Phenotypic and Genotypic Traits of Vancomycin-Resistant Enterococci from Healthy Food-Producing Animals

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Bacterial Isolation
2.2. Multiplex PCR Detection of vanA, vanB, vanC-1, and vanC-2/3 Genes
2.3. Antimicrobial Susceptibility Testing
2.4. Conjugation Experiments
2.5. Whole Genome Sequencing
2.6. Accession Numbers
3. Results
3.1. Prevalence of VRE among Healthy Food-Producing Animals
3.2. Antimicrobial Susceptibility Testing and Detection of Resistance Genes
3.3. Mating Experiments
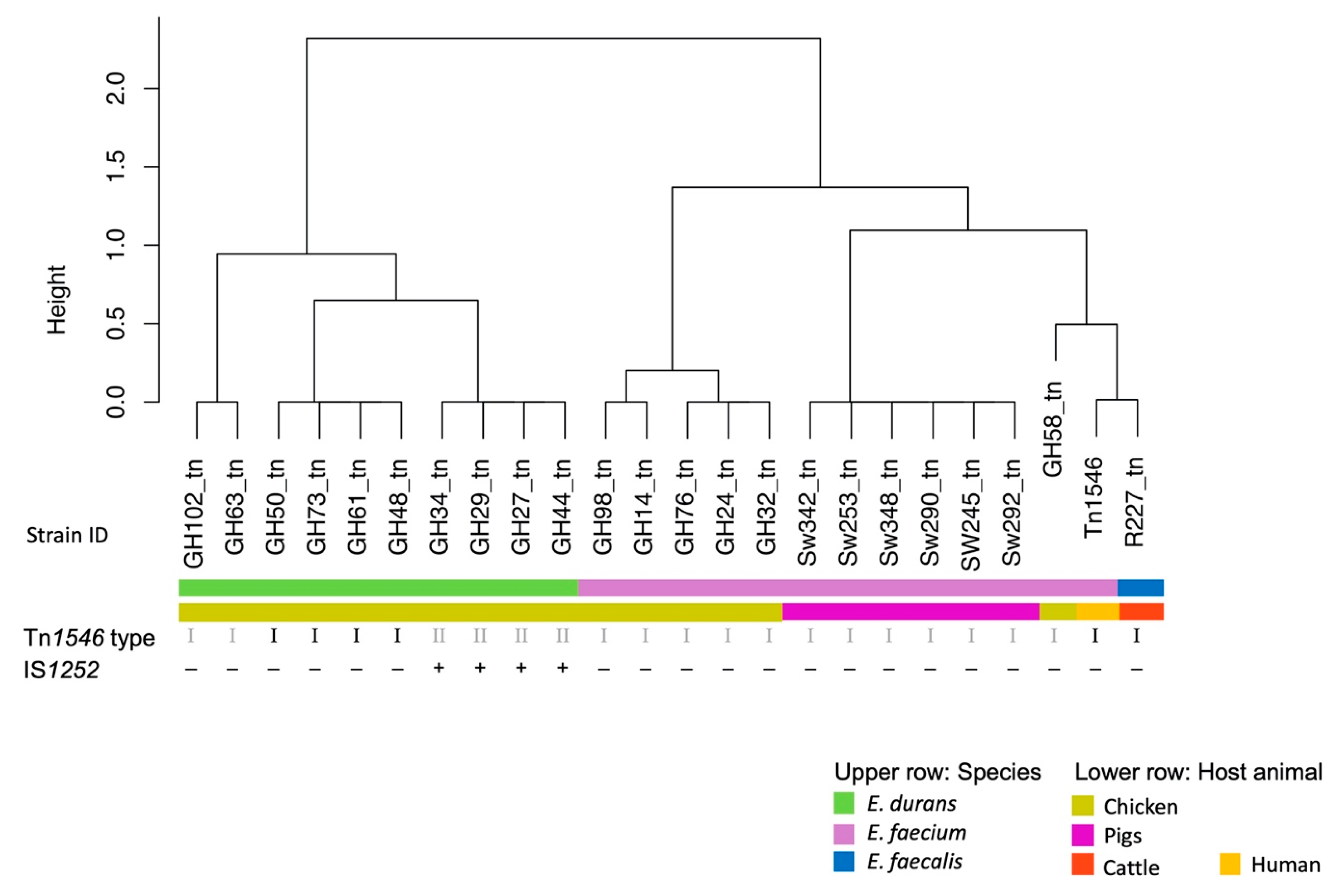
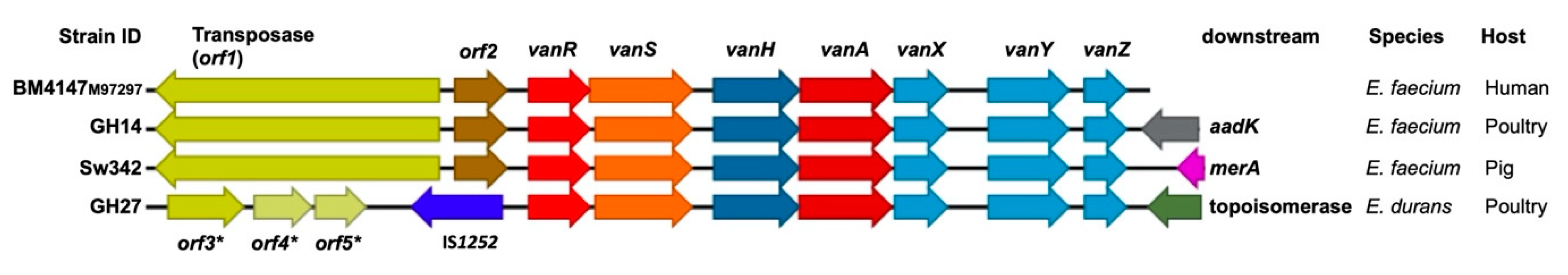
3.4. Characterisation of the Tn1546 Structures
3.5. Multilocus Sequence Typing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Puchter, L.; Chaberny, I.F.; Schwab, F.; Vonberg, R.P.; Bange, F.C.; Ebadi, E. Economic burden of nosocomial infections caused by vancomycin-resistant enterococci. Antimicrob. Resist. Infect. Control 2018, 7, 1. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- ECDC. Surveillance of antimicrobial resistance in Europe 2018; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2019; pp. 57–60. ISBN 978-92-9498-387-9.
- Wassilew, N.; Seth-Smith, H.M.; Rolli, E.; Fietze, Y.; Casanova, C.; Führer, U.; Egli, A.; Marschall, J.; Buetti, N. Outbreak of vancomycin-resistant Enterococcus faecium clone ST796, Switzerland, December 2017 to April 2018. Eurosurveillance 2018, 23, 1800351. [Google Scholar] [CrossRef] [PubMed]
- Howden, B.P.; Holt, K.E.; Lam, M.M.C.; Seemann, T.; Ballard, S.; Coombs, G.W.; Tong, S.Y.C.; Grayson, M.L.; Johnson, P.D.R.; Stinear, T.P. Genomic insights to control the emergence of vancomycin-resistant enterococci. MBio 2013, 4, e00412-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinholt, M.; Gumpert, H.; Bayliss, S.; Nielsen, J.B.; Vorobieva, V.; Pedersen, M.; Feil, E.; Worning, P.; Westh, H. Genomic analysis of 495 vancomycin-resistant Enterococcus faecium reveals broad dissemination of a vanA plasmid in more than 19 clones from Copenhagen, Denmark. J. Antimicrob. Chemother. 2016, 72, 40–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raven, K.E.; Gouliouris, T.; Brodrick, H.; Coll, F.; Brown, N.M.; Reynolds, R.; Reuter, S.; Török, M.E.; Parkhill, J.; Peacock, S.J. Complex routes of nosocomial vancomycin-resistant Enterococcus faecium transmission revealed by genome sequencing. Clin. Infect. Dis. 2017, 64, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.O.; Baptiste, K.E. Vancomycin-resistant enterococci: A review of antimicrobial resistance mechanisms and perspectives of human and animal health. Microb. Drug Resist. 2018, 24, 590–606. [Google Scholar] [CrossRef] [Green Version]
