
Evaluation of Pyrophosphate-Driven Proton Pumps in Saccharomyces cerevisiae under Stress Conditions
 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Maintenance
2.2. Plasmid Construction
2.3. Yeast Strain Engineering
2.4. Microtiter Plate Cultures
2.5. Growth Studies in Bioreactors (or Growth Studies or Cultivations)
2.6. Analytical Methods
2.7. Flow Cytometry
2.8. Calculations
3. Results and Discussions
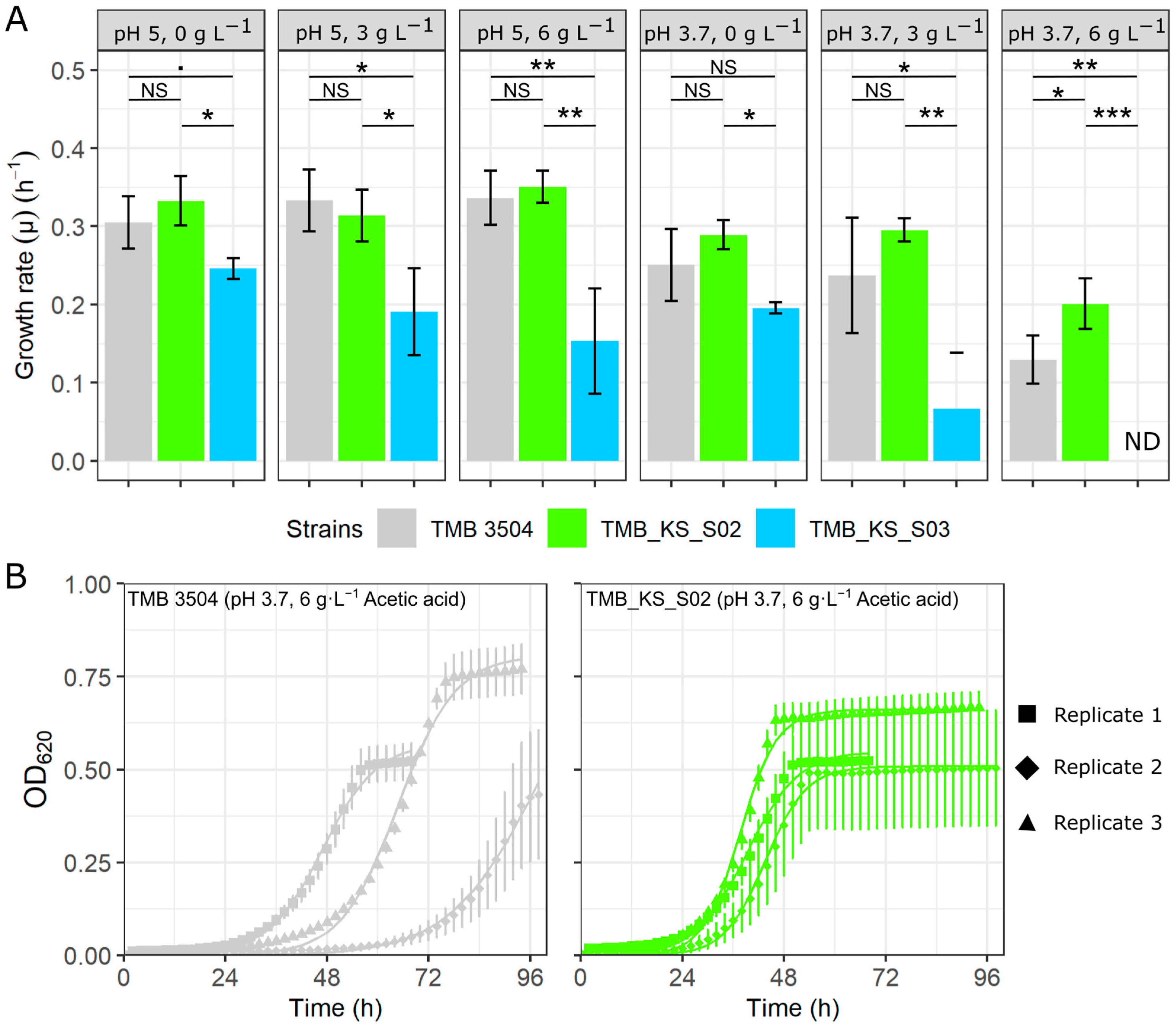
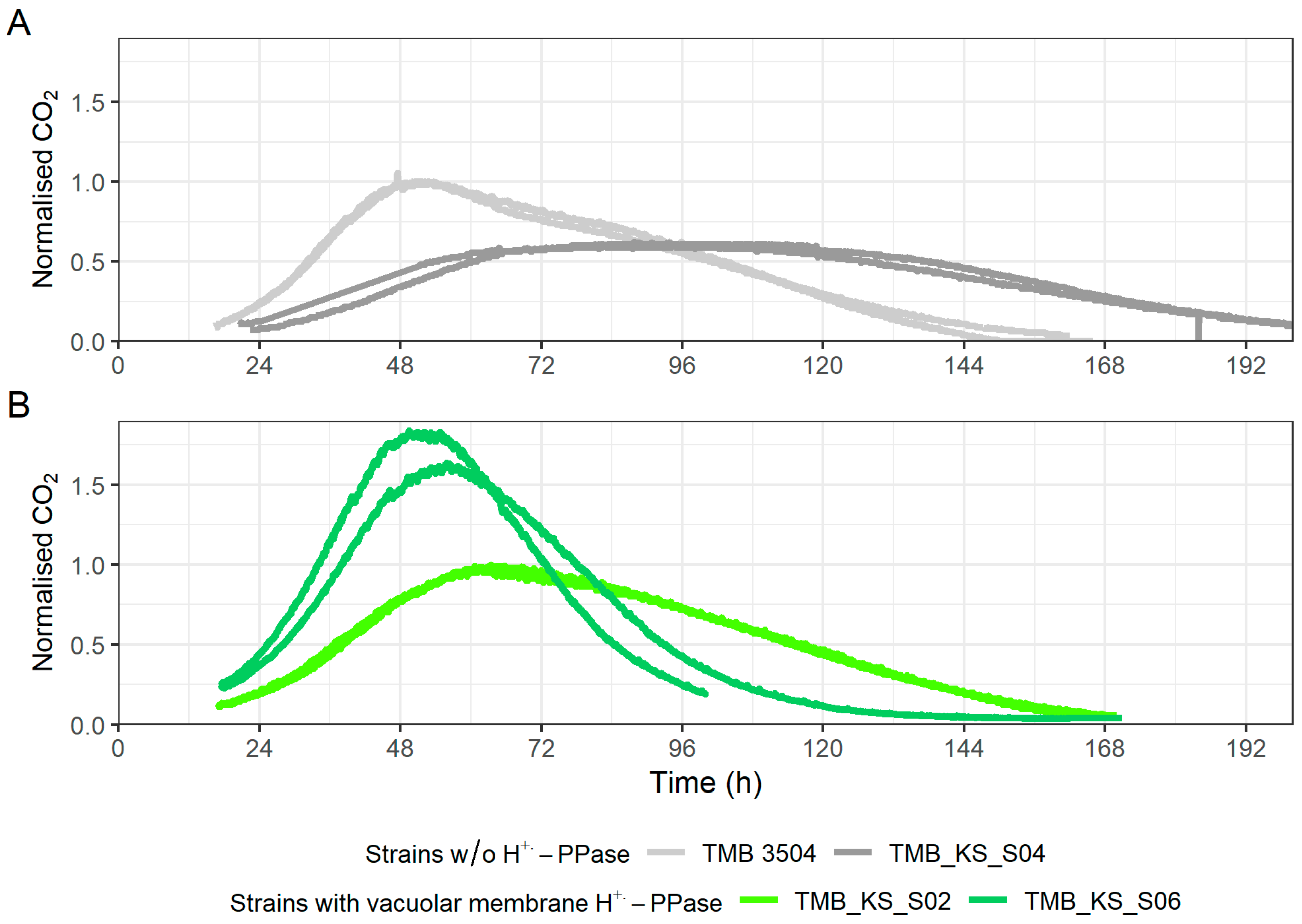
3.1. Vacuolar Membrane H+-PPase Improved Growth Rates at a Low pH and Acetic Acid Stress
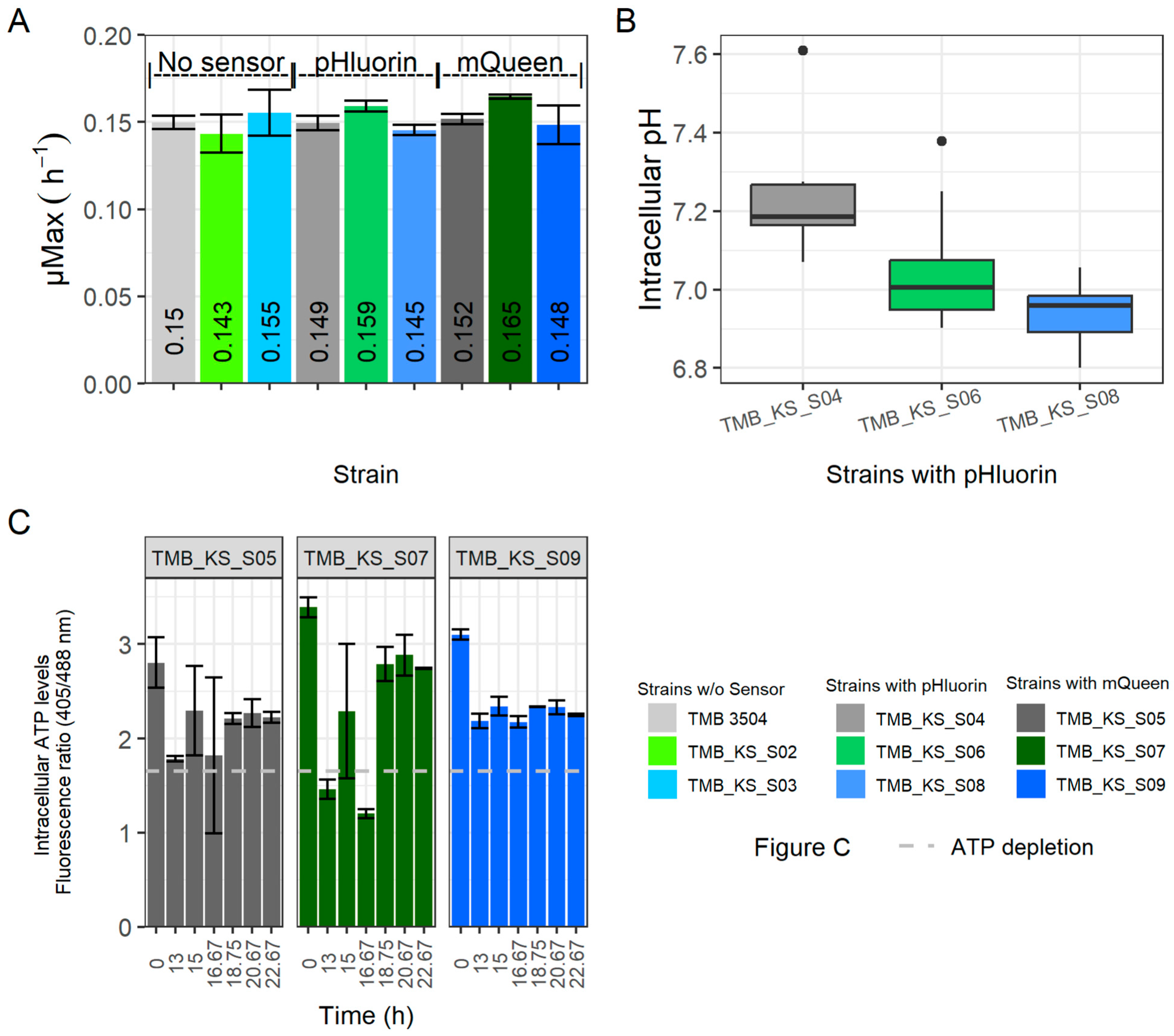
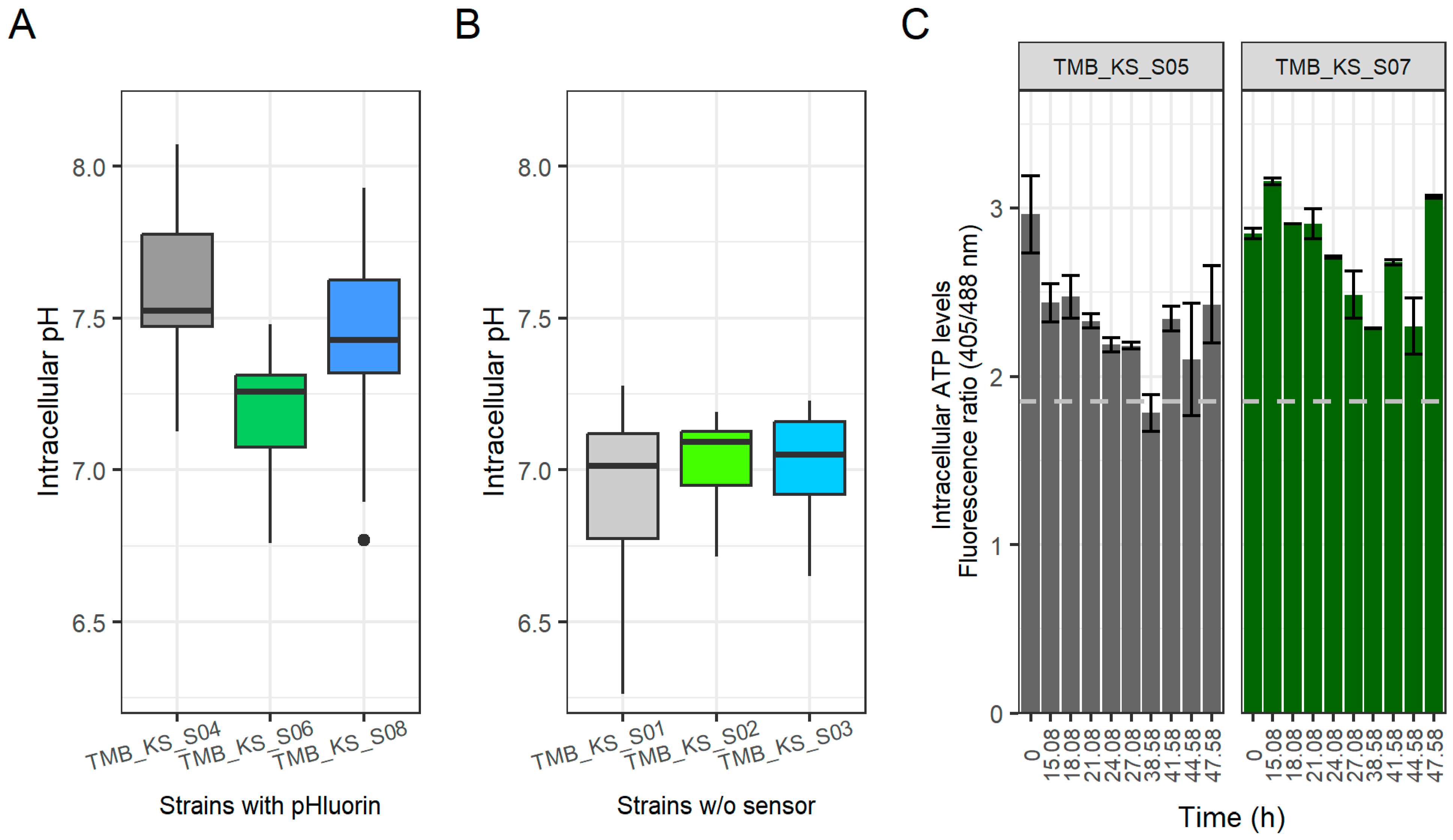
3.2. Expression of H+-PPase Led to an Acidified Cytoplasm during Glucose Fermentations
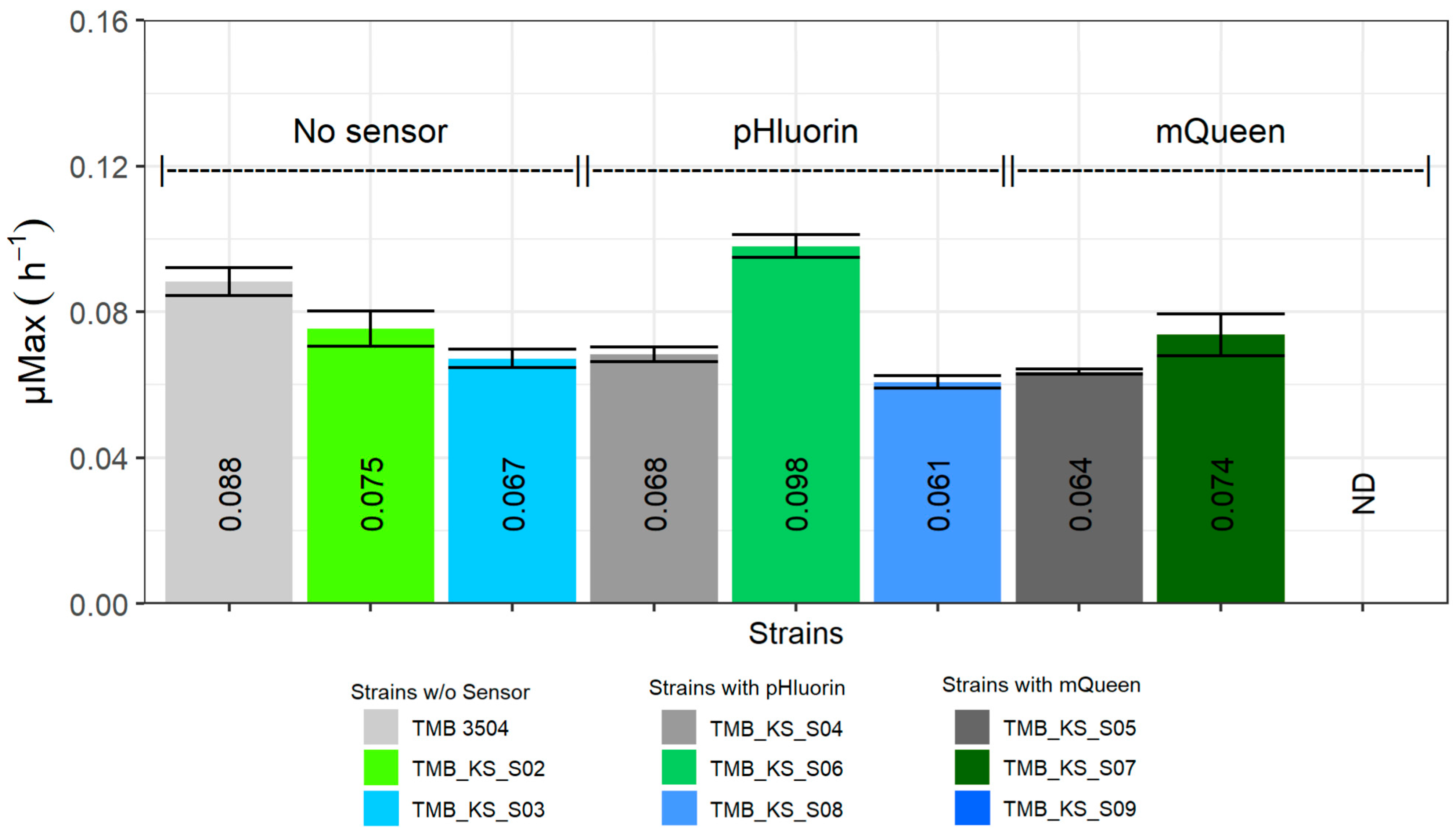
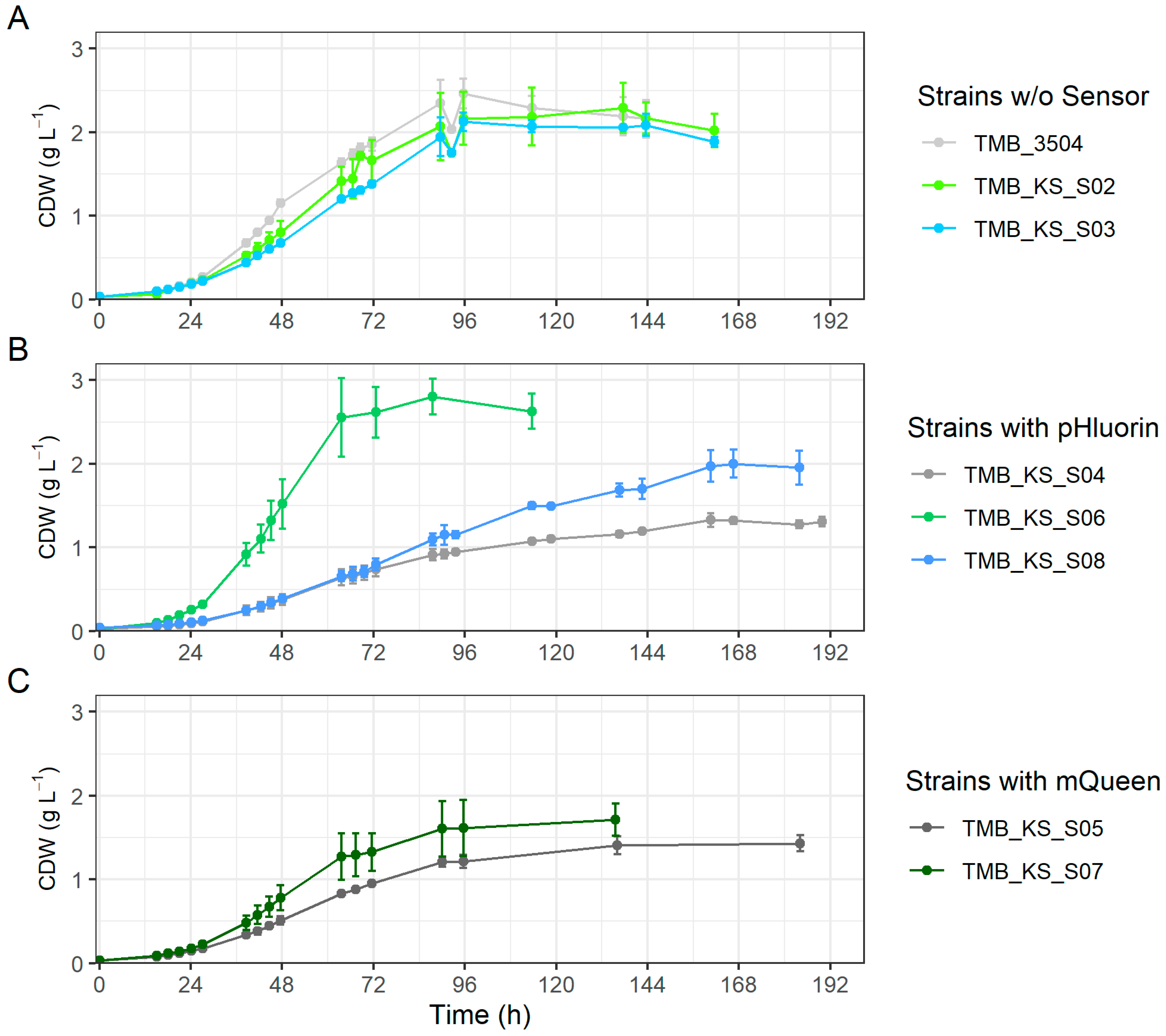
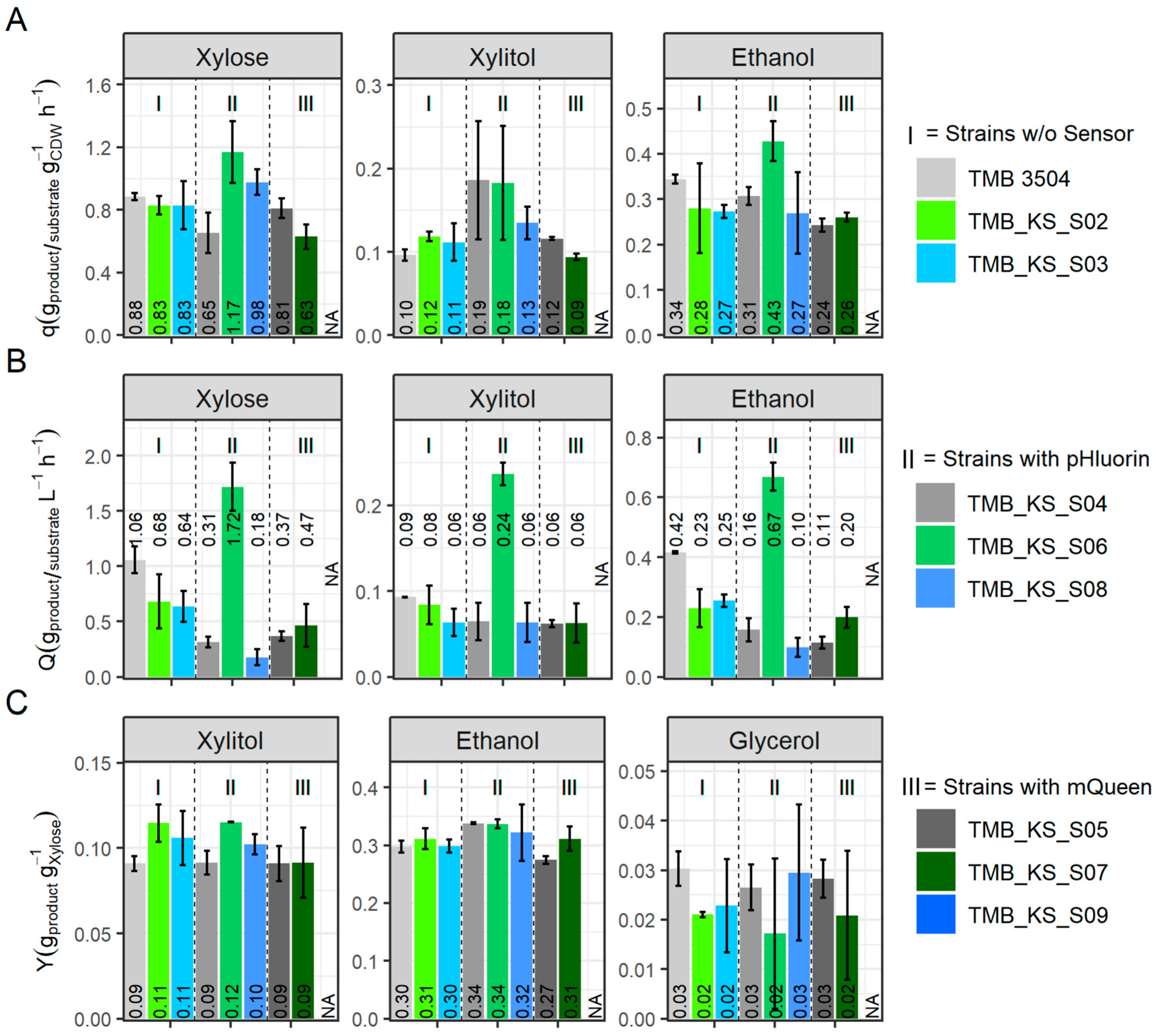
3.3. Vacuolar Membrane H+-PPase Improved Xylose Fermentation When pHluorin Is Co-Expressed
3.4. Cell Morphology Is Influenced by Biosensor Expression on Xylose
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hayakawa, K.; Matsuda, F.; Shimizu, H. Metabolome Analysis of Saccharomyces cerevisiae and Optimization of Culture Medium for S-Adenosyl-l-Methionine Production. AMB Express 2016, 6, 38. [Google Scholar] [CrossRef]
- Xu, R.; Wang, D.; Wang, C.; Zhang, G.; Wei, G. Improved S-Adenosylmethionine and Glutathione Biosynthesis by Heterologous Expression of an ATP6 Gene in Candida utilis. J. Basic Microbiol. 2018, 58, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, D.; Zhang, G.; Wang, C.; Wei, G. Disruption of por1 Gene in Candida utilis Improves Co-Production of S-Adenosylmethionine and Glutathione. J. Biotechnol. 2019, 290, 16–23. [Google Scholar] [CrossRef]
- Chen, Y.; Tan, T. Enhanced S-Adenosylmethionine Production by Increasing ATP Levels in Baker’s Yeast (Saccharomyces cerevisiae). J. Agric. Food Chem. 2018, 66, 5200–5209. [Google Scholar] [CrossRef]
- Sahoo, A.; Das, P.K.; Patra, S.; Veeranki, V.D. Engineered Yeasts for the Production of Biofuel and Platform Chemicals. In Advances in Yeast Biotechnology for Biofuels and Sustainability; Elsevier: Amsterdam, The Netherlands, 2023; pp. 21–46. ISBN 978-0-323-95449-5. [Google Scholar]
- Meadows, A.L.; Hawkins, K.M.; Tsegaye, Y.; Antipov, E.; Kim, Y.; Raetz, L.; Dahl, R.H.; Tai, A.; Mahatdejkul-Meadows, T.; Xu, L.; et al. Rewriting Yeast Central Carbon Metabolism for Industrial Isoprenoid Production. Nature 2016, 537, 694–697. [Google Scholar] [CrossRef]
- Liu, H.; Marsafari, M.; Wang, F.; Deng, L.; Xu, P. Engineering Acetyl-CoA Metabolic Shortcut for Eco-Friendly Production of Polyketides Triacetic Acid Lactone in Yarrowia lipolytica. Metab. Eng. 2019, 56, 60–68. [Google Scholar] [CrossRef]
- Daicho, K.M.; Hirono-Hara, Y.; Kikukawa, H.; Tamura, K.; Hara, K.Y. Engineering Yeast with a Light-Driven Proton Pump System in the Vacuolar Membrane. Microb. Cell Fact. 2024, 23, 4. [Google Scholar] [CrossRef]
