
Butyric Acid Supplementation Reduces Changes in the Taxonomic and Functional Composition of Gut Microbiota Caused by H. pylori Eradication Therapy

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
- (1)
- Patients of both sexes, aged 18 to 65;
- (2)
- Upper gastrointestinal endoscopy and H. pylori detection with at least one method performed within one month prior to enrollment into the study;
- (3)
- Signed written informed consent form;
- (4)
- Informed consent to comply with the same dietary and cooking procedures throughout the study period (all the patients were requested to fill in the questionnaire concerning their daily meals throughout the study period).
- (1)
- Endoscopy-confirmed gastric polyps or cancer/malignancy;
- (2)
- A history of concomitant diseases and conditions that might significantly affect the gut microbiota, such as the following:
- (a)
- Inflammatory bowel diseases;
- (b)
- Malabsorption syndrome associated with a documented disease of the small intestine, pancreas, etc.;
- (c)
- Cancers in any location;
- (d)
- Prior gastrointestinal surgeries (except for appendectomy);
- (e)
- The use of some drugs (immunosuppressive agents, cytostatics, steroids, antibiotics, and pre- and probiotics) within 3 months prior to enrollment into the study;
- (f)
- Functional bowel disorders, celiac disease, allergies, type 1 and type 2 diabetes, metabolic syndrome, non-alcoholic fatty liver disease, psychiatric disorders, etc., which, in the investigator’s view, may cause changes in the intestinal microbiota’s composition.
- (3)
- The use of bismuth-containing drugs (Bismuthate tripotassium dicitrate) for 4 weeks, and proton pump inhibitors (PPI) for 2 weeks, prior to H. pylori detection (except for cases using serology for H. pylori detection);
- (4)
- Alcohol or drug abuse;
- (5)
- Decompensation of chronic diseases (cardiovascular, respiratory, gastrointestinal, endocrine diseases, and liver and kidney failure);
- (6)
- Infectious and parasitic diseases, including human immunodeficiency virus (HIV), viral hepatitis, and tuberculosis;
- (7)
- Diarrhea (bowel movements more than 3 times a day) for at least 3 consecutive days during the last month;
- (8)
- Pregnancy or breastfeeding;
- (9)
