
Increasing the Pentose Phosphate Pathway Flux to Improve Plasmid DNA Production in Engineered E. coli
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids
2.2. Precultures Development
2.3. Culture Conditions
2.4. Quantification of Concentration of Metabolites
2.5. Plasmid DNA Analyses
2.6. Measurement of the Glucose-6-Phosphate 1-Dehydrogenase (G6PDH) Activity
2.7. Calculations
2.8. Flux Balance Analysis
3. Results
3.1. Cultures of Untransformed Strains W3110 and VH34 in Mixtures of Glucose and Glycerol
3.2. Effect of recA Inactivation on pDNA Production in Strain VH34
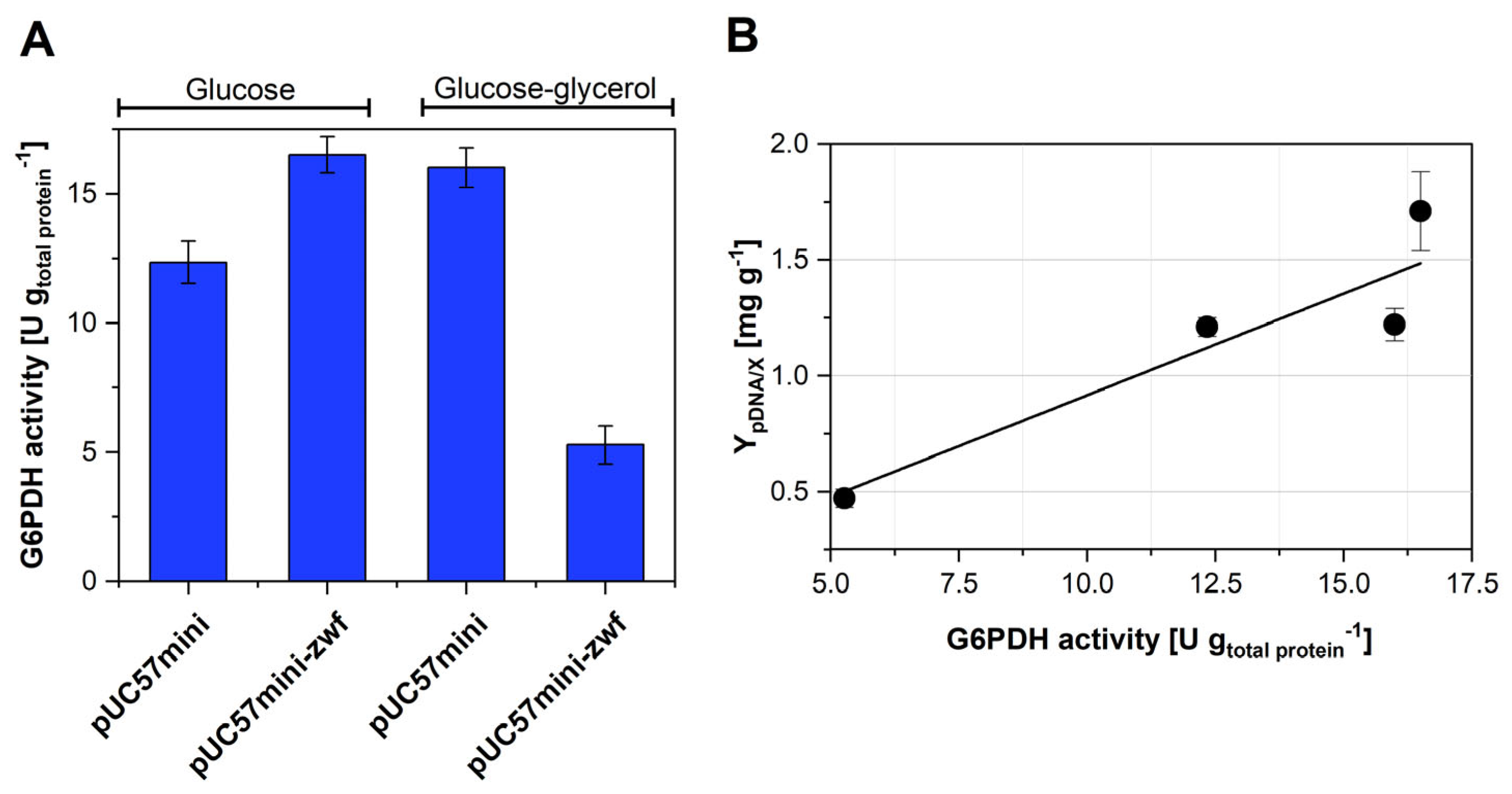
3.3. Strategies to Improve pDNA Production in Strain VH36
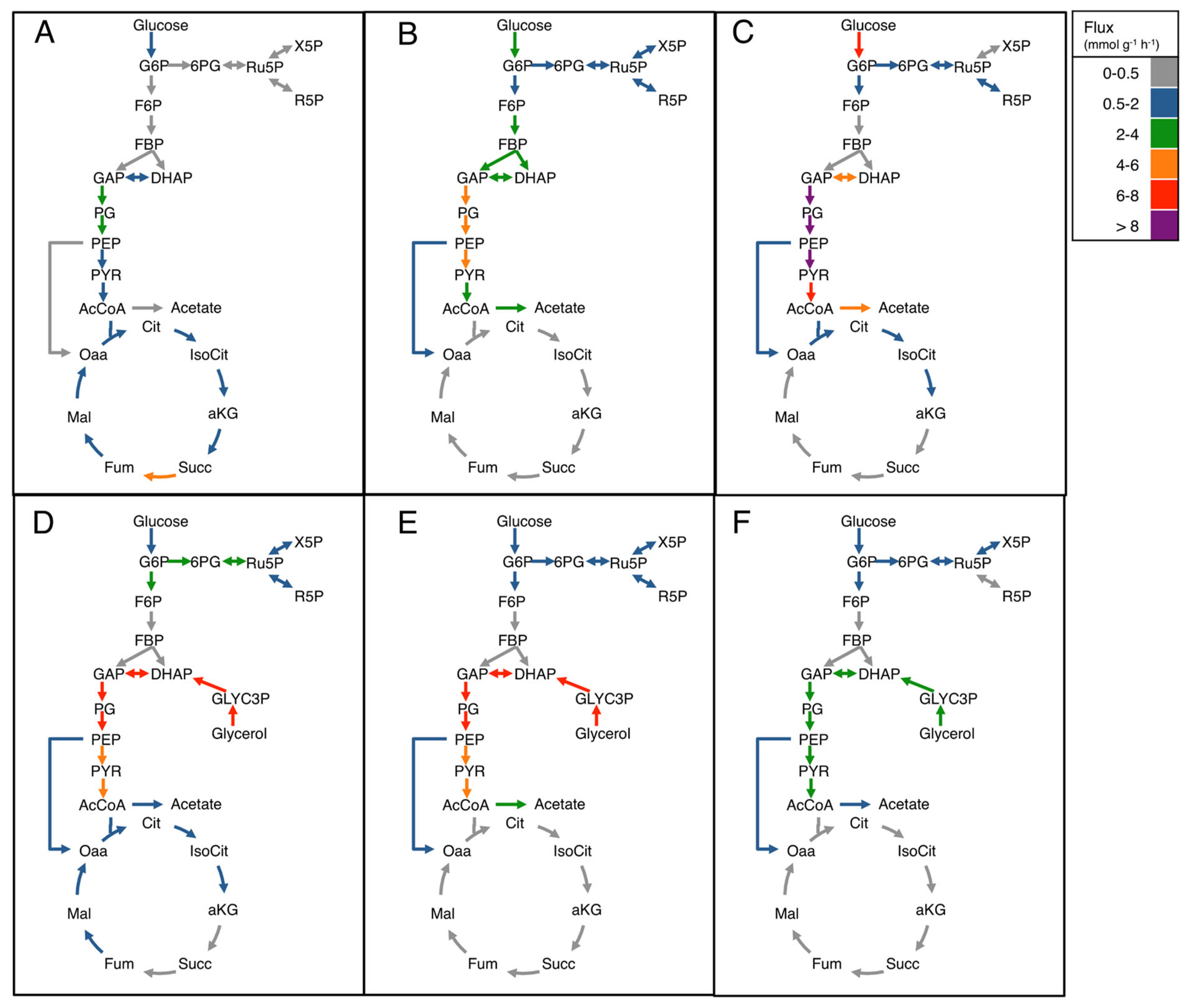
3.4. Flux Balance Analysis of pDNA Production in Strain VH36
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mairhofer, J.; Lara, A.R. Advances in Host and Vector Development for the Production of Plasmid DNA Vaccines. In Cancer Vaccines. Methods in Molecular Biology; Lawman, M., Lawman, P., Eds.; Humana Press: New York, NY, USA, 2014; Volume 1139, pp. 505–541. [Google Scholar]
- Rosa, S.S.; Prazeres, D.M.F.; Azevedo, A.M.; Marques, M.P.C. mRNA vaccines manufacturing: Challenges and bottlenecks. Vaccine 2021, 39, 2190–2200. [Google Scholar] [CrossRef] [PubMed]
- Ohlson, J. Plasmid manufacture is the bottleneck of the genetic medicine revolution. Drug Discov. Today 2020, 25, 1891–1893. [Google Scholar] [CrossRef]
- Eiteman, M.A.; Altman, E. Overcoming acetate in Escherichia coli recombinant protein fermentations. Trends Biotechnol. 2006, 24, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Taymaz-Nikerel, H.; Lara, A.R. Vitreoscilla Haemoglobin: A Tool to Reduce Overflow Metabolism. Microorganisms 2021, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Food and Drugs Administration of the United States of America. Guidance for Industry: Considerations for Plasmid DNA Vaccines for Infectious Disease Indications; Food and Drugs Administration of the United States of America: Washington, DC, USA, 2007. [Google Scholar]
- Gonçalves, G.A.L.; Bower, D.M.; Prazeres, D.M.F.; Monteiro, G.A.; Prather, K.L.J. Rational engineering of Escherichia coli strains for plasmid biopharmaceutical manufacturing. Biotechnol. J. 2012, 7, 251–261. [Google Scholar] [CrossRef]
