
The Microbial Community Structure in the Rhizosphere of Theobroma cacao L. and Euterpe oleracea Mart. Is Influenced by Agriculture System in the Brazilian Amazon
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
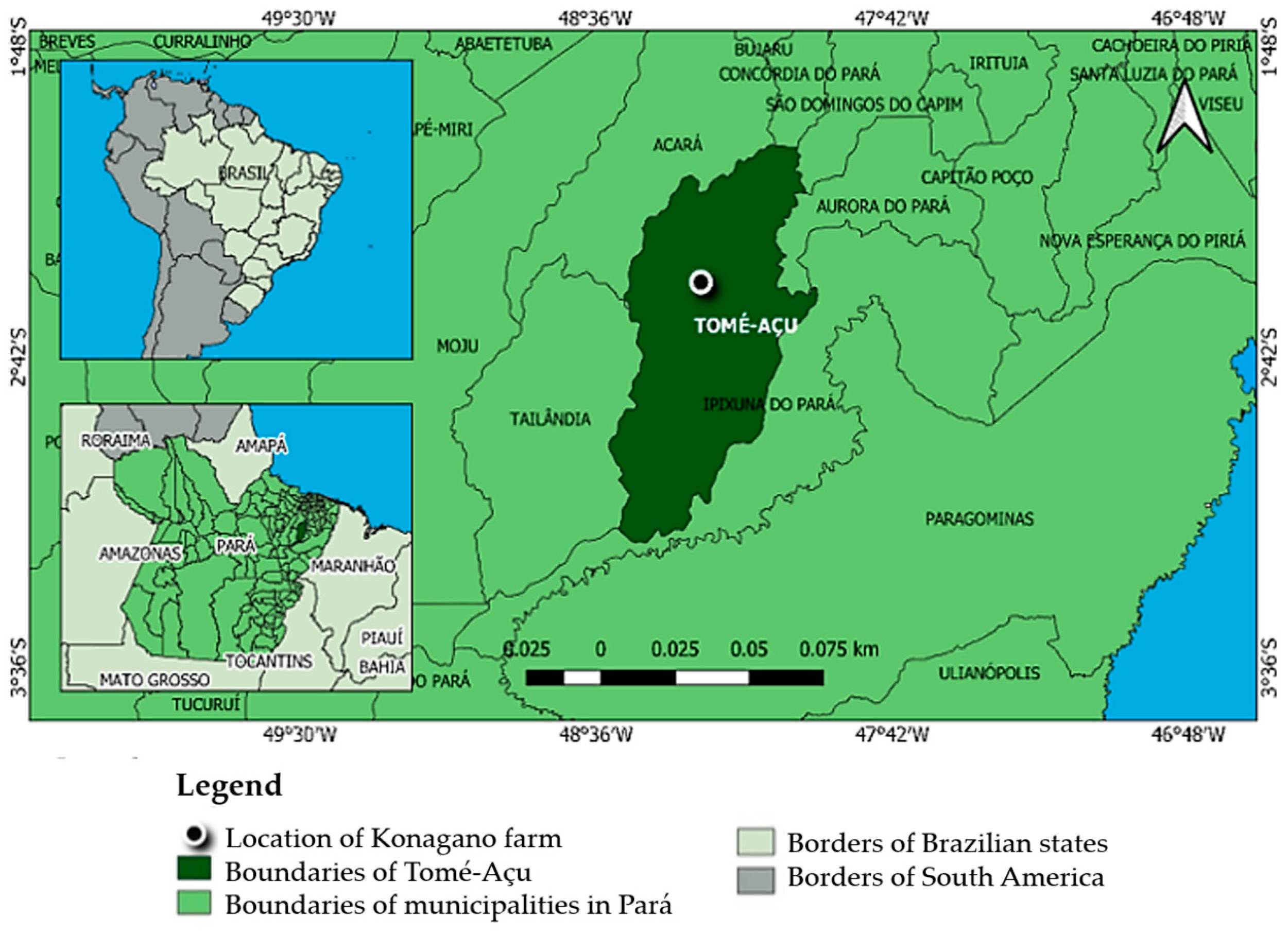
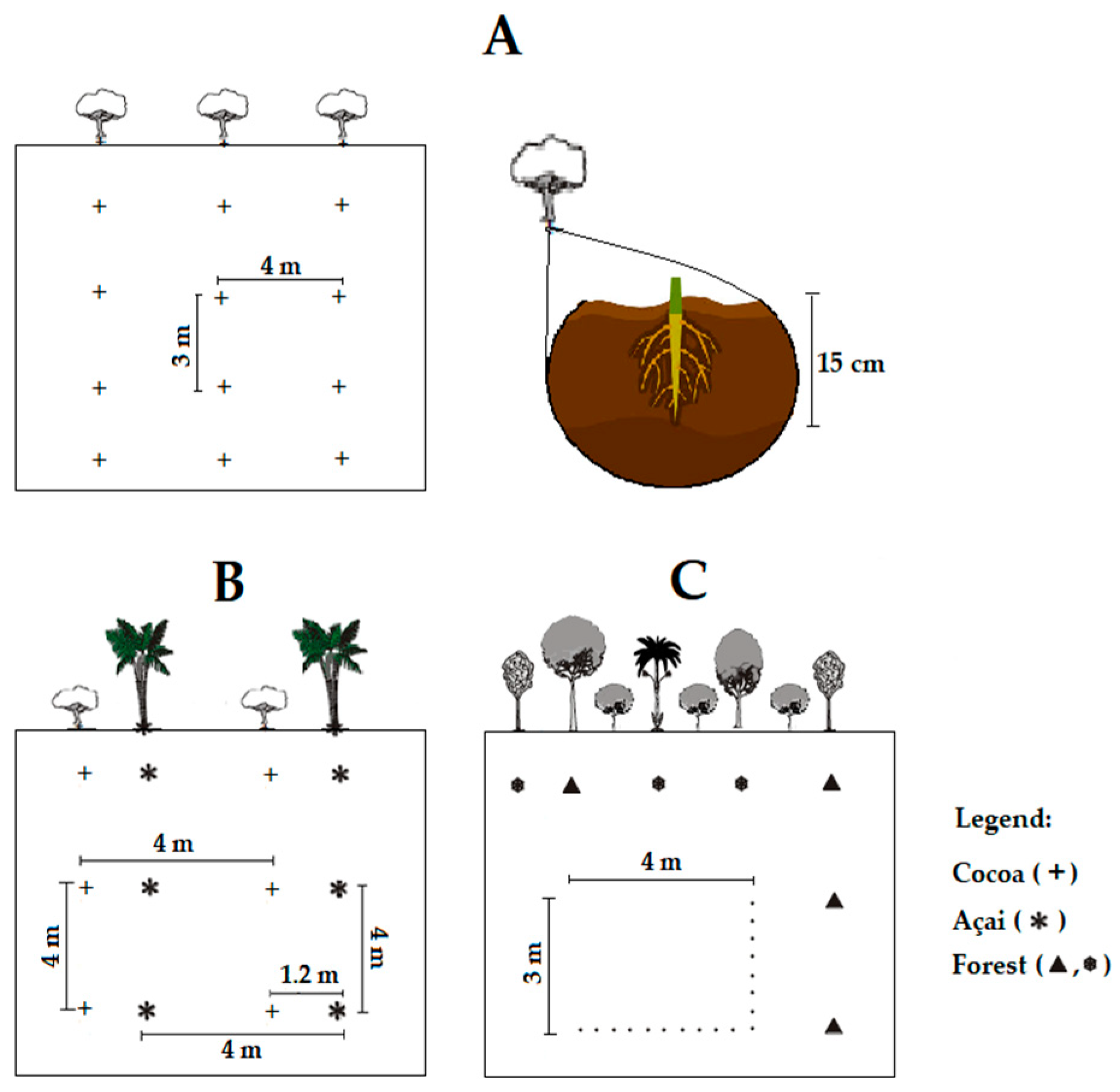
2.1. Field Experiment and Soil Sampling
2.2. TC-DNA Extraction and PCR Amplification
2.3. DGGE and Statistical Analyses
2.4. Isolation of Bacterial Strains
2.5. Molecular Identification of Bacterial Strains
2.6. Production of Siderophores, Indole Acetic Acid (IAA), and Antimicrobial Substances (AMS); Mineralization of Organic Phosphate; and Solubilization of Inorganic Phosphate
3. Results
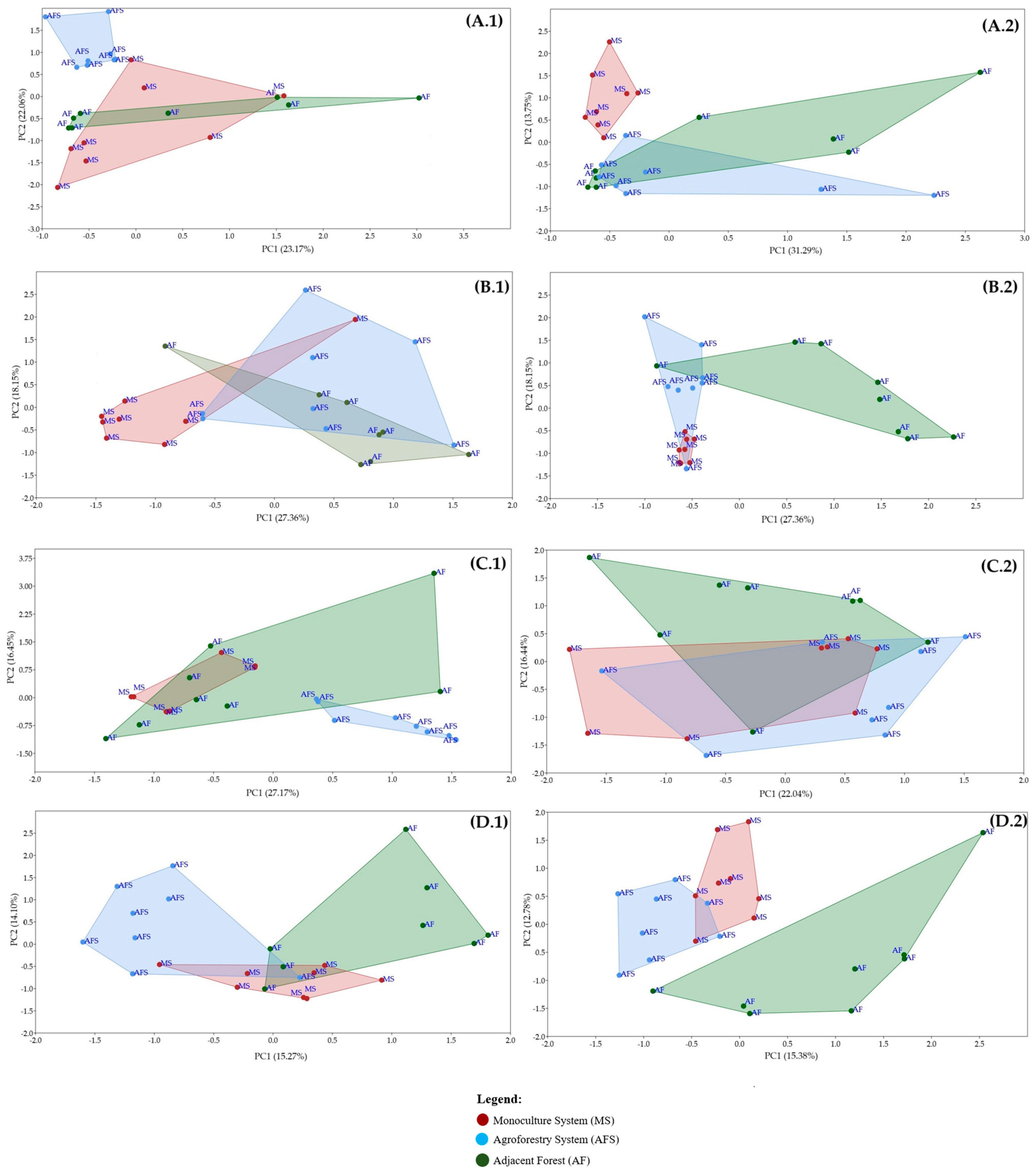
3.1. Analysis of the Microbial Community Structure and Bacterial Guilds Associated with PGPR
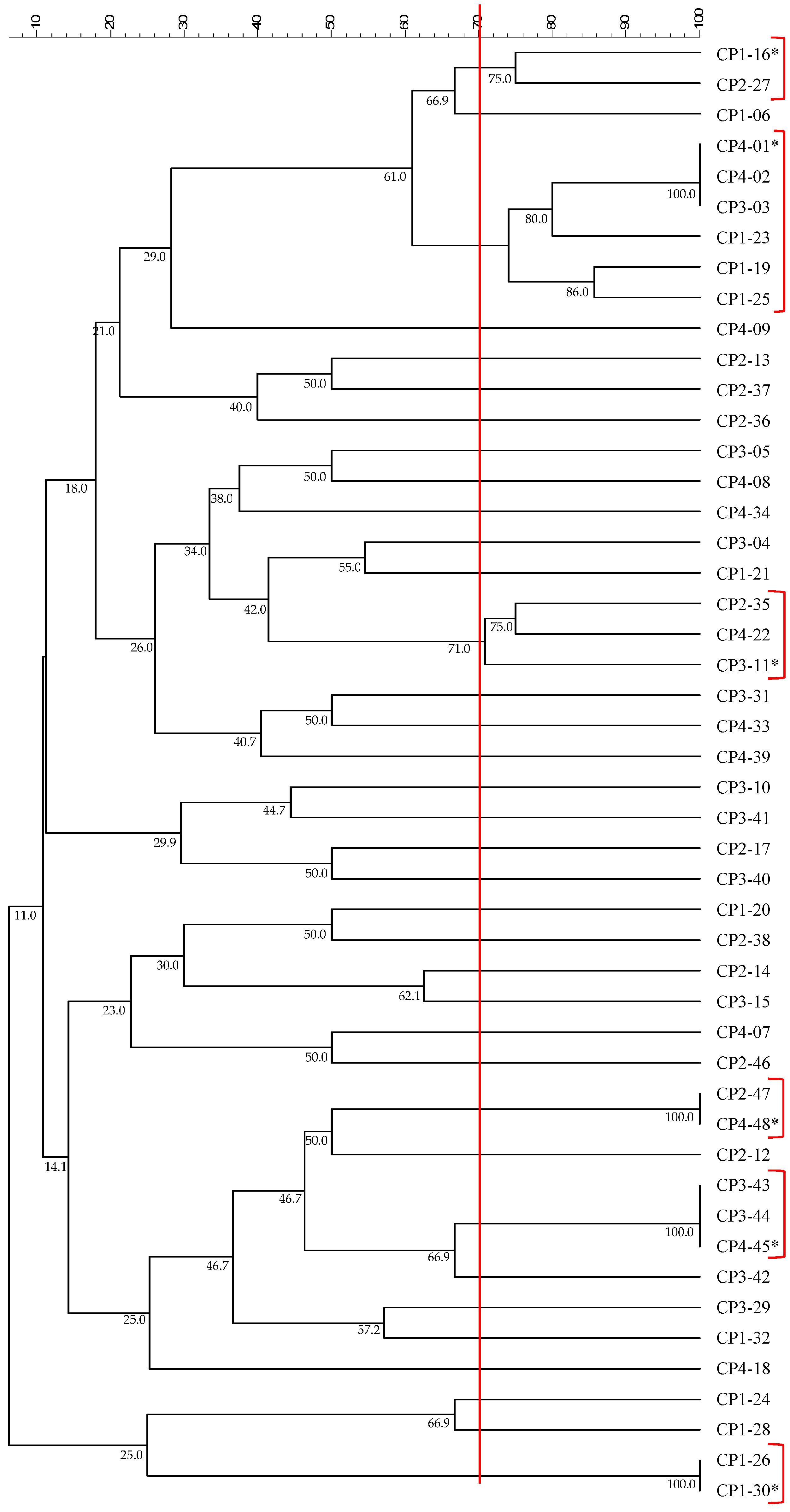
3.2. Isolation, Identification, and Characterization of Bacterial Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Souza, P.A.; Moreira, L.F.; Sarmento, D.H.; Da Costa, F.B. Cacao—Theobroma cacao. In Exotic Fruits; Academic Press: Cambridge, MA, USA, 2018; pp. 69–76. [Google Scholar]
- Chaves, S.F.D.S.; Alves, R.M.; Dias, L.A.D.S. Contribution of breeding to agriculture in the Brazilian Amazon. I. Açai palm and oil palm. Crop Breed. Appl. Biotechnol. 2021, 21. [Google Scholar] [CrossRef]
- Instituto Brasileiro de Geografia e Estatística (IBGE). Levantamento Sistemático de Produção Agrícola. 2022. Available online: https://ibge.gov.br/ (accessed on 7 October 2023).
- Instituto Brasileiro de Geografia e Estatistica (IBGE). Produção Agrícola Municipal. 2020. Available online: https://ibge.gov.br/ (accessed on 18 November 2023).
- Inpe. Monitoramento da Floresta Amazônica Brasileira por Satélite. Available online: http://terrabrasilis.dpi.inpe.br/ (accessed on 10 December 2023).
- Porro, R.; Miller, R.P.; Tito, M.R.; Donovan, J.A.; Vivan, J.L.; Trancoso, R.; Gonçalves, A.L. Agroforestry in the Amazon region: A pathway for balancing conservation and development. In Agroforestry—The Future of Global Land Use; Springer: Berlin/Heidelberg, Germany, 2012; pp. 391–428. [Google Scholar]
- Deheuvels, O.; Rousseau, G.X.; Soto Quiroga, G.; Decker Franco, M.; Cerda, R.; Vílchez Mendoza, S.J.; Somarriba, E. Biodiversity is affected by changes in management intensity of cocoa-based agroforests. AgroForestry Syst. 2014, 88, 1081–1099. [Google Scholar] [CrossRef]
- De Castro, F.; Futemma, C. Farm Knowledge Co-Production at an Old Amazonian Frontier: Case of the Agroforestry System in Tomé-Açu, Brazil. Rural Landsc. Soc. Environ. Hist. 2021, 8, 1081–1099. [Google Scholar] [CrossRef]
- Schmidt, J.E.; Firl, A.; Hamran, H.; Imaniar, N.I.; Crow, T.M.; Forbes, S.J. Impacts of shade trees on the adjacent cacao rhizosphere in a young diversified agroforestry system. Agronomy 2022, 12, 195. [Google Scholar] [CrossRef]
- Cherubin, M.R.; Chavarro-Bermeo, J.P.; Silva-Olaya, A.M. Agroforestry systems improve soil physical quality in northwestern Colombian Amazon. Agrofor. Syst. 2019, 93, 1741–1753. [Google Scholar] [CrossRef]
- Niether, W.; Jacobi, J.; Blaser, W.J.; Andres, C.; Armengot, L. Cocoa agroforestry systems versus monocultures: A multi-dimensional meta-analysis. Environ. Res. Lett. 2020, 15, 104085. [Google Scholar] [CrossRef]
- Banerjee, S.; Baah-Acheamfour, M.; Carlyle, C.N.; Bissett, A.; Richardson, A.E.; Siddique, T.; Chang, S.X. Determinants of bacterial communities in C anadian agroforestry systems. Environ. Microbiol. 2016, 18, 1805–1816. [Google Scholar] [CrossRef]
- Kim, N.; Zabaloy, M.C.; Guan, K.; Villamil, M.B. Do cover crops benefit soil microbiome? A meta-analysis of current research. Soil Biol. Biochem. 2020, 142, 107701. [Google Scholar] [CrossRef]