- A history of first-line eradication therapy;
- (10)
- Inability or unwillingness to comply with the study procedures.
- (1)
- ECAB-14 (n = 11): esomeprazole 20 mg, clarithromycin 500 mg, amoxicillin 1000 mg, and bismuthate tripotassium dicitrate 240 mg, twice daily, per os, for 14 days;
- (2)
- ECAB-Z-14 (n = 11): esomeprazole 20 mg, clarithromycin 500 mg, amoxicillin 1000 mg, bismuthate tripotassium dicitrate 240 mg, twice daily, and butyric acid+inulin (Zacofalk) taken in 2 tablets (one tablet of butyric acid+inulin corresponds to 250 mg of butyric acid and 250 mg of inulin), daily, per os, for 14 days;
- (1)
- Prior to the beginning of eradication therapy to evaluate the baseline composition of the gut microbiota (time point I);
- (2)
- After the end of eradication therapy (within 14 (+3) days from the beginning of eradication therapy) to evaluate the influence of the eradication therapy on the gut microbiota (time point II);
- (3)
- Within a month (+5 days) of the end of eradication therapy to evaluate the long-term effects of H. pylori eradication on the gut microbiota (time point III).
2.2. Metagenomic Sequencing and Bioinformatic Analysis
2.3. Statistical Analysis
3. Results
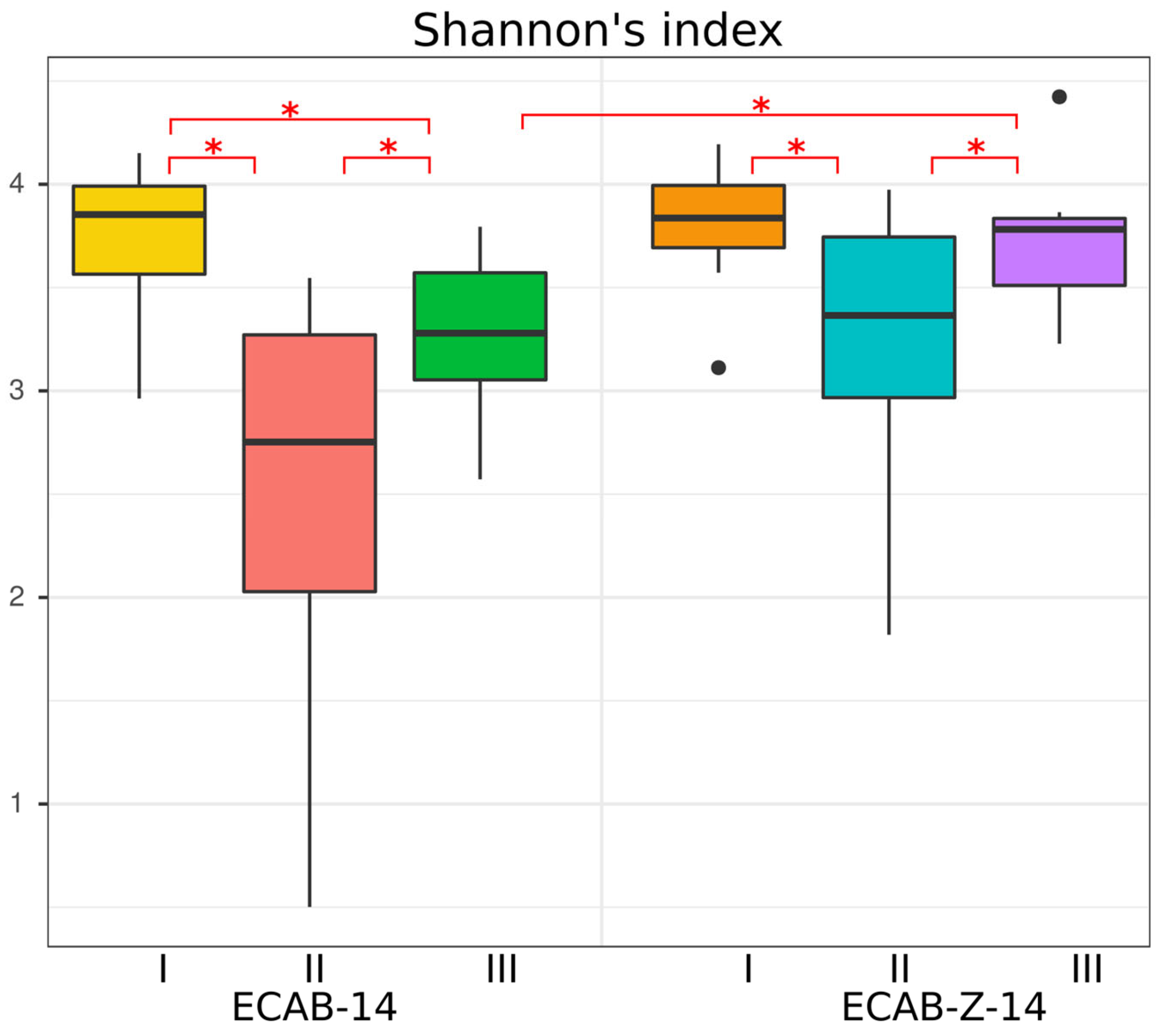
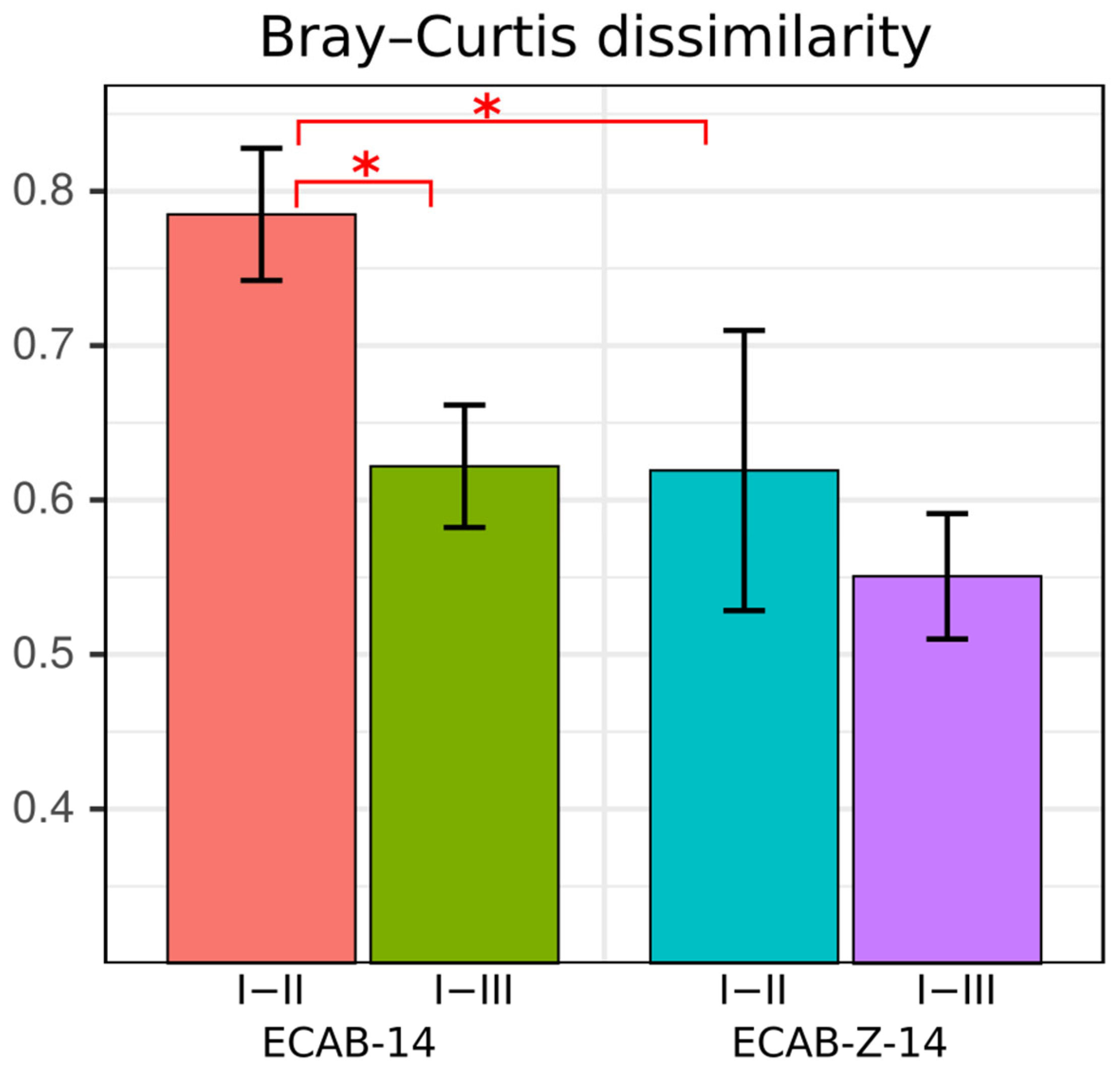
3.1. Diversity and Compositional Analysis
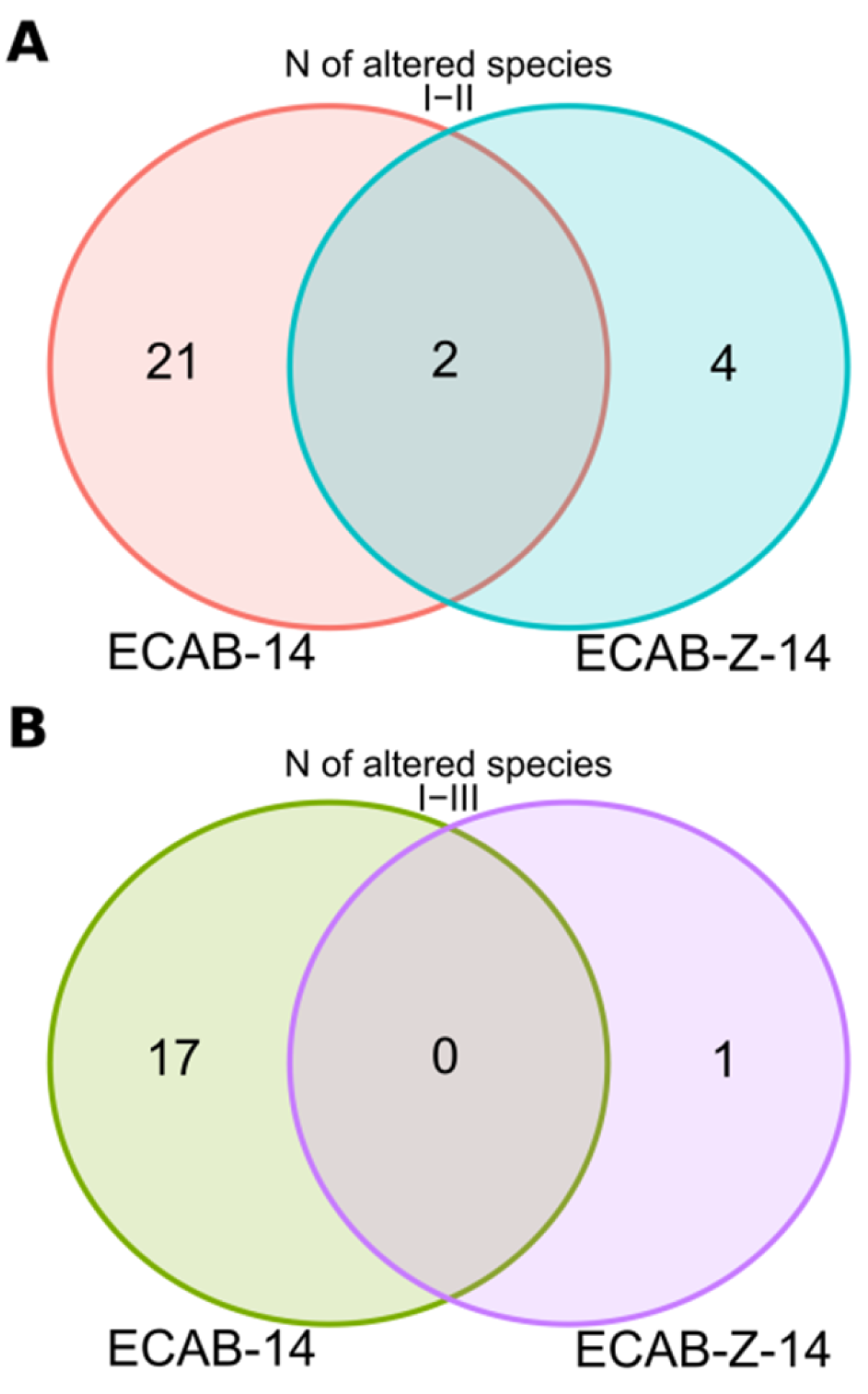
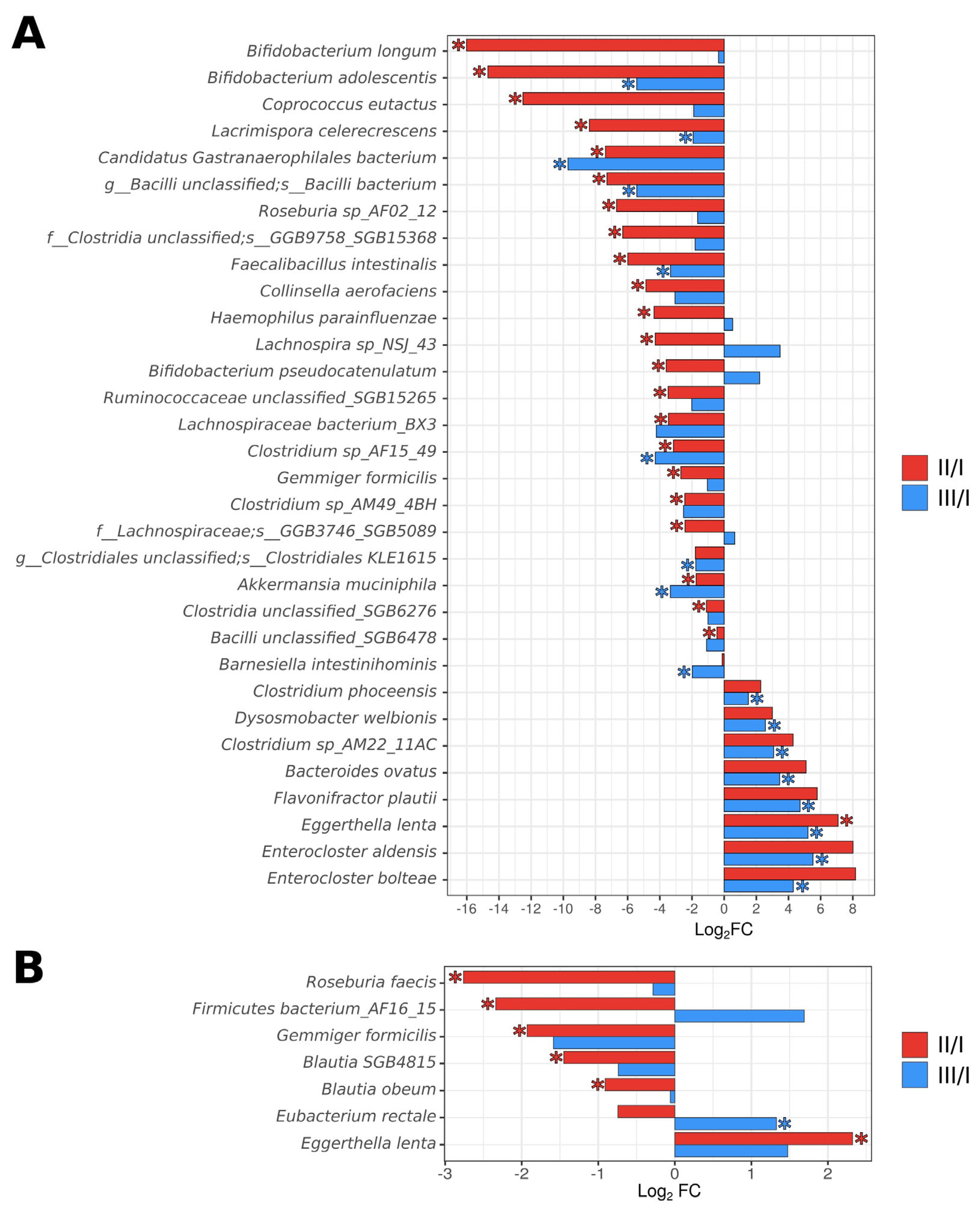
3.2. Analysis of the Taxonomic Composition of the Gut Microbiota
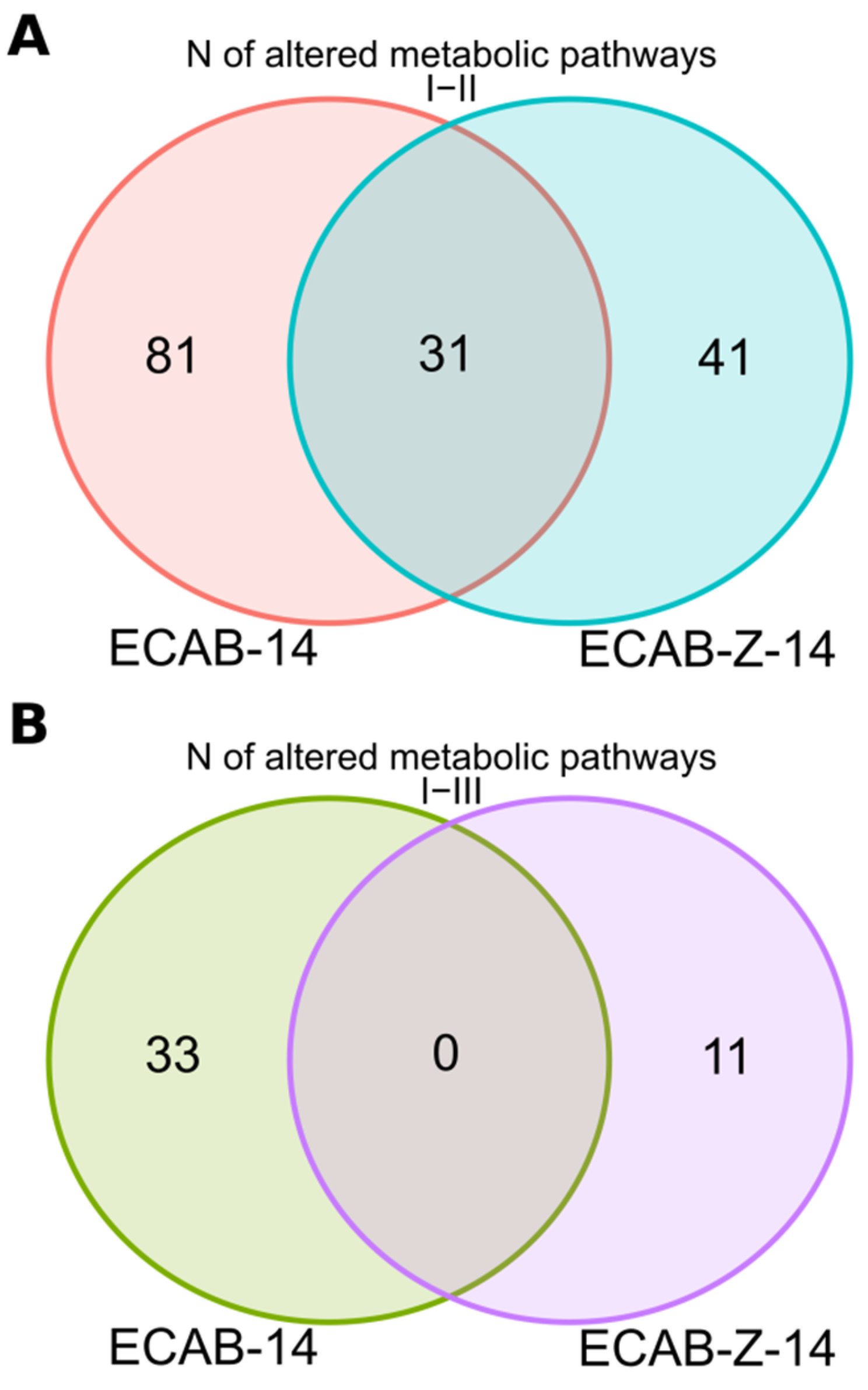
3.3. Functional Analysis
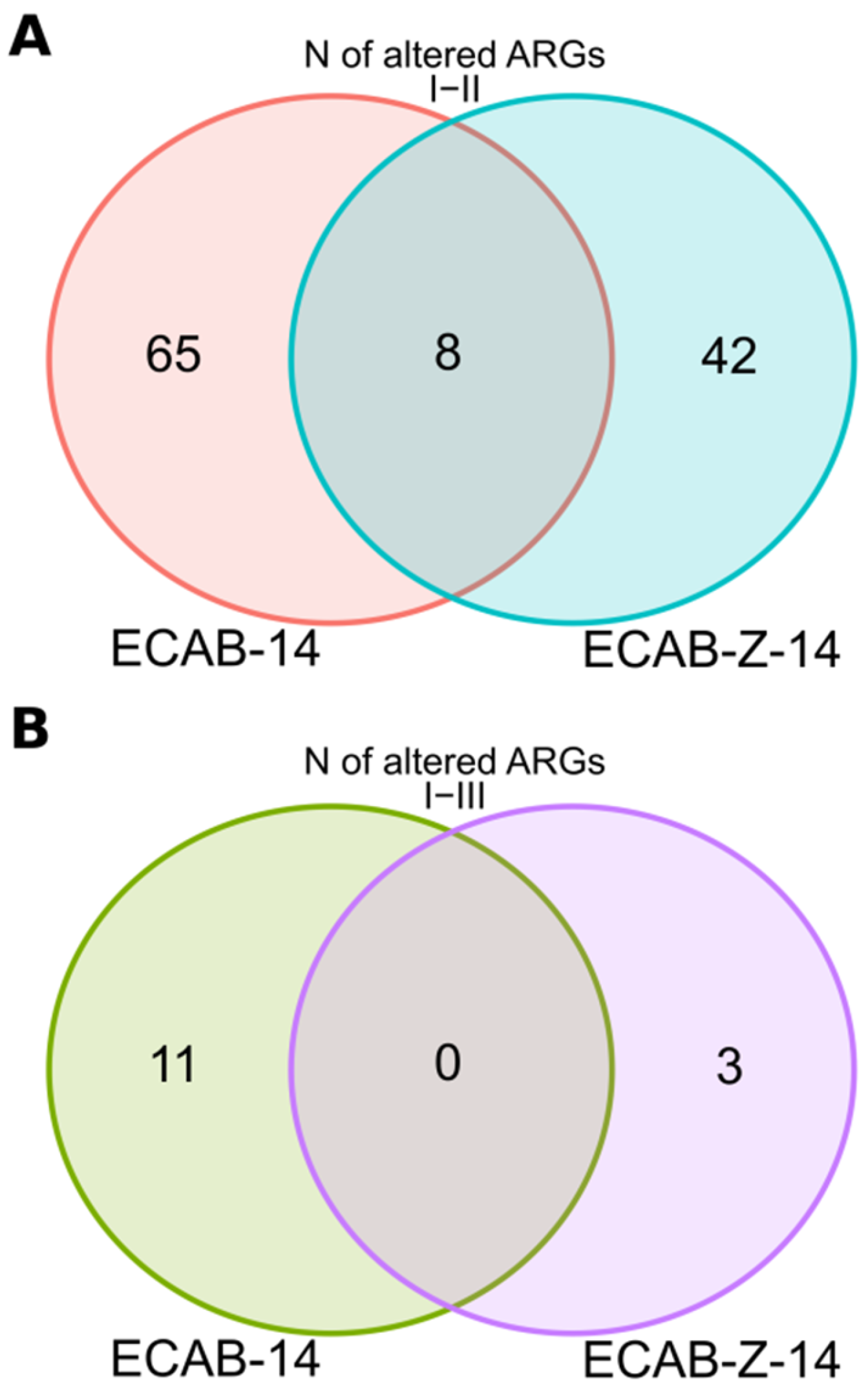
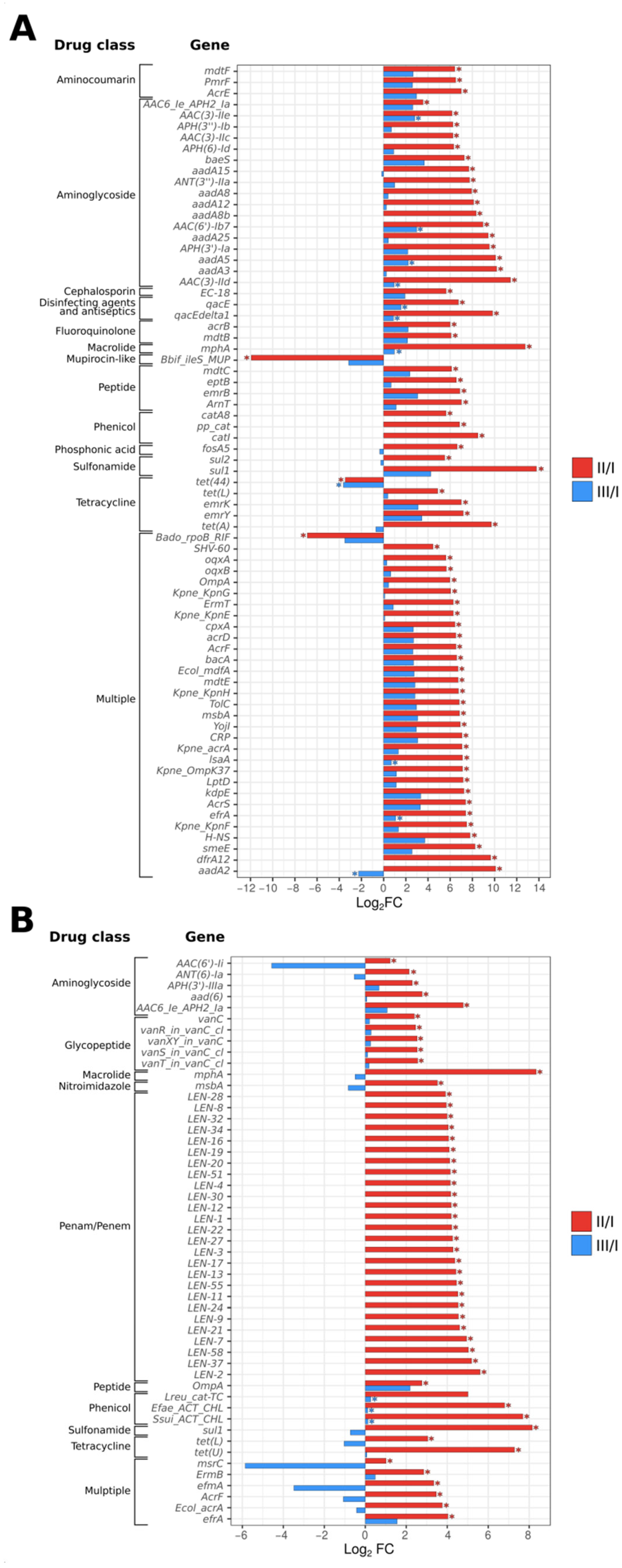
3.4. Gut Resistome Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malfertheiner, P.; Megraud, F.; Rokkas, T.; Gisbert, J.P.; Liou, J.-M.; Schulz, C.; Gasbarrini, A.; Hunt, R.H.; Leja, M.; O’Morain, C. Management of Helicobacter pylori Infection: The Maastricht VI/Florence Consensus Report. Gut 2022, 71, 1724–1762. [Google Scholar] [CrossRef] [PubMed]
- Sugano, K.; Tack, J.; Kuipers, E.J.; Graham, D.Y.; El-Omar, E.M.; Miura, S.; Haruma, K.; Asaka, M.; Uemura, N.; Malfertheiner, P. Kyoto Global Consensus Report on Helicobacter pylori Gastritis. Gut 2015, 64, 1353–1367. [Google Scholar] [CrossRef] [PubMed]