- Courvalin, P. Vancomycin resistance in gram-positive cocci. Clin. Infect. Dis. 2006, 42, S25–S34. [Google Scholar] [CrossRef]
- Sletvold, H.; Johnsen, P.J.; Wikmark, O.-G.; Simonsen, G.S.; Sundsfjord, A.; Nielsen, K.M. Tn 1546 is part of a larger plasmid-encoded genetic unit horizontally disseminated among clonal Enterococcus faecium lineages. J. Antimicrob. Chemother. 2010, 65, 1894–1906. [Google Scholar] [CrossRef] [Green Version]
- Acar, J.; Casewell, M.; Freeman, J.; Friis, C.; Goossens, H. Avoparcin and virginiamycin as animal growth promoters: A plea for science in decision-making. Clin. Microbiol. Infect. 2000, 6, 477–482. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.S.; Seol, S.Y.; Cho, D.T. Diversity of Tn1546-like elements in vancomycin-resistant enterococci isolated from humans and poultry in Korea. J. Clin. Microbiol. 2003, 41, 2641–2643. [Google Scholar] [CrossRef] [Green Version]
- Willems, R.J.L.; Top, J.; van den Braak, N.; van Belkum, A.; Mevius, D.J.; Hendriks, G.; van Santen-Verheuvel, M.; van Embden, J.D.A. Molecular diversity and evolutionary relationships of Tn1546-like elements in enterococci from humans and animals. Antimicrob. Agents Chemother. 1999, 43, 483–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wardal, E.; Kuch, A.; Gawryszewska, I.; Żabicka, D.; Hryniewicz, W.; Sadowy, E. Diversity of plasmids and Tn1546-type transposons among vanA Enterococcus faecium in Poland. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 313–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López, M.; Sáenz, Y.; Alvarez-Martínez, M.J.; Marco, F.; Robredo, B.; Rojo-Bezares, B.; Ruiz-Larrea, F.; Zarazaga, M.; Torres, C. Tn1546 structures and multilocus sequence typing of vanA-containing enterococci of animal, human and food origin. J. Antimicrob. Chemother. 2010, 65, 1570–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayjanov, J.R.; Baan, J.; Rogers, M.R.C.; Troelstra, A.; Willems, R.J.L.; van Schaik, W. Enterococcus faecium genome dynamics during long-term asymptomatic patient gut colonization. Microb. Genom. 2019, 5, e000277. [Google Scholar] [CrossRef]
- Willems, R.J.L.; Top, J.; Marga van Santen, D.; Coque, T.M.; Baquero, F.; Grundmann, H.; Bonten, M.J.M. Global spread of vancomycin-resistant Enterococcus faecium from distinct nosocomial genetic complex. Emerg. Infect. Dis. 2005, 11, 821. [Google Scholar] [CrossRef]
