
The Role of Flagellum and Flagellum-Based Motility on Salmonella Enteritidis and Escherichia coli Biofilm Formation
 , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Plasmids
2.3. Gene Disruption
2.4. Eliminating Antibiotic Resistance Gene
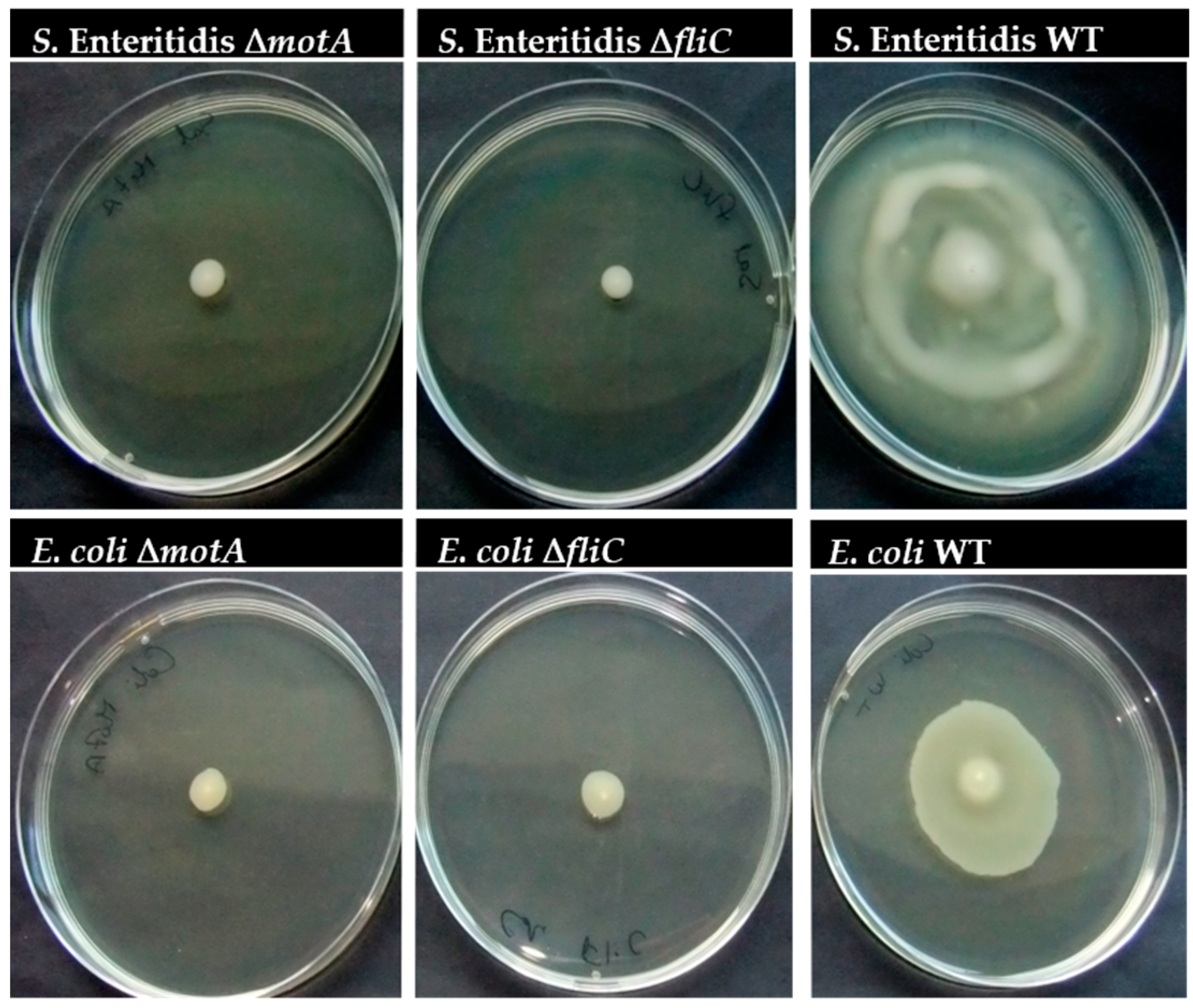
2.5. The Effect of motA or fliC Deletion on S. Enteritidis and E. coli Swarming
2.6. Biofilm Formation
2.7. Cultivability Assessment
2.8. Biomass Quantification by the CV Assay
2.9. Determination of the Fitness and Malthusian Parameter
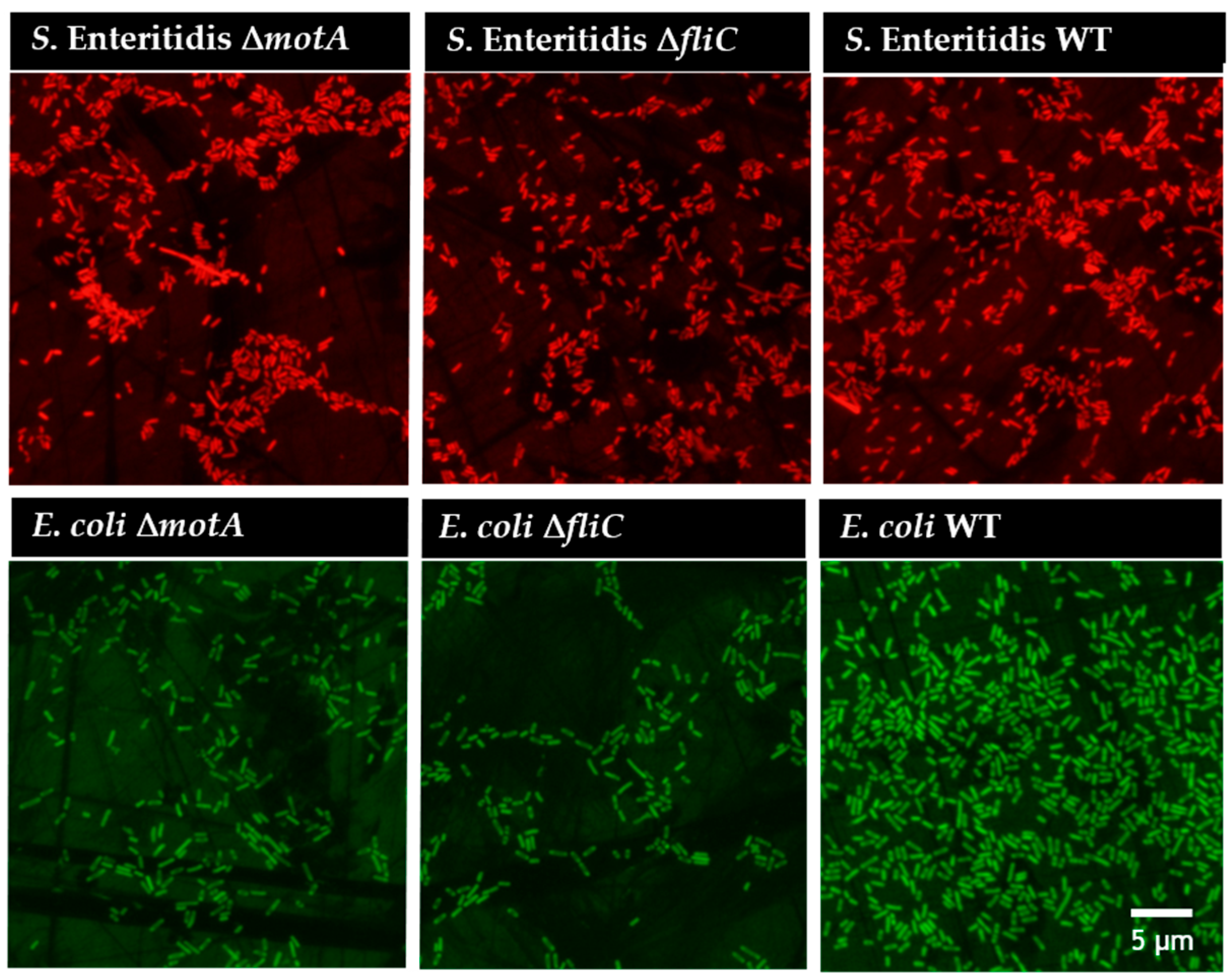
2.10. PNA FISH Hybridization and DAPI Staining
2.11. Statistical Analysis
3. Results
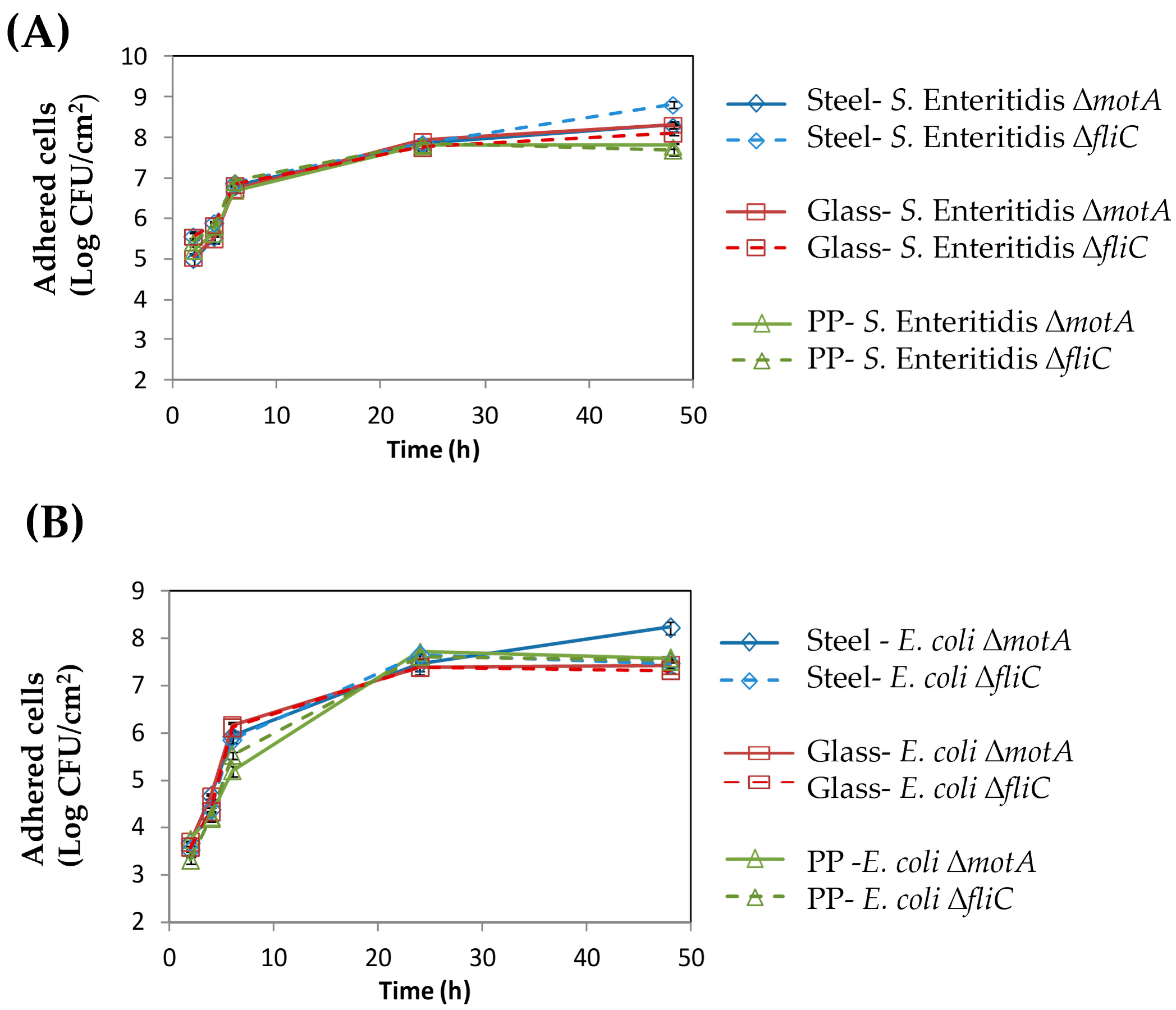
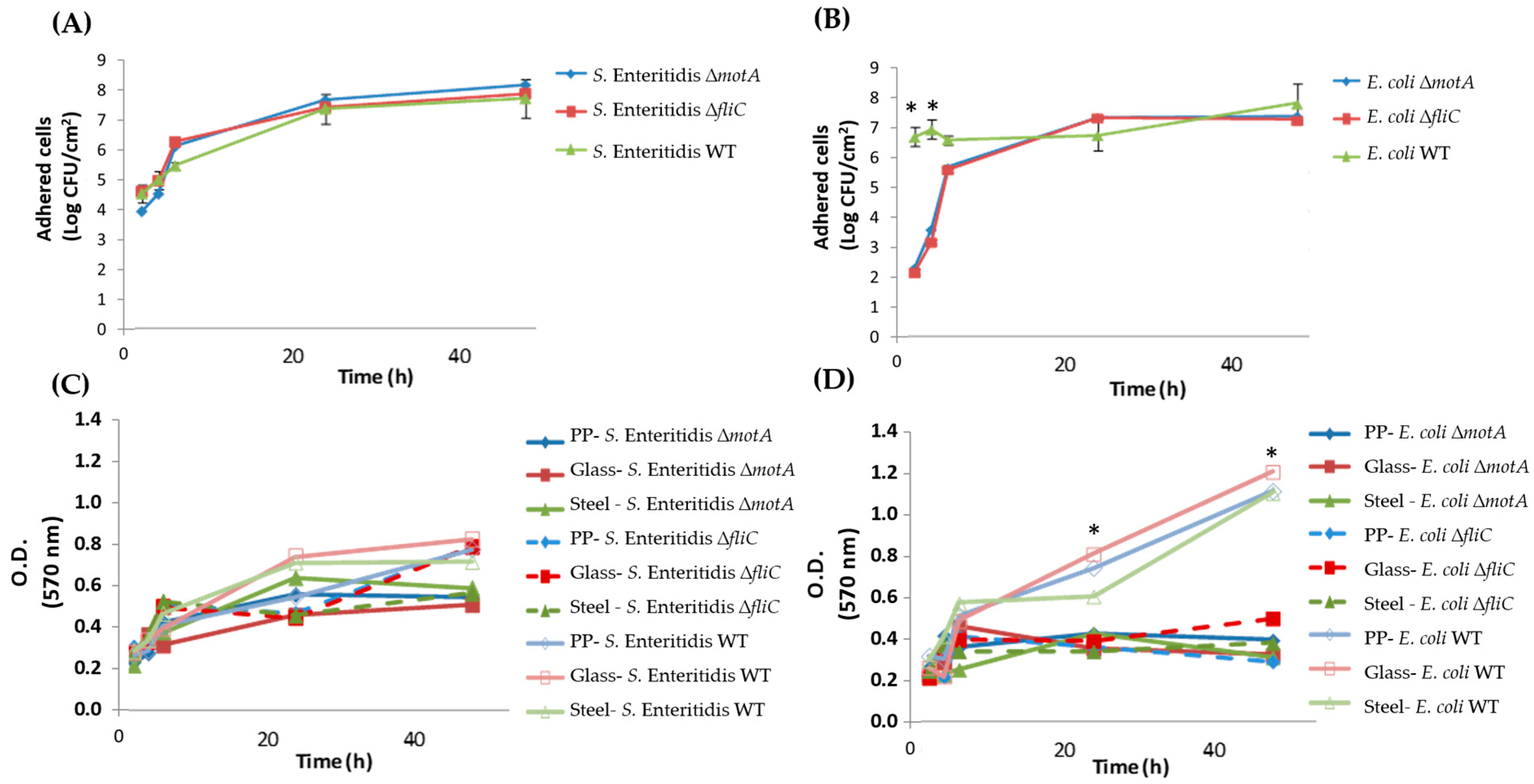
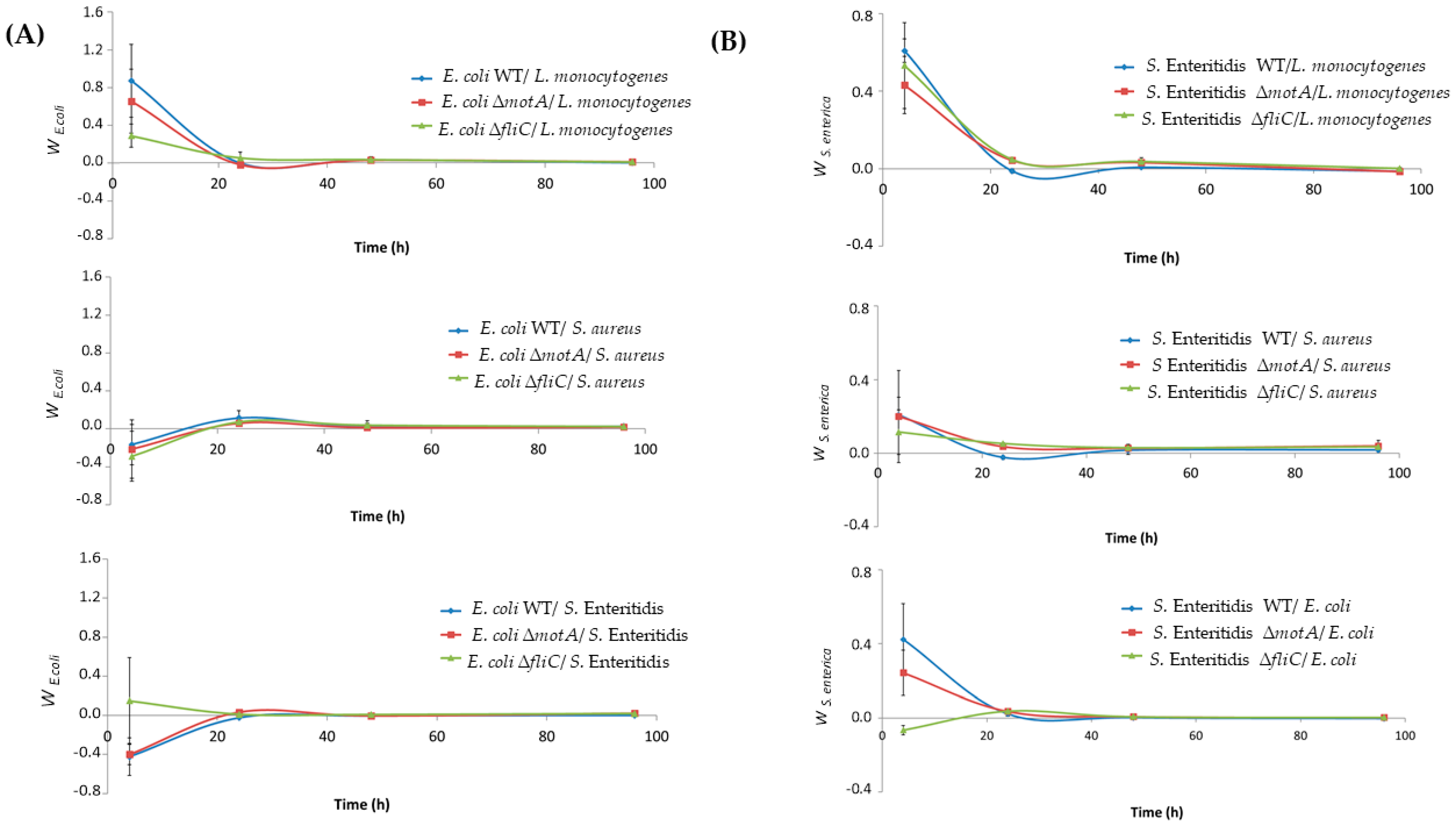
3.1. Flagellum and Flagellum-Based Motility Effect on S. Enteritidis and E. coli Biofilm Formation
3.2. Salmonella and E. coli Mutants Fitness in Dual-Species Biofilms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Stafford, G.P.; Hughes, C. Salmonella Typhimurium FlhE, a Conserved Flagellar Regulon Gene Required for Swarming. Microbiology 2007, 153, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Imada, K. Bacterial Flagellar Axial Structure and Its Construction. Biophys. Rev. 2018, 10, 559–570. [Google Scholar] [CrossRef]
