A Streptomyces sp. NEAU-HV9: Isolation, Identification, and Potential as a Biocontrol Agent against Ralstonia solanacearum of Tomato Plants

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Screening and Isolation of Actinomycetes
2.3. Screening of Antagonistic Actinobacteria Strains
2.4. Morphological and Biochemical Characteristics of NEAU-HV9
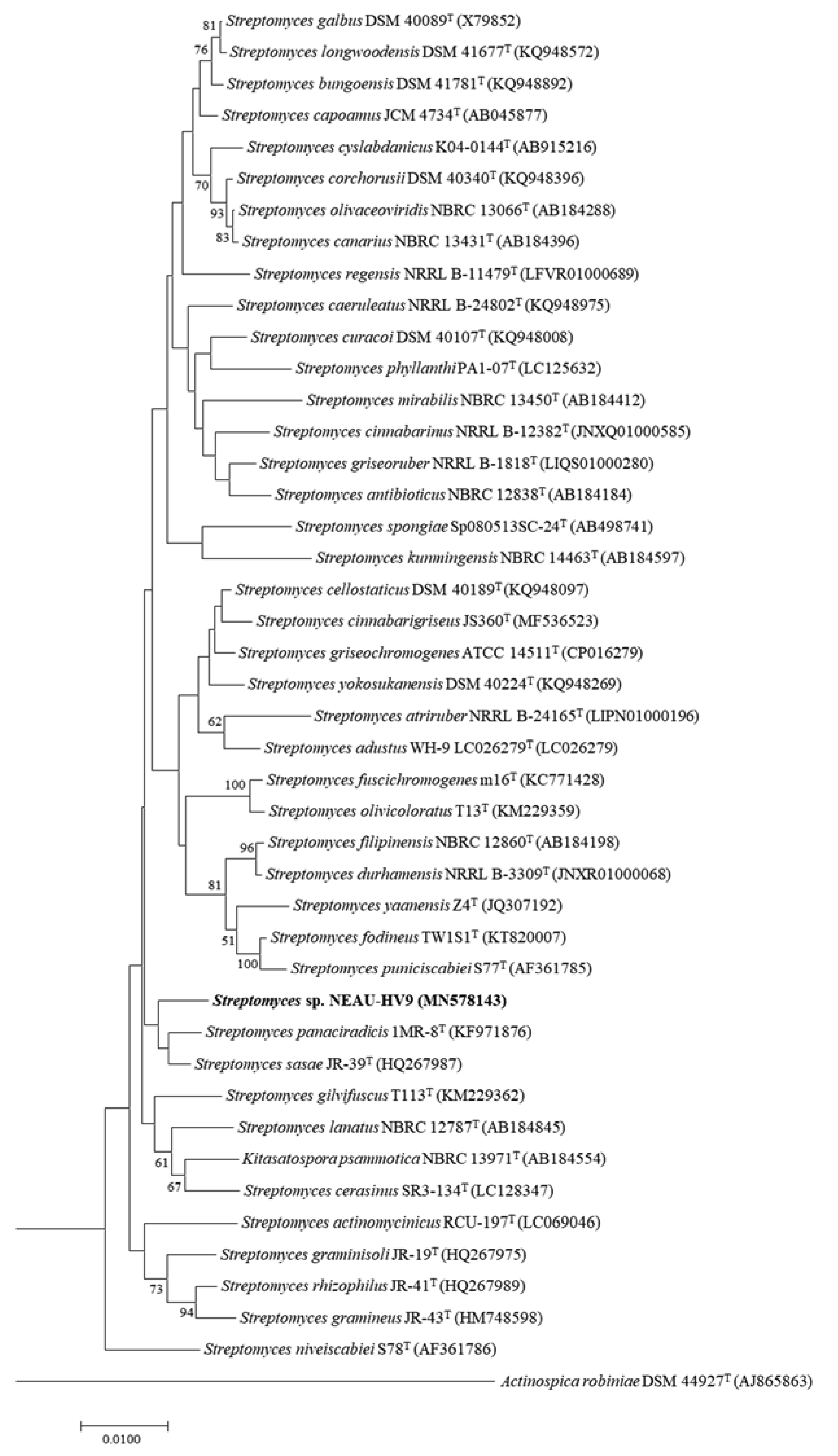
2.5. Phylogenetic Analysis of NEAU-HV9
2.6. Fermentation
2.7. Isolation and Purification of Antibacterial Compounds
2.8. Determination of Minimum Inhibitory Concentration (MIC)
2.9. Biological Assays in Tomato Seedlings
2.10. Pot Culture Experiments
3. Results
3.1. Isolation and Identification of an Antimicrobial Compound Producing Strain
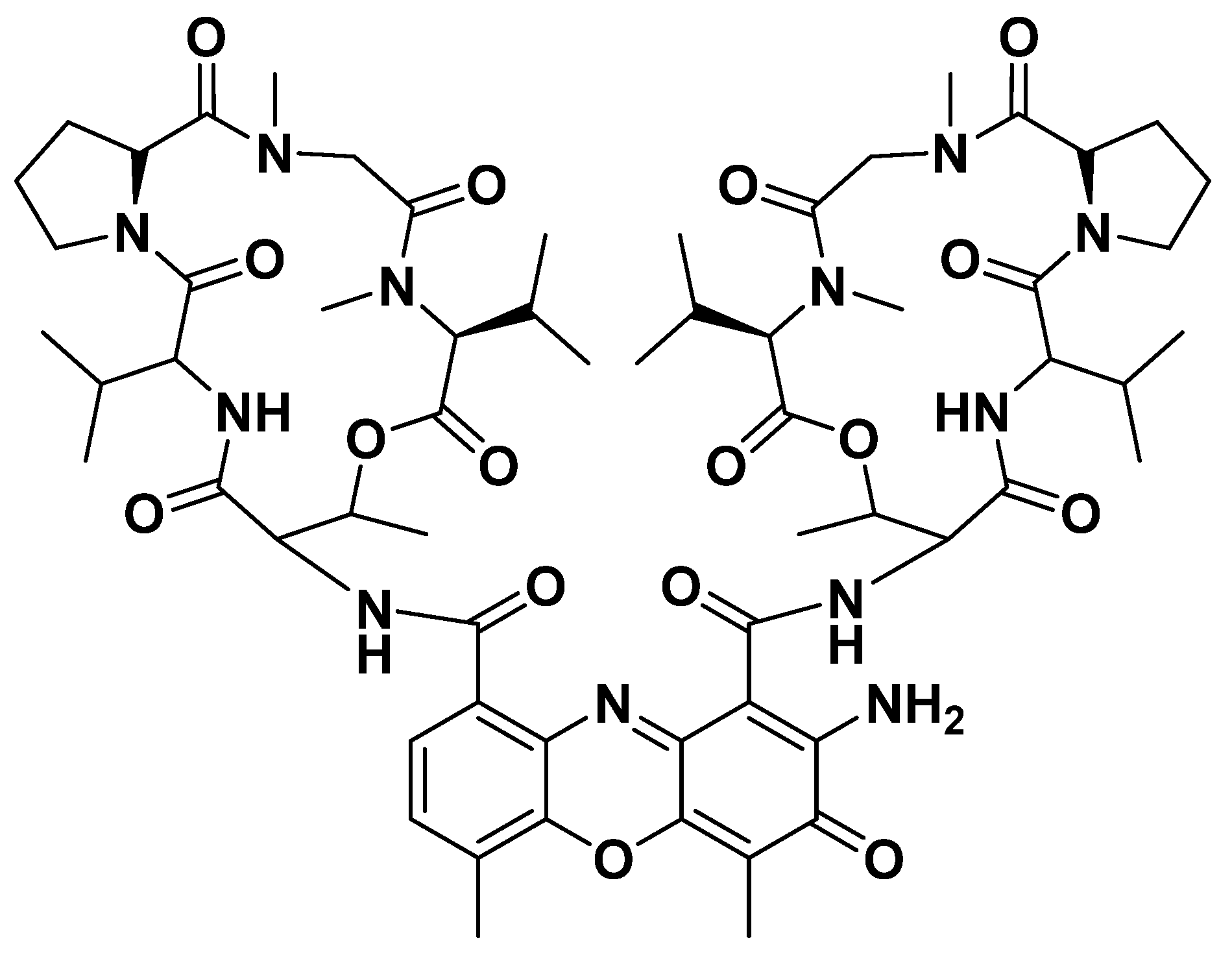
3.2. Structural Characterization of Compound
3.3. Bioactivity of Isolated Compound
3.3.1. Minimum Inhibitory Concentration (MIC)
3.3.2. Biological Assays in Tomato Seedlings
3.4. Pot Culture Experiments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lee, C.G.; Lida, T.; Uwagaki, Y.; Otani, Y.; Nakaho, K.; Ohkuma, M. Comparison of prokaryotic and eukaryotic communities in soil samples with and without tomato bacterial wilt collected from different fields. Microbes Eniviron. 2017, 32, 376–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.K.; Singh, A.K.; Kumar, A. Disease management of tomato through PGPB: Current trends and future perspective. 3Biotech 2017, 7, 255. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.G.; Wu, F.Z. Dynamics of the diversity of fungal and Fusarium communities during continuous cropping of cucumber in the green house. FEMS Microbiol. Ecol. 2012, 802, 469–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yabuuchi, E.; Kosako, Y.; Yano, I.; Hota, H.; Nishiuchi, Y. Transfer of two Burkholderia and an Alcaligenes species to Ralstonia gen. nov., Ralstonia solanacearum (Smith, 1986) comb. nov. Microbiol. Immun. 1995, 39, 897–904. [Google Scholar] [CrossRef]