- Hara, K.Y.; Kondo, A. ATP Regulation Inbioproduction. Microb. Cell Fact. 2015, 14, 198. [Google Scholar] [CrossRef] [PubMed]
- Orij, R.; Brul, S.; Smits, G.J. Intracellular PH Is a Tightly Controlled Signal in Yeast. Biochim. Biophys. Acta (BBA) Gen. Subj. 2011, 1810, 933–944. [Google Scholar] [CrossRef]
- Martínez-Muñoz, G.A.; Kane, P. Vacuolar and Plasma Membrane Proton Pumps Collaborate to Achieve Cytosolic PH Homeostasis in Yeast. J. Biol. Chem. 2008, 283, 20309–20319. [Google Scholar] [CrossRef]
- Pampulha, M.E.; Loureiro-Dias, M.C. Energetics of the Effect of Acetic Acid on Growth of Saccharomyces cerevisiae. FEMS Microbiol. Lett. 2000, 184, 69–72. [Google Scholar] [CrossRef]
- Bielen, A.A.M.; Willquist, K.; Engman, J.; Van Der Oost, J.; Van Niel, E.W.J.; Kengen, S.W.M. Pyrophosphate as a Central Energy Carrier in the Hydrogen-Producing Extremely Thermophilic Caldicellulosiruptor saccharolyticus. FEMS Microbiol. Lett. 2010, 307, 48–54. [Google Scholar] [CrossRef]
- Khadilkar, A.S.; Yadav, U.P.; Salazar, C.; Shulaev, V.; Paez-Valencia, J.; Pizzio, G.A.; Gaxiola, R.A.; Ayre, B.G. Constitutive and Companion Cell-Specific Overexpression of AVP1, Encoding a Proton-Pumping Pyrophosphatase, Enhances Biomass Accumulation, Phloem Loading, and Long-Distance Transport. Plant Physiol. 2016, 170, 401–414. [Google Scholar] [CrossRef]
- Lahti, R. Microbial Inorganic Pyrophosphatases. Microbiol. Rev. 1983, 47, 169–178. [Google Scholar] [CrossRef]
- Lahti, R.; Kolakowski, L.F.; Heinonen, J.; Vihinen, M.; Pohjanoksa, K.; Cooperman, B.S. Conservation of Functional Residues between Yeast and E. coli Inorganic Pyrophosphatases. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 1990, 1038, 338–345. [Google Scholar] [CrossRef]
- Serrano-Bueno, G.; Madroñal, J.M.; Manzano-López, J.; Muñiz, M.; Pérez-Castiñeira, J.R.; Hernández, A.; Serrano, A. Nuclear Proteasomal Degradation of Saccharomyces cerevisiae Inorganic Pyrophosphatase Ipp1p, a Nucleocytoplasmic Protein Whose Stability Depends on Its Subcellular Localization. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2019, 1866, 1019–1033. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Bueno, G.; Hernández, A.; López-Lluch, G.; Pérez-Castiñeira, J.R.; Navas, P.; Serrano, A. Inorganic Pyrophosphatase Defects Lead to Cell Cycle Arrest and Autophagic Cell Death through NAD+ Depletion in Fermenting Yeast. J. Biol. Chem. 2013, 288, 13082–13092. [Google Scholar] [CrossRef]
- da-Silva, W.S.; Bomfim, F.M.; Galina, A.; de Meis, L. Heat of PPi Hydrolysis Varies Depending on the Enzyme Used: Yeast and Corn Vacuolar Pyrophosphatase. J. Biol. Chem. 2004, 279, 45613–45617. [Google Scholar] [CrossRef] [PubMed]
- Potapenko, E.; Cordeiro, C.D.; Huang, G.; Docampo, R. Pyrophosphate Stimulates the Phosphate-Sodium Symporter of Trypanosoma brucei Acidocalcisomes and Saccharomyces cerevisiae Vacuoles. mSphere 2019, 4, e00045-19. [Google Scholar] [CrossRef] [PubMed]
