Post-Vaccination Streptococcus pneumoniae Carriage and Virulence Gene Distribution among Children Less Than Five Years of Age, Cape Coast, Ghana

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Study the Site
2.2. Study Participants
2.3. Sample Collection and Transport
2.4. Pneumococcal Isolation
2.5. Antibiotic Susceptibility Testing
2.6. Characterization of S. pneumoniae Isolates
2.6.1. Serotyping
2.6.2. Determination of Virulence Genes
2.7. Statistical Analysis
2.8. Ethical Approval
3. Results
3.1. Characteristics of the Study Participants
3.2. Pneumococcal Carriage in Children
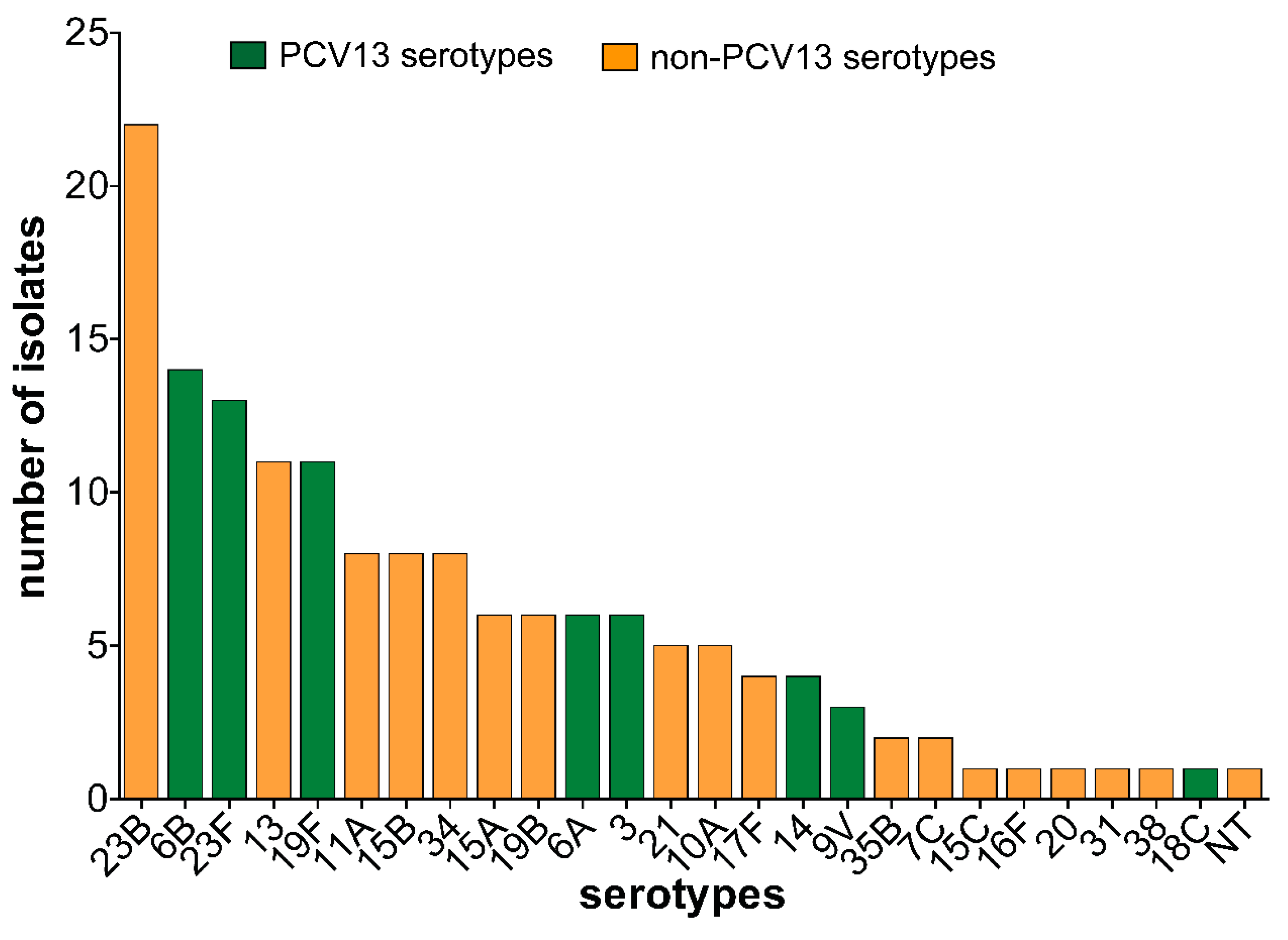
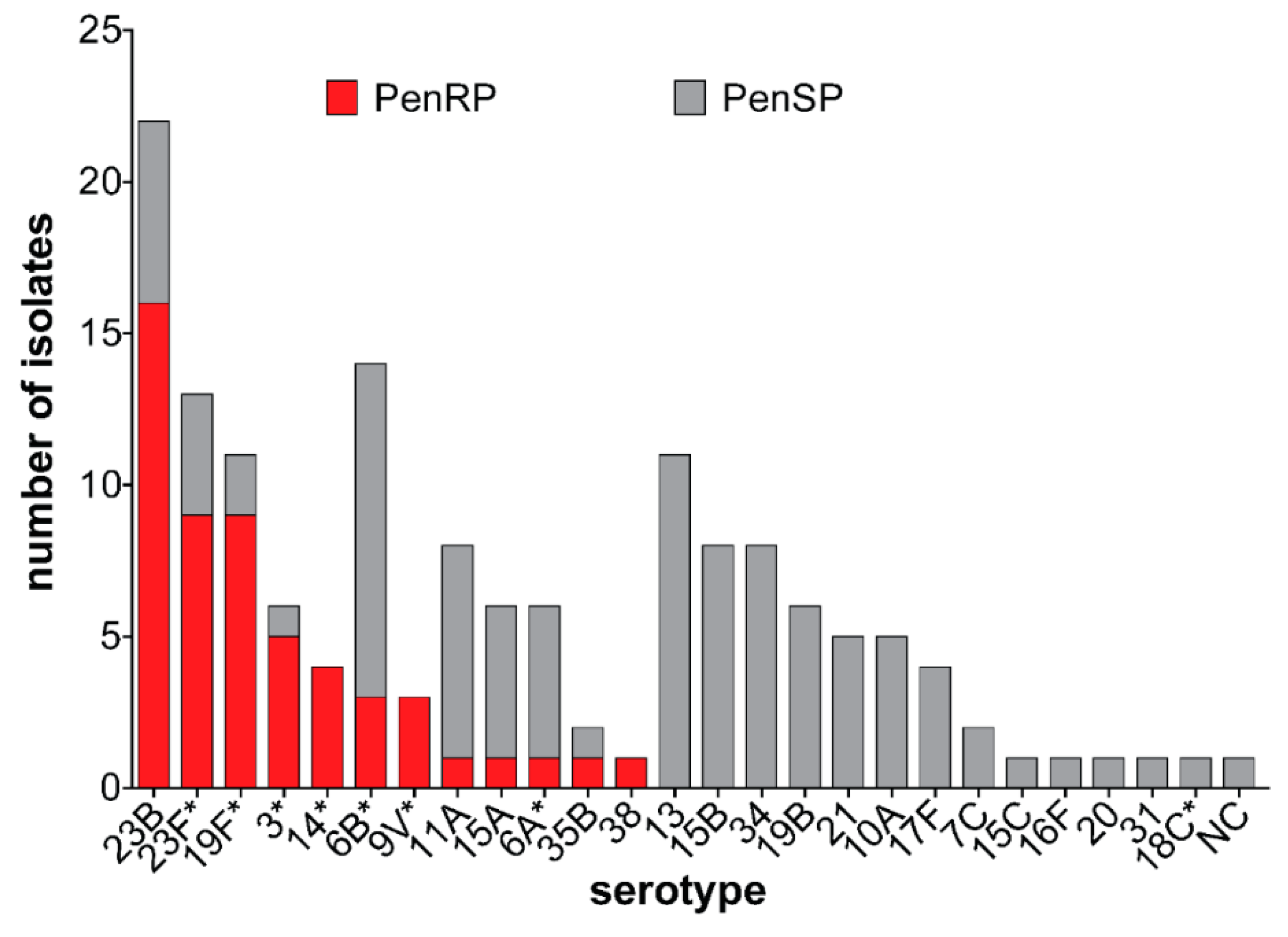
3.3. Pneumococcal Serotypes and PCV13 Serotypes Identified in Carriage Isolates
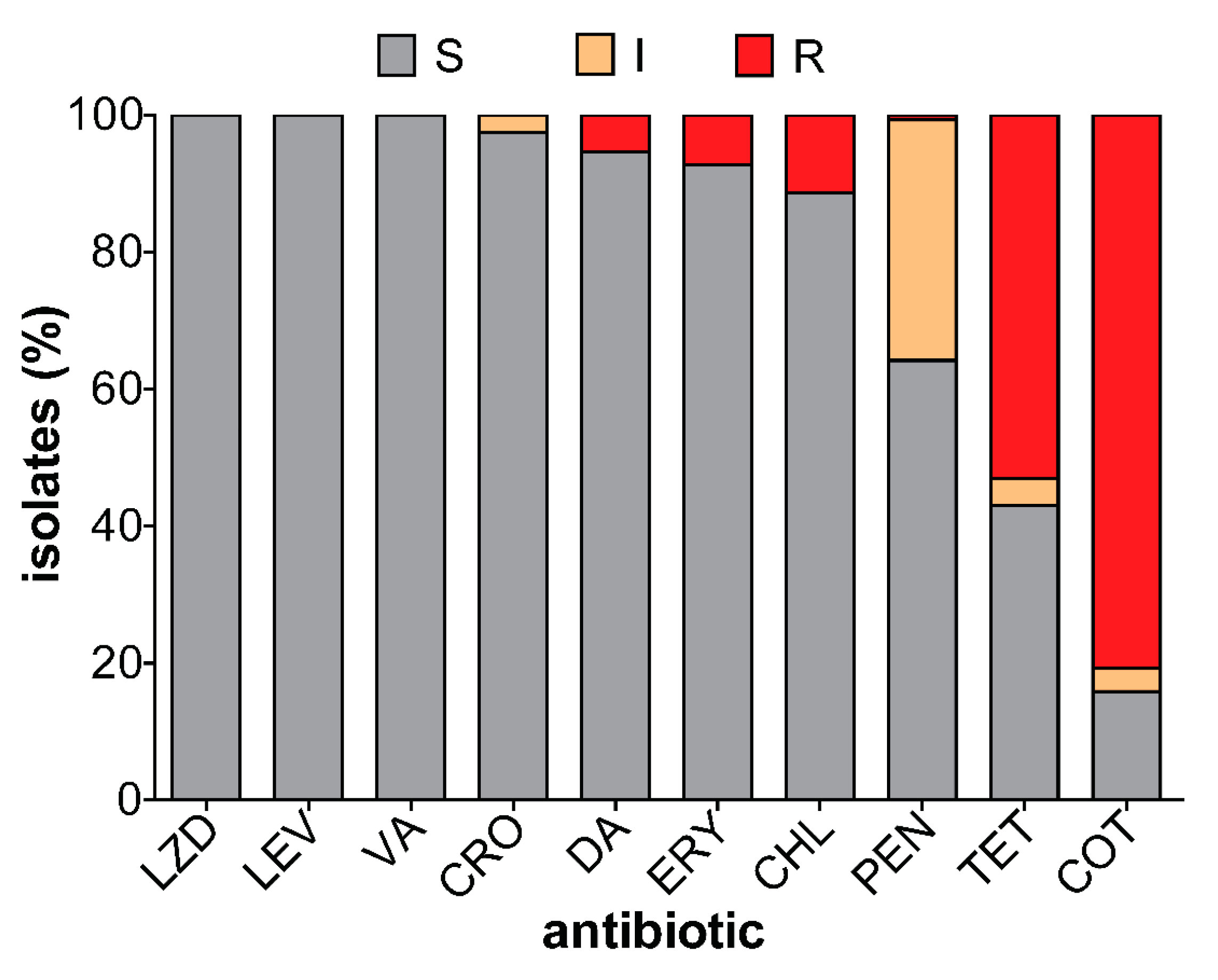
3.4. Antibiotic Susceptibility of Carriage Isolates
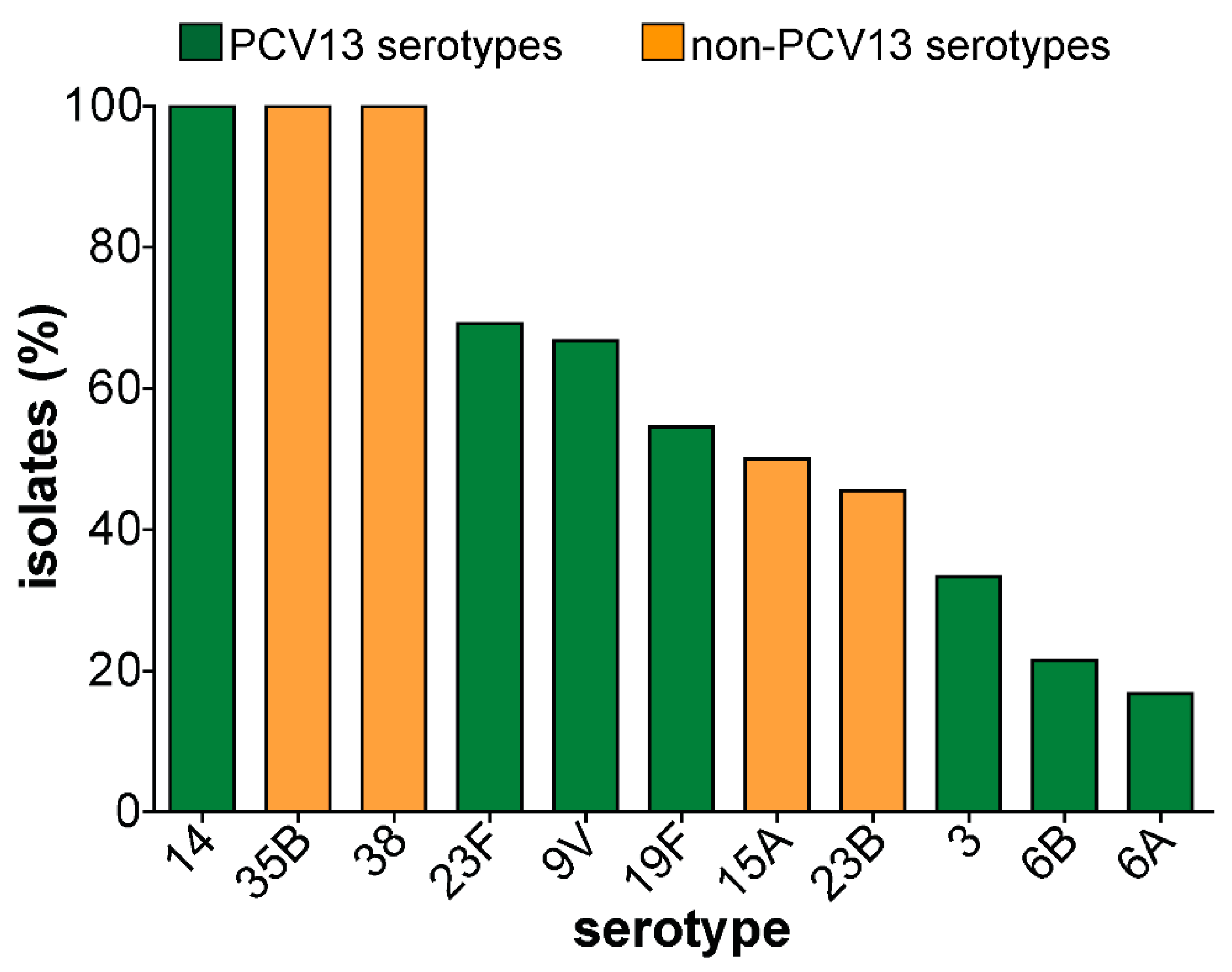
3.5. Virulence Genes Detected in Carriage Isolates
4. Discussion
4.1. Serotype Distribution
4.2. Antibiotic Resistance
4.3. Virulence Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wahl, B.; O’Brien, K.L.; Greenbaum, A.; Majumder, A.; Liu, L.; Chu, Y.; Lukšić, I.; Nair, H.; McAllister, D.A.; Campbell, H.; et al. Burden of Streptococcus