- Duan, Q.; Zhou, M.; Zhu, L.; Zhu, G. Flagella and Bacterial Pathogenicity. J. Basic Microbiol. 2013, 53, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Wang, M.; Liu, Y.; Wu, P.; Yao, T.; Yang, W.; Yang, Q.; Yan, J.; Yang, B. Regulation of Flagellar Motility and Biosynthesis in Enterohemorrhagic Escherichia coli O157:H7. Gut Microbes 2022, 14, 2110822. [Google Scholar] [CrossRef] [PubMed]
- Benyoussef, W.; Deforet, M.; Monmeyran, A.; Henry, N. Flagellar Motility during E. coli Biofilm Formation Provides a Competitive Disadvantage Which Recedes in the Presence of Co-Colonizers. Front. Cell. Infect. Microbiol. 2022, 12, 896898. [Google Scholar] [CrossRef]
- Haiko, J.; Westerlund-Wikström, B. The Role of the Bacterial Flagellum in Adhesion and Virulence. Biology 2013, 2, 1242–1267. [Google Scholar] [CrossRef]
- Mahajan, A.; Currie, C.; Mackie, S.; Tree, J.; McAteer, S.; McKendrick, I.; McNeilly, R.; Woodward, M.; Gally, D.; Smith, D. An Investigation of the Expression and Adhesin function of H7 Flagella in the Interaction of Escherichia coli O157:H7 with Bovine Intestinal Epithelium. Cell Microbiol. 2009, 11, 121–137. [Google Scholar] [CrossRef] [PubMed]