- Babalola, O.O.; Emmanuel, O.C.; Adeleke, B.S.; Odelade, K.A.; Nwachukwu, B.C.; Ayiti, O.E.; Adegboyega, T.T.; Igiehon, N.O. Rhizosphere microbiome cooperations: Strategies for sustainable crop production. Curr. Microbiol. 2021, 78, 1069–1085. [Google Scholar] [CrossRef]
- Thoms, D.; Liang, Y.; Haney, C.H. Maintaining symbiotic homeostasis: How do plants engage with beneficial microorganisms while at the same time restricting pathogens? Mol. Plant Microbe Interact. 2021, 34, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Wen, T.; Zhao, M.; Yuan, J.; Kowalchuk, G.A.; Shen, Q. Root exudates mediate plant defense against foliar pathogens by recruiting beneficial microbes. Soil Ecol. Lett. 2021, 3, 42–51. [Google Scholar] [CrossRef]
- Vives-Peris, V.; De Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep. 2020, 39, 3–17. [Google Scholar] [CrossRef]
- Santoyo, G.; Urtis-Flores, C.A.; Loeza-Lara, P.D.; Orozco-Mosqueda, M.D.C.; Glick, B.R. Rhizosphere colonization determinants by plant growth-promoting rhizobacteria (PGPR). Biology 2021, 10, 475. [Google Scholar] [CrossRef]
- De Castro, G.L.S.; Da Silva Júnior, D.D.; Viana, R.G.; Rêgo, M.C.F.; da Silva, G.B. Photosynthetic apparatus protection and drought effect mitigation in açai palm seedlings by rhizobacteria. Acta Physiol. Plant. 2019, 41, 163. [Google Scholar] [CrossRef]
- Yengwe, J.; Gebremikael, M.T.; Buchan, D.; Lungu, O.; De Neve, S. Effects of Faidherbia albida canopy and leaf litter on soil microbial communities and nitrogen mineralization in selected Zambian soils. Agrofor. Syst. 2018, 92, 349–363. [Google Scholar] [CrossRef]
- Tchakounté, G.V.T.; Berger, B.; Patz, S.; Fankem, H.; Ruppel, S. Community structure and plant growth-promoting potential of cultivable bacteria isolated from Cameroon soil. Microbiol. Res. 2018, 214, 47–59. [Google Scholar] [CrossRef]
- Benidire, L.; Pereira, S.I.A.; Castro, P.M.L.; Boularbah, A. Assessment of plant growth promoting bacterial populations in the rhizosphere of metallophytes from the Kettara mine, Marrakech. Environ. Sci. Pollut. Res. 2016, 23, 21751–21765. [Google Scholar] [CrossRef]
- De Castro, G.L.S.; Rêgo, M.C.F.; Silvestre, W.V.D.; Batista, T.F.V.; da Silva, G.B. Açaí palm seedling growth promotion by rhizobacteria inoculation. Braz. J. Microbiol. 2020, 51, 205–216. [Google Scholar] [CrossRef]
- Falcão, L.L.; Silva-Werneck, J.O.; Vilarinho, B.R.; Da Silva, J.P.; Pomella, A.W.V.; Marcellino, L.H. Antimicrobial and plant growth-promoting properties of the cacao endophyte Bacillus subtilis ALB629. J. Appl. Microbiol. 2014, 116, 1584–1592. [Google Scholar] [CrossRef] [PubMed]
- Claessen, M.E.C. Manual de métodos de análise de solo/Centro Nacional de Pesquisa de Solos.–2 ed. rev. Atual. In Documentos; EMBRAPA-CNPS: Rio de Janeiro, Brazil, 1997; Volume 1, 212p. [Google Scholar]
- Rodrigues, T.E.; Dos Santos, P.L.; Rollim, P.A.M.; Santos, E.; Rego, R.S.; Da Silva, J.M.L.; Gama, J.R.N. Caracterização e Classificação dos Solos do Município de Tomé-Açu, PA; Ministério da Agricultura, Pecuária e Abastecimento: Natal, Brazil, 2001. [Google Scholar]
- Muyzer, G.; De Waal, E.C.; Uitterlinden, A. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Nübel, U.; Engelen, B.; Felske, A.; Snaidr, J.; Wieshuber, A.; Amann, R.I.; Ludwig, W.; Backhaus, H. Sequence heterogeneities of genes encoding 16S rRNAs in Paenibacillus polymyxa detected by temperature gradient gel electrophoresis. J. Bacteriol. 1996, 178, 5636–5643. [Google Scholar] [CrossRef]
- Sakurai, M.; Wasaki, J.; Tomizawa, Y.; Shinano, T.; Osaki, M. Analysis of bacterial communities on alkaline phosphatase genes in soil supplied with organic matter. Soil Sci. Plant Nutr. 2008, 54, 62–71. [Google Scholar] [CrossRef]
- Poly, F.; Monrozier, L.J.; Bally, R. Improvement in the RFLP procedure for studying the diversity of nifH genes in communities of nitrogen fixers in soil. Res. Microbiol. 2001, 152, 95–103. [Google Scholar] [CrossRef]
