

The SIX5 Protein in Fusarium oxysporum f. sp. cepae Acts as an Avirulence Effector toward Shallot (Allium cepa L. Aggregatum Group)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Fungal Strain
2.2. Pathogenicity Test toward Onion and Shallot Plants
2.3. RNA Extraction and Quantitative Reverse-Transcriptase Polymerase Chain Reaction (qRT-PCR)
2.4. Sequence Alignment and Prediction of Signal Peptide
2.5. Generation of Gene Knockout and Complementation Constructs
2.6. Protoplast Preparation
2.7. Fungal Transformation
2.8. Vegetative Growth Assays
2.9. Statistical Analysis
3. Results
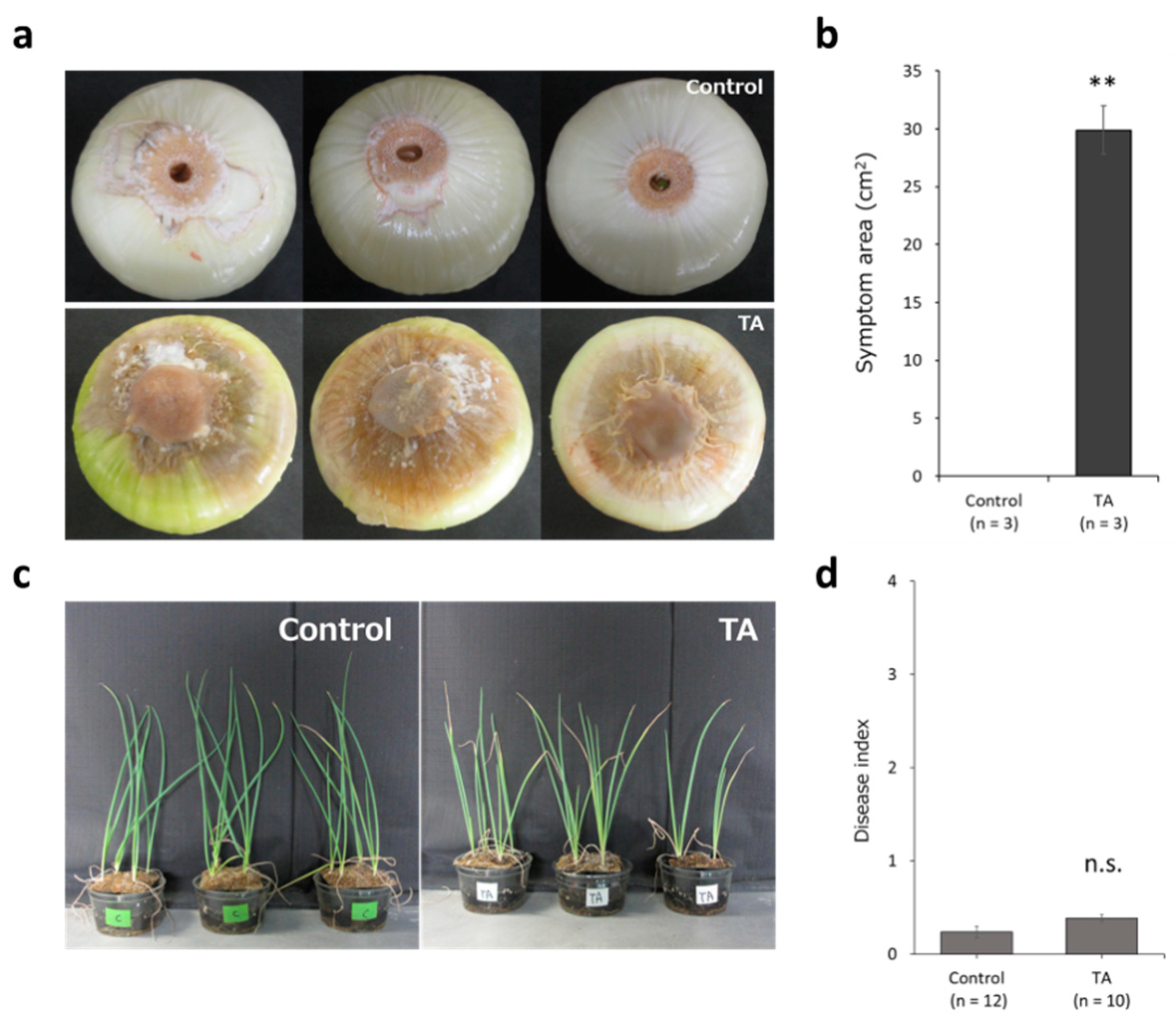
3.1. Confirmation of Pathogenicity of Foc_TA toward Onion and Shallot Plants
3.2. Expression of FocSIX5 Gene in Onion and Shallot Plants during Infection
3.3. Characterization of FocSIX5
3.4. Generation of a FocSIX5 Gene-Modified Mutant
3.5. Mycelial Growth of FocSIX5 Gene-Modified Mutant
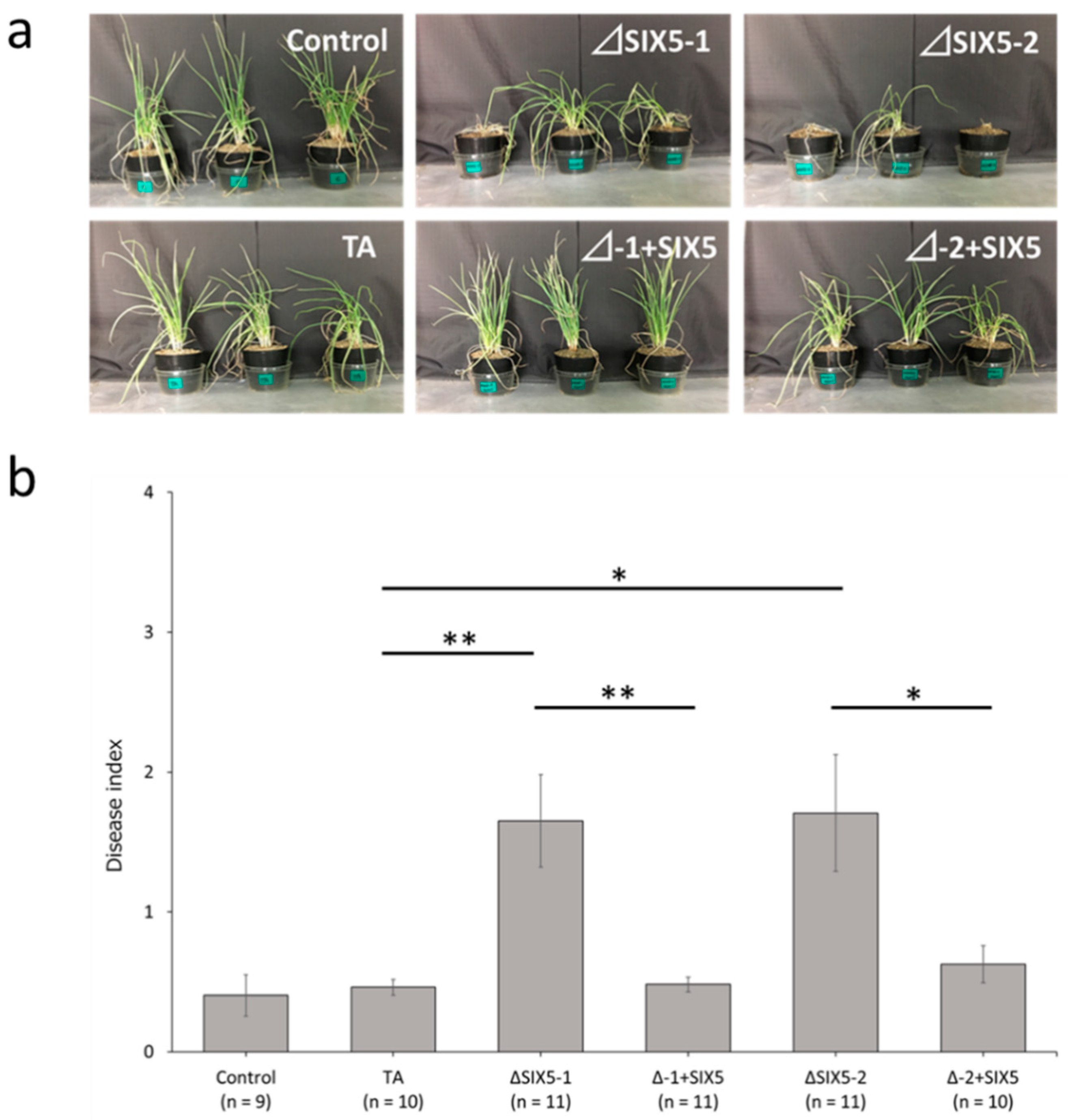
3.6. Effect of the FocSIX5 Gene Modification on Pathogenicity toward Onion and Shallot
3.7. Effect of G200A Mutation on the Pathogenicity toward Shallot
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. Fusarium laboratory workshops-A recent history. Mycotoxin Res. 2006, 22, 73–74. [Google Scholar] [CrossRef]
- Laurence, M.H.; Burgess, L.W.; Summerell, B.A.; Liew, E.C. High levels of diversity in Fusarium oxysporum from non-cultivated ecosystems in Australia. Fungal Biol. 2012, 116, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Michielse, C.B.; Rep, M. Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol. 2009, 10, 311–324. [Google Scholar] [CrossRef]
- Haapalainen, M.; Laitala, E.; Rämö, S.; Latvala, S. Pathogenicity and toxin production of different Fusarium oxysporum isolates infecting onion (Allium cepa L.). Ann. Appl. Biol. 2022, 180, 348–360. [Google Scholar] [CrossRef]
- Sasaki, K.; Nakahara, K.; Tanaka, S.; Shigyo, M.; Ito, S. Genetic and pathogenic variability of Fusarium oxysporum f. sp. cepae isolated from onion and Welsh onion in Japan. Phytopathology 2015, 105, 525–532. [Google Scholar] [CrossRef]