pneumoniae and Haemophilus influenzae type b disease in children in the era of conjugate vaccines: Global, regional, and national estimates for 2000–15. Lancet Glob. Heath 2018, 6, e744–e757. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, S.L.; Barson, W.J.; Ling, L.P.; Romero, J.R.; Bradley, J.S.; Tan, T.Q.; Pannaraj, P.S.; Givner, L.B.; Hulten, K.G. Invasive Pneumococcal Disease in Children’s Hospitals: 2014–2017. Pediatrics 2019, 144, e20190567. [Google Scholar] [CrossRef] [PubMed]
- Weiser, J.N.; Ferreira, D.M.; Paton, J.C. Streptococcus pneumoniae: Transmission, colonization and invasion. Nat. Rev. Genet. 2018, 16, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.; Nguyen, C.D.; Dunne, E.M.; Mulholland, E.K.; Mungun, T.; Pomat, W.S.; Rafai, E.; Satzke, C.; Weinberger, D.M.; Russell, F.M. Using pneumococcal carriage studies to monitor vaccine impact in low- and middle-income countries. Vaccine 2019, 37, 6299–6309. [Google Scholar] [CrossRef]
- Weinberger, D.M.; Bruden, D.T.; Grant, L.R.; Lipsitch, M.; O’Brien, K.L.; Pelton, S.I.; Sanders, E.A.M.; Feikin, D.R. Using Pneumococcal Carriage Data to Monitor Postvaccination Changes in Invasive Disease. Am. J. Epidemiol. 2013, 178, 1488–1495. [Google Scholar] [CrossRef] [Green Version]