- Coloma-Rivero, R.F.; Flores-Concha, M.; Molina, R.E.; Soto-Shara, R.; Cartes, Á.; Oñate, Á.A. Brucella and Its Hidden Flagellar System. Microorganisms 2022, 10, 83. [Google Scholar] [CrossRef]
- Friedlander, R.S.; Vogel, N.; Aizenberg, J. Role of Flagella in Adhesion of Escherichia coli to Abiotic Surfaces. Langmuir 2015, 31, 6137–6144. [Google Scholar] [CrossRef]
- Guttenplan, S.B.; Kearns, D.B. Regulation of Flagellar Motility during Biofilm Formation. FEMS Microbiol. Rev. 2013, 37, 849–871. [Google Scholar] [CrossRef]
- Pratt, L.A.; Kolter, R. Genetic Analysis of Escherichia coli Biofilm Formation: Roles of Flagella, Motility, Chemotaxis and Type I Pili. Mol. Microbiol. 1998, 30, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Crawford, R.W.; Reeve, K.E.; Gunn, J.S. Flagellated but Not Hyperfimbriated Salmonella Enterica Serovar Typhimurium Attaches to and Forms Biofilms on Cholesterol-Coated Surfaces. J. Bacteriol. 2010, 192, 2981–2990. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.H.; Liang, Y.H.; Chen, C.L.; Chiu, C.H. Characterization of Salmonella Resistance to Bile during Biofilm Formation. J. Microbiol. Immunol. Infect. 