- Southwood, M.J.; Viljoen, A.; McLeod, A. Inoculum sources of Fusarium oxysporum f. sp. cepae on onion in the Western Cape Province of South Africa. Crop Prot. 2015, 75, 88–95. [Google Scholar] [CrossRef]
- Taylor, A.; Vágány, V.; Jackson, A.C.; Harrison, R.J.; Rainoni, A.; Clarkson, J.P. Identification of pathogenicity-related genes in Fusarium oxysporum f. sp. cepae. Mol. Plant Pathol. 2016, 17, 1032–1047. [Google Scholar] [CrossRef] [PubMed]
- Teshima, Y.; Ikeda, T.; Imada, K.; Sasaki, K.; El-Sayed, M.A.; Shigyo, M.; Tanaka, S.; Ito, S. Identification and biological activity of antifungal saponins from Shallot (Allium cepa L. aggregatum Group). J. Agric. Food Chem. 2013, 61, 7440–7445. [Google Scholar] [CrossRef]
- Yin, M.-C.; Hsu, P.-C.; Chang, H.-H. In vitro antioxidant and antibacterial activities of Shallot and Scallion. Food Sci. 2003, 68, 281–284. [Google Scholar] [CrossRef]
- Herlina, L.; Reflinur, R.; Sobir, S.; Maharijaya, A.; Wiyono, S. The genetic diversity and population structure of shallots (Allium cepa var. aggregatum) in Indonesia based on R gene-derived markers. Biodivers. J. Biol. Divers. 2019, 20, 696–703. [Google Scholar] [CrossRef]
- Vu, H.Q.; El-Sayed, M.A.; Ito, S.I.; Yamauchi, N.; Shigyo, M. Discovery of a new source of resistance to Fusarium oxysporum, cause of Fusarium wilt in Allium fistulosum, located on chromosome 2 of Allium cepa Aggregatum group. Genome 2012, 55, 797–807. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Sawada, Y.; Nakabayashi, R.; Sato, S.; Hirakawa, H.; El-Sayed, M.; Hirai, M.Y.; Saito, K.; Yamauchi, N.; Shigyo, M. Integrating transcriptome and target metabolome variability in doubled haploids of Allium cepa for abiotic stress protection. Mol. Breed. 2015, 35, 195. [Google Scholar] [CrossRef]
- Astley, D.; Innes, N.L.; Van der Meer, Q.P. Genetic Resources of Allium Species: A Global Report; IBPGR: Rome, Italy, 1982. [Google Scholar]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Rovenich, H.; Boshoven, J.C.; Thomma, B.P. Filamentous pathogen effector functions: Of pathogens, hosts and microbiomes. Curr. Opin. Plant Biol. 2014, 20, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Ngou, B.P.M.; Ding, P.; Xin, X.F. PTI-ETI crosstalk: An integrative view of plant immunity. Curr. Opin. Plant Biol. 2021, 62, 102030. [Google Scholar] [CrossRef] [PubMed]