- Swarthout, T.D.; Fronterre, C.; Lourenço, J.; Obolski, U.; Gori, A.; Bar-Zeev, N.; Everett, D.; Kamng’Ona, A.W.; Mwalukomo, T.S.; Mataya, A.A.; et al. High residual carriage of vaccine-serotype Streptococcus pneumoniae after introduction of pneumococcal conjugate vaccine in Malawi. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Renner, L.A.; Usuf, E.; Mohammed, N.I.; Ansong, D.; Dankwah, T.; Kusah, J.T.; Owusu, S.K.; Awunyo, M.; Arhin, B.; Addo, Y.; et al. Hospital-based Surveillance for Pediatric Bacterial Meningitis in the Era of the 13-Valent Pneumococcal Conjugate Vaccine in Ghana. Clin. Infect. Dis. 2019, 69, S89–S96. [Google Scholar] [CrossRef]
- Sigurdsson, S.; Erlendsdóttir, H.; Quirk, S.J.; Kristjánsson, J.; Hauksson, K.; Andrésdóttir, B.D.I.; Jónsson, A.J.; Halldórsson, K.H.; Sæmundsson, Á.; Ólason, Ó.H.; et al. Pneumococcal vaccination: Direct and herd effect on carriage of vaccine types and antibiotic resistance in Icelandic children. Vaccine 2017, 35, 5242–5248. [Google Scholar] [CrossRef]
- WHO. Who Vaccine-Preventable Diseases: Monitoring System. 2020 Global Summary. Available online: https://apps.who.int/immunization_monitoring/globalsummary (accessed on 29 July 2020).