2020, 53, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Bowen, W.H.; Burne, R.A.; Wu, H.; Koo, H. Oral Biofilms: Pathogens, Matrix, and Polymicrobial Interactions in Microenvironments. Trends Microbiol. 2018, 26, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Rath, S.; Bal, S.C.B.; Dubey, D. Oral Biofilm: Development Mechanism, Multidrug Resistance, and Their Effective Management with Novel Techniques. Rambam Maimonides Med. J. 2021, 12, e0004. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-Step Inactivation of Chromosomal Genes in Escherichia coli K-12 Using PCR Products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef]
- Kalai Chelvam, K.; Chai, L.C.; Thong, K.L. Variations in Motility and Biofilm Formation of Salmonella Enterica Serovar Typhi. Gut Pathog. 2014, 6, 1–10. [Google Scholar] [CrossRef]
- Azevedo, N.F.; Pacheco, A.P.; Keevil, C.W.; Vieira, M.J. Adhesion of Water Stressed Helicobacter pylori to Abiotic Surfaces. J. Appl. Microbiol. 2006, 101, 718–724. [Google Scholar] [CrossRef]
- Almeida, C.; Azevedo, N.F.; Santos, S.; Keevil, C.W.; Vieira, M.J. Discriminating Multi-Species Populations in Biofilms with Peptide Nucleic Acid Fluorescence in Situ Hybridization (PNA FISH). PLoS ONE 2011, 6, e14786. [Google Scholar] [CrossRef]