- Hayward, A.C. Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum. Ann. Rev. Phytopathol. 1991, 29, 65–87. [Google Scholar] [CrossRef]
- Vu, T.T.; Kim, J.C.; Choi, Y.H.; Choi, G.J.; Jang, K.S.; Choi, T.H.; Yoon, T.M.; Lee, S.W. Effect of Gallotannins Derived from Sedum takesimense on Tomato Bacterial Wilt. Plant Dis. 2013, 97, 1593–1598. [Google Scholar] [CrossRef] [Green Version]
- Genin, S.; Boucher, C. Lessons learned from the genome analysis of Ralstonia solanacearum. Ann. Rev. Phytopathol. 2004, 42, 107–134. [Google Scholar] [CrossRef]
- Ciampi-Panno, L.; Fernandez, C.; Bustamante, P.; Andrade, N.; Ojeda, S.; Conteras, A. Biological control of bacterial wilt of potatoes caused by Pseudomonas solanacearum. Am. Potato J. 1989, 66, 315–332. [Google Scholar] [CrossRef]
- McLaughlin, R.J.; Sequeira, L.; Weingartner, D.P. Biocontrol of bacterial wilt of potato with an avirulent strain of Pseudomonas solanacearum: Interactions with root-knot nematodes. Am. Potato J. 1990, 67, 93–107. [Google Scholar] [CrossRef]
- Toyota, K.; Kimura, M. Suppresision of Ralstonia solanacearum in soil following colonization by other strains of R. solanacearum. Soil Sci. Plant Nutr. 2000, 46, 449–459. [Google Scholar]
- Viaene, T.; Langendries, S.; Beirinckx, S.; Maes, M.; Goormachtig, S. Streptomyces as a plant’s best friend? FEMS Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, M.P.; Bastos, M.S.; Xavier, V.B.; Cassel, E.; Astarita, L.V.; Santarémm, E.R. Plant growth and resistance promoted by Streptomyces spp. in tomato. Plant Physiol. Biochem. 2017, 118, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Xio, K.; Kinkel, L.L.; Samac, D.A. Biological control of Phytophthora root rots on alfalfa and soybean with Streptomyces. Biol. Control 2002, 23, 285–295. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; Sivasithamparam, K. Non-streptomycete Actinobacteria as biocontrol agents of soil-borne fungal plant pathogens and as plant growth promoters. Soil Biol. Biochem. 2006, 38, 1505–1520. [Google Scholar] [CrossRef]