- Chey, W.D.; Leontiadis, G.I.; Howden, C.W.; Moss, S.F. ACG Clinical Guideline: Treatment of Helicobacter pylori Infection. Off. J. Am. Coll. Gastroenterol. ACG 2017, 112, 212–239. [Google Scholar] [CrossRef] [PubMed]
- Ivashkin, V.T.; Lapina, T.L.; Maev, I.V.; Drapkina, O.M.; Kozlov, R.S.; Sheptulin, A.A.; Trukhmanov, A.S.; Abdulkhakov, S.R.; Alekseeva, O.P.; Alekseenko, S.A.; et al. Clinical Practice Guidelines of Russian Gastroenterological Association, Scientific Society for the Clinical Study of Human Microbiome, Russian Society for the Prevention of Non-Communicable Diseases, Interregional Association for Clinical Microbiology and Antimicrobial Chemotherapy for H. pylori Diagnostics and Treatment in Adults. Russ. J. Gastroenterol. Hepatol. Coloproctol. 2022, 32, 72–93. [Google Scholar] [CrossRef]
- IARC; WHO. Helicobacter pylori Eradication as a Strategy for Preventing Gastric Cancer in 1994; International Agency for Research on Cancer: Lyon, France; World Health Organisation: Geneva, Switzerland, 2014; Available online: https://www.iarc.fr/en/media-centre/pr/2014/pdfs/pr227_E.pdf/ (accessed on 9 December 2023).