- Ermakova, S.A.; Mansurova, S.E.; Kalebina, T.S.; Lobakova, E.S.; Selyach, I.O.; Kulaev, I.S. Accumulation of Pyrophosphate and Other Energy-Rich Phosphorus Compounds under Various Conditions of Yeast Growth. Arch. Microbiol. 1981, 128, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Carmelo, V.; Santos, H.; Sá-Correia, I. Effect of Extracellular Acidification on the Activity of Plasma Membrane ATPase and on the Cytosolic and Vacuolar PH of Saccharomyces cerevisiae. Biochim. Biophys. Acta (BBA) Biomembr. 1997, 1325, 63–70. [Google Scholar] [CrossRef]
- Carmelo, V.; Bogaerts, P.; Sá-Correia, I. Activity of Plasma Membrane H+-ATPase and Expression of PMA1 and PMA2 Genes in Saccharomyces cerevisiae Cells Grown at Optimal and Low PH. Arch. Microbiol. 1996, 166, 315–320. [Google Scholar] [CrossRef]
- van Niel, E.W.J.; Bergdahl, B.; Hahn-Hägerdal, B. Close to the Edge: Growth Restrained by the NAD(P)H/ATP Formation Flux Ratio. Front. Microbiol. 2017, 8, 01149. [Google Scholar] [CrossRef] [PubMed]
- Sonderegger, M.; Jeppsson, M.; Hahn-Hagerdal, B.; Sauer, U. Molecular Basis for Anaerobic Growth of Saccharomyces cerevisiae on Xylose, Investigated by Global Gene Expression and Metabolic Flux Analysis. Appl. Environ. Microbiol. 2004, 70, 2307–2317. [Google Scholar] [CrossRef]
- Klinke, H.B.; Thomsen, A.B.; Ahring, B.K. Inhibition of Ethanol-Producing Yeast and Bacteria by Degradation Products Produced during Pre-Treatment of Biomass. Appl. Microbiol. Biotechnol. 2004, 66, 10–26. [Google Scholar] [CrossRef] [PubMed]
- Guaragnella, N.; Bettiga, M. Acetic Acid Stress in Budding Yeast: From Molecular Mechanisms to Applications. Yeast 2021, 38, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Nasution, O.; Lee, Y.M.; Kim, E.; Choi, W.; Kim, W. Overexpression of PMA1 Enhances Tolerance to Various Types of Stress and Constitutively Activates the SAPK Pathways in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2017, 101, 229–239. [Google Scholar] [CrossRef]
- Sànchez, I.; Nogué, V.; Narayanan, V.; Gorwa-Grauslund, M.F. Short-Term Adaptation Improves the Fermentation Performance of Saccharomyces cerevisiae in the Presence of Acetic Acid at Low PH. Appl. Microbiol. Biotechnol. 2013, 97, 7517–7525. [Google Scholar] [CrossRef]
- Moysés, D.N.; Reis, V.C.B.; Almeida, J.R.M.d.; de Moraes, L.M.P.; Torres, F.A.G. Xylose Fermentation by Saccharomyces cerevisiae: Challenges and Prospects. Int. J. Mol. Sci. 2016, 17, 207. [Google Scholar] [CrossRef]
- Serrano, R. In Vivo Glucose Activation of the Yeast Plasma Membrane ATPase. FEBS Lett. 1983, 156, 11–14. [Google Scholar] [CrossRef]
- Nijland, J.G.; Zhang, X.; Driessen, A.J.M. D-Xylose Accelerated Death of Pentose Metabolizing Saccharomyces cerevisiae. Biotechnol. Biofuels Bioprod. 2023, 16, 67. [Google Scholar] [CrossRef]
- Peng, B.; Williams, T.C.; Henry, M.; Nielsen, L.K.; Vickers, C.E. Controlling Heterologous Gene Expression in yeast Cell Factories on Different Carbon Substrates and across the Diauxic Shift: Acomparison of Yeast Promoter Activities. Microb. Cell Fact. 2015, 14, 91. [Google Scholar] [CrossRef] [PubMed]
