce Plant Growth-Promoting Effects" Microorganisms 9, no. 6: 1130. https://doi.org/10.3390/microorganisms9061130