- Taechowisan, T.; Chuaychot, N.; Chanaphat, S.; Wanbanjob, A.; Tantiwachwutikul, P. Antagonistic effects of Streptomyces sp. SRM1 on colletotrichum musae. Biotechnology 2009, 8, 86–92. [Google Scholar] [CrossRef]
- Bentley, S.D.; Chater, K.F.; Cerden, A.M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3. Nature 2002, 417, 141–147. [Google Scholar] [CrossRef]
- Brockmann, H. Die actinomycine. Angew. Chem. 1960, 87, 1767–1947. [Google Scholar]
- Farber, S.; D’Angio, G.; Evans, A.; Mitus, A. Clinical studies of actinomycin D with special reference to Wilms’ tumor in children. J. Urol. 2002, 168, 2560–2562. [Google Scholar] [CrossRef]
- Womer, R.B. Soft tissue sarcomas. Eur. J. Cancer 1997, 33, 2230–2234. [Google Scholar] [CrossRef]
- Monteiro, F.; Genin, S.; van, D.I.; Valls, M. A luminescent reporter evidences active expression of Ralstonia solanacearum type III secretion system genes throughout plant infection. Microbiology 2012, 158, 2107–2116. [Google Scholar] [CrossRef] [Green Version]
- Guidot, A.; Jiang, W.; Ferdy, J.B.; Thébaud, C.; Barberis, P.; Gouzy, J.; Genin, S. Multihost experimental evolution of the pathogen Ralstonia solanacearum unveils genes involved in adaptation to plants. Mol. Biol. Evol. 2014, 11, 2913–2928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Phukan, T.; Sharma, P.L.; Kabyashree, K.; Barman, A.; Kumar, R.; Sonti, R.V.; Genin, S.; Ray, S.K. An Innovative Root Inoculation Method to Study Ralstonia solanacearum Pathogenicity in Tomato Seedlings. Phytopathology 2018, 108, 436–442. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R. Studying Virulence Functions of Ralstonia solanacearum, the Causal Agent of Bacterial Wilt in Plants. Ph.D. Thesis, Tezpur University, Tezpur, India, 2014. [Google Scholar]
- van Elsas, J.D.; Kastelein, P.; de Vries, P.M.; van Overbeek, L.S. Effects of ecological factors on the survival and physiology of Ralstonia solanacearum bv 2 in irrigation water. Can. J. Microbiol. 2001, 47, 842–854. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, M.; Nonomura, H. Humic acid-vitamin agar, a new medium for selective isolation of soil actinomycetes. J. Ferment. Technol. 1987, 65, 501–509. [Google Scholar] [CrossRef]