- Drake, R.; Serrano, A.; Pérez-Castiñeira, J.R. N-Terminal Chimaeras with Signal Sequences Enhance the Functional Expression and Alter the Subcellular Localization of Heterologous Membrane-Bound Inorganic Pyrophosphatases in Yeast. Biochem. J. 2010, 426, 147–157. [Google Scholar] [CrossRef]
- Yoon, H.-S.; Kim, S.-Y.; Kim, I.-S. Stress Response of Plant H+-PPase-Expressing Transgenic Escherichia coli and Saccharomyces cerevisiae: A Potentially Useful Mechanism for the Development of Stress-Tolerant Organisms. J. Appl. Genet. 2013, 54, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.; Herrera-Palau, R.; Madroñal, J.M.; Albi, T.; López-Lluch, G.; Perez-Castiñeira, J.R.; Navas, P.; Valverde, F.; Serrano, A. Vacuolar H+-Pyrophosphatase AVP1 Is Involved in Amine Fungicide Tolerance in Arabidopsis thaliana and Provides Tridemorph Resistance in Yeast. Front. Plant Sci. 2016, 7, 00085. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Zhen, R.G.; Rea, P.A. Heterologous Expression of Plant Vacuolar Pyrophosphatase in Yeast Demonstrates Sufficiency of the Substrate-Binding Subunit for Proton Transport. Proc. Natl. Acad. Sci. USA 1994, 91, 6128–6132. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Inuzuka, M.; Ingi, T. Cellular Signaling Mediated by Calphoglin-Induced Activation of IPP and PGM. Biochem. Biophys. Res. Commun. 2004, 325, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Orij, R.; Postmus, J.; Ter Beek, A.; Brul, S.; Smits, G.J. In Vivo Measurement of Cytosolic and Mitochondrial PH Using a PH-Sensitive GFP Derivative in Saccharomyces cerevisiae Reveals a Relation between Intracellular PH and Growth. Microbiology 2009, 155, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Yaginuma, H.; Kawai, S.; Tabata, K.V.; Tomiyama, K.; Kakizuka, A.; Komatsuzaki, T.; Noji, H.; Imamura, H. Diversity in ATP Concentrations in a Single Bacterial Cell Population Revealed by Quantitative Single-Cell Imaging. Sci. Rep. 2014, 4, 6522. [Google Scholar] [CrossRef]
- Takaine, M.; Ueno, M.; Kitamura, K.; Imamura, H.; Yoshida, S. Reliable Imaging of ATP in Living Budding and Fission Yeast. J. Cell Sci. 2019, 132, jcs230649. [Google Scholar] [CrossRef]
- Inoue, H.; Nojima, H.; Okayama, H. High Efficiency Transformation of Escherichia coli with Plasmids. Gene 1990, 96, 23–28. [Google Scholar] [CrossRef]
- Cadete, R.M.; de las Heras, A.M.; Sandström, A.G.; Ferreira, C.; Gírio, F.; Gorwa-Grauslund, M.-F.; Rosa, C.A.; Fonseca, C. Exploring Xylose Metabolism in Spathaspora Species: XYL1.2 from Spathaspora passalidarum as the Key for Efficient Anaerobic Xylose Fermentation in Metabolic Engineered Saccharomyces cerevisiae. Biotechnol. Biofuels 2016, 9, 167. [Google Scholar] [CrossRef] [PubMed]
- Lennox, E.S. Transduction of Linked Genetic Characters of the Host by Bacteriophage P1. Virology 1955, 1, 190–206. [Google Scholar] [CrossRef]