- Lebreton, F.; van Schaik, W.; McGuire, A.M.; Godfrey, P.; Griggs, A.; Mazumdar, V.; Corander, J.; Cheng, L.; Saif, S.; Young, S. Emergence of epidemic multidrug-resistant Enterococcus faecium from animal and commensal strains. MBio 2013, 4, e00534-13. [Google Scholar] [CrossRef] [Green Version]
- Hammerum, A.M.; Baig, S.; Kamel, Y.; Roer, L.; Pinholt, M.; Gumpert, H.; Holzknecht, B.; Røder, B.; Justesen, U.S.; Samulioniené, J.; et al. Emergence of vanA Enterococcus faecium in Denmark, 2005–2015. J. Antimicrob. Chemother. 2017, 72, 2184–2190. [Google Scholar] [CrossRef]
- Mahony, A.A.; Buultjens, A.H.; Ballard, S.A.; Grabsch, E.A.; Xie, S.; Seemann, T.; Stuart, R.L.; Kotsanas, D.; Cheng, A.; Heffernan, H.; et al. Vancomycin-resistant Enterococcus faecium sequence type 796—Rapid international dissemination of a new epidemic clone. Antimicrob. Resist. Infect. Control 2018, 7, 44. [Google Scholar] [CrossRef]
- Buultjens, A.H.; Lam, M.M.; Ballard, S.; Monk, I.R.; Mahony, A.A.; Grabsch, E.A.; Grayson, M.L.; Pang, S.; Coombs, G.W.; Robinson, J.O.; et al. Evolutionary origins of the emergent ST796 clone of vancomycin resistant Enterococcus faecium. Peer J. 2017, 5, e2916. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, O. Vancomycin resistant enterococci in farm animals—Occurrence and importance. Infect. Ecol. Epidemiol. 2012, 2, 16959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual (3-Volume Set), 4th ed.; Cold Spring Harbor Laboratory Press: Cold Spring Habor, NY, USA, 2001; pp. 11–28. [Google Scholar]
- Dutka-Malen, S.; Evers, S.; Courvalin, P. Detection of glycopeptide resistance genotypes and identification to the species level of clinically relevant enterococci by PCR. J. Clin. Microbiol. 1995, 33, 1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wambui, J.; Tasara, T.; Njage, P.M.K.; Stephan, R. Species distribution and antimicrobial profiles of Enterococcus spp. isolates from Kenyan small and medium enterprise slaughterhouses. J. Food Prot. 2018, 81, 1445–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; CLSI supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018; Volume 38, pp. 1–198. ISBN 1-56238-839-8. [Google Scholar]
- Jacob, A.E.; Hobbs, S.J. Conjugal transfer of plasmid-borne multiple antibiotic resistance in Streptococcus faecalis var. zymogenes. J. Bacteriol. 1974, 117, 360–372. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Shovill. Available online: https://githubcom/tseemann/shovill (accessed on 15 January 2019).