- Shirling, E.B.; Gottlieb, D. Methods for characterization of Streptomyces species. Int. J. Syst. Bacteriol. 1966, 16, 313–340. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.C.; Brouchkov, A.; Griva, G.; Schinner, F.; Margesin, R. Isolation and characterization of bacteria from ancient siberian permafrost sediment. Biology 2013, 2, 85–106. [Google Scholar] [CrossRef]
- Fu, Y.S.; Yan, R.; Liu, D.; Zhao, J.W.; Song, J.; Wang, X.J.; Cui, L.; Zhang, J.; Xiang, W.S. Characterization of Sinomonas gamaensis sp. nov., a Novel Soil Bacterium with Antifungal Activity against Exserohilum turcicum. Microorganisms 2019, 7, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Wu, Y.; Wang, M. Identification and antifungal activity of an actinomycete strain against Alternaria spp. Span. J. Agric. Res. 2014, 12, 1158–1165. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.Y.; Zhao, Y.; Song, W.; Duan, L.P.; Jiang, S.W.; Wang, X.J.; Zhao, J.W.; Xiang, W.S. Streptomyces inhibens sp. nov., a novel actinomycete isolated from rhizosphere soil of wheat (Triticum aestivum L.). Int. J. Syst. Evol. Microbiol. 2019, 69, 688–695. [Google Scholar] [CrossRef]
- Waksman, S.A. The Actinomycetes. A Summary of Current Knowledge; Ronald: New York, NY, USA, 1967. [Google Scholar]
- Jones, K.L. Fresh isolates of actinomycetes in which the presence of sporogenous aerial mycelia is a fluctuating characteristic. J. Bacteriol. 1949, 57, 141–145. [Google Scholar] [CrossRef] [Green Version]
- Waksman, S.A. Classification, identification and descriptions of genera and species. In The Actinomycetes; Williams and Wilkins: Baltimore, MD, USA, 1961; Volume 2. [Google Scholar]
- Kelly, K.L. Inter-Society Color Council-National Bureau of Standards Color-Name Charts Illustrated with Centroid Colors; Government Printing Office: Washington, DC, USA, 1964.