- Kamada, T.; Kurose, H.; Yamanaka, Y.; Manabe, N.; Kusunoki, H.; Shiotani, A.; Inoue, K.; Hata, J.; Matsumoto, H.; Akiyama, T. Relationship between Gastroesophageal Junction Adenocarcinoma and Helicobacter pylori Infection in Japan. Digestion 2012, 85, 256–260. [Google Scholar] [CrossRef]
- Wong, B.C.-Y.; Lam, S.K.; Wong, W.M.; Chen, J.S.; Zheng, T.T.; Feng, R.E.; Lai, K.C.; Hu, W.H.C.; Yuen, S.T.; Leung, S.Y. Helicobacter pylori Eradication to Prevent Gastric Cancer in a High-Risk Region of China: A Randomized Controlled Trial. JAMA 2004, 291, 187–194. [Google Scholar] [CrossRef]
- Rugge, M.; Genta, R.M.; Graham, D.Y.; Di Mario, F.; Coelho, L.G.V.; Kim, N.; Malfertheiner, P.; Sugano, K.; Tsukanov, V.; Correa, P. Chronicles of a Cancer Foretold: 35 Years of Gastric Cancer Risk Assessment. Gut 2016, 65, 721–725. [Google Scholar] [CrossRef]
- Wang, J.; Xu, L.; Shi, R.; Huang, X.; Li, S.W.H.; Huang, Z.; Zhang, G. Gastric Atrophy and Intestinal Metaplasia before and after Helicobacter pylori Eradication: A Meta-Analysis. Digestion 2011, 83, 253–260. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, Y.; Gerhard, M.; Gao, J.-J.; Mejias-Luque, R.; Zhang, L.; Vieth, M.; Ma, J.-L.; Bajbouj, M.; Suchanek, S. Effect of Helicobacter pylori on Gastrointestinal Microbiota: A Population-Based Study in Linqu, a High-Risk Area of Gastric Cancer. Gut 2020, 69, 1598–1607. [Google Scholar] [CrossRef]
- Zhou, Y.; Ye, Z.; Lu, J.; Miao, S.; Lu, X.; Sun, H.; Wu, J.; Wang, Y.; Huang, Y. Long-term Changes in the Gut Microbiota after 14-day Bismuth Quadruple Therapy in Penicillin-allergic Children. Helicobacter 2020, 25, e12721. [Google Scholar] [CrossRef]
- Hsu, P.; Pan, C.; Kao, J.Y.; Tsay, F.; Peng, N.; Kao, S.; Wang, H.; Tsai, T.; Wu, D.; Chen, C. Helicobacter pylori Eradication with Bismuth Quadruple Therapy Leads to Dysbiosis of Gut Microbiota with an Increased Relative Abundance of Proteobacteria and Decreased Relative Abundances of Bacteroidetes and Actinobacteria. Helicobacter 2018, 23, e12498. [Google Scholar] [CrossRef]
- Liou, J.-M.; Chen, C.-C.; Chang, C.-M.; Fang, Y.-J.; Bair, M.-J.; Chen, P.-Y.; Chang, C.-Y.; Hsu, Y.-C.; Chen, M.-J.; Chen, C.-C. Long-Term Changes of Gut Microbiota, Antibiotic Resistance, and Metabolic Parameters after Helicobacter pylori Eradication: A Multicentre, Open-Label, Randomised Trial. Lancet Infect. Dis. 2019, 19, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.; Pan, C.; Kao, J.Y.; Tsay, F.; Peng, N.; Kao, S.; Chen, Y.; Tsai, T.; Wu, D.; Tsai, K. Short-term and Long-term Impacts of Helicobacter pylori Eradication with Reverse Hybrid Therapy on the Gut Microbiota. J. Gastroenterol. Hepatol. 2019, 34, 1968–1976. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yao, H.; Tong, T.; Lau, K.; Leung, S.Y.; Ho, J.W.K.; Leung, W.K. Dynamic Changes in Antibiotic Resistance Genes and Gut Microbiota after Helicobacter pylori Eradication Therapies. Helicobacter 2022, 27, e12871. [Google Scholar] [CrossRef] [PubMed]
- Oh, B.; Kim, J.W.; Kim, B. Changes in the Functional Potential of the Gut Microbiome Following Probiotic Supplementation during Helicobacter pylori Treatment. Helicobacter 2016, 21, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Olekhnovich, E.I.; Manolov, A.I.; Samoilov, A.E.; Prianichnikov, N.A.; Malakhova, M.V.; Tyakht, A.V.; Pavlenko, A.V.; Babenko, V.V.; Larin, A.K.; Kovarsky, B.A. Shifts in the Human Gut Microbiota Structure Caused by Quadruple Helicobacter pylori Eradication Therapy. Front. Microbiol. 2019, 10, 1902. [Google Scholar] [CrossRef] [PubMed]
- Khusnutdinova, D.; Grigoryeva, T.; Abdulkhakov, S.; Safina, D.; Siniagina, M.; Markelova, M.; Boulygina, E.; Malanin, S.; Tyakht, A.; Kovarsky, B. Gut Microbiome Shotgun Sequencing in Assessment of Microbial Community Changes Associated with H. Pylori Eradication Therapy. Bionanoscience 2016, 6, 585–587. [Google Scholar] [CrossRef]
- Butorova, L.I.; Ardatskaya, M.D.; Osadchuk, M.A.; Kadnikova, N.G.; Lukianova, E.I.; Plavnik, R.G.; Sayutina, E.V.; Topchiy, T.B.; Tuayeva, E.M. Comparison of Clinical-Metabolic Efficacy of Pre- and Probiotics in the Conducted Optimized Protocols of Eradication Therapy of Helicobacter pylori Infection. Ter. Arkhiv 2020, 92, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Ianiro, G.; Bibbo, S.; Gasbarrini, A.; Cammarota, G. Therapeutic Modulation of Gut Microbiota: Current Clinical Applications and Future Perspectives. Curr. Drug Targets 2014, 15, 762–770. [Google Scholar] [CrossRef]
- Ianiro, G.; Tilg, H.; Gasbarrini, A. Antibiotics as Deep Modulators of Gut Microbiota: Between Good and Evil. Gut 2016, 65, gutjnl-2016. [Google Scholar] [CrossRef]
- Chen, L.; Xu, W.; Lee, A.; He, J.; Huang, B.; Zheng, W.; Su, T.; Lai, S.; Long, Y.; Chu, H. The Impact of Helicobacter pylori Infection, Eradication Therapy and Probiotic Supplementation on Gut Microenvironment Homeostasis: An Open-Label, Randomized Clinical Trial. EBioMedicine 2018, 35, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas, P.A.; Garcés, D.; Prado-Vivar, B.; Flores, N.; Fornasini, M.; Cohen, H.; Salvador, I.; Cargua, O.; Baldeón, M.E. Effect of Saccharomyces Boulardii CNCM I-745 as Complementary Treatment of Helicobacter pylori Infection on Gut Microbiome. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Tang, L.; Huang, C.; Tian, C.; Chen, L.; He, Z.; Yang, G.; Zuo, L.; Zhao, G.; Liu, E. The Effect of Probiotics Supplementation on Gut Microbiota after Helicobacter pylori Eradication: A Multicenter Randomized Controlled Trial. Infect. Dis. Ther. 2021, 10, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Liang, L.; Lv, P.; Liu, L.; Wang, S.; Wang, Z.; Chen, Y. Effects of Non-viable Lactobacillus Reuteri Combining with 14-day Standard Triple Therapy on Helicobacter pylori Eradication: A Randomized Double-blind Placebo-controlled Trial. Helicobacter 2021, 26, e12856. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Xiao, S.; Li, S.; Suo, B.; Wang, Y.; Meng, L.; Liu, Z.; Yin, Z.; Xue, Y.; Zhou, L. The Impact of Helicobacter pylori Infection, Eradication Therapy, and Probiotics Intervention on Gastric Microbiota in Young Adults. Helicobacter 2021, 26, e12848. [Google Scholar] [CrossRef]
