Molecular Prevalence of Blastocystis sp. from Patients with Diarrhea in the Republic of Korea


Division of Vectors and Parasitic Diseases, Korea Disease Control and Prevention Agency, 187 Osongsaenmyeong 2-ro, Osong-eup, Heungdeok-gu, Cheongju 28159, Chungcheongbuk-do, Republic of Korea
*
Author to whom correspondence should be addressed.
Microorganisms 2024, 12(3), 523; https://doi.org/10.3390/microorganisms12030523
Submission received: 6 February 2024
/
Revised: 26 February 2024
/
Accepted: 4 March 2024
/
Published: 5 March 2024
(This article belongs to the Section Parasitology)
Abstract
:Blastocystis sp. is the most common intestinal protozoan affecting human health worldwide. Several studies have reported the prevalence of Blastocystis sp. in various regions of the Republic of Korea. However, limited data are available on the prevalence and subtype (ST) distribution of this parasite among regions. Therefore, we investigated the prevalence and ST distributions of this parasite in the Republic of Korea. For this purpose, 894 stool specimens were collected from patients with diarrhea and tested for the presence of Blastocystis sp. using PCR analysis. The isolates were subsequently subtyped. The overall prevalence was 11.6%. Of the 104 isolates, ST3 was the most prevalent, followed by ST1. Additionally, a single case of the rare subtype ST8 was identified, representing the first reported case in the Republic of Korea. The results suggested that the predominance of ST3 observed in this study reflects human-to-human transmission with low genetic diversity within the ST, while ST1 transmission is likely correlated with animals. In the future, to better understand Blastocystis sp. transmission dynamics, human, animal, and environmental factors should be studied from a “One Health” perspective.
1. Introduction
Blastocystis sp. is a protozoan parasite that resides within the gastrointestinal tract of a wide range of hosts, including mammals, birds, fish, reptiles, insects, and humans [1,2]. This enteric microorganism is a causative agent of waterborne and foodborne diseases. The infective cysts are transmitted through the fecal–oral route [3,4]. An estimated more than 1 billion people are infected with this parasite worldwide, with a prevalence of 10–15% in developed countries and >50% in developing nations [5]. Although most infected people are asymptomatic carriers, some vulnerable individuals develop symptoms, including diarrhea, abdominal pain, gas, and nausea, as well as extraintestinal symptoms such as urticaria and rash [6,7,8,9,10]. Blastocystis sp. colonization is reportedly associated with a healthy gut microbiome rather than intestinal dysbacteriosis [6]. However, in vitro and in vivo studies combined with genomic analyses have identified various virulence factors that may be related to the genetic variation in Blastocystis isolates [2,11].
The genetic variation in Blastocystis sp. in birds and mammals (including humans) is categorized into at least 28 subtypes (STs), including ST1–ST17, ST21, and ST23–ST32, based on polymorphisms in the small subunit of the rRNA (SSU rRNA) gene [12]. Among these, ST1–10, ST12, ST14, ST16, and ST23 have been reported in humans, with ST1–ST4 representing >90% of subtyped human isolates [13,14]. Accordingly, most ST1–ST4 infections are transmitted via human-to-human routes, while the other STs predominate in certain host groups, such as mammals or birds, and are transmitted through human–animal pathways [15,16].
Genetic variant studies using Multilocus Sequence Typing (MLST) schemes have revealed intra-genetic variation among the STs [14,17]. More specifically, following isolation from symptomatic patients, Blastocystis sp. subtypes and 18S alleles have been reported in several countries [9,18,19,20,21]. In Iran, the predominant Blastocystis subtypes responsible for causing diarrhea, bloating, nausea, and urticaria are ST1 (allele 4), ST2 (alleles 9, 11, and 12), ST3 (alleles 34, 37, and 52), and ST6 (allele 122) [19,20]. In Colombia, those causing diarrhea were classified as ST2 (allele 9), and those associated with irritable bowel syndrome (IBS) were classified as ST3 (allele 34) [21].
Although studies in the Republic of Korea (ROK) have reported the STs of Blastocystis sp. [22,23,24,25], 18S alleles have not been reported for Blastocystis STs isolated from humans. Therefore, the primary aim of the current study was to evaluate the genetic variation and geographical distribution of subtypes and 18S alleles in patients with diarrhea in the ROK.
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
In total, 894 stool samples were collected from patients with diarrhea via the Enteric Pathogens Active Surveillance Network (Enter-Net) of the Korea Disease Control and Prevention Agency (KDCA) in 2022. Enter-Net is organized by the KDCA and comprised of 11 local Public Health Institutes and 73 participating hospitals. Samples were collected from patients suspected of waterborne and foodborne infectious diseases to isolate and identify the causative agent. The samples were collected from the following 11 regions in the ROK with 10 mL screw-cap tubes: Incheon, Seoul, Chungcheong buk-do, Gyeongsangbuk-do, Daejeon, Daegu, Jeollabuk-do, Gyeongsangnam-do, Busan, Jeollanam-do, and Jeju Island (Table 1). The samples were stored at −20 °C before DNA extraction. Total DNA was extracted from 250 mg (or 250 µL for watery samples) of fecal samples using the QIAcube HT apparatus and the DNeasy® PowerSoil® Pro Kit (QIAGEN, Hilden, Germany), following the manufacturer’s protocol. To enhance the extraction efficacy, the FastPrep-24™ 5G (MP Biomedicals, Solon, OH, USA) was employed for physical disruption at 6 m/s for 40 s. The extracted DNA was preserved at −20 °C until PCR analysis. The samples were processed to detect viruses, bacteria, and parasitic protozoa.
Ethical approval was not required because this study managed to assess public welfare via a fact-finding survey (Infectious Disease Control and Prevention Act).
2.2. PCR Amplification
PCR amplification of Blastocystis sp. was carried out using barcoding region primers BhRDr (5′-GAGCTTTTTAACTGCAACAACG-3′) and RD5 (5′-ATCTGGTTGATCCTGCCAGT-3′) for SSU rRNA [26]. PCR amplification included the following steps: initial denaturing at 94 °C for 5 min and 35 cycles of 94 °C for 1 min, 59 °C for 1 min, and 1 min at 72 °C; a final extension at 72 °C was performed for 2 min. A negative control was used to detect contamination in each reaction. The amplified products (600 bp specific band) were visualized using a Qiagen QIAxcel Capillary Electrophoresis system (QIAGEN, Hilden, Germany).