- Simonet, P.; Grosjean, M.C.; Misra, A.K.; Nazaret, S.; Cournoyer, B.; Normand, P. Frankia genus-specific characterization by polymerase chain reaction. Appl. Environ. Microbiol. 1991, 57, 3278–3286. [Google Scholar] [CrossRef]
- Smit, E.; Leeflang, P.; Glandorf, B.; Dirk Van Elsas, J.; Wernars, K. Analysis of fungal diversity in the wheat rhizosphere by sequencing of cloned PCR-amplified genes encoding 18S rRNA and temperature gradient gel electrophoresis. Appl. Environ. Microbiol. 1999, 65, 2614–2621. [Google Scholar] [CrossRef]
- White, T.J.B.T. Analysis of phylogenetic relationships by amplification and direct sequencing of ribosomal RNA genes. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Heuer, H.; Wieland, G.; Schönfeld, J.; Schönwälder, A.; Gomes, N.C.M.; Smalla, K. Bacterial community profiling using DGGE or TGGE analysis. Environ. Mol. Microbiol. Protoc. Appl. 2001, 9, 177–190. [Google Scholar]
- Hammer, Ø.; Harper, D.A. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1. [Google Scholar]
- Marques, J.M.; da Silva, T.F.; Vollú, R.E.; De Lacerda, J.R.M.; Blank, A.F.; Smalla, K.; Seldin, L. Bacterial endophytes of sweet potato tuberous roots affected by the plant genotype and growth stage. Appl. Soil Ecol. 2015, 96, 273–281. [Google Scholar] [CrossRef]
- Seldin, L.; Dubnau, D. Deoxyribonucleic acid homology among Bacillus polymyxa, Bacillus macerans, Bacillus azotofixans, and other nitrogen-fixing Bacillus strains. Int. J. Syst. Evol. Microbiol. 1985, 35, 151–154. [Google Scholar] [CrossRef]
- Rademaker, J.L.W.; Louws, F.J.; Rossbach, U.; Vinuesa, P.; De Bruijn, F.J. Computer-assisted pattern analysis of molecular fingerprints and database construction. In Molecular Microbial Ecology Manual; Akkermans, A.D.L., van Elsas, J.D., de Bruijn, F.J., Eds.; Kluwer Academic Publishers: Amsterdam, The Netherlands, 1999; pp. 1–33. [Google Scholar]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: Hoboken, NJ, USA, 1991; pp. 115–147. [Google Scholar]
- Turner, S.; Pryer, K.M.; Miao, V.P.; Palmer, J.D. Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis 1. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.W.; Bonner, J. The enzymatic inactivation of indoleacetic acid; some characteristics of the enzyme contained in pea seedlings. Arch. Biochem. 1947, 13, 11–25. [Google Scholar] [PubMed]
- Mew, T.W.; Rosales, A.M. Bacterization of rice plants for control of sheath blight caused by Rhizoctonia solani. Phytopathology 1986, 76, 1260–1264. [Google Scholar] [CrossRef]
- Goswami, M.; Deka, S. Isolation of a novel rhizobacteria having multiple plant growth promoting traits and antifungal activity against certain phytopathogens. Microbiol. Res. 2020, 240, 126516. [Google Scholar] [CrossRef]
- Rosado, S. Phenotypic and genetic diversity of Paenibacillus azotofixans strains isolated from the rhizoplane or rhizosphere soil of different grasses. J. Appl. Microbiol. 1998, 84, 216–226. [Google Scholar] [CrossRef]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Ramos, H.M.N.; Vasconcelos, S.S.; Kato, O.R.; Castellani, D.C. Above-and belowground carbon stocks of two organic, agroforestry-based oil palm production systems in eastern Amazonia. Agrofor. Syst. 2018, 92, 221–237. [Google Scholar] [CrossRef]
- Prakash, D.; Benbi, D.K.; Saroa, G.S. Land-use effects on phosphorus fractions in Indo-Gangetic alluvial soils. Agrofor. Syst. 2018, 92, 437–448. [Google Scholar] [CrossRef]
- Hailemariam, M.; Birhane, E.; Gebresamuel, G.; Gebrekiros, A.; Desta, Y.; Alemayehu, A.; Muruts, H.; Araya, T.; Norgrove, L. Arbuscular mycorrhiza effects on Faidherbia albida (Del.) A. Chev. growth under varying soil water and phosphorus levels in Northern Ethiopia. Agrofor. Syst. 2018, 92, 485–498. [Google Scholar] [CrossRef]