- Mumberg, D.; Müller, R.; Funk, M. Yeast Vectors for the Controlled Expression of Heterologous Proteins in Different Genetic Backgrounds. Gene 1995, 156, 119–122. [Google Scholar] [CrossRef]
- Jessop-Fabre, M.M.; Jakočiūnas, T.; Stovicek, V.; Dai, Z.; Jensen, M.K.; Keasling, J.D.; Borodina, I. EasyClone-MarkerFree: A Vector Toolkit for Marker-less Integration of Genes into Saccharomyces cerevisiae via CRISPR-Cas9. Biotechnol. J. 2016, 11, 1110–1117. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. High-Efficiency Yeast Transformation Using the LiAc/SS Carrier DNA/PEG Method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef]
- Verduyn, C.; Postma, E.; Scheffers, W.A.; Van Dijken, J.P. Effect of Benzoic Acid on Metabolic Fluxes in Yeasts: A Continuous-Culture Study on the Regulation of Respiration and Alcoholic Fermentation. Yeast 1992, 8, 501–517. [Google Scholar] [CrossRef] [PubMed]
- Sprouffske, K.; Wagner, A. Growthcurver: An R Package for Obtaining Interpretable Metrics from Microbial Growth Curves. BMC Bioinform. 2016, 17, 172. [Google Scholar] [CrossRef]
- Rice, J.F.; Sullivan, T.R.; Helbert, J.R. A Rapid Method for the Determination of Yeast Dry Weight Concentration. J. Am. Soc. Brew. Chem. 1980, 38, 142–145. [Google Scholar] [CrossRef]
- Borgström, C.; Persson, V.C.; Rogova, O.; Osiro, K.O.; Lundberg, E.; Spégel, P.; Gorwa-Grauslund, M. Using Phosphoglucose Isomerase-Deficient (pgi1Δ) Saccharomyces cerevisiae to Map the Impact of Sugar Phosphate Levels on d-Glucose and d-Xylose Sensing. Microb. Cell Fact. 2022, 21, 253. [Google Scholar] [CrossRef]
- Torello Pianale, L.; Rugbjerg, P.; Olsson, L. Real-Time Monitoring of the Yeast Intracellular State During Bioprocesses with a Toolbox of Biosensors. Front. Microbiol. 2021, 12, 802169. [Google Scholar] [CrossRef] [PubMed]
- Lecchi, S.; Nelson, C.J.; Allen, K.E.; Swaney, D.L.; Thompson, K.L.; Coon, J.J.; Sussman, M.R.; Slayman, C.W. Tandem Phosphorylation of Ser-911 and Thr-912 at the C Terminus of Yeast Plasma Membrane H+-ATPase Leads to Glucose-Dependent Activation. J. Biol. Chem. 2007, 282, 35471–35481. [Google Scholar] [CrossRef] [PubMed]
- Tenreiro, S.; Rosa, P.C.; Viegas, C.A.; Sá-Correia, I. Expression of the AZR1 Gene (ORF YGR224w), Encoding a Plasma Membrane Transporter of the Major Facilitator Superfamily, Is Required for Adaptation to Acetic Acid and Resistance to Azoles in Saccharomyces cerevisiae. Yeast 2000, 16, 1469–1481. [Google Scholar] [CrossRef]
- Scholz-Starke, J.; Primo, C.; Yang, J.; Kandel, R.; Gaxiola, R.A.; Hirschi, K.D. The Flip Side of the Arabidopsis Type I Proton-Pumping Pyrophosphatase (AVP1): Using a Transmembrane H+ Gradient to Synthesize Pyrophosphate. J. Biol. Chem. 2019, 294, 1290–1299. [Google Scholar] [CrossRef]
- de Ruijter, J.C.; Koskela, E.V.; Nandania, J.; Frey, A.D.; Velagapudi, V. Understanding the Metabolic Burden of Recombinant Antibody Production in Saccharomyces cerevisiae Using a Quantitative Metabolomics Approach. Yeast 2018, 35, 331–341. [Google Scholar] [CrossRef]