- Houterman, P.M.; Speijer, D.; Dekker, H.L.; De Koster, C.G.; Cornelissen, B.J.C.; Rep, M. The mixed xylem sap proteome of Fusarium oxysporum-infected tomato plants. Mol. Plant Pathol. 2007, 8, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.M.; Houterman, P.M.; Schreiver, I.; Ma, L.; Amyotte, S.; Chellappan, B.; Boeren, S.; Takken, F.L.W.; Rep, M. MITEs in the promoters of effector genes allow prediction of novel virulence genes in Fusarium Oxysporum. BMC Genom. 2013, 14, 119. [Google Scholar] [CrossRef]
- Catanzariti, A.M.; Lim, G.T.T.; Jones, D.A. The tomato I-3 gene: A novel gene for resistance to Fusarium wilt disease. New Phytol. 2015, 207, 106–118. [Google Scholar] [CrossRef]
- Gawehns, F.; Houterman, P.M.; Ichou, F.A.; Michielse, C.B.; Hijdra, M.; Cornelissen, B.J.C.; Rep, M.; Takken, F.L. The Fusarium oxysporum effector Six6 contributes to virulence and suppresses I-2-mediated cell death. Mol. Plant Microbe Interact. 2014, 27, 336–348. [Google Scholar] [CrossRef]
- Houterman, P.M.; Ma, L.; van Ooijen, G.; de Vroomen, M.J.; Cornelissen, B.J.C.; Takken, F.L.W.; Rep, M. The effector protein Avr2 of the xylem-colonizing fungus Fusarium oxysporum activates the tomato resistance protein I-2 intracellularly. Plant J. 2009, 58, 970–978. [Google Scholar] [CrossRef]
- Ma, L.; Houterman, P.M.; Gawehns, F.; Cao, L.; Sillo, F.; Richter, H.; Clavijo-Ortiz, M.J.; Schmidt, S.M.; Boeren, S.; Vervoort, J.; et al. The AVR 2–SIX 5 gene pair is required to activate I-2-mediated immunity in tomato. New Phytol. 2015, 208, 507–518. [Google Scholar] [CrossRef]
- Rep, M.; Van Der Does, H.C.; Meijer, M.; Van Wijk, R.; Houterman, P.M.; Dekker, H.L.; De Koster, C.G.; Cornelissen, B.J.C. A small, cysteine-rich protein secreted by Fusarium oxysporum during colonization of xylem vessels is required for I-3-mediated resistance in tomato. Mol. Microbiol. 2004, 53, 1373–1383. [Google Scholar] [CrossRef]