- Mills, R.O.; Twum-Danso, K.; Owusu-Agyei, S.; Donkor, E.S. Epidemiology of pneumococcal carriage in children under five years of age in Accra, Ghana. Infect. Dis. 2015, 47, 326–331. [Google Scholar] [CrossRef]
- Dayie, N.T.K.D.; Arhin, R.E.; Newman, M.J.; Dalsgaard, A.; Bisgaard, M.; Frimodt-Møller, N.; Slotved, H.-C. Penicillin resistance and serotype distribution of Streptococcus pneumoniae in Ghanaian children less than six years of age. BMC Infect. Dis. 2013, 13, 490. [Google Scholar] [CrossRef] [Green Version]
- Negash, A.A.; Asrat, D.; Abebe, W.; Hailemariam, T.; Gebre, M.; Verhaegen, J.; Aseffa, A.; Vaneechoutte, M. Pneumococcal Carriage, Serotype Distribution, and Risk Factors in Children With Community-Acquired Pneumonia, 5 Years After Introduction of the 10-Valent Pneumococcal Conjugate Vaccine in Ethiopia. Open Forum Infect. Dis. 2019, 6, 259. [Google Scholar] [CrossRef] [PubMed]
- Sidorenko, S.; Rennert, W.; Lobzin, Y.; Briko, N.; Kozlov, R.; Namazova-Baranova, L.S.; Tsvetkova, I.; Ageevets, V.; Nikitina, E.; Ardysheva, A.; et al. Multicenter study of serotype distribution of Streptococcus pneumoniae nasopharyngeal isolates from healthy children in the Russian Federation after introduction of PCV13 into the National Vaccination Calendar. Diagn. Microbiol. Infect. Dis. 2020, 96, 114914. [Google Scholar] [CrossRef] [PubMed]
- Odutola, A.; Ota, M.O.C.; Antonio, M.; Ogundare, E.O.; Saidu, Y.; Owiafe, P.K.; Worwui, A.; Idoko, O.T.; Owolabi, O.; Kampmann, B.; et al. Immunogenicity of Pneumococcal Conjugate Vaccine Formulations Containing Pneumococcal Proteins, and Immunogenicity and Reactogenicity of Co-Administered Routine Vaccines—A Phase II, Randomised, Observer-Blind Study in Gambian Infants. Vaccine 2019, 37, 2586–2599. [Google Scholar] [CrossRef] [PubMed]
- Rose, L.; Shivshankar, P.; Hinojosa, E.; Rodriguez, A.; Sanchez, C.J.; Orihuela, C.J. Antibodies against PsrP, a NovelStreptococcus pneumoniaeAdhesin, Block Adhesion and Protect Mice against Pneumococcal Challenge. J. Infect. Dis. 2008, 198, 375–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornick, J.E.; Bishop, Ö.T.; Yalcin, F.; Kiran, A.M.; Kumwenda, B.; Chaguza, C.; Govindpershad, S.; Ousmane, S.; Senghore, M.; Du Plessis, M.; et al. The global distribution and diversity of protein vaccine candidate antigens in the highly virulent Streptococcus pnuemoniae serotype 1. Vaccine 2017, 35, 972–980. [Google Scholar]
- Selva, L.; Ciruela, P.; Blanchette, K.; Del Amo, E.; Pallares, R.; Orihuela, C.J.; Muñoz-Almagro, C. Prevalence and Clonal Distribution of pcpA, psrP and Pilus-1 among Pediatric Isolates of Streptococcus pneumoniae. PLoS ONE 2012, 7, e41587. [Google Scholar] [CrossRef] [Green Version]
- Hilleringmann, M.; Kohler, S.; Gàmez, G.; Hammerschmidt, S. Pneumococcal Pili and Adhesins. In Streptococcus Pneumoniae: Molecular Mechanisms of Host-Pathogen Interactions; Hammerschmidt, J., Brown, S., Orihuela, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Kasher, M.; Roizin, H.; Cohen, A.; Jaber, H.; Mikhailov, S.; Rubin, C.; Doron, D.; Rahav, G.; Regev-Yochay, G. The Impact of Pcv7/13 on the Distribution of Carried Pneumococcal Serotypes and on Pilus Prevalence; 14 Years of Repeated Cross-Sectional Surveillance. Vaccine 2020, 38, 3591–3599. [Google Scholar] [CrossRef]
- Dayie, N.T.K.D.; Tettey, E.Y.; Newman, M.J.; Bannerman, E.; Donkor, E.S.; Labi, A.-K.; Slotved, H.-C. Pneumococcal carriage among children under five in Accra, Ghana, five years after the introduction of pneumococcal conjugate vaccine. BMC Pediatr. 2019, 19, 316. [Google Scholar] [CrossRef]
- Ghana Districts. Available online: http://www.ghanadistricts.com (accessed on 11 August 2020).
- Satzke, C.; Turner, P.; Virolainen-Julkunen, A.; Adrian, P.V.; Antonio, M.; Hare, K.M.; Henao-Restrepo, A.M.; Leach, A.J.; Klugman, K.P.; Porter, B.D.; et al. Pneumococcal Carriage Working Group. Standard Method for Detecting Upper Respiratory Carriage of Streptococcus Pneumoniae: Updated Recommendations from the World Health Organization Pneumococcal Carriage Working Group. Vaccine 2013, 32, 165–179. [Google Scholar] [CrossRef]
- CLSI. M100 Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017; Volume 37. [Google Scholar]
- CDC. Protocol for Multiplex Pcr–S. Pneumoniae Serotyping–Clinical Specimens and Pneumococcal Isolates–African Set. Available online: www.cdc.gov/streplab/pneumococcus/resources.html (accessed on 18 March 2018).