- Zhao, J.W.; Han, L.Y.; Yu, M.Y.; Cao, P.; Li, D.M.; Guo, X.W.; Liu, Y.Q.; Wang, X.J.; Xiang, W.S. Characterization of Streptomyces sporangiiformans sp. nov., a Novel Soil Actinomycete with Antibacterial Activity against Ralstonia solanacearum. Microorganisms. 2019, 7, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, R.E.; Barnett, D.A.; Handerhan, J.E.; Pang, C. Nocardia coeliaca, Nocardia autotrophica, and the nocardin strain. Int. J. Syst. Bacteriol. 1974, 24, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Yokota, A.; Tamura, T.; Hasegawa, T.; Huang, L.H. Catenuloplanes japonicas gen. nov., sp. nov., nom. rev., a new genus of the order Actinomycetales. Int. J. Syst. Bacteriol. 1993, 43, 805–812. [Google Scholar] [CrossRef] [Green Version]
- Woese, C.R.; Gutell, R.; Gupta, R.; Noller, H.F. Detailed analysis of the higher-order structure of 16S-like ribosomal ribonucleic acids. Microbiol. Rev. 1983, 47, 621–669. [Google Scholar] [CrossRef] [PubMed]
- Springer, N.; Ludwig, W.; Amann, R.; Schmidt, H.J.; Görtz, H.D.; Schleifer, K.H. Occurrence of fragmented 16S rRNA in an obligate bacterial endosymbiont of Paramecium caudatum. Proc. Natl. Acad. Sci. USA 1993, 90, 9892–9895. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA and whole genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Rathod, B.B.; Korasapati, R.; Sripadi, P.; Reddy, S.P. Novel actinomycin group compound from newly isolated Streptomyces sp. RAB12: Isolation, characterization, and evaluation of antimicrobial potential. Appl. Microbiol. Biotechnol. 2018, 102, 1241–1250. [Google Scholar] [CrossRef]
- Guo, J.H.; Qi, H.Y.; Guo, Y.H.; Ge, H.; Gong, L.Y.; Zhang, L.X.; Sun, P.H. Biocontrol of tomato wilt by growth-promoting rhizobacteria. Biol. Control 2004, 29, 66–72. [Google Scholar] [CrossRef]
- Roy, N.; Choi, K.; Khan, R.; Lee, S.W. Culturing Simpler and Bacterial Wilt Suppressive Microbial Communities from Tomato Rhizosphere. Plant Pathol. J. 2019, 35, 362–371. [Google Scholar] [PubMed]
- Roberts, D.P.; Denny, T.P.; Schell, M.A. Cloning of the egl gene of Pseudomonas solanacearum and analysis of its role in phytopathogenicity. J. Bacteriol. 1988, 170, 1445–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukuda, M.; Kitahara, K.; Miyazaki, K. Comparative RNA function analysis reveals high functional similarity between distantly related bacterial 16 S rRNAs. Sci. Rep. 2017, 7, 9993. [Google Scholar] [CrossRef] [Green Version]
- Woese, C.R. Bacterial evolution. Microbiol. Rev. 1987, 51, 221–2711. [Google Scholar] [CrossRef]
- Kulkarni, M.; Gorthi, S.; Banerjee, G.; Chattopadhyay, P. Production, characterization and optimization of actinomycin D from Streptomyces hydrogenans IB310, a(n antagonistic bacterium against phytopathogens. Biocatal. Agric. Biotechnol. 2017, 10, 69–74. [Google Scholar] [CrossRef]
- Zhang, L.L.; Wan, C.X.; Luo, X.X.; Lv, L.L.; Wang, X.P.; Xia, Z.F. Manufacture of Actinomycin D with Streptomyces mutabilis for Controlling Plant Pathogenic Bacteria, Cow Mastitis, and Female Colpitis. China Patent CN104450580A, 25 March 2015. [Google Scholar]