- Gonçalves, G.A.L.; Bower, D.M.; Prazeres, D.M.F.; Monteiro, G.A.; Prather, K.L.J. De novo creation of MG1655-derived E. coli strains specifically designed for plasmid DNA production. Appl. Microbiol. Biotechnol. 2013, 97, 611–620. [Google Scholar] [CrossRef]
- Flores, S.; De Anda-Herrera, R.; Gosset, G.; Bolívar, F.G. Growth-rate recovery of Escherichia coli cultures carrying a multicopy plasmid, by engineering of the pentose-phosphate pathway. Biotechnol. Bioeng. 2004, 87, 485–494. [Google Scholar] [CrossRef]
- Wang, Z.; Xiang, L.; Shao, J.; Wegrzyn, A.; Wegrzyn, G. Effects of the presence of ColE1 plasmid DNA in Escherichia coli on the host cell metabolism. Microb. Cell Fact. 2006, 5, 34. [Google Scholar] [CrossRef]
- Williams, J.A.; Luke, J.; Langtry, S.; Anderson, S.; Hodgson, C.P.; Carnes, A.E. Generic plasmid DNA production platform incorporating low metabolic burden seed-stock and fed-batch fermentation processes. Biotechnol. Bioeng. 2009, 103, 1129–1143. [Google Scholar] [CrossRef]
- Soto, R.; Caspeta, L.; Barrón, B.L.; Gosset, G.; Ramírez, O.T.; Lara, A.R. High cell-density cultivation in batch mode for plasmid DNA vaccine production by a metabolically engineered E. coli strain with minimized overflow metabolism. Biochem. Eng. J. 2011, 56, 165–171. [Google Scholar] [CrossRef]
- Borja, M.G.; Meza, E.; Gosset, G.; Ramírez, O.T.; Lara, A.R. Engineering E. coli to increase plasmid DNA production in high cell-density cultivations in batch mode. Microb. Cell Fact. 2012, 11, 132. [Google Scholar] [CrossRef] [PubMed]
- Meza, E.; Becker, J.; Bolivar, F.; Gosset, G.; Wittmann, C. Consequences of phosphoenolpyruvate:sugar phosphotranferase system and pyruvate kinase isozymes inactivation in central carbon metabolism flux distribution in Escherichia coli. Microb. Cell Fact. 2012, 11, 127. [Google Scholar] [CrossRef] [PubMed]
- Wunderlich, M.; Taymaz-Nikerel, H.; Gosset, G.; Ramírez, O.T.; Lara, A.R. Effect of growth rate on plasmid DNA production and metabolic performance of engineered Escherichia coli strains. J. Biosci. Bioeng. 2014, 117, 336–342. [Google Scholar] [CrossRef] [PubMed]