2.3. DNA Sequencing Analysis
All positive PCR products were purified and sequenced. Nucleotide sequences were analyzed and trimmed using BioEdit version 7.2.5 (Ibis Therapeutics Inc., Carlsbad, CA, USA). The resulting nucleotide sequences were subjected to BLAST searches (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 20 August 2023) using the data available on GenBank (http://www.ncbinlm.nih.gov/genbank/, accessed on 20 August 2023). STs were determined based on an exact match or an identity ≥99% against all known Blastocystis STs, with a query coverage of ≥99%.
2.4. Phylogenetic Analysis and Allele Discrimination
Multiple sequence alignments were performed using ClustalW BioEdit version 7.2.5 (Ibis Therapeutics Inc., Carlsbad, CA, USA). The Mega Align program (DNASTAR, Madison, WI, USA) was used to determine the similarity and difference rates between sequences. Finally, phylogenetic analysis was performed with MEGA software (version 5.02) using neighbor-joining analysis with a Kimura 2-parameter model estimated using bootstrap analysis with 1000 replicates. Each sequence was identified based on the accession number, host, origin, and genotype. All established sequences were submitted to the MLST database (https://pubmlst.org/blastocystis/, accessed on 20 August 2023) for ST corroboration and relevant allele identification [26].
2.5. Statistical Analysis
The relationships between Blastocystis sp. and sex or age groups were assessed using the Chi-square (X2) test or Fisher’s exact test. A p value < 0.05 was considered significant. All statistical analyses were performed using GraphPad Prism 5 software 5.01.
3. Results
3.1. Regional Distribution of Blastocystis sp.
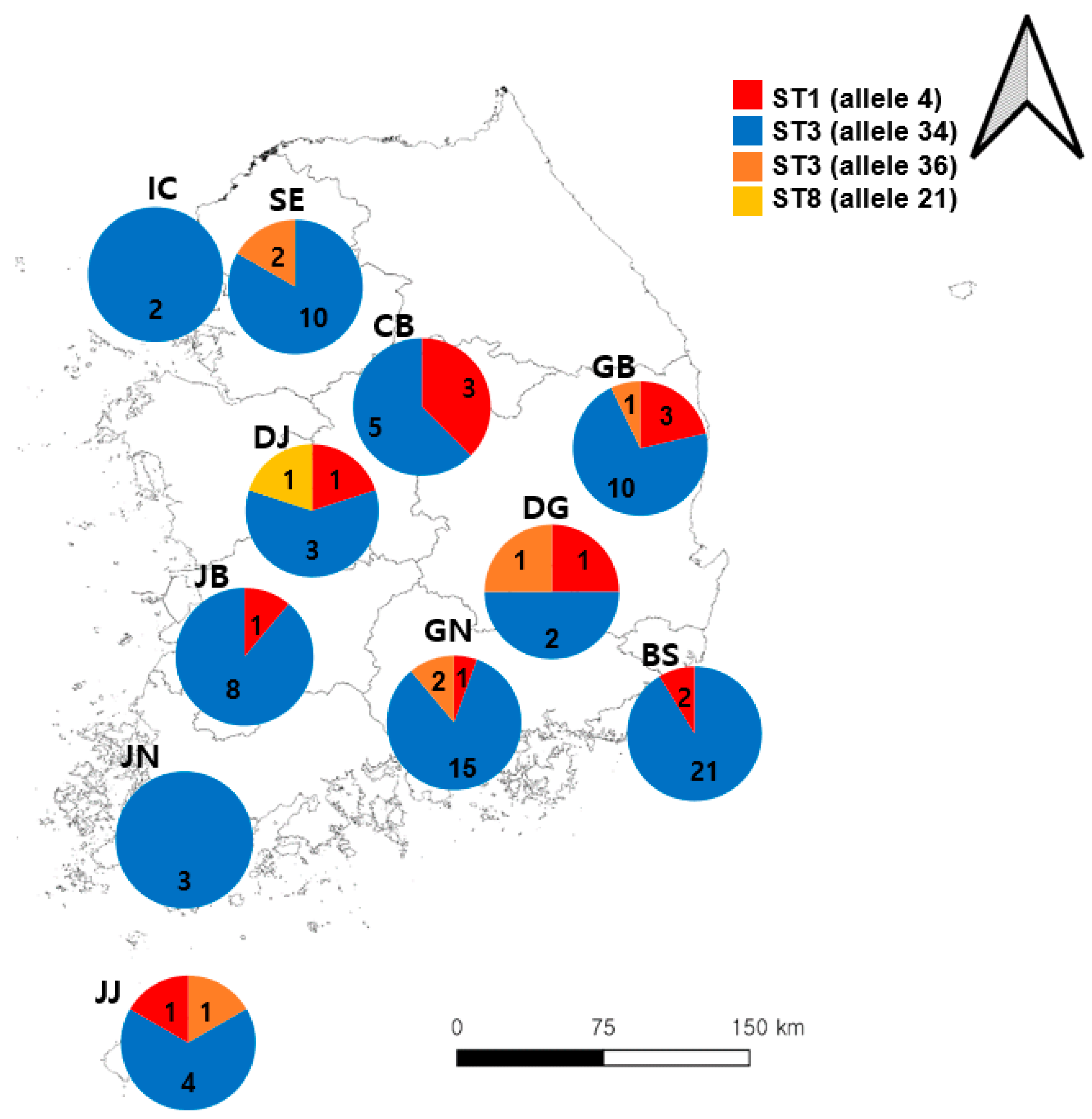
In total, 894 samples were collected from patients with diarrhea in this study, 104 of which were positive for Blastocystis sp. (Table 1). Busan had the highest number of positive cases, followed by Gyeongsangnam-do and Seoul. Three different Blastocystis sp. STs were identified, including ST1 (11/104, 10.6%), ST3 (92/104, 88.5%), and ST8 (1/104, 1.0%). ST3 (allele34) was found to be the most common Blastocystis sp., which was detected in all sample collection areas, but ST3 (allele36) was detected only in five areas: Seoul, Gyeongsangbuk-do, Daegu, Gyeongsangnam-do, and Jeju Island. The prevalence of Blastocystis sp. ST1 was identified in Chungcheongbuk-do, Daejeon, Daegu, Gyeongsangbuk-do, Gyeongsangnam-do, Jeollabuk-do, and Jeju Island (Figure 1). Of the 104 Blastocystis sp. cases, 18 were confirmed to be co-infected with Bacillus cereus, Clostridium perfringens, Escherichia coli, Salmonella spp., and Shigella spp. Mixed ST infections with Blastocystis sp. were not detected.