- Jacobs, M.R.; Bajaksouzian, S.; Bonomo, R.A.; Good, C.E.; Windau, A.R.; Hujer, A.M.; Massire, C.; Melton, R.; Blyn, L.B.; Ecker, D.J.; et al. Occurrence, Distribution, and Origins of Streptococcus pneumoniae Serotype 6C, a Recently Recognized Serotype. J. Clin. Microbiol. 2008, 47, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Hadjipanayis, A.; Efstathiou, E.; Alexandrou, M.; Panayiotou, L.; Zachariadou, C.; Petrou, P.; Papaevangelou, V. Nasopharyngeal Pneumococcal Carriage among Healthy Children in Cyprus Post Widespread Simultaneous Implementation of PCV10 and PCV13 Vaccines. PLoS ONE 2016, 11, e0163269. [Google Scholar] [CrossRef] [PubMed]
- Lindstrand, A.; Galanis, I.; Darenberg, J.; Morfeldt, E.; Naucler, P.; Blennow, M.; Alfvén, T.; Henriques-Normark, B.; Örtqvist, Å. Unaltered pneumococcal carriage prevalence due to expansion of non-vaccine types of low invasive potential 8 years after vaccine introduction in Stockholm, Sweden. Vaccine 2016, 34, 4565–4571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emgård, M.; Msuya, S.E.; Nyombi, B.M.; Mosha, D.; Gonzales-Siles, L.; Nordén, R.; Geravandi, S.; Mosha, V.; Blomqvist, J.; Franzén, S.; et al. Carriage of penicillin-non-susceptible pneumococci among children in northern Tanzania in the 13-valent pneumococcal vaccine era. Int. J. Infect. Dis. 2019, 81, 156–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanik-Yuksek, S.; Tezer, H.; Gülhan, B.; Ozkaya-Parlakay, A.; Güldemir, D.; Coskun-Ari, F.F.; Demirdag, T.B.; Uzun, A.K.; Kızılgün, M.; Solmaz, S.; et al. Nasopharyngeal pneumococcal carriage in healthy Turkish children after 13-valent conjugated pneumococcal vaccine implementation in the national immunization program. J. Infect. Public Health 2020, 13, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Spijkerman, J.; van Gils, E.J.M.; Veenhoven, R.H.; Hak, E.; Yzerman, F.; van der Ende, A.; Wijmenga-Monsuur, A.J.; van den Dobbelsteen, G.P.J.M.; Sanders, E.A.M. Carriage Ofstreptococcus Pneumoniae3 Years after Start of Vaccination Program, the Netherlands. Emerg. Infect. Dis. 2011, 17, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Adetifa, I.M.O.; Adamu, A.L.; Karani, A.; Waithaka, M.; Odeyemi, K.A.; Okoromah, C.A.N.; Bello, M.M.; Abubakar, I.S.; Inem, V.; Scott, J.A.G. Nasopharyngeal Pneumococcal Carriage in Nigeria: A two-site, population-based survey. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Løvlie, A.; Vestrheim, D.F.; Aaberge, I.S.; Steens, A. Changes in pneumococcal carriage prevalence and factors associated with carriage in Norwegian children, four years after introduction of PCV13. BMC Infect. Dis. 2020, 20, 29. [Google Scholar] [CrossRef]
- Satzke, C.; Dunne, E.M.; Choummanivong, M.; Ortika, B.D.; Neal, E.F.; Pell, C.L.; Nation, M.L.; Fox, K.K.; Nguyen, C.D.; Gould, K.A.; et al. Pneumococcal carriage in vaccine-eligible children and unvaccinated infants in Lao PDR two years following the introduction of the 13-valent pneumococcal conjugate vaccine. Vaccine 2019, 37, 296–305. [Google Scholar] [CrossRef]
- Collins, D.A.; Hoskins, A.; Snelling, T.; Senasinghe, K.; Bowman, J.; Stemberger, N.A.; Leach, A.J.; Lehmann, D. Predictors of pneumococcal carriage and the effect of the 13-valent pneumococcal conjugate vaccination in the Western Australian Aboriginal population. Pneumonia 2017, 9, 14. [Google Scholar] [CrossRef] [Green Version]
- Fjeldhøj, S.; Laursen, R.P.; Larnkjær, A.; Mølgaard, C.; Fuursted, K.; Krogfelt, K.A.; Slotved, H.-C. Probiotics and carriage of Streptococcus pneumoniae serotypes in Danish children, a double-blind randomized controlled trial. Sci. Rep. 2018, 8, 15258. [Google Scholar] [CrossRef]
- Roca, A.; Bojang, A.; Bottomley, C.; Gladstone, R.A.; Adetifa, J.U.; Egere, U.; Burr, S.; Antonio, M.; Bentley, S.D.; Kampmann, B.; et al. Effect on nasopharyngeal pneumococcal carriage of replacing PCV7 with PCV13 in the Expanded Programme of Immunization in The Gambia. Vaccine 2015, 33, 7144–7151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, H.L.; Deloria-Knoll, M.; Levine, O.S.; Stoszek, S.K.; Hance, L.F.; Reithinger, R.; Muenz, L.R.; O’Brien, K.L. Systematic Evaluation of Serotypes Causing Invasive Pneumococcal Disease among Children Under Five: The Pneumococcal Global Serotype Project. PLoS Med. 2010, 7, e1000348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricketson, L.J.; Wood, M.L.; Vanderkooi, O.G.; Macdonald, J.; Martin, I.E.; Demczuk, W.; Kellner, J.D. Trends in Asymptomatic Nasopharyngeal Colonization With Streptococcus pneumoniae After Introduction of the 13-valent Pneumococcal Conjugate Vaccine in Calgary, Canada. Pediatr. Infect. Dis. J. 2014, 33, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Van Der Linden, M.; Perniciaro, S.; Imöhl, M. Increase of serotypes 15A and 23B in IPD in Germany in the PCV13 vaccination era. BMC Infect. Dis. 2015, 15, 207. [Google Scholar] [CrossRef] [Green Version]