- de la Cruz, M.; Ramírez, E.A.; Sigala, J.C.; Utrilla, J.; Lara, A.R. Plasmid DNA Production in Proteome-Reduced Escherichia coli. Microorganisms 2020, 8, 1444. [Google Scholar] [CrossRef] [PubMed]
- Anderlei, T.; Zang, W.; Papaspyrou, M.; Büchs, J. Online respiration activity measurement (OTR, CTR, RQ) in shake flasks. Biochem. Eng. J. 2004, 17, 187–194. [Google Scholar] [CrossRef]
- Schmidt, T.; Friehs, K.; Flaschel, E. Structures of plasmid DNA. In Plasmids for Therapy and Vaccination; Schleef, M., Ed.; Wiley-VCH Verlag GmBH: Weinheim, Germany, 2001; pp. 29–43. [Google Scholar]
- Kielkopf, C.L.; Bauer, W.; Urbatsch, I.L. Bradford Assay for Determining Protein Concentration. Cold Spring Harb. Protoc. 2020, 4, 102269. [Google Scholar] [CrossRef]
- Orth, J.D.; Conrad, T.M.; Na, J.; Lerman, J.A.; Nam, H.; Feist, A.M.; Palsson, B. A comprehensive genome-scale reconstruction of Escherichia coli metabolism. Mol. Syst. Biol. 2011, 7, 535. [Google Scholar] [CrossRef]
- Schellenberger, J.; Que, R.; Fleming, R.; Thiele, I.; Orth, J.; Feist, A.; Zielinski, D.; Bordbar, A.; Lewis, N.; Rahmanian, S.; et al. Quantitative prediction of cellular metabolism with constraint-based models: The COBRA Toolbox v2.0. Nat. Protoc. 2011, 6, 1290–1307. [Google Scholar] [CrossRef]
- Taymaz-Nikerel, H.; Borujeni, A.E.; Verheijen, P.J.; Heijnen, J.J.; van Gulik, W.M. Genome-derived minimal metabolic models for Escherichia coli MG1655 with estimated in vivo respiratory ATP stoichiometry. Biotechnol. Bioeng. 2010, 107, 369–381. [Google Scholar] [CrossRef]
- Gonzalez, R.; Murarka, A.; Dharmadi, Y.; Yazdani, S.S. A new model for the anaerobic fermentation of glycerol in enteric bacteria: Trunk and auxiliary pathways in Escherichia coli. Metab. Eng. 2008, 10, 234–245. [Google Scholar] [CrossRef]
- Okano, H.; Hermsen, R.; Kochanowski, K.; Hwa, T. Regulation underlying hierarchical and simultaneous utilization of carbon substrates by flux sensors in Escherichia coli. Nat. Microbiol. 2020, 5, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Xiong, D.; Hu, H.; Wakayama, M.; Yu, W.; Zhang, X.; Shimizu, K. Elucidation of the co-metabolism of glycerol and glucose in Escherichia coli by genetic engineering, transcription profiling, and (13)C metabolic flux analysis. Biotechnol. Biofuels 2016, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Jaén, K.E.; Velazquez, D.; Delvigne, F.; Sigala, J.C.; Lara, A.R. Engineering E. coli for improved microaerobic pDNA production. Bioprocess Biosyst. Eng. 2019, 42, 1457–1466. [Google Scholar] [CrossRef] [PubMed]