3.2. Prevalence of Blastocystis sp.
Blastocystis sp. was detected in 45 male (45/461, 9.8%) and 59 female (59/433, 13.6%) patients. However, infection was not associated with sex (p = 0.118, odds ratio = 1.395, 95% confidence interval = 0.9247–2.106; Table 2).
The ages of the patients ranged from 1 month to 97 years (mean: 49 years; SD: ±30 years). The patients were divided into eight categories based on age; Blastocystis sp. was detected in all age groups. The largest number of positive cases was detected within the 0–9 age group, with the associated infection rate significantly higher in the other age groups (Chi-square = 23.09, p = 0.001; Table 2).
3.3. Phylogenetic Analysis
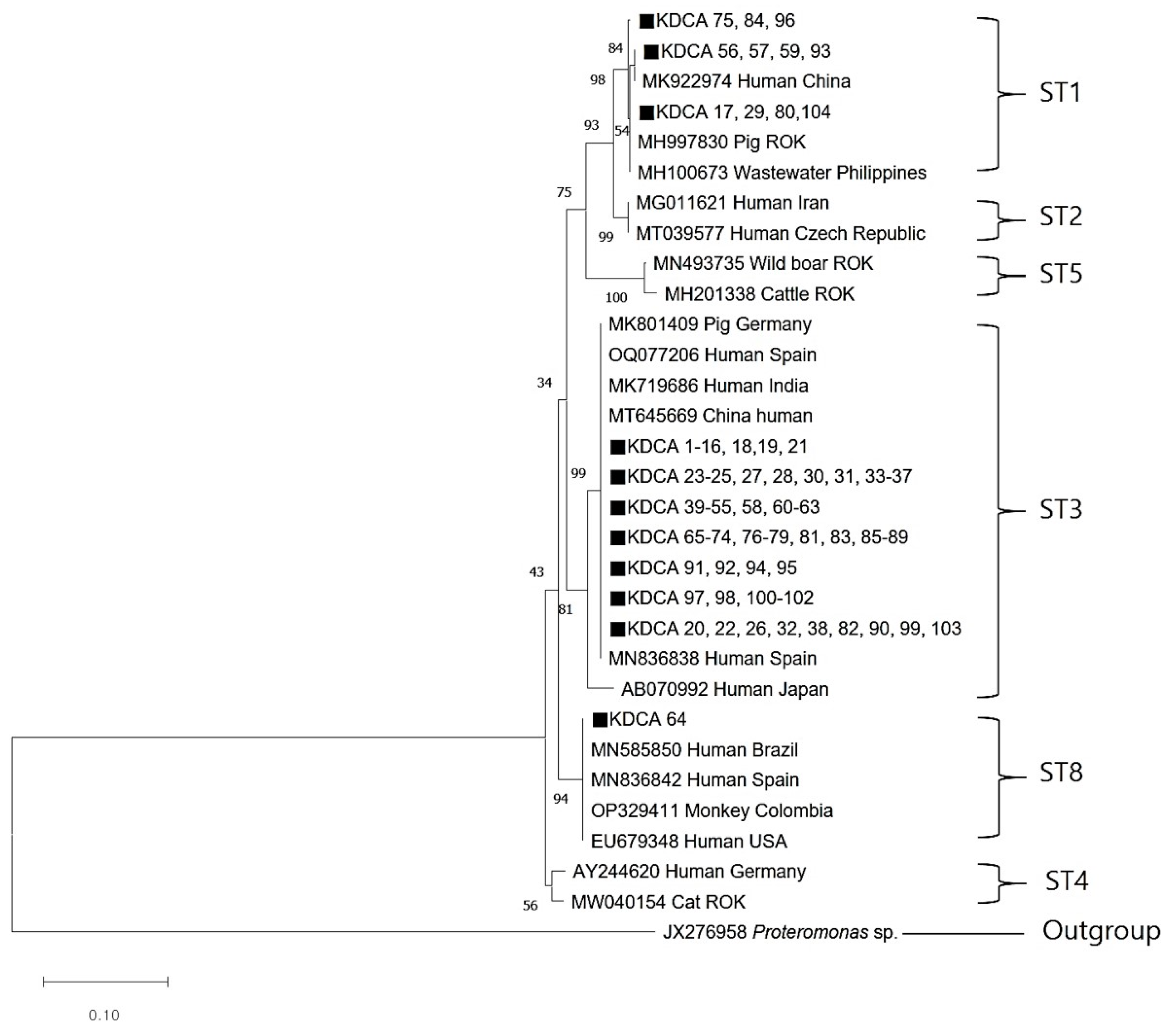
All 104 Blastocystis-positive samples were successfully sequenced. The ST1 sequences (KDCA 56, 57, 59, and 93) were closely related to the ST1 sequence (MK922974) isolated from humans in China. In addition, ST1 sequences (KDCA 17, 29, 80, and 104) were similar to an ST1 sequence (MH997830) obtained from pigs in the ROK. The ST3 sequence showed a close genetic relationship with human ST3 sequences from Spain, India, and China. The ST8 sequence (KDCA 64) obtained in this study was identical to those from humans in Brazil (MN585850), Spain (MN836842), and the USA (EU679348), as well as those from monkeys in Colombia (OP329411) (Figure 2). Excluding the previously registered sequences, all sequences obtained in this study were deposited into GenBank under accession number OR447547.