- Stevens, M.J.A.; Cernela, N.; Corti, S.; Stephan, R. Draft genome sequence of Streptococcus parasuis 4253, the first available for the species. Microbiol. Res. Announc. 2019, 8, 18. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.V.; Cosentino, S.; Lukjancenko, O.; Saputra, D.; Rasmussen, S.; Hasman, H.; Sicheritz-Pontén, T.; Aarestrup, F.M.; Ussery, D.W.; Lund, O. Benchmarking of methods for genomic taxonomy. J. Clin. Microbiol. 2014, 52, 1529–1539. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2016, gkw1004. [Google Scholar] [CrossRef]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; De Pascale, G.; Ejim, L. The comprehensive antibiotic resistance database. Antimicrob. Agents Chemother. 2013, 57, 3348–3357. [Google Scholar] [CrossRef] [Green Version]
- Arthur, M.; Depardieu, F.; Molinas, C.; Reynolds, P.; Courvalin, P. The vanZ gene of Tn1546 from Enterococcus faecium BM4147 confers resistance to teicoplanin. Gene 1995, 154, 87–92. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. PNAS 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, L.; Glover, R.H.; Humphris, S.; Elphinstone, J.G.; Toth, I.K. Genomics and taxonomy in diagnostics for food security: Soft-rotting enterobacterial plant pathogens. Anal. Methods 2016, 8, 12–24. [Google Scholar] [CrossRef]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K. Cluster: Cluster Analysis Basics and Extensions, R package version 2.1.0. 2019.
- Arnold, S.; Gassner, B.; Giger, T.; Zwahlen, R. Banning antimicrobial growth promoters in feedstuffs does not result in increased therapeutic use of antibiotics in medicated feed in pig farming. Pharmacoepidemiol. Drug Saf. 2004, 13, 323–331. [Google Scholar] [CrossRef] [PubMed]
- FOPH. Swiss Antibiotic Resistance report 2018. Usage of Antibiotics and Occurrence of Antibiotic Resistance in Bacteria from Humans and Animals in Switzerland. November 2018; Federal Office of Public Health and Federal Food Safety and Veterinary Office: Berne, Switzerland, 2018; pp. 98–102, Publication number:2018-OEG-87. [Google Scholar]
- FOPH. Swiss Antibiotic Resistance Report 2016. Usage of Antibiotics and Occurrence of Antibiotic Resistance in Bacteria from Humans and Animals in Switzerland. November 2016; Federal Office of Public Health and Federal Food Safety and Veterinary Office: Berne, Switzerland, 2016; pp. 80–87, Publication number:2016-OEG-30. [Google Scholar]
- Aarestrup, F.M. Characterization of glycopeptide-resistant Enterococcus faecium (GRE) from broilers and pigs in Denmark: Genetic evidence that persistence of GRE in pig herds is associated with coselection by resistance to macrolides. J. Clin. Microbiol. 2000, 38, 2774–2777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattoir, V.; Leclercq, R. Twenty-five years of shared life with vancomycin-resistant enterococci: Is it time to divorce? J. Antimicrob. Chemother. 2013, 68, 731–742. [Google Scholar] [CrossRef]
- Hammerum, A.M. Enterococci of animal origin and their significance for public health. Clin. Microbiol. Infect. 2012, 18, 619–625. [Google Scholar] [CrossRef]
- Silva, V.; Igrejas, G.; Carvalho, I.; Peixoto, F.; Cardoso, L.; Pereira, J.E.; del Campo, R.; Poeta, P. Genetic characterization of vanA Enterococcus faecium isolates from wild red-legged partridges in Portugal. Microb. Drug Resist. 2018, 24, 89–94. [Google Scholar] [CrossRef]
- Lozano, C.; Gonzalez-Barrio, D.; Camacho, M.C.; Lima-Barbero, J.F.; de la Puente, J.; Höfle, U.; Torres, C. Characterization of faecal vancomycin-resistant enterococci with acquired and intrinsic resistance mechanisms in wild animals, Spain. Microb. Ecol. 2016, 72, 813–820. [Google Scholar] [CrossRef]