- Heyland, J.; Fu, J.; Blank, L.M.; Schmid, A. Carbon Metabolism Limits Recombinant Protein Production in Pichia pastoris. Biotechnol. Bioeng. 2011, 108, 1942–1953. [Google Scholar] [CrossRef]
- Krogh, A.M.; Beck, V.; Christensen, L.H.; Henriksen, C.M.; Møller, K.; Olsson, L. Adaptation of Saccharomyces cerevisiae Expressing a Heterologous Protein. J. Biotechnol. 2008, 137, 28–33. [Google Scholar] [CrossRef]
- Kastberg, L.L.B.; Ard, R.; Jensen, M.K.; Workman, C.T. Burden Imposed by Heterologous Protein Production in Two Major Industrial Yeast Cell Factories: Identifying Sources and Mitigation Strategies. Front. Fungal. Biol. 2022, 3, 827704. [Google Scholar] [CrossRef]
- Isom, D.G.; Page, S.C.; Collins, L.B.; Kapolka, N.J.; Taghon, G.J.; Dohlman, H.G. Coordinated Regulation of Intracellular PH by Two Glucose-Sensing Pathways in Yeast. J. Biol. Chem. 2018, 293, 2318–2329. [Google Scholar] [CrossRef]
- Dechant, R.; Binda, M.; Lee, S.S.; Pelet, S.; Winderickx, J.; Peter, M. Cytosolic PH Is a Second Messenger for Glucose and Regulates the PKA Pathway through V-ATPase. EMBO J. 2010, 29, 2515–2526. [Google Scholar] [CrossRef] [PubMed]
- Serrano, R. Energy Requirements for Maltose Transport in Yeast. Eur. J. Biochem. 1977, 80, 97–102. [Google Scholar] [CrossRef]
- Hervé, M.; Wietzerbin, J.; Lebourguais, O.; Tran-Dinh, S. Effects of 2-Deoxy-d-Glucose on the Glucose Metabolism in Saccharomyces cerevisiae Studied by Multinuclear-NMR Spectroscopy and Biochemical Methods. Biochimie 1992, 74, 1103–1115. [Google Scholar] [CrossRef] [PubMed]
- Kafri, M.; Metzl-Raz, E.; Jona, G.; Barkai, N. The Cost of Protein Production. Cell Rep. 2015, 14, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Zeng, Y.; Tang, R.-Q.; Alper, H.S.; Bai, F.-W.; Zhao, X.-Q. Condition-Specific Promoter Activities in Saccharomyces cerevisiae. Microb. Cell Fact. 2018, 17, 58. [Google Scholar] [CrossRef] [PubMed]
- Osiro, K.O.; Brink, D.P.; Borgström, C.; Wasserstrom, L.; Carlquist, M.; Gorwa-Grauslund, M.F. Assessing the Effect of D-Xylose on the Sugar Signaling Pathways of Saccharomyces cerevisiae in Strains Engineered for Xylose Transport and Assimilation. FEMS Yeast Res. 2018, 18, fox096. [Google Scholar] [CrossRef] [PubMed]
- Brink, D.P.; Borgström, C.; Tueros, F.G.; Gorwa-Grauslund, M.F. Real-Time Monitoring of the Sugar Sensing in Saccharomyces cerevisiae Indicates Endogenous Mechanisms for Xylose Signaling. Microb. Cell Fact. 2016, 15, 183. [Google Scholar] [CrossRef] [PubMed]
- Bakker, B.M.; Overkamp, K.M.; van Maris, A.J.A.; Kötter, P.; Luttik, M.A.H.; van Dijken, J.P.; Pronk, J.T. Stoichiometry and Compartmentation of NADH Metabolism in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2001, 25, 15–37. [Google Scholar] [CrossRef]
- Lohmeier-Vogel, E.M.; Hahn-Hägerdal, B.; Vogel, H.J. Phosphorus-31 and Carbon-13 Nuclear Magnetic Resonance Studies of Glucose and Xylose Metabolism in Candida tropicalis Cell Suspensions. Appl. Environ. Microbiol. 1995, 61, 1414–1419. [Google Scholar] [CrossRef] [PubMed]