3.4. Distribution of 18S Alleles and Genotypes
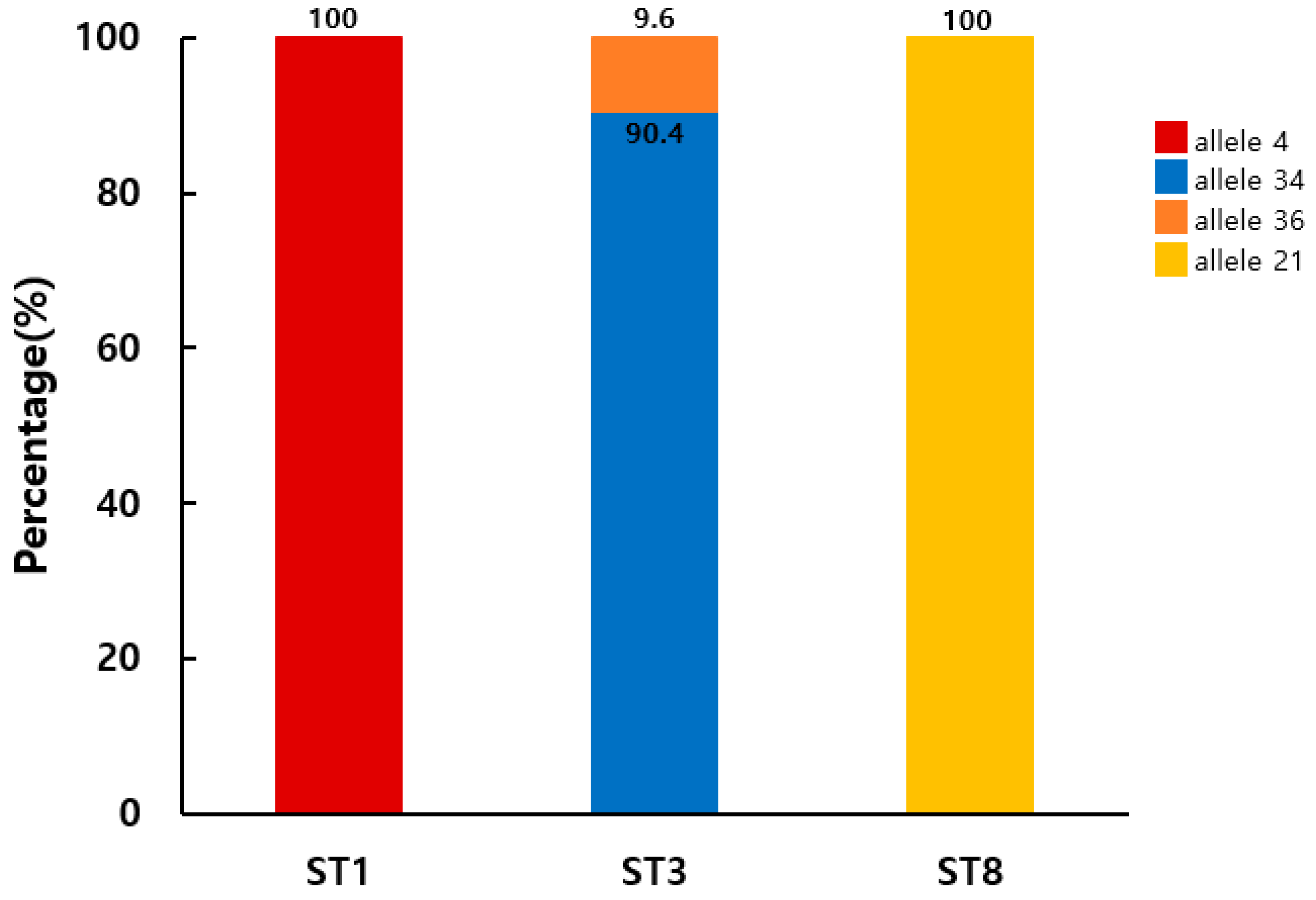
Allele analysis of the Blastocystis SSU rRNA gene revealed four variants in the positive samples. Based on the Blastocystis 18S database, the detected alleles were identified as allele 4 (100%; 11/11) for ST1, alleles 34 (90.4%; 83/92) and 36 (9.6%; 9/92) for ST3, and allele 21 (100%; 1/1) for ST8 (Figure 3). To evaluate the intra-ST diversity and identify genotypes, partial SSU rDNA gene sequences belonging to ST1 or ST3 were aligned (Figure 4). This comparison identified three ST1 (ST1-1 to ST1-3) and two ST3 (ST3-1 and ST3-2) genotypes in patients with diarrhea in the ROK. All ST1 isolates (ST1-1 to ST1-3) belonged to allele 4, with one or four variable positions. Meanwhile, ST3-1 belonged to allele 34, and ST3-2 to allele 36, with a single variable position.
4. Discussion
Blastocystis is one of the most common genera of intestinal eukaryotes found in humans worldwide [16]. The prevalence of Blastocystis sp. reportedly ranges from 0.5% in Thailand to 100% in Senegal [12,27]. Several Asian studies have determined the average prevalence of Blastocystis sp. in China (11.62%), Japan (1.0%), Malaysia (19.25%), Nepal (25.2%), Singapore (3.3%), the Philippines (49.1%), and Indonesia (34.25%) [16,28]. The prevalence of Blastocystis sp. is particularly high within marginalized areas in terms of public health, industrialization, and socioeconomic status [29]. In China, the prevalence of Blastocystis sp. reaches 32.6% in less industrialized and socio-economically marginalized areas, whereas in industrialized urban areas, it is as low as 1.9% [30]. Similarly, in Malaysia, low household income and untreated tap water use were identified as important risk factors for Blastocystis sp. infection [31]. In Nepal, infection is widespread in rural communities due to the consumption of untreated water [32]. Meanwhile, in the Philippines, the prevalence of Blastocystis sp. infection was high among pet owners [33]. The high prevalence was found to have poor hygiene and close contact between humans and animals in the area surveyed [34]. In contrast, Japan reported a decrease from the previously reported prevalence (0.5–2.5%) due to improved public health [28]. Comparatively, Singapore is an urbanized city-state with a low prevalence rate [35]. In previous studies, the prevalence of Blastocystis sp. in the ROK was 5.6–9.2%, while the current study detected a rate of 11.6% [22,23,24,25]. This discrepancy may stem from the differences in sample size and breadth of sample collection locations. That is, the current study included more than 500 stool samples collected across various regions and age groups. In terms of distribution by age, the prevalence of Blastocystis sp. was high in those aged 0–9 and >60 years, presumably due to relatively weakened immunity compared with the other groups [36].
One of the most common symptoms associated with Blastocystis sp. is diarrhea, with several studies reporting a correlation between diarrhea and specific subtypes. For example, in Iran, ST1, ST2, ST3, and ST6 were detected in symptomatic patients, while ST2 and ST3 were detected in Colombian patients with diarrhea [19,20,21]. Similarly, a Saudi Arabian study reported that ST3 predominated in patients with abdominal pain and diarrhea [37]. However, ST1, ST2, and ST3 have also been reported in asymptomatic carriers. Hence, further investigation into the pathological role of Blastocystis subtypes is needed.