- Torres-Gómez, M.; Garibay-Orijel, R.; Casas, A.; Pérez-Salicrup, D.R. Ectomycorrhizal trees intermingled within Cupressus lusitanica plantations sustain the diversity and availability of edible mushrooms. Agrofor. Syst. 2018, 92, 575–588. [Google Scholar] [CrossRef]
- Beule, L.; Vaupel, A.; Moran-Rodas, V.E. Abundance, diversity, and function of soil microorganisms in temperate alley-cropping agroforestry systems: A review. Microorganisms 2022, 10, 616. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.W.; Gama-Rodrigues, A.C. Sistemas agroflorestais com cacaueiro. In Ciência Tecnol. E Manejo Do Cacaueiro; CEPLAC/CEPEC: Ilhéus, Brazil, 2007; pp. 246–271. [Google Scholar]
- Xue, P.P.; Carrillo, Y.; Pino, V.; Minasny, B.; McBratney, A.B. Soil properties drive microbial community structure in a large scale transect in South Eastern Australia. Sci. Rep. 2018, 8, 11725. [Google Scholar] [CrossRef] [PubMed]
- Arévalo-Gardini, E.; Canto, M.; Alegre, J.; Arévalo-Hernández, C.O.; Loli, O.; Julca, A.; Baligar, V. Cacao agroforestry management systems effects on soil fungi diversity in the Peruvian Amazon. Ecol. Indic. 2020, 115, 106404. [Google Scholar] [CrossRef]
- Lori, M.; Armengot, L.; Schneider, M.; Schneidewind, U.; Bodenhausen, N.; Mäder, P.; Krause, H.M. Organic management enhances soil quality and drives microbial community diversity in cocoa production systems. Sci. Total Environ. 2022, 834, 155223. [Google Scholar] [CrossRef] [PubMed]
- De Vries, F.T.; Shade, A. Controls on soil microbial community stability under climate change. Front. Microbiol. 2013, 4, 265. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, Z.; Yuan, J. Impact of agro-farming activities on microbial diversity of acidic red soils in a Camellia Oleifera Forest. Rev. Bras. Ciência Solo 2019, 43, e0190044. [Google Scholar] [CrossRef]
- Wang, J.; Zou, Y.; Di Gioia, D.; Singh, B.K.; Li, Q. Conversion to agroforestry and monoculture plantation is detrimental to the soil carbon and nitrogen cycles and microbial communities of a rainforest. Soil Biol. Biochem. 2020, 147, 107849. [Google Scholar] [CrossRef]
- Buyer, J.S.; Baligar, V.C.; He, Z.; Arévalo-Gardini, E. Soil microbial communities under cacao agroforestry and cover crop systems in Peru. Appl. Soil Ecol. 2017, 120, 273–280. [Google Scholar] [CrossRef]
- Wang, J.; Ren, C.; Cheng, H.; Zou, Y.; Bughio, M.A.; Li, Q. Conversion of rainforest into agroforestry and monoculture plantation in China: Consequences for soil phosphorus forms and microbial community. Sci. Total Environ. 2017, 595, 769–778. [Google Scholar] [CrossRef]
- Tomich, T.P.; Noordwijk, M.V.; Budidarsono, S.; Gillison, A.; Kusumanto, T.; Murdiyarso, D.; Fagi, A.M. Agricultural intensification, deforestation and the environment: Assessing tradeoffs in Sumatra, Indonesia. In Tradeoffs or Synergies? Agricultural Intensification, Economic Development and the Environment; CABI Publishing: Wallingford, UK, 2001; pp. 221–244. [Google Scholar]
- Lacombe, S.; Bradley, R.L.; Hamel, C.; Beaulieu, C. Do tree-based intercropping systems increase the diversity and stability of soil microbial communities? Agric. Ecosyst. Environ. 2009, 131, 25–31. [Google Scholar] [CrossRef]
- Chifflot, V.; Rivest, D.; Olivier, A.; Cogliastro, A.; Khasa, D. Molecular analysis of arbuscular mycorrhizal community structure and spores distribution in tree-based intercropping and forest systems. Agric. Ecosyst. Environ. 2009, 131, 32–39. [Google Scholar] [CrossRef]
- Arévalo-Gardini, E.; Canto, M.; Alegre, J.; Loli, O.; Julca, A.; Baligar, V. Influence of agroforestry systems with cacao on soil properties (physical, chemical and microbiological) in Peruvian Amazon. In International Symposium on Cocoa Research (ISCR), Lima, Peru, 13–17 November 2017; International Cocoa Organization (ICCO): Abidjan, Côte d’Ivoire, 2018. [Google Scholar]