- Azevedo, A.S.; Almeida, C.; Melo, L.F.; Azevedo, N.F. Interaction between Atypical Microorganisms and E. coli in Catheter-Associated Urinary Tract Biofilms. Biofouling 2014, 30, 893–902. [Google Scholar] [CrossRef]
- Almeida, C.; Azevedo, N.F.; Fernandes, R.M.; Keevil, C.W.; Vieira, M.J. Fluorescence in Situ Hybridization Method Using a Peptide Nucleic Acid Probe for Identification of Salmonella Spp. in a Broad Spectrum of Samples. Appl. Environ. Microbiol. 2010, 76, 4476–4485. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, A.S.; Almeida, C.; Pereira, B.; Madureira, P.; Wengel, J.; Azevedo, N.F. Detection and Discrimination of Biofilm Populations Using Locked Nucleic Acid/2′-O-Methyl-RNA Fluorescence in Situ Hybridization (LNA/2′OMe-FISH). Biochem. Eng. J. 2015, 104, 64–73. [Google Scholar] [CrossRef]
- Liu, N.T.; Bauchan, G.R.; Francoeur, C.B.; Shelton, D.R.; Lo, Y.M.; Nou, X. Ralstonia Insidiosa Serves as Bridges in Biofilm Formation by Foodborne Pathogens Listeria monocytogenes, Salmonella Enterica, and Enterohemorrhagic Escherichia coli. Food Control 2016, 65, 14–20. [Google Scholar] [CrossRef]