- Kwambana-Adams, B.A.; Asiedu-Bekoe, F.; Sarkodie, B.; Afreh, O.K.; Kuma, G.K.; Owusu-Okyere, G.; Foster-Nyarko, E.; Ohene, S.-A.; Okot, C.; Worwui, A.K.; et al. An outbreak of pneumococcal meningitis among older children (≥5 years) and adults after the implementation of an infant vaccination programme with the 13-valent pneumococcal conjugate vaccine in Ghana. BMC Infect. Dis. 2016, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bozio, C.H.; Abdul-Karim, A.; Abenyeri, J.; Abubakari, B.; Ofosu, W.; Zoya, J.; Ouattara, M.; Srinivasan, V.; Vuong, J.T.; Opare, D.; et al. Continued occurrence of serotype 1 pneumococcal meningitis in two regions located in the meningitis belt in Ghana five years after introduction of 13-valent pneumococcal conjugate vaccine. PLoS ONE 2018, 13, e0203205. [Google Scholar] [CrossRef]
- Dayie, N.T.K.D.; Tetteh-Ocloo, G.; Labi, A.-K.; Olayemi, E.; Slotved, H.-C.; Lartey, M.; Donkor, E.S. Pneumococcal carriage among sickle cell disease patients in Accra, Ghana: Risk factors, serotypes and antibiotic resistance. PLoS ONE 2018, 13, e0206728. [Google Scholar] [CrossRef]
- Dayie, N.T.; Baffuor-Asare, M.; Labi, A.-K.; Obeng-Nkrumah, N.; Olayemi, E.; Lartey, M.; Slotved, H.-C.; Donkor, E.S. Epidemiology of Pneumococcal Carriage among HIV-Infected Individuals in the Conjugate Vaccine Era: A Study in Southern Ghana. BioMed Res. Int. 2019, 2019, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Heinsbroek, E.; Tafatatha, T.; Phiri, A.; Swarthout, T.D.; Alaerts, M.; Crampin, A.C.; Chisambo, C.; Mwiba, O.; Read, J.M.; French, N. Pneumococcal carriage in households in Karonga District, Malawi, before and after introduction of 13-valent pneumococcal conjugate vaccination. Vaccine 2018, 36, 7369–7376. [Google Scholar] [CrossRef]
- Usuf, E.; Bottomley, C.; Bojang, E.; Cox, I.; Bojang, A.; Gladstone, R.; Kampmann, B.; Hill, P.C.; Roca, A. Persistence of Nasopharyngeal Pneumococcal Vaccine Serotypes and Increase of Nonvaccine Serotypes Among Vaccinated Infants and Their Mothers 5 Years After Introduction of Pneumococcal Conjugate Vaccine 13 in The Gambia. Clin. Infect. Dis. 2019, 68, 1512–1521. [Google Scholar] [CrossRef]
- Jayasinghe, S.; Chiu, C.; Quinn, H.; Menzies, R.; Gilmour, R.; McIntyre, P. Effectiveness of 7- and 13-Valent Pneumococcal Conjugate Vaccines in a Schedule Without a Booster Dose: A 10-Year Observational Study. Clin. Infect. Dis. 2018, 67, 367–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, M.; Conklin, L.M.; Bigogo, G.; Jagero, G.; Hampton, L.M.; Fleming-Dutra, K.E.; Junghae, M.; Carvalho, M.D.G.S.; Pimenta, F.; Beall, B.; et al. Pneumococcal Carriage and Antibiotic Susceptibility Patterns from Two Cross-Sectional Colonization Surveys among Children Aged <5 Years Prior to the Introduction of 10-Valent Pneumococcal Conjugate Vaccine—Kenya, 2009–2010. BMC Infect. Dis. 2017, 17, 25. [Google Scholar]
- Jensen, V.V.S.; Furberg, A.-S.; Slotved, H.-C.; Bazhukova, T.; Haldorsen, B.; Caugant, D.A.; Sundsfjord, A.; Valentiner-Branth, P.; Simonsen, G.S. Epidemiological and molecular characterization of Streptococcus pneumoniae carriage strains in pre-school children in Arkhangelsk, northern European Russia, prior to the introduction of conjugate pneumococcal vaccines. BMC Infect. Dis. 2020, 20, 1–14. [Google Scholar]
- Dayie, N.T.K.D.; Arhin, R.E.; Newman, M.J.; Dalsgaard, A.; Bisgaard, M.; Frimodt-Møller, N.; Slotved, H.-C. Multidrug-Resistant Streptococcus pneumoniae Isolates from Healthy Ghanaian Preschool Children. Microb. Drug Resist. 2015, 21, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Labi, A.-K.; Obeng-Nkrumah, N.; Nartey, E.T.; Bjerrum, S.; Adu-Aryee, N.A.; Ofori-Adjei, Y.A.; Yawson, A.E.; Newman, M.J. Antibiotic use in a tertiary healthcare facility in Ghana: A point prevalence survey. Antimicrob. Resist. Infect. Control. 2018, 7, 15. [Google Scholar] [CrossRef]
- Kawaguchiya, M.; Urushibara, N.; Aung, M.S.; Shinagawa, M.; Takahashi, S.; Kobayashi, N. Serotype distribution, antimicrobial resistance and prevalence of pilus islets in pneumococci following the use of conjugate vaccines. J. Med. Microbiol. 2017, 66, 643–650. [Google Scholar] [CrossRef]
- Adetifa, I.M.O.; Antonio, M.; Okoromah, C.A.N.; Ebruke, C.; Inem, V.; Nsekpong, D.; Bojang, A.; Adegbola, R.A. Pre-Vaccination Nasopharyngeal Pneumococcal Carriage in a Nigerian Population: Epidemiology and Population Biology. PLoS ONE 2012, 7, e30548. [Google Scholar] [CrossRef]
- Afari-Asiedu, S.; Kinsman, J.; Boamah, E.A.; Abdulai, M.A.; Gyapong, M.; Sankoh, O.; Hulscher, M.E.J.L.; Asante, K.P.; Wertheim, H. To sell or not to sell; the differences between regulatory and community demands regarding access to antibiotics in rural Ghana. J. Pharm. Policy Pr. 2018, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mbeye, N.M.; Ter Kuile, F.; Davies, M.-A.; Phiri, K.; Egger, M.; Wandeler, G.; Africa, I.-S. Cotrimoxazole prophylactic treatment prevents malaria in children in sub-Saharan Africa: Systematic review and meta-analysis. Trop. Med. Int. Health 2014, 19, 1057–1067. [Google Scholar] [CrossRef] [Green Version]
- Mwenya, D.M.; Charalambous, B.M.; Phillips, P.P.J.; Mwansa, J.C.L.; Batt, S.L.; Nunn, A.J.; Walker, S.; Gibb, D.M.; Gillespie, S.H. Impact of Cotrimoxazole on Carriage and Antibiotic Resistance of Streptococcus pneumoniae and Haemophilus influenzae in HIV-Infected Children in Zambia. Antimicrob. Agents Chemother. 2010, 54, 3756–3762. [Google Scholar] [CrossRef] [Green Version]