As previously reported, ST3 of Blastocystis sp. is the predominant subtype globally [14,38]. This aligned with the findings of the current study, with ST3 as the dominant subtype in the ROK, followed by ST1. Our phylogenetic analysis revealed that the ST3 sequence shared sequence similarity with other human isolates from around the world, whereas no similar sequences have been reported in domestic animals. Hence, ST3 is transmitted via human-to-human routes. Moreover, ST3 was detected in all study areas, suggesting its broad distribution among humans in the ROK. ST1 is the predominant subtype isolated from humans [14,39]. In the current study, most ST1 sequences were phylogenetically similar to others reportedly isolated from humans. Meanwhile, certain ST1 sequences were closely related to ST1 sequences isolated in domestic pigs (MH997827). This suggests indirect zoonotic transmission caused by exposure to contaminated water, food, or aerosols [38,40]. In addition, although ST2 has been previously reported in the ROK [22], it was not detected in the current study. Meanwhile, we identified Blastocystis ST8 in the ROK for the first time. ST8 is typically isolated from non-human primates and rarely humans [15]. Although it has been reported in animals and contaminated water sources in Asia, it has not been previously reported in East Asian populations [41]. In fact, Blastocystis ST8 has only been previously isolated in humans within the Americas and Europe [38]. The nucleotide sequence of ST8 detected in this study was 100% consistent with that of the American and European strains. Although the patient’s overseas travel history could not be ascertained, overseas inflow was suspected.
DNA barcoding is a useful method for determining intra-ST diversity [21]. According to allelic discrimination related to the identified subtypes in humans, globally, the most frequent alleles in each ST are as follows: ST1 (alleles 4 and 2), ST2 (allele 9), ST3 (alleles 34, 36, and 37), ST4 (alleles 42 and 91), and ST8 (allele 21) [2,42]. Genetic analysis in this study revealed that all four alleles detected in the Blastocystis sp. cross-corresponded with previously reported alleles. Analysis of the variable positions within the alignment of each ST revealed three ST1 genotypes in 11 isolates. In addition, two ST3 genotypes were identified. Although ST3 was detected most frequently, its genetic diversity was low. Considering that ST3 has never been reported in livestock or farm animals, the low genetic diversity could be associated with it being transmitted exclusively via human-to-human routes in the ROK. Among the ST genotypes, ST3-1 (79.8% of all ST isolates) predominated, representing the main cause of Blastocystis sp. transmission in the ROK.
Certain limitations were noted in the current study. No data were available on symptoms other than diarrhea, nor was information related to environmental or animal contact provided. In the future, multicenter epidemiological studies are needed to evaluate the role of environmental factors, animal contact, and overseas travel history to confirm the active circulation of Blastocystis sp. in the ROK. In addition, studies from the “One Health” perspective would help develop a more comprehensive understanding of Blastocystis sp. transmission and prevalence.
Author Contributions
Conceptualization, J.-W.J.; methodology, J.-Y.K. and J.-H.C.; investigation and data analysis, J.-Y.K. and H.-I.L.; performing the experiments and resource collection, J.-Y.K. and J.-H.C.; writing—original draft preparation, J.-Y.K.; writing—review and editing, M.-R.L., J.-W.J. and H.-I.L.; supervision, M.-R.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by a grant from the Korea Disease Control and Prevention Agency (KDCA; 6331-311-210-13) of the Republic of Korea.
Data Availability Statement
Patient consent was not required for this study as it involved a retrospective analysis of specimens collected for legally stipulated research survey projects, and no further investigations were conducted with the submitted specimens.
Acknowledgments
We appreciate the 11 regional Institute of Health & Environment members for collecting samples nationwide, including Incheon, Seoul, Chungcheongbuk-do, Gyeongsangbuk-do, Daejeon, Daegu, Jeollabuk-do, Gyeongsangnam-do, Busan, Jeollanam-do and Jeju Island Institute of Health & Environment. We also appreciate the valuable assistance provided by Bo Gyeong Han (Korea Disease Control and Prevention Agency) in creating the map illustrated in Figure 1.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Garcia-Montoya, G.M.; Galvan-Diaz, A.L.; Alzate, J.F. Metataxomics Reveals Blastocystis Subtypes Mixed Infections in Colombian Children. Infect. Genet. Evol. 2023, 113, 105478. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.D.N.; Gantois, N.; Hoang, T.T.; Do, B.T.; Desramaut, J.; Naguib, D.; Tran, T.N.; Truong, A.D.; Even, G.; Certad, G.; et al. First Epidemiological Survey on the Prevalence and Subtypes Distribution of the Enteric Parasite Blastocystis sp. in Vietnam. Microorganisms 2023, 11, 731. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, H.; Abe, N.; Wu, Z. PCR-Based Identification of Zoonotic Isolates of Blastocystis from Mammals and Birds. Microbiology 2004, 150, 1147–1151. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.I.; Chye, T.T.; Karmacharya, B.M.; Govind, S.K. Blastocystis sp.: Waterborne Zoonotic Organism, a Possibility? Parasit. Vectors 2012, 5, 130. [Google Scholar] [CrossRef]