- Ma, A.; Neumann, N.; Chui, L. Phenotypic and Genetic Determination of Biofilm Formation in Heat Resistant Escherichia coli Possessing the Locus of Heat Resistance. Microorganisms 2021, 9, 403. [Google Scholar] [CrossRef] [PubMed]
- Białucha, A.; Gospodarek-Komkowska, E.; Kwiecińska-Piróg, J.; Skowron, K. Influence of Selected Factors on Biofilm Formation by Salmonella Enterica Strains. Microorganisms 2021, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Simões, L.C.; Simões, M.; Oliveira, R.; Vieira, M.J. Potential of the Adhesion of Bacteria Isolated from Drinking Water to Materials. J. Basic Microbiol. 2007, 47, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, D.; Chitolina, G.Z.; Wilsmann, D.E.; Lucca, V.; de Emery, B.D.; Borges, K.A.; Furian, T.Q.; Salle, C.T.P.; de Souza Moraes, H.L.; do Nascimento, V.P. Adhesion Capacity of Salmonella Enteritidis, Escherichia coli and Campylobacter jejuni on Polystyrene, Stainless Steel, and Polyethylene Surfaces. Food Microbiol. 2023, 114, 104280. [Google Scholar] [CrossRef]
- Huber, B.; Riedel, K.; Köthe, M.; Givskov, M.; Molin, S.; Eberl, L. Genetic Analysis of Functions Involved in the Late Stages of Biofilm Development in Burkholderia cepacia H111. Mol. Microbiol. 2002, 46, 411–426. [Google Scholar] [CrossRef]