- Velazquez, D.; Sigala, J.C.; Martínez, L.M.; Gaytán, P.; Gosset, G.; Lara, A.R. Glucose transport engineering allows mimicking fed-batch performance in batch mode and selection of superior producer strains. Microb. Cell Fact. 2022, 21, 183. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.M.; Pedro, A.Q.; Oppolzer, D.; Sousa, F.; Queiroz, J.A.; Passarinha, L.A. Enhanced biosynthesis of plasmid DNA from Escherichia coli VH33 using Box–Behnken design associated to aromatic amino acids pathway. Biochem. Eng. J. 2015, 98, 117–126. [Google Scholar] [CrossRef]
- Ow, D.S.W.; Nissom, P.M.; Philp, R.; Oh, S.K.W.; Yap, M.G.S. Global transcriptional analysis of metabolic burden due to plasmid maintenance in Escherichia coli DH5α during batch fermentation. Enzym. Microb. Technol. 2006, 39, 391–398. [Google Scholar] [CrossRef]
- Kloska, S.M.; Pałczyński, K.; Marciniak, T.; Talaśka, T.; Miller, M.; Wysocki, B.J.; Davis, P.; Wysocki, T.A. Queueing theory model of pentose phosphate pathway. Sci. Rep. 2022, 12, 4601. [Google Scholar] [CrossRef]
- Bertels, L.K.; Fernández Murillo, L.; Heinisch, J.J. The Pentose Phosphate Pathway in Yeasts-More Than a Poor Cousin of Glycolysis. Biomolecules 2021, 11, 725. [Google Scholar] [CrossRef]
- Lin, P.C.; Saha, R.; Zhang, F.; Pakrasi, H.B. Metabolic engineering of the pentose phosphate pathway for enhanced limonene production in the cyanobacterium Synechocystis sp. PCC 6803. Sci. Rep. 2017, 7, 17503. [Google Scholar] [CrossRef]
- Cunningham, D.S.; Koepsel, R.R.; Ataai, M.M.; Domach, M.M. Factors affecting plasmid production in Escherichia coli from a resource allocation standpoint. Microb. Cell Fact. 2009, 8, 27. [Google Scholar] [CrossRef]
- Meade, J.; Bartlow, P.; Trivedi, R.N.; Akhtar, P.; Ataai, M.M.; Khan, S.A.; Domach, M.M. Effect of plasmid replication deregulation via inc mutations on E. coli proteome & simple flux model analysis. Microb. Cell Fact. 2015, 14, 31. [Google Scholar] [PubMed]