- Matovelle, C.; Tejedor, M.T.; Monteagudo, L.V.; Beltrán, A.; Quílez, J. Prevalence and Associated Factors of Blastocystis sp. Infection in Patients with Gastrointestinal Symptoms in Spain: A Case-Control Study. Trop. Med. Infect. Dis. 2022, 7, 226. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, P.D.; Stensvold, C.R.; Rajilić-Stojanović, M.; Heilig, H.G.; De Vos, W.M.; O’Toole, P.W.; Cotter, P.D. The Microbial Eukaryote Blastocystis Is a Prevalent and Diverse Member of the Healthy Human Gut Microbiota. FEMS Microbiol. Ecol. 2014, 90, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Angelici, M.C.; Nardis, C.; Scarpelli, R.; Ade, P. Blastocystis hominis Transmission by Non-potable Water: A Case Report in Italy. New Microbiol. 2018, 41, 173–177. [Google Scholar]
- Fréalle, E.; El Safadi, D.; Cian, A.; Aubry, E.; Certad, G.; Osman, M.; Wacrenier, A.; Dutoit, E.; Creusy, C.; Dubos, F.; et al. Acute Blastocystis-associated appendicular peritonitis in a child, Casablanca, Morocco. Emerg Infect Dis. 2015, 21, 91. [Google Scholar] [CrossRef]
- Aykur, M.; Calıskan Kurt, C.; Dirim Erdogan, D.; Biray Avcı, C.; Vardar, R.; Aydemir, S.; Girginkardesler, N.; Gunduz, C.; Dagci, H. Distribution and Phylogenetic Analysis of Subtypes and Alleles of Blastocystis sp. in the Stool Samples Collected from Patients with Gastrointestinal Complaints in İzmir, Turkey. Acta Parasitol. 2023, 68, 304–316. [Google Scholar] [CrossRef]
- Casero, R.D.; Mongi, F.; Sánchez, A.; Ramírez, J.D. Blastocystis and Urticaria: Examination of Subtypes and Morphotypes in an Unusual Clinical Manifestation. Acta Trop. 2015, 148, 156–161. [Google Scholar] [CrossRef]
- Ajjampur, S.S.; Tan, K.S.W. Pathogenic Mechanisms in Blastocystis spp.—Interpreting Results from in Vitro and in Vivo Studies. Parasitol. Int. 2016, 65, 772–779. [Google Scholar]
- Buthaina, D.; Ghalia, A.; Samar, A.N. Molecular Characterization of Blastocystis Subtypes in Symptomatic Patients from the Southern Region of Syria. PLoS ONE 2023, 18, e02832. [Google Scholar]
- Stensvold, C.R.; Christiansen, D.B.; Olsen, K.E.P.; Nielsen, H.V. Blastocystis sp. Subtype 4 Is Common in Danish Blastocystis-Positive Patients Presenting with Acute Diarrhea. Am. J. Trop. Med. Hyg. 2011, 84, 883–885. [Google Scholar] [CrossRef]
- Alfellani, M.A.; Stensvold, C.R.; Vidal-Lapiedra, A.; Onuoha, E.S.; Fagbenro-Beyioku, A.F.; Clark, C.G. Variable Geographic Distribution of Blastocystis Subtypes and Its Potential Implications. Acta Trop. 2013, 126, 11–18. [Google Scholar] [CrossRef]
- Gong, B.; Liu, X.; Wu, Y.; Xu, N.; Xu, M.; Yang, F.; Tong, L.; Zhou, K.; Cao, J.; Liu, A.; et al. Prevalence and Subtype Distribution of Blastocystis in Ethnic Minority Groups on Both Sides of the China-Myanmar Border, and Assessment of Risk Factors. Parasite 2019, 26, 46. [Google Scholar] [CrossRef] [PubMed]
- Nemati, S.; Zali, M.R.; Johnson, P.; Mirjalali, H.; Karanis, P. Molecular Prevalence and Subtype Distribution of Blastocystis sp. in Asia and in Australia. J. Water Health 2021, 19, 687–704. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Alfellani, M.; Clark, C.G. Levels of Genetic Diversity Vary Dramatically Between Blastocystis Subtypes. Infect. Genet. Evol. 2012, 12, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, J.D.; Flórez, C.; Olivera, M.; Bernal, M.C.; Giraldo, J.C. Blastocystis Subtyping and Its Association with Intestinal Parasites in Children from Different Geographical Regions of Colombia. PLoS ONE 2017, 12, e0172586. [Google Scholar] [CrossRef] [PubMed]
- Alinaghizade, A.; Mirjalali, H.; Mohebali, M.; Stensvold, C.R.; Rezaeian, M. Inter- and Intra-Subtype Variation of Blastocystis Subtypes Isolated from Diarrheic and Non-Diarrheic Patients in Iran. Infect. Genet. Evol. 2017, 50, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Rezaei Riabi, T.; Mirjalali, H.; Haghighi, A.; Rostami Nejad, M.; Pourhoseingholi, M.A.; Poirier, P.; Delbac, F.; Wawrzyniak, I.; Zali, M.R. Genetic Diversity Analysis of Blastocystis Subtypes from Both Symptomatic and Asymptomatic Subjects Using a Barcoding Region from the 18S rRNA Gene. Infect. Genet. Evol. 2018, 61, 119–126. [Google Scholar] [CrossRef]
- Ramírez, J.D.; Sánchez, L.V.; Bautista, D.C.; Corredor, A.F.; Flórez, A.C.; Stensvold, C.R. Blastocystis Subtypes Detected in Humans and Animals from Colombia. Infect. Genet. Evol. 2014, 22, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Won, E.J.; Kim, S.H.; Shin, J.H.; Chai, J.Y. Molecular Detection and Subtyping of Human Blastocystis and the Clinical Implications: Comparisons Between Diarrheal and Non-diarrheal Groups in Korean Populations. Korean J. Parasitol. 2020, 58, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.H.; Jung, B.K.; Shin, H.J.; Hong, S.J.; Ryoo, S.W.; Lee, J.G.; Han, S.H.; Park, S.M.; Rhee, M.S.; Kimm, H.J.; et al. Genotypes of Blastocystis sp. Among Elderly Health Checkup People in South Korea with a Questionnaire on Risk Factors. Parasitol. Res. 2021, 120, 3297–3306. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Byun, J.H.; Oh, Y.J.; Liu, C.S.; Bae, M.H.; Won, E.J. Epidemiology and Subtype Distribution of Blastocystis Among Asymptomatic Volunteers in South Korea: A Multicenter Study. Ann. Clin. Microbiol. 2023, 26, 11–17. [Google Scholar]
- Won, E.J.; Kim, S.H.; Kee, S.J.; Shin, J.H.; Suh, S.P.; Chai, J.Y.; Ryang, D.W.; Shin, M.G. Multiplex Real-Time PCR Assay Targeting Eight Parasites Customized to the Korean Population: Potential Use for Detection in Diarrheal Stool Samples from Gastroenteritis Patients. PLoS ONE 2016, 11, e0166957. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Clark, C.G. Molecular Identification and Subtype Analysis of Blastocystis. Curr. Protoc. Microbiol. 2016, 43, 20A.2.1–20A.2.10. [Google Scholar] [CrossRef]