- Widyati, E.; Siarudin, M.; Indrajaya, Y. The dynamic of functional microbes community under auri (Acacia auriculiformis Cunn. Ex Benth) agroforestry system. J. Manaj. Hutan Trop. 2022, 28, 119. [Google Scholar] [CrossRef]
- Wang, H.; Cao, X.; Fan, W.; Deng, P.; Xu, X. Effects of intercropping systems of Phyllostachys edulis and Bletilla striata on soil bacterial community composition and function. Agrofor. Syst. 2023, 97, 617–630. [Google Scholar] [CrossRef]
- Masson-Boivin, C.; Giraud, E.; Perret, X.; Batut, J. Establishing nitrogen-fixing symbiosis with legumes: How many rhizobium recipes? Trends Microbiol. 2009, 17, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Leite, M.F.A.; Liu, B.; Gómez Cardozo, E.; Silva, H.R.E.; Luz, R.L.; Muchavisoy, K.H.M.; Kuramae, E.E. Microbiome resilience of Amazonian forests: Agroforest divergence to bacteria and secondary forest succession convergence to fungi. Glob. Change Biol. 2023, 29, 1314–1327. [Google Scholar] [CrossRef] [PubMed]
- McGrath, D.A.; Duryea, M.L.; Cropper, W.P. Soil phosphorus availability and fine root proliferation in Amazonian agroforests 6 years following forest conversion. Agric. Ecosyst. Environ. 2001, 83, 271–284. [Google Scholar] [CrossRef]
- Bell, M.C.; Raczkowski, C.W. Soil property indices for assessing short-term changes in soil quality. Renew. Agric. Food Syst. 2008, 23, 70–79. [Google Scholar] [CrossRef]
- Yao, M.K.; Koné, A.W.; Otinga, A.N.; Kassin, E.K.; Tano, Y. Carbon and nutrient cycling in tree plantations vs. natural forests: Implication for an efficient cocoa agroforestry system in West Africa. Reg. Environ. Change 2021, 21, 44. [Google Scholar] [CrossRef]
- De Schrijver, A.; Mertens, J.; Geudens, G.; Staelens, J.; Campforts, E.; Luyssaert, S.; Verheyen, K. Acidification of forested podzols in North Belgium during the period 1950–2000. Sci. Total Environ. 2006, 361, 189–195. [Google Scholar] [CrossRef]
- Kotowska, M.M.; Leuschner, C.; Triadiati, T.; Hertel, D. Conversion of tropical lowland forest reduces nutrient return through litterfall, and alters nutrient use efficiency and seasonality of net primary production. Oecologia 2016, 180, 601–618. [Google Scholar] [CrossRef] [PubMed]
- Suárez, L.R.; Salazar, J.C.S.; Casanoves, F.; Bieng, M.A.N. Cacao agroforestry systems improve soil fertility: Comparison of soil properties between forest, cacao agroforestry systems, and pasture in the Colombian Amazon. Agric. Ecosyst. Environ. 2021, 314, 107349. [Google Scholar] [CrossRef]
- Havlin, J.L.; Beaton, J.D.; Tisdale, S.L.; Nelson, W.L. Soil Fertility and Fertilizers: An Introduction to Nutrient Management, 7th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2005. [Google Scholar]
- Liu, C.; Jin, Y.; Hu, Y.; Tang, J.; Xiong, Q.; Xu, M.; Beng, K.C. Drivers of soil bacterial community structure and diversity in tropical agroforestry systems. Agric. Ecosyst. Environ. 2019, 278, 24–34. [Google Scholar] [CrossRef]
- Utomo, B.; Prawoto, A.A.; Bonnet, S.; Bangviwat, A.; Gheewala, S.H. Environmental performance of cocoa production from monoculture and agroforestry systems in Indonesia. J. Clean. Prod. 2016, 134, 583–591. [Google Scholar] [CrossRef]
- Ribaski, J.; Montoya Vilcahuaman, L.J.; Rodigheri, H.R. Sistemas agroflorestais: Aspectos ambientais e socioeconômicos. Inf. Agropecu. 2001, 22, 61–67. [Google Scholar]
- Munroe, J.W.; Isaac, M.E. N 2-fixing trees and the transfer of fixed-N for sustainable agroforestry: A review. Agron. Sustain. Dev. 2014, 34, 417–427. [Google Scholar] [CrossRef]
- Munroe, J.W.; Soto, G.; Virginio Filho, E.D.M.; Fulthorpe, R.; Isaac, M.E. Soil microbial and nutrient properties in the rhizosphere of coffee under agroforestry management. Appl. Soil Ecol. 2015, 93, 40–46. [Google Scholar] [CrossRef]
- Gao, P.; Zheng, X.; Wang, L.; Liu, B.; Zhang, S. Changes in the soil bacterial community in a chronosequence of temperate walnut-based intercropping systems. Forests 2019, 10, 299. [Google Scholar] [CrossRef]