- Wang, F.; Deng, L.; Huang, F.; Wang, Z.; Lu, Q.; Xu, C. Flagellar Motility Is Critical for Salmonella Enterica Serovar Typhimurium Biofilm Development. Front. Microbiol. 2020, 11, 499150. [Google Scholar] [CrossRef]
- Prouty, A.M.; Gunn, J.S. Comparative Analysis of Salmonella Enterica Serovar Typhimurium Biofilm Formation on Gallstones and on Glass. Infect. Immun. 2003, 71, 7154. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) * | |
|---|---|---|
| Recombination primers | ||
| E. coli | FliC_Fw | GCAGCAGAGACAGAACCTGCTGCGGTACCTGGTTAGCTTTGTGTAGGCTGGAGCTGCTTC |
| FliC_Rv | ATGGCACAAGTCATTAATACCAACAGCCTCTCGCTGATCAATGGGAATTAGCCATGGTCC | |
| MotAB_Fw | GTTTTCATGCAAAATGGCCTGTTCGGCTTGTTTGTTCAGTGTGTAGGCTGGAGCTGCTTC | |
| MotAB_Rv | GTGCTTATCTTATTAGGTTACCTGGTTGTTCTCGGTACAGATGGGAATTAGCCATGGTCC | |
| S. Enteritidis | FljB_Fw | TTAACGCAGTAAAGAGAGGACGTTTTGCGGAACCTGGTTGTGTAGGCTGGAGCTGCTTC |
| FljB_Rv | ATGGCACAAGTCATTAATACAAACAGCCTGTCGCTGTTGAATGGGAATTAGCCATGGTCC | |
| MotBA_Fw | GTTTTCATGCAAAATGGCCTGTTCCGCCTGTTTGTTTAACGTGTAGGCTGGAGCTGCTTC | |
| MotBA_Rv | GTGCTTATCTTATTAGGTTACCTGGTGGTTATCGGTACAGATGGGAATTAGCCATGGTCC | |
| Confirmation primers | ||
| E. coli | FliC Fw | TCAGGCAATTTGGCGTTGCCGTC |
| FliC Rv | CAGACGATAACAGGGTTGACGGC | |
| MotAB Fw | GCCAACAGTTCGTCCGCTTC | |