- Takken, F.; Rep, M. The arms race between tomato and Fusarium Oxysporum. Mol. Plant Pathol. 2010, 11, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Huang, C.; Lindhout, P. Screening for resistance in wild Lycopersicon species to Fusarium oxysporum f. sp. lycopersici race 1 and race 2. Euphytica 1997, 93, 145–153. [Google Scholar] [CrossRef]
- Lievens, B.; Houterman, P.M.; Rep, M. Effector gene screening allows unambiguous identification of Fusarium oxysporum f. sp. lycopersici races and discrimination from other formae speciales. FEMS Microbiol. Lett. 2009, 300, 201–215. [Google Scholar] [CrossRef]
- Armitage, A.D.; Taylor, A.; Sobczyk, M.K.; Baxter, L.; Greenfield, B.P.J.; Bates, H.J.; Wilson, F.; Jackson, A.C.; Ott, S.; Harrison, R.J.; et al. Characterisation of pathogen-specific regions and novel effector candidates in Fusarium oxysporum f. sp. cepae. Sci. Rep. 2018, 8, 13530. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Herlina, L.; Istiaji, B.; Wiyono, S. The causal agent of Fusarium disease infested Shallots in Java islands of Indonesia. E3S Web Conf. 2021, 232, 03003. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. Clustal W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H.; Signal, P. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Kuwayama, H.; Obara, S.; Morio, T.; Katoh, M.; Urushihara, H.; Tanaka, Y. PCR-mediated generation of a gene disruption construct without the use of DNA ligase and plasmid vectors. Nucleic Acids Res. 2002, 30, E2. [Google Scholar] [CrossRef] [PubMed]
- van der Does, H.C.; Duyvesteijn, R.G.; Goltstein, P.M.; van Schie, C.C.; Manders, E.M.; Cornelissen, B.J.; Rep, M. Expression of effector gene SIX1 of Fusarium oxysporum requires living plant cells. Fungal Genet. Biol. 2008, 45, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Catlett, N.L.; Lee, B.-N.; Yoder, O.C.; Turgeon, B.G. Split-marker recombination for efficient targeted deletion of fungal genes. Fungal Genet. Rep. 2003, 50, 9–11. [Google Scholar] [CrossRef]
- Namiki, F.; Matsunaga, M.; Okuda, M.; Inoue, I.; Nishi, K.; Fujita, Y.; Tsuge, T. Mutation of an arginine biosynthesis gene causes reduced pathogenicity in Fusarium oxysporum f. sp. melonis. Mol. Plant Microbe Interact. 2001, 14, 580–584. [Google Scholar] [CrossRef]