- Chen, C.; Song, F.; Wang, Q.; Abdel-Mageed, W.M.; Guo, H.; Fu, C.; Hou, W.; Dai, H.; Liu, X.; Yang, N.; et al. A marine-derived Streptomyces sp. MS449 produces high yield of actinomycin X2 and actinomycin D with potent anti-tuberculosis activity. Appl. Microbiol. Biotechnol. 2012, 95, 919–927. [Google Scholar] [CrossRef]
- Chen, Y.F.; Zhou, D.B.; Qi, D.F.; Gao, Z.F.; Xie, J.H.; Luo, Y.P. Growth promotion and disease suppression ability of a streptomyces sp. CB-75 from banana rhizosphere soil. Front. Microbiol. 2018, 8, 2704. [Google Scholar] [CrossRef]
- Crawford, D.L.; Lynch, J.M.; Whipps, J.M.; Ousley, M.A. Isolation and characterization of actionomycete antagonists of a fungal root pathogen. Appl. Environ. Microb. 1993, 59, 3889–3905. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Qiu, Z.; You, J.; Tan, H.; Zhou, S. Isolation and characterization of endophytic Streptomyces strains from surface-sterilized tomato (Lycopersicon esculentum) roots. Lett. Appl. Microbiol. 2004, 39, 425–430. [Google Scholar]
- Gopalakrishnan, S.; Pande, S.; Sharma, M.; Humayun, P.; Kiran, B.K.; Sandeep, D.; Vidya, M.S.; Deepthi, K.; Rupela, O. Evaluation of actinomycete isolates obtained from herbal vermicompost for biological control of Fusarium wilt of chickpea. Crop Protect. 2011, 30, 1070–1078. [Google Scholar] [CrossRef] [Green Version]
- Bull, C.T. Relationship between root colonization and suppression of Gaeumannomyces graminis var tritici by Pseudomonas fluorescens strain 2–79. Phytopathology 1991, 81, 954–959. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Leeman, M.; van Oorschot, M.M.P.; van der Sluis, I.; Schippers, B.; Bakker, P.A.H.M. Dose-response relationships in biological-control of Fusarium-wilt of radish by Pseudomonas spp. Phytopathology 1995, 85, 1075–1081. [Google Scholar] [CrossRef]
- Wang, J.J.; Zhao, Y.; Ruan, Y.Z. Effects of bio-organic fertilizers produced by four Bacillus amyloliquefaciens strains on banana Fusarium wilt Disease. Compost Sci. Util. 2015, 23, 185–198. [Google Scholar] [CrossRef]
- Hong, K.; Gao, A.H.; Xie, Q.Y.; Gao, H.; Zhuang, L.; Lin, H.P.; Yu, H.P.; Li, J.; Yao, X.S.; Goodfellow, M.; et al. Actino-mycetes for marine drug discovery isolated from man-grove soils and plants in China. Mar. Drugs 2009, 7, 24–44. [Google Scholar] [CrossRef]
- Sasaki, T.; Igarashi, Y.; Ogawa, M.; Furumai, T. Identification of 6-prenylindole as an antifungal metabolite of Streptomyces sp. TP-A0595 and synthesis and bioactivity of 6-substituted indoles. J. Antibiot. 2002, 55, 1009–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, X.H.; Naing, K.W.; Lee, Y.S.; Kim, Y.H.; Moon, J.H.; Kim, K.Y. Antagonism of antifungal metabolites from Streptomyces griseus H7602 against Phytophthora capsici. J. Basic Microbiol. 2015, 55, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Kakinuma, K.; Hanson, C.A.; Rinehart, K.L., Jr. Spectinabilin, a new nitro-containing metabolite isolated from Streptomyces spectabilis. Tetrahedron 1976, 32, 217–222. [Google Scholar] [CrossRef]