- Oppong, F.B.; Gyaase, S.; Zandoh, C.; Nettey, O.E.A.; Amenga-Etego, S.; Anane, E.A.; Adda, R.; Dosoo, D.K.; Owusu-Agyei, S.; Asante, K.P. Intermittent preventive treatment of pregnant women in Kintampo area of Ghana with sulphadoxine-pyrimethamine (SP): Trends spanning 2011 and 2015. BMJ Open 2019, 9, e027946. [Google Scholar] [CrossRef]
- Feikin, D.R.; Dowell, S.F.; Nwanyanwu, O.C.; Klugman, K.P.; Kazembe, P.N.; Barat, L.M.; Graf, C.; Bloland, P.B.; Ziba, C.; Huebner, R.E.; et al. Increased Carriage of Trimethoprim/Sulfamethoxazole-Resistantstreptococcus Pneumoniaein Malawian Children after Treatment for Malaria with Sulfadoxine/Pyrimethamine. J. Infect. Dis. 2000, 181, 1501–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sader, H.S.; Mendes, R.E.; Le, J.; Denys, G.; Flamm, R.K.; Jones, R.N. Antimicrobial Susceptibility of Streptococcus pneumoniae from North America, Europe, Latin America, and the Asia-Pacific Region: Results From 20 Years of the SENTRY Antimicrobial Surveillance Program (1997–2016). Open Forum Infect. Dis. 2019, 6, S14–S23. [Google Scholar] [CrossRef] [PubMed]
- Afari-Asiedu, S.; Hulscher, M.; Abdulai, M.A.; Boamah-Kaali, E.; Asante, K.P.; Wertheim, H.F.L. Every medicine is medicine; exploring inappropriate antibiotic use at the community level in rural Ghana. BMC Public Health 2020, 20, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Birindwa, A.M.; Emgård, M.; Nordén, R.; Samuelsson, E.; Geravandi, S.; Gonzales-Siles, L.; Muhigirwa, B.; Kashosi, T.; Munguakonkwa, E.; Manegabe, J.T.; et al. High rate of antibiotic resistance among pneumococci carried by healthy children in the eastern part of the Democratic Republic of the Congo. BMC Pediatr. 2018, 18, 361. [Google Scholar] [CrossRef] [PubMed]
- Prah, J.; Kizzie-Hayford, J.; Walker, E.; Ampofo-Asiama, A. Antibiotic prescription pattern in a Ghanaian primary health care facility. Pan. Afr. Med. J. 2017, 28. [Google Scholar] [CrossRef]
- Abdulai, A.A.; Agana-Nsiire, P.; Biney, F.; Kwakye-Maclean, C.; Kyei-Faried, S.; Amponsa-Achiano, K.; Simpson, S.V.; Bonsu, G.; Ohene, S.-A.; Ampofo, W.K.; et al. Community-based mass treatment with azithromycin for the elimination of yaws in Ghana—Results of a pilot study. PLoS Negl. Trop. Dis. 2018, 12, e0006303. [Google Scholar] [CrossRef] [Green Version]
- Hema-Ouangraoua, S.; Maiga, A.A.; Cairns, M.; Zongo, I.; Frédéric, N.; Yerbanga, R.S.; Tamboura, B.; Badji, H.; Gore-Langton, G.; Kuepfer, I.; et al. Impact of the addition of azithromycin to antimalarials used for seasonal malaria chemoprevention on antimicrobial resistance of Streptococcus pneumoniae. Trop. Med. Int. Health 2019, 24, 1442–1454. [Google Scholar] [CrossRef]
- Kaur, R.; Pham, M.; Yu, K.O.A.; Pichichero, M.E. Rising Pneumococcal Antibiotic Resistance in the Post 13-Valent Pneumococcal Conjugate Vaccine Era in Pediatric Isolates From a Primary Care Setting. Clin. Infect. Dis. 2020, 157, 1–9. [Google Scholar] [CrossRef]
- Neves, F.P.G.; Cardoso, N.T.; Cardoso, C.A.; Teixeira, L.M.; Riley, L.W. Direct effect of the 13-valent pneumococcal conjugate vaccine use on pneumococcal colonization among children in Brazil. Vaccine 2019, 37, 5265–5269. [Google Scholar] [CrossRef]
- Dewe, T.C.M.; D’Aeth, J.C.; Croucher, N.J. Genomic Epidemiology of Penicillin-Non-Susceptible Streptococcus Pneumoniae. Microb. Genom 2019, 5, e000305. [Google Scholar] [CrossRef]
- Voß, F.; Kohler, T.P.; Meyer, T.; Abdullah, M.R.; Van Opzeeland, F.J.; Saleh, M.; Michalik, S.; Van Selm, S.; Schmidt, F.; De Jonge, M.I.; et al. Intranasal Vaccination With Lipoproteins Confers Protection Against Pneumococcal Colonisation. Front. Immunol. 2018, 9, 2405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, F.; Talekar, S.J.; Klugman, K.P.; Vidal, J.E. Expression of Streptococcus pneumoniae Virulence-Related Genes in the Nasopharynx of Healthy Children. PLoS ONE 2013, 8, e67147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blumental, S.; Granger-Farbos, A.A.; Moïsi, J.J.; Soullié, B.B.; Leroy, P.P.; Njanpop-Lafourcade, B.M.B.; Yaro, S.S.; Nacro, B.; Hallin, M.; Koeck, J.L.J. Virulence Factors of Streptococcus pneumoniae. Comparison between African and French Invasive Isolates and Implication for Future Vaccines. PLoS ONE 2015, 10, e0133885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz-Almagro, C.; Selva, L.; Sanchez, C.J.; Esteva, C.; De Sevilla, M.F.; Pallares, R.; Orihuela, C.J. PsrP, a Protective Pneumococcal Antigen, Is Highly Prevalent in Children with Pneumonia and Is Strongly Associated with Clonal Type. Clin. Vaccine Immunol. 2010, 17, 1672–1678. [Google Scholar] [CrossRef] [Green Version]
- Basset, A.; Trzcinski, K.; Hermos, C.; O’Brien, K.L.; Reid, R.; Santosham, M.; McAdam, A.J.; Lipsitch, M.; Malley, R. Association of the Pneumococcal Pilus with Certain Capsular Serotypes but Not with Increased Virulence. J. Clin. Microbiol. 2007, 45, 1684–1689. [Google Scholar] [CrossRef] [Green Version]