- Yazdankhah, S.; Rudi, K.; Bernhoft, A. Zinc and copper in animal feed—Development of resistance and co-resistance to antimicrobial agents in bacteria of animal origin. Microb. Ecol. Health Dis. 2014, 25, 25862. [Google Scholar] [CrossRef] [Green Version]
- Wright, G.D.; Ladak, P. Overexpression and characterization of the chromosomal aminoglycoside 6′-N-acetyltransferase from Enterococcus faecium. Antimicrob. Agents Chemother. 1997, 41, 956–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Campo, R.; Galán, J.C.; Tenorio, C.; Ruiz-Garbajosa, P.; Zarazaga, M.; Torres, C.; Baquero, F. New aac (6′)-I genes in Enterococcus hirae and Enterococcus durans: Effect on β-lactam/aminoglycoside synergy. J. Antimicrob. Chemother. 2005, 55, 1053–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isnard, C.; Malbruny, B.; Leclercq, R.; Cattoir, V. Genetic basis for in vitro and in vivo resistance to lincosamides, streptogramins A, and pleuromutilins (LSAP phenotype) in Enterococcus faecium. Antimicrob. Agents Chemother. 2013, 57, 4463–4469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bérenger, R.; Bourdon, N.; Auzou, M.; Leclercq, R.; Cattoir, V. In vitro activity of new antimicrobial agents against glycopeptide-resistant Enterococcus faecium clinical isolates from France between 2006 and 2008. Med. Mal. Infect. 2011, 41, 405–409. [Google Scholar] [CrossRef]
- Bourgeois-Nicolaos, N.; Moubareck, C.; Mangeney, N.; Butel, M.; Doucet-Populaire, F. Comparative study of vanA gene transfer from Enterococcus faecium to Enterococcus faecalis and to Enterococcus faecium in the intestine of mice. FEMS Microbiol. Lett. 2006, 254, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Novais, C.; Freitas, A.R.; Sousa, J.C.; Baquero, F.; Coque, T.M.; Peixe, L.V. Diversity of Tn1546 and its role in the dissemination of vancomycin-resistant enterococci in Portugal. Antimicrob. Agents Chemother. 2008, 52, 1001–1008. [Google Scholar] [CrossRef] [Green Version]
- Freitas, A.R.; Coque, T.M.; Novais, C.; Hammerum, A.M.; Lester, C.H.; Zervos, M.J.; Donabedian, S.; Jensen, L.B.; Francia, M.V.; Baquero, F. Human and swine hosts share vancomycin-resistant Enterococcus faecium CC17 and CC5 and Enterococcus faecalis CC2 clonal clusters harboring Tn1546 on indistinguishable plasmids. J. Clin. Microbiol. 2011, 49, 925–931. [Google Scholar] [CrossRef] [Green Version]
- Torres, C.; Alonso, C.A.; Ruiz-Ripa, L.; León-Sampedro, R.; Del Campo, R.; Coque, T.M. Antimicrobial Resistance in Enterococcus spp. of animal origin. Microbiol. Spectr. 2018, 6, 4. [Google Scholar] [CrossRef]
- Guardabassi, L. Dalsgaard, A. Occurrence, structure, and mobility of Tn1546-like elements in environmental isolates of vancomycin-resistant enterococci. Appl. Environ. Microbiol. 2004, 70, 984–990. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.O.; Yoo, J.I.; Kim, H.K.; Kim, H.S.; Yoo, J.S.; Lee, Y.S.; Jung, Y.H. Diversity of Tn1546 in vanA-positive Enterococcus faecium clinical isolates with VanA, VanB, and VanD phenotypes and susceptibility to vancomycin. J. Appl. Microbiol. 2013, 115, 969–976. [Google Scholar]