- Lohmeier-Vogel, E.M.; McIntyre, D.D.; Vogel, H.J. Phosphorus-31 and Carbon-13 Nuclear Magnetic Resonance Studies of Glucose and Xylose Metabolism in Cell Suspensions and Agarose-Immobilized Cultures of Pichia stipitis and Saccharomyces cerevisiae. Appl. Enviorn. Microbiol. 1996, 62, 2832–2838. [Google Scholar] [CrossRef]
- Eskes, E.; Deprez, M.-A.; Wilms, T.; Winderickx, J. PH Homeostasis in Yeast; the Phosphate Perspective. Curr. Genet. 2018, 64, 155–161. [Google Scholar] [CrossRef]
- Toivari, M.; Nygård, Y.; Kumpula, E.-P.; Vehkomäki, M.-L.; Benčina, M.; Valkonen, M.; Maaheimo, H.; Andberg, M.; Koivula, A.; Ruohonen, L.; et al. Metabolic Engineering of Saccharomyces cerevisiae for Bioconversion of D-Xylose to d-Xylonate. Metab. Eng. 2012, 14, 427–436. [Google Scholar] [CrossRef]
- Ostergaard, S.; Olsson, L.; Nielsen, J. In Vivo Dynamics of Galactose Metabolism in Saccharomyces cerevisiae: Metabolic Fluxes and Metabolite Levels. Biotechnol. Bioeng. 2001, 73, 412–425. [Google Scholar] [CrossRef]
- Maslanka, R.; Zadrag-Tecza, R. Reproductive Potential of Yeast Cells Depends on Overall Action of Interconnected Changes in Central Carbon Metabolism, Cellular Biosynthetic Capacity, and Proteostasis. Int. J. Mol. Sci. 2020, 21, 7313. [Google Scholar] [CrossRef]
- Vemuri, G.N.; Eiteman, M.A.; McEwen, J.E.; Olsson, L.; Nielsen, J. Increasing NADH Oxidation Reduces Overflow Metabolism in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2007, 104, 2402–2407. [Google Scholar] [CrossRef]
- van den Brink, J.; Canelas, A.B.; van Gulik, W.M.; Pronk, J.T.; Heijnen, J.J.; de Winde, J.H.; Daran-Lapujade, P. Dynamics of Glycolytic Regulation during Adaptation of Saccharomyces cerevisiae to Fermentative Metabolism. Appl. Environ. Microbiol. 2008, 74, 5710–5723. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, Y.; Makanae, K.; Hasunuma, T.; Ishibashi, Y.; Kito, K.; Moriya, H. Estimating the Protein Burden Limit of Yeast Cells by Measuring the Expression Limits of Glycolytic Proteins. eLife 2018, 7, e34595. [Google Scholar] [CrossRef]
- Kintaka, R.; Makanae, K.; Moriya, H. Cellular Growth Defects Triggered by an Overload of Protein Localization Processes. Sci. Rep. 2016, 6, 31774. [Google Scholar] [CrossRef] [PubMed]
- Bergdahl, B.; Heer, D.; Sauer, U.; Hahn-Hägerdal, B.; van Niel, E.W. Dynamic Metabolomics Differentiates between Carbon and Energy Starvation in Recombinant Saccharomyces cerevisiae Fermenting Xylose. Biotechnol. Biofuels 2012, 5, 34. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.R.; Sá-Correia, I. Transcription Patterns of PMA1 and PMA2 Genes and Activity of Plasma Membrane H+-ATPase in Saccharomyces cerevisiae during Diauxic Growth and Stationary Phase. Yeast 2003, 20, 207–219. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Designation | Strain Name | Relevant Genotype | References |
|---|---|---|---|