- Nabavi-Rad, A.; Sadeghi, A.; Asadzadeh Aghdaei, H.; Yadegar, A.; Smith, S.M.; Zali, M.R. The Double-Edged Sword of Probiotic Supplementation on Gut Microbiota Structure in Helicobacter pylori Management. Gut Microbes 2022, 14, 2108655. [Google Scholar] [CrossRef]
- Biswas, I.; Mohapatra, P.K. Das Recent Advancement in Metabiotics: A Consortium with Bioactive Molecules after Fermentation by Probiotic Bacteria with Multidisciplinary Application Potential and Future Solution in Health Sector. Bioresour. Technol. Rep. 2023, 23, 101583. [Google Scholar] [CrossRef]
- Sharma, M.; Shukla, G. Metabiotics: One Step Ahead of Probiotics; an Insight into Mechanisms Involved in Anticancerous Effect in Colorectal Cancer. Front. Microbiol. 2016, 7, 1940. [Google Scholar] [CrossRef]
- Yang, N.; Lan, T.; Han, Y.; Zhao, H.; Wang, C.; Xu, Z.; Chen, Z.; Tao, M.; Li, H.; Song, Y. Tributyrin Alleviates Gut Microbiota Dysbiosis to Repair Intestinal Damage in Antibiotic-Treated Mice. PLoS ONE 2023, 18, e0289364. [Google Scholar] [CrossRef]
- Blaak, E.E.; Canfora, E.E.; Theis, S.; Frost, G.; Groen, A.K.; Mithieux, G.; Nauta, A.; Scott, K.; Stahl, B.; Van Harsselaar, J. Short Chain Fatty Acids in Human Gut and Metabolic Health. Benef. Microbes 2020, 11, 411–455. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of Short Chain Fatty Acids by the Gut Microbiota and Their Impact on Human Metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef]
- Huang, C.; Song, P.; Fan, P.; Hou, C.; Thacker, P.; Ma, X. Dietary Sodium Butyrate Decreases Postweaning Diarrhea by Modulating Intestinal Permeability and Changing the Bacterial Communities in Weaned Piglets. J. Nutr. 2015, 145, 2774–2780. [Google Scholar] [CrossRef]
- Mowat, A.M.; Agace, W.W. Regional Specialization within the Intestinal Immune System. Nat. Rev. Immunol. 2014, 14, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Meijer, K.; de Vos, P.; Priebe, M.G. Butyrate and Other Short-Chain Fatty Acids as Modulators of Immunity: What Relevance for Health? Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Vishwakarma, V.; Singhal, B. Metabiotics: The Functional Metabolic Signatures of Probiotics: Current State-of-Art and Future Research Priorities—Metabiotics: Probiotics Effector Molecules. Adv. Biosci. Biotechnol. 2018, 9, 147–189. [Google Scholar] [CrossRef]
- Tang, T.W.H.; Chen, H.-C.; Chen, C.-Y.; Yen, C.Y.T.; Lin, C.-J.; Prajnamitra, R.P.; Chen, L.-L.; Ruan, S.-C.; Lin, J.-H.; Lin, P.-J. Loss of Gut Microbiota Alters Immune System Composition and Cripples Postinfarction Cardiac Repair. Circulation 2019, 139, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Schropp, N.; Stanislas, V.; Michels, K.B.; Thriene, K. How Do Prebiotics Affect Human Intestinal Bacteria?—Assessment of Bacterial Growth with Inulin and XOS In Vitro. Int. J. Mol. Sci. 2023, 24, 12796. [Google Scholar] [CrossRef] [PubMed]
- Kruse, H.-P.; Kleessen, B.; Blaut, M. Effects of Inulin on Faecal Bifidobacteria in Human Subjects. Br. J. Nutr. 1999, 82, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Kolida, S.; Tuohy, K.; Gibson, G.R. Prebiotic Effects of Inulin and Oligofructose. Br. J. Nutr. 2002, 87, S193–S197. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, D.; Kathagen, G.; D’hoe, K.; Vieira-Silva, S.; Valles-Colomer, M.; Sabino, J.; Wang, J.; Tito, R.Y.; De Commer, L.; Darzi, Y. Quantitative Microbiome Profiling Links Gut Community Variation to Microbial Load. Nature 2017, 551, 507–511. [Google Scholar] [CrossRef]
- da Silva, T.F.; Casarotti, S.N.; de Oliveira, G.L.V.; Penna, A.L.B. The Impact of Probiotics, Prebiotics, and Synbiotics on the Biochemical, Clinical, and Immunological Markers, as Well as on the Gut Microbiota of Obese Hosts. Crit. Rev. Food Sci. Nutr. 2021, 61, 337–355. [Google Scholar] [CrossRef]
- Amabebe, E.; Robert, F.O.; Agbalalah, T.; Orubu, E.S.F. Microbial Dysbiosis-Induced Obesity: Role of Gut Microbiota in Homoeostasis of Energy Metabolism. Br. J. Nutr. 2020, 123, 1127–1137. [Google Scholar] [CrossRef]
- Le Bastard, Q.; Chapelet, G.; Javaudin, F.; Lepelletier, D.; Batard, E.; Montassier, E. The Effects of Inulin on Gut Microbial Composition: A Systematic Review of Evidence from Human Studies. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 403–413. [Google Scholar] [CrossRef]
- Jung, T.-H.; Jeon, W.-M.; Han, K.-S. In Vitro Effects of Dietary Inulin on Human Fecal Microbiota and Butyrate Production. J. Microbiol. Biotechnol. 2015, 25, 1555–1558. [Google Scholar] [CrossRef]
- Nista, E.C.; Candelli, M.; Finizio, R.; Cazzato, A.; Sparano, L.; Cammarota, G.; Gasbarrini, G.; Gasbarrini, A. Effect of Butyric Acid and Inulin Supplementation on Side Effects of Anti-H. Pylori Therapy: Preliminary Data. Dig. Liver Dis. 2006, 38, S72. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 9 December 2023).