- Knupp-Pereira, P.A.; Marques, N.T.C.; Teixeira, L.M.; Póvoa, H.C.C.; Neves, F.P.G. Prevalence of PspA families and pilus islets among Streptococcus pneumoniae colonizing children before and after universal use of pneumococcal conjugate vaccines in Brazil. Braz. J. Microbiol. 2019, 51, 419–425. [Google Scholar] [CrossRef]
- Hjálmarsdóttir, M.Á.; Pétursdóttir, B.; Erlendsdóttir, H.; Haraldsson, G.; Kristinsson, K.G. Prevalence of pilus genes in pneumococci isolated from healthy preschool children in Iceland: Association with vaccine serotypes and antibiotic resistance. J. Antimicrob. Chemother. 2015, 70, 2203–2208. [Google Scholar] [CrossRef] [Green Version]
- Kulohoma, B.W.; Gray, K.; Kamng’Ona, A.; Cornick, J.; Bentley, S.D.; Heyderman, R.S.; Everett, D.B. Piliation of Invasive Streptococcus pneumoniae Isolates in the Era before Pneumococcal Conjugate Vaccine Introduction in Malawi. Clin. Vaccine Immunol. 2013, 20, 1729–1735. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Number of Children | Number of Carriers (%, 95% CI) | OR * (95% CI)(p-Value) |
|---|---|---|---|
| Total number of children | 513 | 151 (29.4, 26–34) | |
| Gender | |||
| Males | 262 | 74 (28.2, 23–34) | 1 |
| Females | 251 | 77 (30.7, 25–37 | 1.124 (0.77–1.64) (p = 0.546) |
| Source of participants | |||
| Immunization clinic | 100 | 27 (27, 18–36) | 1 |
| Kindergarten | 413 | 124 (30, 26–34) | 1.160 (0.71–1.89) (p = 0.552) |
| Age group (months) | |||
| ≤24 | 162 | 50 (30.9, 0.24–0.38) | 1 |
| >24 | 351 | 101 (28.8, 0.24–0.34) | 0.905 (0.60–1.36) (p = 0.677) |
| 1–6 | 6 | 2 (33, 21–88) | 1 |
| 7–12 | 47 | 14 (30, 16–43) | 1.29 (0.21–7.83) (p = 0.785) |
| 13–18 | 54 | 18 (33, 20–46) | 1.09 (0.45–2.63) (p = 0.846) |
| 19–24 | 55 | 16 (29, 17–41) | 1.29 (0.56–2.97) (p = 0.556) |
| 25–30 | 98 | 22 (22, 14–31) | 1.06 (0.45–2.46) (p = 0.902) |
| 31–36 | 79 | 23 (29, 19–39) | 0.74 (0.32–1.62) (p = 0.457) |
| 37–42 | 26 | 8 (31, 12–50) | 1.06 (0.48–2.32) (p = 0.892) |
| 43–48 | 45 | 17 (38, 23–53) | 1.14 (0.41–3.22) (p = 0.801) |
| 49–54 | 53 | 17 (32, 19–45) | 1.56 (0.66–3.70) (p = 0.312) |
| 55–59 | 50 | 14 (28, 15–41) | 1.21 (0.52–2.83) (p = 0.652) |
| Total | 513 | 151 (29.4, 25–34) |
| Variable | No./Total (%) | Crude OR * (95% CI) | p-Value | Adjusted OR * (95% CI *) | p-Value |
|---|---|---|---|---|---|
| Sex | |||||
| Male | 17/74 (23) | ||||
| Female | 26/77 (33.8) | 1.709 (0.83–3.50) | 0.153 | ||
| Age group | |||||
| ≤24 months | 15/50 (30) | ||||
| ≥24 months | 28/101 (72.3) | 0.895 (0.43–1.89) | 0.849 | ||
| Vaccine type | |||||
| non-PCV13 | 16/93 (17.2) | ||||
| PCV13 | 27/58 (46.6) | 4.192 (1.99–8.84) | 0.000 | 3.945 (1.68–9.26) | 0.002 |
| Virulence genes | |||||
| pcpA negative | 6/19 (31.6) | ||||
| pcpA positive | 37/132 (28.0) | 0.844 (0.29–2.34) | 0.788 | 1.036 (0.29–3.62) | 0.956 |
| psrP negative | 26/56 (46.4) | ||||
| psrP positive | 17/95 (17.9) | 0.251 (0.12–0.53) | 0.001 | 0.244 (0.11–0.55) | 0.001 |
| PI-1 negative | 35/133 (26.3) | ||||
| PI-1 positive | 8/18 (44.4) | 2.240 (0.82–6.13) | 0.161 | 1.168 (0.33–4.12) | 0.809 |
| PI-2 negative | 37/141 (26.2) | ||||
| PI-2 positive | 6/10 (60) | 4.216 (1.13–15.78) | 0.032 | 1.690 (0.38–7.48) | 0.489 |
| Serotype | pcpA | psrP | PI-1 | PI-2 | Total Number |
|---|---|---|---|---|---|
| 23B | 21 | 8 | 0 | 1 | 22 |
| 6B | 7 | 11 | 7 | 0 | 14 |
| 23F | 11 | 5 | 0 | 0 | 13 |
| 13 | 11 | 10 | 1 | 0 | 11 |
| 19F | 9 | 6 | 5 | 9 | 11 |
| 11A | 8 | 7 | 1 | 0 | 8 |
| 15B | 8 | 6 | 0 | 0 | 8 |
| 34 | 6 | 7 | 0 | 0 | 8 |
| 15A | 5 | 6 | 0 | 0 | 6 |
| 19B | 6 | 5 | 0 | 0 | 6 |
| 3 | 6 | 2 | 0 | 0 | 6 |
| 6A | 6 | 6 | 1 | 0 | 6 |
| 21 | 4 | 5 | 0 | 0 | 5 |
| 10A | 5 | 1 | 0 | 0 | 5 |
| 14 | 4 | 2 | 0 | 0 | 4 |
| 17F | 4 | 0 | 0 | 0 | 4 |
| 9V | 3 | 1 | 3 | 0 | 3 |
| 35B | 2 | 2 | 0 | 0 | 2 |
| 7C | 0 | 2 | 0 | 0 | 2 |
| 15C | 1 | 0 | 0 | 0 | 1 |
| 16F | 1 | 1 | 0 | 0 | 1 |
| 18C | 1 | 1 | 0 | 0 | 1 |
| 20 | 1 | 1 | 0 | 0 | 1 |
| 31 | 1 | 1 | 0 | 0 | 1 |
| 38 | 1 | 1 | 0 | 0 | 1 |
| NT | 0 | 0 | 0 | 0 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mills, R.O.; Abdullah, M.R.; Akwetey, S.A.; Sappor, D.C.; Cole, I.; Baffuor-Asare, M.; Bolivar, J.A.; Gámez, G.; van der Linden, M.P.G.; Hammerschmidt, S. Post-Vaccination Streptococcus pneumoniae Carriage and Virulence Gene Distribution among Children Less Than Five Years of Age, Cape Coast, Ghana. Microorganisms 2020, 8, 1987. https://doi.org/10.3390/microorganisms8121987
Mills RO, Abdullah MR, Akwetey SA, Sappor DC, Cole I, Baffuor-Asare M, Bolivar JA, Gámez G, van der Linden MPG, Hammerschmidt S. Post-Vaccination Streptococcus pneumoniae Carriage and Virulence Gene Distribution among Children Less Than Five Years of Age, Cape Coast, Ghana. Microorganisms. 2020; 8(12):1987. https://doi.org/10.3390/microorganisms8121987
Chicago/Turabian StyleMills, Richael O., Mohammed R. Abdullah, Samuel A. Akwetey, Dorcas C. Sappor, Isaac Cole, Michael Baffuor-Asare, Johan A. Bolivar, Gustavo Gámez, Mark P. G. van der Linden, and Sven Hammerschmidt. 2020. "Post-Vaccination Streptococcus pneumoniae Carriage and Virulence Gene Distribution among Children Less Than Five Years of Age, Cape Coast, Ghana" Microorganisms 8, no. 12: 1987. https://doi.org/10.3390/microorganisms8121987