| TMB 3504 | Parent Strain | CEN.PK 2-1C; MATa; ura3-52; Δgre3; his3::HIS3 PGK1p-XKS1-PGK1t; TAL1::PGK1p-TAL1-PGK1t; TKL1::PGK1p-TKL1-PGK1t; RKI1::PGK1p-RKI1-PGK1t; RPE1::PGK1p-RPE1-PGK1t; ura3::YIpRC5p | [43] |
| TMB_KS_S02 | Vacuolar membrane H+-PPase strain | TMB 3504; XI-3::TEF1p-(Tc)AVP1-CYC7t | This study |
| TMB_KS_S03 | Cell membrane H+-PPase strain | TMB 3504; XI-3::TEF1p-(Suc2)AVP1-CYC7t | This study |
| TMB_KS_S04 | Parent strain with pHluorin | TMB 3504; X-4::GPD1p-pHluorin-CYC7t | This study |
| TMB_KS_S05 | Parent strain with mQueen | TMB 3504; X-4::GPD1p-mQueen2m-CYC7t | This study |
| TMB_KS_S06 | Vacuolar membrane H+-PPase strain with pHluorin | TMB KS S02; X-4::GPD1p-pHluorin-CYC7t | This study |
| TMB_KS_S07 | Vacuolar membrane H+-PPase strain with mQueen | TMB KS S02; X-4::GPD1p-mQueen2m-CYC7t | This study |
| TMB_KS_S08 | Cell membrane H+-PPase strain with pHluorin | TMB KS S03; X-4::GPD1p-pHluorin-CYC7t | This study |
| TMB_KS_S09 | Cell membrane H+-PPase strain with mQueen | TMB KS S03; X-4::GPD1p-mQueen2m-CYC7t | This study |
| Name | Relevant Genotype | Source |
|---|---|---|
| pRSET-QUE2m | ColE1; AmpR; T7p-QUEEN-2m-T7t | [40] [Addgene; #129350] |
| pUC57-VP-Suc2PSP-AVP1 | AmpR; M13p-TEF1p-(Tc)(Suc2)AVP1-CYC7t | This study |
| pYES-pACT1-pHluorin | AmpR; URA3; ACT1p-pHluorin-CYC1t | [39] |
| p426-GPDp | URA3; AmpR | [45] |
| pTMB_KS_036 | AmpR; URA3; GPD1p-pHluorin-CYC7t; | This Study |
| pTMB_KS_038 | AmpR; URA3; GPD1p-Queen-2m-CYC7t; | This Study |
| pCFB2312 | KanR; pTEF1p-Cas9-CYC1t | [46] |
| pCFB3035 | gRNA_X-4; natMX | [46] |
| pCFB3042 | X-4 MarkerFree backbone; Geneticin | [46] |
| pCFB2904 | gRNA_XI-3; natMX | [46] |
| pCFB3045 | XI-3 MarkerFree backbone; Geneticin | [46] |
| pTMB_KS_040 | pCFB3042; GPD1p-pHluorin-CYC7t | This Study |
| pTMB_KS_041 | pCFB3042; GPD1p-Queen-2m-CYC7t | This Study |
| pTMB_KS_042 | AmpR; M13p-TEF1p-(Tc)AVP1-CYC7t | This Study |
| pTMB_KS_043 | AmpR; M13p-TEF1p-(Suc2)AVP1-CYC7t | This Study |
| pTMB_KS_044 | pCFB3045; TEF1p-(Tc)AVP1-CYC7t | This Study |
| pTMB_KS_045 | pCFB3045; TEF1p-(Suc2)AVP1-CYC7t | This Study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sreenivas, K.; Eisentraut, L.; Brink, D.P.; Persson, V.C.; Carlquist, M.; Gorwa-Grauslund, M.F.; van Niel, E.W.J. Evaluation of Pyrophosphate-Driven Proton Pumps in Saccharomyces cerevisiae under Stress Conditions. Microorganisms 2024, 12, 625. https://doi.org/10.3390/microorganisms12030625
Sreenivas K, Eisentraut L, Brink DP, Persson VC, Carlquist M, Gorwa-Grauslund MF, van Niel EWJ. Evaluation of Pyrophosphate-Driven Proton Pumps in Saccharomyces cerevisiae under Stress Conditions. Microorganisms. 2024; 12(3):625. https://doi.org/10.3390/microorganisms12030625
Chicago/Turabian StyleSreenivas, Krishnan, Leon Eisentraut, Daniel P. Brink, Viktor C. Persson, Magnus Carlquist, Marie F. Gorwa-Grauslund, and Ed W. J. van Niel. 2024. "Evaluation of Pyrophosphate-Driven Proton Pumps in Saccharomyces cerevisiae under Stress Conditions" Microorganisms 12, no. 3: 625. https://doi.org/10.3390/microorganisms12030625