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Truong, D.T.; Franzosa, E.A.; Tickle, T.L.; Scholz, M.; Weingart, G.; Pasolli, E.; Tett, A.; Huttenhower, C.; Segata, N. MetaPhlAn2 for Enhanced Metagenomic Taxonomic Profiling. Nat. Methods 2015, 12, 902–903. [Google Scholar] [CrossRef] [PubMed]
- Beghini, F.; McIver, L.J.; Blanco-Míguez, A.; Dubois, L.; Asnicar, F.; Maharjan, S.; Mailyan, A.; Manghi, P.; Scholz, M.; Thomas, A.M. Integrating Taxonomic, Functional, and Strain-Level Profiling of Diverse Microbial Communities with BioBakery 3. Elife 2021, 10, e65088. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef] [PubMed]
- Guillemard, E.; Poirel, M.; Schäfer, F.; Quinquis, L.; Rossoni, C.; Keicher, C.; Wagner, F.; Szajewska, H.; Barbut, F.; Derrien, M. A Randomised, Controlled Trial: Effect of a Multi-Strain Fermented Milk on the Gut Microbiota Recovery after Helicobacter pylori Therapy. Nutrients 2021, 13, 3171. [Google Scholar] [CrossRef] [PubMed]
- Quin, C.; Estaki, M.; Vollman, D.M.; Barnett, J.A.; Gill, S.K.; Gibson, D.L. Probiotic Supplementation and Associated Infant Gut Microbiome and Health: A Cautionary Retrospective Clinical Comparison. Sci. Rep. 2018, 8, 8283. [Google Scholar] [CrossRef] [PubMed]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; Van Immerseel, F. From the Gut to the Peripheral Tissues: The Multiple Effects of Butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.A.; Ward, L.C.; Brown, L. Inulin Oligofructose Attenuates Metabolic Syndrome in High-Carbohydrate, High-Fat Diet-Fed Rats. Br. J. Nutr. 2016, 116, 1502–1511. [Google Scholar] [CrossRef]
- De Cossio, L.F.; Fourrier, C.; Sauvant, J.; Everard, A.; Capuron, L.; Cani, P.D.; Layé, S.; Castanon, N. Impact of Prebiotics on Metabolic and Behavioral Alterations in a Mouse Model of Metabolic Syndrome. Brain Behav. Immun. 2017, 64, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Sheng, W.; Ji, G.; Zhang, L. Immunomodulatory Effects of Inulin and Its Intestinal Metabolites. Front. Immunol. 2023, 14, 1224092. [Google Scholar] [CrossRef] [PubMed]
- Moens, F.; De Vuyst, L. Inulin-Type Fructan Degradation Capacity of Clostridium Cluster IV and XIVa Butyrate-Producing Colon Bacteria and Their Associated Metabolic Outcomes. Benef. Microbes 2017, 8, 473–490. [Google Scholar] [CrossRef]
- Wang, Y.; Wan, X.; Wu, X.; Zhang, C.; Liu, J.; Hou, S. Eubacterium rectale Contributes to Colorectal Cancer Initiation via Promoting Colitis. Gut Pathog. 2021, 13, 2. [Google Scholar] [CrossRef]
- Ye, L.; Chan, E.W.C.; Chen, S. Selective and Suppressive Effects of Antibiotics on Donor and Recipient Bacterial Strains in Gut Microbiota Determine Transmission Efficiency of Bla NDM-1-Bearing Plasmids. J. Antimicrob. Chemother. 2019, 74, 1867–1875. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdulkhakov, S.; Markelova, M.; Safina, D.; Siniagina, M.; Khusnutdinova, D.; Abdulkhakov, R.; Grigoryeva, T. Butyric Acid Supplementation Reduces Changes in the Taxonomic and Functional Composition of Gut Microbiota Caused by H. pylori Eradication Therapy. Microorganisms 2024, 12, 319. https://doi.org/10.3390/microorganisms12020319
Abdulkhakov S, Markelova M, Safina D, Siniagina M, Khusnutdinova D, Abdulkhakov R, Grigoryeva T. Butyric Acid Supplementation Reduces Changes in the Taxonomic and Functional Composition of Gut Microbiota Caused by H. pylori Eradication Therapy. Microorganisms. 2024; 12(2):319. https://doi.org/10.3390/microorganisms12020319
Chicago/Turabian StyleAbdulkhakov, Sayar, Maria Markelova, Dilyara Safina, Maria Siniagina, Dilyara Khusnutdinova, Rustam Abdulkhakov, and Tatiana Grigoryeva. 2024. "Butyric Acid Supplementation Reduces Changes in the Taxonomic and Functional Composition of Gut Microbiota Caused by H. pylori Eradication Therapy" Microorganisms 12, no. 2: 319. https://doi.org/10.3390/microorganisms12020319