- Sagnuankiat, S.; Wanichsuwan, M.; Bhunnachet, E.; Jungarat, N.; Panraksa, K.; Komalamisra, C.; Maipanich, W.; Yoonuan, T.; Pubampen, S.; Adisakwattana, P.; et al. Health Status of Immigrant Children and Environmental Survey of Child Daycare Centers in Samut Sakhon Province, Thailand. J. Immigr. Minor. Health 2016, 18, 21–27. [Google Scholar] [CrossRef]
- Hirata, T.; Nakamura, H.; Kinjo, N.; Hokama, A.; Kinjo, F.; Yamane, N.; Fujita, J. Prevalence of Blastocystis hominis and Strongyloides stercoralis Infection in Okinawa, Japan. Parasitol. Res. 2007, 101, 1717–1719. [Google Scholar] [CrossRef]
- Yoshikawa, H.; Wu, Z.; Kimata, I.; Iseki, M.; Ali, I.K.M.; Hossain, M.B.; Zaman, V.; Haque, R.; Takahashi, Y. Polymerase chain reaction-based genotype classification among human Blastocystis hominis populations isolated from different countries. Parasitol. Res. 2004, 92, 22–29. [Google Scholar]
- Li, L.H.; Zhang, X.P.; Lv, S.; Zhang, L.; Yoshikawa, H.; Wu, Z.; Steinmann, P.; Utzinger, J.; Tong, X.M.; Chen, S.H.; et al. Cross-sectional surveys and subtype classification of human Blastocystis isolates from four epidemiological settings in China. Parasitol. Res. 2007, 102, 83–90. [Google Scholar] [CrossRef]
- Noradilah, S.A.; Moktar, N.; Anuar, T.S.; Lee, I.L.; Salleh, F.M.; Manap, S.N.A.A.; Mohtar, N.S.H.M.; Azrul, S.M.; Abdullah, W.O.; Nordin, A.; et al. Molecular epidemiology of blastocystosis in Malaysia: Does seasonal variation play an important role in determining the distribution and risk factors of Blastocystis subtype infections in the Aboriginal community? Parasit. Vectors 2017, 10, 360. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.L.; Tan, T.C.; Tan, P.C.; Nanthiney, D.R.; Biraj, M.K.; Surendra, K.M.; Suresh, K.G. Predominance of Blastocystis sp. subtype 4 in rural communities, Nepal. Parasitol. Res. 2012, 110, 1553–1562. [Google Scholar] [CrossRef] [PubMed]
- Belleza, M.L.B.; Reyes, J.C.B.; Tongol-Rivera, P.N.; Rivera, W.L. Subtype analysis of Blastocystis sp. isolates from human and canine hosts in an urban community in the Philippines. Parasitol. Int. 2016, 65, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, H.; Tokoro, M.; Nagamoto, T.; Arayama, S.; Asih, P.B.; Rozi, I.E.; Syafruddin, D. Molecular survey of Blastocystis sp. from humans and associated animals in an Indonesian community with poor hygiene. Parasitol. Int. 2016, 65, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Ng, G.C.; Lin, R.T.; Yoshikawa, H.; Taylor, M.B.; Tan, K.S. Predominance of subtype 3 among Blastocystis isolates from a major hospital in Singapore. Parasitol. Res. 2008, 102, 663–670. [Google Scholar] [CrossRef]
- Paik, S.; Jung, B.Y.; Lee, H.; Hwang, M.H.; Han, J.E.; Rhee, M.H.; Kim, T.H.; Kwon, O.D.; Kwak, D. Molecular Detection and Subtyping of Blastocystis in Korean Pigs. Korean J. Parasitol. 2019, 57, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, R.T.; El-Bali, M.A.; Mohamed, A.A.; Abdel-Fatah, M.A.; El-Malky, M.A.; Mowafy, N.M.; Zaghlool, D.A.; Bakri, R.A.; Al-Harthi, S.A. Subtyping of Blastocystis sp. isolated from symptomatic and asymptomatic individuals in Makkah, Saudi Arabia. Parasit. Vectors 2017, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- Popruk, S.; Adao, D.E.V.; Rivera, W.L. Epidemiology and Subtype Distribution of Blastocystis in Humans: A Review. Infect. Genet. Evol. 2021, 95, 105085. [Google Scholar] [CrossRef]
- Rudzińska, M.; Sikorska, K. Epidemiology of Blastocystis Infection: A Review of Data from Poland in Relation to Other Reports. Pathogens 2023, 12, 1050. [Google Scholar] [CrossRef]
- Taamasri, P.; Mungthin, M.; Rangsin, R.; Tongupprakarn, B.; Areekul, W.; Leelayoova, S. Transmission of Intestinal Blastocystosis Related to the Quality of Drinking Water. Southeast Asian J. Trop. Med. Public Health 2000, 31, 112–117. [Google Scholar]
- Rauff-Adedotun, A.A.; Meor Termizi, F.H.; Shaari, N.; Lee, I.L. The Coexistence of Blastocystis spp. in Humans, Animals and Environmental Sources from 2010–2021 in Asia. Biology 2021, 10, 990. [Google Scholar] [CrossRef] [PubMed]
- Kumarasamy, V.; Rajamanikam, A.; Anbazhagan, D.; Atroosh, W.M.; Azzani, M.; Subramaniyan, V.; Abdullah, S.R. Systematic Review and Meta-analysis: Epidemiology of Human Blastocystis spp. Infection in Malaysia. Trop. Med. Infect. Dis. 2023, 8, 415. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Geographical frequency of Blastocystis sp. subtypes (STs) detected in 104 human fecal samples from the Republic of Korea (ROK). The pie charts represent the proportion of subtypes (alleles) detected in each area; the Arabic numerals represent the number of cases detected. Abbreviations: BS, Busan; CB, Chungcheongbuk-do; DG, Daegu; DJ, Daejeon; GB, Gyeongsangbuk-do; GN, Gyeongsangnam-do; IC, Incheon; JB, Jeollabuk-do; JJ, Jeju Island; JN, Jeollanam-do; SE, Seoul.
Figure 1.
Geographical frequency of Blastocystis sp. subtypes (STs) detected in 104 human fecal samples from the Republic of Korea (ROK). The pie charts represent the proportion of subtypes (alleles) detected in each area; the Arabic numerals represent the number of cases detected. Abbreviations: BS, Busan; CB, Chungcheongbuk-do; DG, Daegu; DJ, Daejeon; GB, Gyeongsangbuk-do; GN, Gyeongsangnam-do; IC, Incheon; JB, Jeollabuk-do; JJ, Jeju Island; JN, Jeollanam-do; SE, Seoul.