- Reckinger, A.R.; Jeong, K.S.; Khodursky, A.B.; Hiasa, H. RecA can stimulate the relaxation activity of topoisomerase I: Molecular basis of topoisomerase-mediated genome-wide transcriptional responses in Escherichia coli. Nucleic Acids Res. 2007, 35, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.C.; Kowalczykowski, S.C. RecA: Regulation and Mechanism of a Molecular Search Engine. Trends Biochem. Sci. 2016, 41, 491–507. [Google Scholar] [CrossRef]
- Courcelle, J.; Donaldson, J.R.; Chow, K.H.; Courcelle, C.T. DNA damage-induced replication fork regression and processing in Escherichia coli. Science 2003, 299, 1064–1067. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, S.; Gubaev, A.; Klostermeier, D. Binding and Hydrolysis of a Single ATP Is Sufficient for N-Gate Closure and DNA Supercoiling by Gyrase. J. Mol. Biol. 2017, 429, 3717–3729. [Google Scholar] [CrossRef] [PubMed]
- Reece, R.J.; Maxwell, A. DNA gyrase: Structure and function. Crit. Rev. Biochem. Mol. Biol. 1991, 26, 335–375. [Google Scholar] [CrossRef]
- Fishel, R.A.; James, A.A.; Kolodner, R. recA-independent general genetic recombination of plasmids. Nature 1981, 294, 184–186. [Google Scholar] [CrossRef]
- Krishna Rao, D.V.; Ramu, C.T.; Rao, J.V.; Narasu, M.L.; Bhujanga Rao, A.K. Impact of dissolved oxygen concentration on some key parameters and production of rhG-CSF in batch fermentation. J. Ind. Microbiol. Biotechnol. 2008, 35, 991–1000. [Google Scholar] [CrossRef]
- Goyal, D.; Sahni, G.; Sahoo, D.K. Enhanced production of recombinant streptokinase in Escherichia coli using fed-batch culture. Bioresour. Technol. 2009, 100, 4468–4474. [Google Scholar] [CrossRef]
- Silva, F.; Queiroz, J.A.; Domingues, F.C. Evaluating metabolic stress and plasmid stability in plasmid DNA production by Escherichia coli. Biotechnol. Adv. 2012, 30, 691–708. [Google Scholar] [CrossRef]
- Crozat, E.; Fournes, F.; Cornet, F.; Hallet, B.; Rousseau, P. Resolution of Multimeric Forms of Circular Plasmids and Chromosomes. Microbiol. Spectr. 2014, 2, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Summers, D.K.; Beton, C.W.H.; Withers, H.L. Multicopy plasmid instability: The dimer catastrophe hypothesis. Mol. Microbiol. 