- Akamatsu, H.; Taga, M.; Kodama, M.; Johnson, R.; Otani, H.; Kohmoto, K. Molecular karyotypes for Alternaria plant pathogens known to produce host-specific toxins. Curr. Genet. 1999, 35, 647–656. [Google Scholar] [CrossRef]
- Kawabe, M.; Mizutani, K.; Yoshida, T.; Teraoka, T.; Yoneyama, K.; Yamaguchi, I.; Arie, T. Cloning of the pathogenicity-related gene FPD1 in Fusarium oxysporum f. sp. lycopersici. J. Gen. Plant Pathol. 2004, 70, 16–20. [Google Scholar] [CrossRef]
- Saitoh, K.I.; Togashi, K.; Arie, T.; Teraoka, T. A simple method for a mini-preparation of fungal DNA. J. Gen. Plant Pathol. 2006, 72, 348–350. [Google Scholar] [CrossRef]
- Hammond-Kosack, K.E.; Jones, J.D.G. Plant disease resistance genes. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 575–607. [Google Scholar] [CrossRef]
- Martin, G.B.; Bogdanove, A.J.; Sessa, G. Understanding the functions of plant disease resistance proteins. Annu. Rev. Plant Biol. 2003, 54, 23–61. [Google Scholar] [CrossRef]
- Niu, X.; Zhao, X.; Ling, K.S.; Levi, A.; Sun, Y.; Fan, M. The FonSIX6 gene acts as an avirulence effector in the Fusarium oxysporum f. sp. niveum—Watermelon pathosystem. Sci. Rep. 2016, 6, 28146. [Google Scholar] [CrossRef]
- Rouxel, T.; Balesdent, M.H. Avirulence genes. In Encyclopedia of Life Sciences (ELS); John Wiley & Sons, Ltd.: Chichester, UK, 2010. [Google Scholar] [CrossRef]
- Stephens, C.; Ölmez, F.; Blyth, H.; McDonald, M.; Bansal, A.; Turgay, E.B.; Hahn, F.; Saintenac, C.; Nekrasov, V.; Solomon, P.; et al. Remarkable recent changes in the genetic diversity of the avirulence gene AvrStb6 in global populations of the wheat pathogen Zymoseptoria tritici. Mol. Plant Pathol. 2021, 22, 1121–1133. [Google Scholar] [CrossRef]
- Sasaki, K.; Nakahara, K.; Shigyo, M.; Tanaka, S.; Ito, S. Detection and quantification of onion isolates of Fusarium oxysporum f. sp. cepae in onion plant. J. Gen. Plant Pathol. 2015, 81, 232–236. [Google Scholar] [CrossRef]
- Ellis, J.G.; Dodds, P.N.; Lawrence, G.J. Flax rust resistance gene specificity is based on direct resistance-avirulence protein interactions. Annu. Rev. Phytopathol. 2007, 45, 289–306. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; McAdams, S.A.; Bryan, G.T.; Hershey, H.P.; Valent, B. Direct interaction of resistance gene and avirulence gene products confers rice blast resistance. EMBO J. 2000, 19, 4004–4014. [Google Scholar] [CrossRef] [PubMed]