- Gouliouris, T.; Raven, K.E.; Ludden, C.; Blane, B.; Corander, J.; Horner, C.S.; Hernandez-Garcia, J.; Wood, P.; Hadjirin, N.F.; Radakovic, M. Genomic surveillance of Enterococcus faecium reveals limited sharing of strains and resistance genes between livestock and humans in the United Kingdom. MBio 2018, 9, e01780-18. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, O.; Greko, C.; Top, J.; Franklin, A.; Bengtsson, B. Spread without known selective pressure of a vancomycin-resistant clone of Enterococcus faecium among broilers. J. Antimicrob. Chemother. 2009, 63, 868–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, O.; Alm, E.; Greko, C.; Bengtsson, B. The rise and fall of a vancomycin-resistant clone of Enterococcus faecium among broilers in Sweden. J. Glob. Antimicrob. Resist. 2019, 17, 233–235. [Google Scholar] [CrossRef] [PubMed]
- Robredo, B.; Torres, C.; Singh, K.V.; Murray, B. Molecular analysis of Tn1546 in vanA-containing Enterococcus spp. isolated from humans and poultry. Antimicrob. Agents Chemother. 2000, 44, 2588–2589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vignaroli, C.; Zandri, G.; Aquilanti, L.; Pasquaroli, S.; Biavasco, F. Multidrug-resistant enterococci in animal meat and faeces and co-transfer of resistance from an Enterococcus durans to a human Enterococcus faecium. Curr. Microbiol. 2011, 62, 1438–1447. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Host/Species | No. of Strains | Resistance Phenotype | Resistance Genotype | MLST | ||
|---|---|---|---|---|---|---|
| MIC [µg/mL] Vancomycin | Additional Resistances1 | Resistance Genes | Tn1546 Type | |||
| Cattle | ||||||
| E. faecalis | 1 | ≥128 | – | dfrE, emeA, efrA, efrB, lsaA, vanA | I | 29 |
| Pigs | ||||||
| E. faecium | 1 | ≥256 | PEN, ERY, TE | aac(6′)-Ii, eat(A)v, cadA, cadC, copZ, czrA, merA, merR, tetW/N/W, vanA, zosA | I | 133 |
| E. faecium | 5 | ≥256 | PEN, TE | aac(6′)-Ii, eat(A)v, cadA, cadC, copZ, czrA, merA, merR, tetW/N/W, vanA, zosA | I | 133 |
| Poultry | ||||||
| E. faecium | 1 | ≥256 | ERY | aac(6′)-Ii, aadK, eat(A)v, vanA | I | 13 |
| E. faecium | 1 | ≥256 | PEN | aac(6′)-Ii, aadK, eat(A)v, vanA | I | 157 |
| E. faecium | 1 | ≥256 | – | aac(6′)-Ii, aadK, eat(A)v, vanA | I | 157 |
| E. faecium | 3 | ≥256 | ERY | aac(6′)-Ii, aadK, eat(A)v, vanA | I | 310 |
| E. durans | 1 | ≥256 | TE | aac(6′)-Iid, tetW/N/W, vanA | I | – |
| E. durans | 2 | ≥256 | ERY, TE | aac(6′)-Iid, ermB, vanA | I | – |
| E. durans | 1 | 256 | ERY, TE | aac(6′)-Iid, ermB tetW/N/W, vanA | I | – |
| E. durans | 1 | ≥256 | TE | aac(6′)-Iid, ermB, tetW/N/W, vanA | I | – |
| E. durans | 1 | ≥256 | ERY, TE | aac(6′)-Iid, ermB tetW/N/W, vanA | I | – |
| E. durans | 3 | ≥256 | ERY, TE | aac(6′)-Iid, ermB tetW/N/W, vanA | II | – |
| E. durans | 1 | ≥256 | TE | aac(6′)-Iid, tetW/N/W, vanA | II | – |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wist, V.; Morach, M.; Schneeberger, M.; Cernela, N.; Stevens, M.J.A.; Zurfluh, K.; Stephan, R.; Nüesch-Inderbinen, M. Phenotypic and Genotypic Traits of Vancomycin-Resistant Enterococci from Healthy Food-Producing Animals. Microorganisms 2020, 8, 261. https://doi.org/10.3390/microorganisms8020261
Wist V, Morach M, Schneeberger M, Cernela N, Stevens MJA, Zurfluh K, Stephan R, Nüesch-Inderbinen M. Phenotypic and Genotypic Traits of Vancomycin-Resistant Enterococci from Healthy Food-Producing Animals. Microorganisms. 2020; 8(2):261. https://doi.org/10.3390/microorganisms8020261
Chicago/Turabian StyleWist, Valerie, Marina Morach, Marianne Schneeberger, Nicole Cernela, Marc J.A. Stevens, Katrin Zurfluh, Roger Stephan, and Magdalena Nüesch-Inderbinen. 2020. "Phenotypic and Genotypic Traits of Vancomycin-Resistant Enterococci from Healthy Food-Producing Animals" Microorganisms 8, no. 2: 261. https://doi.org/10.3390/microorganisms8020261