1993, 8, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Saraswat, V.; Kim, D.Y.; Lee, J.; Park, Y.-H. Effect of specific production rate of recombinant protein on multimerization of plasmid vector and gene expression level. FEMS Microbiol. Lett. 1999, 179, 367–373. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C-Source | Strain | µ [h−1] | qglucose [g g−1 h−1] | qglycerol [g g−1 h−1] | qacetate [g g−1 h−1] | RQ [mol mol−1] |
|---|---|---|---|---|---|---|
| Glucose | W3110 | 0.53 ± 0.02 | 1.35 ± 0.44 | - | 0.17 ± 0.04 | 0.92 ± 0.05 |
| VH34 | 0.31 ± 0.01 | 0.55 ± 0.17 | - | 0.00 ± 0.00 | 1.11 ± 0.03 | |
| Glycerol | W3110 | 0.43 ± 0.02 | - | 2.06 ± 0.23 | 0.07 ± 0.00 | 0.79 ± 0.01 |
| VH34 | 0.27 ± 0.02 | - | 0.78 ± 0.12 | 0.00 ± 0.00 | 0.76 ± 0.12 | |
| Glucose + glycerol | W3110 | 0.46 ± 0.01 | 1.35 ± 0.17 | - | 0.26 ± 0.02 | 1.11 ± 0.02 |
| VH34 | 0.35 ± 0.01 | 0.48 ± 0.11 | 0.21 ± 0.08 | 0.04 ± 0.02 | 0.92 ± 0.06 |
| Strain | µ [h−1] | qglucose [g g−1 h−1] | qacetate [g g−1 h−1] | YpDNA/X [mg g−1] | qpDNA [mg g−1 h−1] | SCF [%] |
|---|---|---|---|---|---|---|
| W3110recA− | 0.59 ± 0.01 | 1.47 ± 0.12 | 0.30 ± 0.02 | 1.68 ± 0.08 | 0.99 ± 0.13 | 78 ± 6 |
| VH34 | 0.17 ± 0.00 | 0.50 ± 0.05 | 0.00 ± 0.00 | 1.92 ± 0.01 | 0.32 ± 0.00 | 4 ± 1 |
| VH36 | 0.17 ± 0.00 | 0.31 ± 0.03 | 0.01 ± 0.00 | 1.21 ± 0.04 | 0.20 ± 0.01 | 39 ± 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de la Cruz, M.; Kunert, F.; Taymaz-Nikerel, H.; Sigala, J.-C.; Gosset, G.; Büchs, J.; Lara, A.R. Increasing the Pentose Phosphate Pathway Flux to Improve Plasmid DNA Production in Engineered E. coli. Microorganisms 2024, 12, 150. https://doi.org/10.3390/microorganisms12010150
de la Cruz M, Kunert F, Taymaz-Nikerel H, Sigala J-C, Gosset G, Büchs J, Lara AR. Increasing the Pentose Phosphate Pathway Flux to Improve Plasmid DNA Production in Engineered E. coli. Microorganisms. 2024; 12(1):150. https://doi.org/10.3390/microorganisms12010150
Chicago/Turabian Stylede la Cruz, Mitzi, Flavio Kunert, Hilal Taymaz-Nikerel, Juan-Carlos Sigala, Guillermo Gosset, Jochen Büchs, and Alvaro R. Lara. 2024. "Increasing the Pentose Phosphate Pathway Flux to Improve Plasmid DNA Production in Engineered E. coli" Microorganisms 12, no. 1: 150. https://doi.org/10.3390/microorganisms12010150