- Gama-Rodrigues, E.F.; Ramachandran Nair, P.K.; Nair, V.D.; Gama-Rodrigues, A.C.; Baligar, V.C.; Machado, R.C. Carbon storage in soil size fractions under two cacao agroforestry systems in Bahia, Brazil. Environ. Manag. 2010, 45, 274–283. [Google Scholar] [CrossRef]
- Arévalo-Gardini, E.; Canto, M.; Alegre, J.; Loli, O.; Julca, A.; Baligar, V. Changes in soil physical and chemical properties in long term improved natural and traditional agroforestry management systems of cacao genotypes in Peruvian Amazon. PLoS ONE 2015, 10, e0132147. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, G.; Wu, Z.; Wen, X.; Zhong, H.; Zhong, Z.; Gai, X. Agroforestry alters the rhizosphere soil bacterial and fungal communities of moso bamboo plantations in subtropical China. Appl. Soil Ecol. 2019, 143, 192–200. [Google Scholar]
- Wartenberg, A.C.; Blaser, W.J.; Gattinger, A.; Roshetko, J.M.; Van Noordwijk, M.; Six, J. Does shade tree diversity increase soil fertility in cocoa plantations? Agric. Ecosyst. Environ. 2017, 248, 190–199. [Google Scholar] [CrossRef]
- Qu, Z.; Liu, B.; Ma, Y.; Sun, H. Differences in bacterial community structure and potential functions among Eucalyptus plantations with different ages and species of trees. Appl. Soil Ecol. 2020, 149, 103515. [Google Scholar] [CrossRef]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef]
- Luziatelli, F.; Gatti, L.; Ficca, A.G.; Medori, G.; Silvestri, C.; Melini, F.; Ruzzi, M. Metabolites secreted by a plant-growth-promoting Pantoea agglomerans strain improved rooting of Pyrus communis L. cv Dar Gazi cuttings. Front. Microbiol. 2020, 11, 539359. [Google Scholar] [CrossRef] [PubMed]
- Govindasamy, V.; Senthilkumar, M.; Magheshwaran, V.; Kumar, U.; Bose, P.; Sharma, V.; Annapurna, K. Bacillus and Paenibacillus spp.: Potential PGPR for sustainable agriculture. In Plant Growth and Health Promoting Bacteria; Springer: Berlin/Heidelberg, Germany, 2011; pp. 333–364. [Google Scholar]
- Kashyap, B.K.; Solanki, M.K.; Pandey, A.K.; Prabha, S.; Kumar, P.; Kumari, B. Bacillus as plant growth promoting rhizobacteria (PGPR): A promising green agriculture technology. In Plant Health Under Biotic Stress; Springer: Berlin/Heidelberg, Germany, 2019; Volume 2, pp. 219–236. [Google Scholar]
- Stoica, R.M.; Moscovici, M.; Tomulescu, C.; Cășărică, A.; Băbeanu, N.; Popa, O.; Kahraman, H.A. Antimicrobial compounds of the genus Bacillus: A review. Rom. Biotechnol. Lett. 2019, 24, 1111–1119. [Google Scholar] [CrossRef]
- Santos, J.F.D.; Sacramento, B.L.D.; Mota, K.N.A.B.; Souza, J.T.D.; Azevedo Neto, A.D.D. Crescimento de girassol em função da inoculação de sementes com bactérias endofíticas. Pesqui. Agropecu. Trop. 2014, 44, 142–150. [Google Scholar] [CrossRef]
- Om, A.C.; Ghazali, A.H.A.; Keng, C.L.; Ishak, Z. Microbial inoculation improves growth of oil palm plants (Elaeis guineensis Jacq.). Trop. Life Sci. Res. 2009, 20, 71. [Google Scholar] [PubMed]
- George, P.; Gupta, A.; Gopal, M.; Thomas, L.; Thomas, G.V. Multifarious beneficial traits and plant growth promoting potential of Serratia marcescens KiSII and Enterobacter sp. RNF 267 isolated from the rhizosphere of coconut palms (Cocos nucifera L.). World J. Microbiol. Biotechnol. 2013, 29, 109–117. [Google Scholar] [CrossRef]
- Belimov, A.A.; Hontzeas, N.; Safronova, V.I.; Demchinskaya, S.V.; Piluzza, G.; Bullitta, S.; Glick, B.R. Cadmium-tolerant plant growth-promoting bacteria associated with the roots of Indian mustard (Brassica juncea L. Czern.). Soil Biol. Biochem. 2005, 37, 241–250. [Google Scholar] [CrossRef]
- Tsavkelova, E.A.; Cherdyntseva, T.A.; Netrusov, A.I. Auxin production by bacteria associated with orchid roots. Microbiology 2005, 74, 46–53. [Google Scholar] [CrossRef]