| MotAB Rv | CTGTCATGGTCAACAGTGGAAG | |
| S. Enteritidis | FliC Fw | GCAGGTTCAGTGACGGTGATT |
| FliC Rv | CGAAATTCAGGTGCCGATACA | |
| MotBA Fw | GCACCAAATCCAGCAGATGCTG | |
| MotBA_Rv | TTGCCTTGCCTTCGCGTTAATC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilas Boas, D.; Castro, J.; Araújo, D.; Nóbrega, F.L.; Keevil, C.W.; Azevedo, N.F.; Vieira, M.J.; Almeida, C. The Role of Flagellum and Flagellum-Based Motility on Salmonella Enteritidis and Escherichia coli Biofilm Formation. Microorganisms 2024, 12, 232. https://doi.org/10.3390/microorganisms12020232
Vilas Boas D, Castro J, Araújo D, Nóbrega FL, Keevil CW, Azevedo NF, Vieira MJ, Almeida C. The Role of Flagellum and Flagellum-Based Motility on Salmonella Enteritidis and Escherichia coli Biofilm Formation. Microorganisms. 2024; 12(2):232. https://doi.org/10.3390/microorganisms12020232
Chicago/Turabian StyleVilas Boas, Diana, Joana Castro, Daniela Araújo, Franklin L. Nóbrega, Charles W. Keevil, Nuno F. Azevedo, Maria João Vieira, and Carina Almeida. 2024. "The Role of Flagellum and Flagellum-Based Motility on Salmonella Enteritidis and Escherichia coli Biofilm Formation" Microorganisms 12, no. 2: 232. https://doi.org/10.3390/microorganisms12020232