- Ivanova, V. New macrolactone of the desertomycin family from Streptomyces spectabilis. Prep. Biochem. Biotechnol. 1997, 27, 19–38. [Google Scholar] [CrossRef]
- Kim, K.R.; Kim, T.J.; Suh, J.W. The gene cluster for spectinomycin biosynthesis and the aminoglycoside-resistance function of spcm in Streptomyces spectabilis. Curr. Microbiol. 2008, 57, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Selvakumar, J.; Chandrasekaran, S.; Vaithilingam, M. Bio prospecting of marine-derived Streptomyces spectabilis VITJS10 and exploring its cytotoxicity against human liver cancer celllines. Pharmacogn. Mag. 2015, 11, 469–473. [Google Scholar]
- Waksman, S.A.; Woodruff, H.B. Bacteriostatic and bactericidal substances produced by a soil actinomyces. Proc. Soc. Exp. Biol. Med. 1940, 45, 609–614. [Google Scholar] [CrossRef]
- Vu, T.T.; Kim, H.; Tran, V.K.; Vu, H.D.; Hoang, T.X.; Han, J.W.; Choi, Y.H.; Jang, K.S.; Choi, G.J.; Kim, J.J. Antibacterial activity of tannins isolated from Sapium baccatum extract and use for control of tomato bacterial wilt. PLoS ONE 2017, 12, e0181499. [Google Scholar] [CrossRef]
- Zheng, S.J.; Zhu, R.; Tang, B.; Chen, L.Z.; Bai, H.J.; Zhang, J.W. Synthesis and biological evaluations of a series of calycanthaceous analogues as antifungal agents. Nat. Prod. Res. 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Methanol Extract of Cell Pellet | Supernatant | |
|---|---|---|
| Inhibitory zone diameters (mm) | 30.5 | 32.8 |
| Pathogen | MIC (mg/L) |
|---|---|
| R. solanacearum | 0.6 ± 0.2 |
| Treatment | Wilt Incidence (%) | Control Efficacy (%) |
|---|---|---|
| NEAU-HV9 (107 cfu mL−1) | 100 ± 0 a | 0 ± 0 a |
| NEAU-HV9 (108 cfu mL−1) | 93 ± 11.6 b | 6.6 ± 11.5 b |
| NEAU-HV9 (109 cfu mL−1) | 0 ± 0 c | 100 ± 0 c |
| Actinomycin D (1 × MIC) | 0 ± 0 c | 100 ± 0 c |
| Actinomycin D (2 × MIC) | 0 ± 0 c | 100 ± 0 c |
| Control | 100 ± 0 a | … |
| Treatment | Wilt Incidence (%) | Control Efficacy (%) |
|---|---|---|
| NEAU-HV9 | 13.3 ± 5.8 b | 82 ± 6 a |
| Actinomycin D | 0 ± 0 b | 100 ± 0 a |
| Control | 73.9 ± 6.6 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ling, L.; Han, X.; Li, X.; Zhang, X.; Wang, H.; Zhang, L.; Cao, P.; Wu, Y.; Wang, X.; Zhao, J.; et al. A Streptomyces sp. NEAU-HV9: Isolation, Identification, and Potential as a Biocontrol Agent against Ralstonia solanacearum of Tomato Plants. Microorganisms 2020, 8, 351. https://doi.org/10.3390/microorganisms8030351
Ling L, Han X, Li X, Zhang X, Wang H, Zhang L, Cao P, Wu Y, Wang X, Zhao J, et al. A Streptomyces sp. NEAU-HV9: Isolation, Identification, and Potential as a Biocontrol Agent against Ralstonia solanacearum of Tomato Plants. Microorganisms. 2020; 8(3):351. https://doi.org/10.3390/microorganisms8030351
Chicago/Turabian StyleLing, Ling, Xiaoyang Han, Xiao Li, Xue Zhang, Han Wang, Lida Zhang, Peng Cao, Yutong Wu, Xiangjing Wang, Junwei Zhao, and et al. 2020. "A Streptomyces sp. NEAU-HV9: Isolation, Identification, and Potential as a Biocontrol Agent against Ralstonia solanacearum of Tomato Plants" Microorganisms 8, no. 3: 351. https://doi.org/10.3390/microorganisms8030351