- Berg, G.; Eberl, L.; Hartmann, A. The rhizosphere as a reservoir for opportunistic human pathogenic bacteria. Environ. Microbiol. 2005, 7, 1673–1685. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Ahmed, T.; Noman, M.; Javed, M.T.; Javed, M.R.; Tahir, M.; Shah, S.M. Non-pathogenic Staphylococcus strains augmented the maize growth through oxidative stress management and nutrient supply under induced salt stress. Ann. Microbiol. 2019, 69, 727–739. [Google Scholar] [CrossRef]
- Tullio, H.E. Potencial de Bactérias Endofíticas do Cacau para o Controle de Fungos de Solo e Promoção de Crescimento Radicular na Cultura da Soja. Master’s Thesis, Universidade Estadual de Ponta Grossa, Ponta Grossa, Brazil, 2017. [Google Scholar]
- Haidar, B.; Ferdous, M.; Fatema, B.; Ferdous, A.S.; Islam, M.R.; Khan, H. Population diversity of bacterial endophytes from jute (Corchorus olitorius) and evaluation of their potential role as bioinoculants. Microbiol. Res. 2018, 208, 43–53. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cropping System | Sampling Time | C | MO | N | N | Ratio C/N | P | K | Na | Al | Ca | Ca + Mg | pH |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| *g·Kg−1 | % | **mg·dm−3 | ***cmolc·dm−3 | H2O | |||||||||
| Monoculture (MS) | t1 | 15.2 | 26.2 | 0.8 | 0.1 | 20.2 | 86 | 91 | 8 | 0.1 | 2.3 | 3.2 | 5.1 |
| t2 | 9.7 | 16.7 | 0.3 | 0.03 | 32 | 73 | 86 | 3 | 0.04 | 2.15 | 3.07 | 5.1 | |
| Intercropping (AFS) | t1 | 10.8 | 18.7 | 0.9 | 0.1 | 12 | 27 | 45 | 11 | 0.1 | 4 | 5.1 | 6.2 |
| t2 | 15.5 | 15.5 | 0.8 | 0.08 | 11.9 | 17 | 33 | 4 | 0.07 | 3.07 | 4.2 | 5.09 | |
| Forest (AF) | t1 | 5.2 | 9 | 0.6 | 0.1 | 8.9 | 5 | 28 | 7 | 0.2 | 1.4 | 1.9 | 5.7 |
| t2 | 7.9 | 13.7 | 0.6 | 0.06 | 12.3 | 19 | 15 | 6 | 0.09 | 2.02 | 2.84 | 4.9 | |
| Sampling Time | rrs | alp | nifH | ITS |
|---|---|---|---|---|
| MS × MS (t1 × t2) | 0.0008 * | 0.0457 * | 0.0003 * | 0.0001 * |
| AFS × AFS (t1 × t2) | 0.0002 * | 0.0007 * | 0.0004 * | 0.0006 * |
| Cropping system | ||||
| MS × AF (t1) | 0.0843 | 0.0033 * | 0.1356 | 0.0006 * |
| MS × AFS (t1) | 0.0015 * | 0.0129 * | 0.0003 * | 0.0015 * |
| AFS × AF (t1) | 0.0006 * | 0.9033 | 0.012 * | 0.0006 * |
| MS × AF (t2) | 0.0027 * | 0.0015 * | 0.1212 | 0.0009 * |
| MS × AFS (t2) | 0.0012 * | 0.0015 * | 0.2019 | 0.003 * |
| AFS × AF (t2) | 0.3702 | 0.0048 * | 0.0063 * | 0.0015 * |
| BOX-PCR Group | Isolate | Closest Database Match (Accession Number), Identity (%) | Siderophore | PO4 org | PO4 inorg | IAA | AMS | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Curvularia | Fusarium | Pestalotiopsis | Pythium | Rhizoctonia | |||||||
| 1 | CP1-16 * | Rhodococcus pedocola (KT301938.1), 99 | + | - | - | + | - | - | - | - | - |
| CP2-27 | + | - | - | - | - | - | - | - | - | ||
| 2 | CP1-06 | Rhodococcus phenolicus (AY533293.1), 100 | + | - | - | - | - | - | - | - | - |
| 3 | CP4-01 * | Rhodococcus sp. (KT301938.1), 99 | + | - | - | - | - | - | - | - | - |
| CP4-02 | - | - | - | - | - | - | - | - | - | ||
| CP3-03 | + | + | + | - | - | - | - | - | - | ||
| CP1-19 | - | - | - | - | nd | nd | nd | nd | nd | ||
| CP1-23 | - | - | - | - | - | - | - | - | - | ||
| CP1-25 | + | - | - | - | - | - | - | - | - | ||
| 4 | CP4-09 | Agromyces indicus (HM036655.2), 99 | - | + | + | - | - | - | - | - | - |
| 5 | CP2-13 | Bacillus proteolyticus (KJ812418.1), 100 | - | - | - | - | + | + | + | + | + |
| 6 | CP2-37 | Bacillus paranthracis (KJ812420.1) 100 | + | - | - | - | - | - | - | - | - |
| 7 | CP2-36 | Staphylococcus hominis (X66101.1), 100 | - | - | - | - | - | - | - | - | - |
| 8 | CP3-05 | Lysinibacillus massilienses (AY677116.1), 100 | + | - | - | - | - | - | - | - | - |
| 9 | CP4-08 | Pantoea dispersa (DQ504305.1), 99 | + | + | - | - | - | - | - | - | - |
| 10 | CP4-34 | Leifsonia aquática (D45057.1), 99 | - | - | - | - | - | - | - | - | - |
| 11 | CP3-04 | Roseomonas rosea (AJ488505.1), 100 | + | - | - | - | nd | nd | nd | nd | nd |
| 12 | CP1-21 | Staphylococcus epidermidis (D83363.1), 100 | - | - | - | - | - | - | - | - | - |