Figure 2.
Phylogenetic relationships between Blastocystis sp. The relationships between the genotypes of Blastocystis sp. identified in this study and known genotypes published in GenBank were inferred using a neighbor-joining analysis of SSU rRNA gene sequences based on genetic distances calculated by the Kimura 2-parameter model. The numbers on the branches represent percent bootstrapping values from 1000 replicates. The closed black squares represent each genotype identified in this study. Scale bar = 0.10 replacements per nucleotide site.
Figure 2.
Phylogenetic relationships between Blastocystis sp. The relationships between the genotypes of Blastocystis sp. identified in this study and known genotypes published in GenBank were inferred using a neighbor-joining analysis of SSU rRNA gene sequences based on genetic distances calculated by the Kimura 2-parameter model. The numbers on the branches represent percent bootstrapping values from 1000 replicates. The closed black squares represent each genotype identified in this study. Scale bar = 0.10 replacements per nucleotide site.

Figure 3.
Distribution and frequency of Blastocystis sp. SSU rRNA alleles among subtypes isolated from 104 stool samples of patients in the Republic of Korea.
Figure 3.
Distribution and frequency of Blastocystis sp. SSU rRNA alleles among subtypes isolated from 104 stool samples of patients in the Republic of Korea.

Figure 4.
Alignment of partial SSU rRNA gene sequences from Blastocystis sp. ST1 (A) and ST3 (B) isolates. The variable positions for the reference sequences (genotype ST1-1, ST3-1) are indicated above the alignment (vertical number). Within each ST, the identified genotypes are indicated on the left side of the alignment, and the corresponding reference nucleotide sequences are indicated by dashes. On the right side of each alignment are the total number and percentage of isolates identified in this study.
Figure 4.
Alignment of partial SSU rRNA gene sequences from Blastocystis sp. ST1 (A) and ST3 (B) isolates. The variable positions for the reference sequences (genotype ST1-1, ST3-1) are indicated above the alignment (vertical number). Within each ST, the identified genotypes are indicated on the left side of the alignment, and the corresponding reference nucleotide sequences are indicated by dashes. On the right side of each alignment are the total number and percentage of isolates identified in this study.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Blastocystis sp. detection in fecal samples from patients with diarrhea in 11 regions of the Republic of Korea.
Table 1.
Blastocystis sp. detection in fecal samples from patients with diarrhea in 11 regions of the Republic of Korea.
| Region (Abbreviation) | Samples (n) | Blastocystis sp.-Positive | ||||
|---|---|---|---|---|---|---|
| Positive Samples (n) | Prevalence (%) | ST1 (n) | ST3 (n) | ST8 (n) | ||
| Incheon (IC) | 35 | 2 | 5.7 | - | 2 | - |
| Seoul (SE) | 66 | 12 | 18.2 | - | 12 | - |
| Chungcheongbuk-do (CB) | 94 | 8 | 8.5 | 3 | 5 | - |
| Gyeongsangbuk-do (GB) | 98 | 14 | 14.3 | 3 | 11 | - |
| Daejeon (DJ) | 47 | 5 | 10.6 | 1 | 3 | 1 |
| Daegu (DG) | 118 | 4 | 3.4 | 1 | 3 | - |
| Jeollabuk-do (JB) | 100 | 9 | 9.0 | 1 | 8 | - |
| Gyeongsangnam-do (GN) | 99 | 18 | 18.2 | 1 | 17 | - |
| Busan (BS) | 106 | 23 | 21.7 | - | 23 | - |
| Jeollanam-do (JN) | 37 | 3 | 8.1 | - | 3 | - |
| Jeju Island (JJ) | 94 | 6 | 6.4 | 1 | 5 | - |
| Total | 894 | 104 | 11.6 | 11 | 92 | 1 |
ST, subtype.
Table 2.
Distribution of Blastocystis sp. infection among patients with diarrhea in the Republic of Korea.
Table 2.
Distribution of Blastocystis sp. infection among patients with diarrhea in the Republic of Korea.
| Variable | Samples (n) | Blastocystis sp.-Positive | |||||
|---|---|---|---|---|---|---|---|
| Positive Samples (n) | Prevalence (%) | ST1 (n) | ST3 (n) | ST8 (n) | p Value | ||
| Number of samples | 894 | 104 | 11.6 | 11 | 92 | 1 | |
| Sex | |||||||
| Male | 461 | 46 | 10.0 | 5 | 41 | - | 0.118 |
| Female | 433 | 58 | 13.4 | 6 | 51 | 1 | |
| Age group (years) | |||||||
| 0–9 | 188 | 39 | 20.7 | 1 | 38 | - | 0.001 |
| 10–19 | 54 | 3 | 5.6 | - | 3 | - | |
| 20–29 | 32 | 4 | 12.5 | - | 4 | - | |
| 30–39 | 30 | 3 | 10.0 | - | 3 | - | |
| 40–49 | 48 | 4 | 8.3 | - | 4 | - | |
| 50–59 | 97 | 7 | 7.2 | 1 | 5 | 1 | |
| 60–69 | 149 | 10 | 6.7 | 2 | 8 | - | |
| ≥70 | 296 | 34 | 11.5 | 7 | 27 | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kwon, J.-Y.; Choi, J.-H.; Lee, H.-I.; Ju, J.-W.; Lee, M.-R. Molecular Prevalence of Blastocystis sp. from Patients with Diarrhea in the Republic of Korea. Microorganisms 2024, 12, 523. https://doi.org/10.3390/microorganisms12030523
AMA Style
Kwon J-Y, Choi J-H, Lee H-I, Ju J-W, Lee M-R. Molecular Prevalence of Blastocystis sp. from Patients with Diarrhea in the Republic of Korea. Microorganisms. 2024; 12(3):523. https://doi.org/10.3390/microorganisms12030523
Chicago/Turabian StyleKwon, Ji-Young, Jong-Hoon Choi, Hee-Il Lee, Jung-Won Ju, and Myoung-Ro Lee. 2024. "Molecular Prevalence of Blastocystis sp. from Patients with Diarrhea in the Republic of Korea" Microorganisms 12, no. 3: 523. https://doi.org/10.3390/microorganisms12030523
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.