| 13 | CP3-11 * | Leifsonia poae (AF116342.1), 99 | - | - | - | - | - | - | - | - | - |
| CP4-22 | - | - | - | - | - | - | - | - | - | ||
| CP2-35 | - | - | - | - | - | - | - | - | - | ||
| 14 | CP3-31 | Agromyces indicus (HM036655.2), 99 | - | - | - | - | - | - | - | - | - |
| 15 | CP4-33 | Leifsonia aquática (D45057.1), 99 | - | - | - | - | - | - | - | - | - |
| 16 | CP4-39 | Staphylococcus hominis (X66101.1), 98 | - | + | - | - | - | - | - | - | - |
| 17 | CP3-41 | Pantoea dispersa (Q504305.1), 98 | + | + | + | - | + | - | - | - | - |
| 18 | CP2-17 | Staphylococcus warneri (L37603.1), 99 | - | - | - | - | - | - | - | - | - |
| 19 | CP3-40 | Bacillus clarus (KNR180213.1), 99 | - | - | - | - | + | + | + | + | + |
| 20 | CP1-20 | Paenibacillus terrigena (AB248087.1), 99 | - | - | - | - | - | - | - | - | - |
| 21 | CP2-38 | Bacillus paranthracis (KJ812420.1), 100 | - | - | - | - | + | + | + | + | + |
| 22 | CP2-14 | Staphylococcus warneri (L37603.1), 99 | - | + | - | + | - | - | - | - | - |
| 23 | CP3-15 | Pantoea dispersa (DQ504305.1), 99 | - | - | - | - | - | - | - | - | - |
| 24 | CP4-07 | Staphylococccus epidermidis. (D83363.1), 99 | - | + | + | - | - | - | - | - | - |
| 25 | CP2-46 | Bacillus aerophilus (AJ831844.2), 99 | - | + | - | - | - | - | - | - | - |
| 26 | CP2-47 | Bacillus thuringiensis (D16281.1), 100 | - | - | - | - | + | + | + | + | + |
| CP4-48 * | - | + | - | - | - | - | - | - | - | ||
| 27 | CP2-12 | Lysinibacillus xylanilyticus (FJ477040.1), 100 | - | - | - | - | nd | nd | nd | nd | nd |
| 28 | CP3-43 | Staphylococcus sp. (D83363.1), 99 | - | + | - | - | - | - | - | - | - |
| CP3-44 | - | - | - | - | - | - | - | - | - | ||
| CP4-45 * | - | + | - | - | - | - | - | - | - | ||
| 29 | CP3-42 | Paenibacillus glucanolyticus (AB073189.1), 100 | - | - | - | - | - | - | - | - | - |
| 30 | CP3-29 | Bacillus aerophilus (AJ831844.2), 100 | + | - | - | - | - | - | - | - | - |
| 31 | CP1-32 | Microbacterium testaceum (X77445.1), 99 | - | - | - | - | + | + | + | + | + |
| 32 | CP4-18 | Staphylococcus epidermidis (D83363.1), 99 | - | + | - | - | - | - | - | - | - |
| 33 | CP1-24 | Bacillus tropicus (KJ812435.1), 99 | - | + | - | - | - | - | - | - | - |
| 34 | CP1-28 | Bacillus tropicus (KJ812435.1), 98 | - | + | - | - | + | + | + | + | + |
| 35 | CP2-50 | Staphylococcus hominis (X66101.1), 98 | - | - | - | - | - | - | - | - | - |
| 36 | CP1-26 | Bacillus wiedmannii (KU198626.1), 99 | - | - | - | - | - | - | - | - | - |
| CP1-30 * | - | - | - | - | + | + | + | - | + | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, R.d.S.d.R.d.; Lima, G.V.S.; Garcias, J.T.; Gomes, G.d.O.; Mateus, J.R.; Madeira, L.D.P.d.S.; Seldin, L.; Rogez, H.L.G.; Marques, J.M. The Microbial Community Structure in the Rhizosphere of Theobroma cacao L. and Euterpe oleracea Mart. Is Influenced by Agriculture System in the Brazilian Amazon. Microorganisms 2024, 12, 398. https://doi.org/10.3390/microorganisms12020398
Sousa RdSdRd, Lima GVS, Garcias JT, Gomes GdO, Mateus JR, Madeira LDPdS, Seldin L, Rogez HLG, Marques JM. The Microbial Community Structure in the Rhizosphere of Theobroma cacao L. and Euterpe oleracea Mart. Is Influenced by Agriculture System in the Brazilian Amazon. Microorganisms. 2024; 12(2):398. https://doi.org/10.3390/microorganisms12020398
Chicago/Turabian StyleSousa, Rosiane do Socorro dos Reis de, Giulia Victória Silva Lima, Josinete Torres Garcias, Graziane de Oliveira Gomes, Jackeline Rossetti Mateus, Lucimar Di Paula dos Santos Madeira, Lucy Seldin, Hervé Louis Ghislain Rogez, and Joana Montezano Marques. 2024. "The Microbial Community Structure in the Rhizosphere of Theobroma cacao L. and Euterpe oleracea Mart. Is Influenced by Agriculture System in the Brazilian Amazon" Microorganisms 12, no. 2: 398. https://doi.org/10.3390/microorganisms12020398