- Sintayehu, A.; Fininsa, C.; Ahmed, S.; Sakhuja, P.K. Evaluations of shallot genotypes for resistance against fusarium basal rot (Fusarium oxysporum f. sp. cepae) disease. Crop Prot. 2011, 30, 1210–1215. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions, and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of the MDPI and/or editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′–3′) | Purpose | Reference |
|---|---|---|---|
| SIX5-C-F | GCGCTTCGAGTACATCTCTG | Detection of FocSIX5 | This study |
| SIX5-C-R | CTAGGATGCATCACAATAGA | Detection of FocSIX5 | This study |
| SIX5-Q-F | TGCCACCACTCAGCTTCAGA | Quantification of FocSIX5 | This study |
| SIX5-Q-R | TGAAATGTGGACCAAGTGCTCTA | Quantification of FocSIX5 | This study |
| SIX5-split-F1 | GGGATAGGTAAGCAAGCAGCTTG | Disruption and complementation of FocSIX5 | This study |
| SIX5-split-F2 | GTCGTGACTGGGAAAACCCTGG CGGTGATGAAGAGTAGTAGAG | Disruption of FocSIX5 | This study |
| SIX5-split-F3 | TCCTGTGTGAAATTGTTATCCG CTTCTGTCATTGTGACCAGTG | Disruption of FocSIX5 Verification of FocSIX5 gene knockout | This study |
| SIX5-split-F4 | ATGTCAAGAGCGCGCGAAGCTC | Disruption and complementation of FocSIX5 Verification of FocSIX5 gene knockout | This study |
| FoTEF-Q2-F | CATCGGCCACGTCGACTCT | Quantification of EF-1α | [34] |
| FoTEF-Q2-R | AGAACCCAGGCGTACTTGAA | Quantification of EF-1α | [34] |
| M13F | CGCCAGGGTTTTCCCAGTCACGAC | Creation of hph construct | [35] |
| M13R | AGCGGATAACAATTCACACAGGA | Creation of hph construct | [35] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakane, K.; Kunimoto, M.; Furumoto, K.; Shigyo, M.; Sasaki, K.; Ito, S.-i. The SIX5 Protein in Fusarium oxysporum f. sp. cepae Acts as an Avirulence Effector toward Shallot (Allium cepa L. Aggregatum Group). Microorganisms 2023, 11, 2861. https://doi.org/10.3390/microorganisms11122861
Sakane K, Kunimoto M, Furumoto K, Shigyo M, Sasaki K, Ito S-i. The SIX5 Protein in Fusarium oxysporum f. sp. cepae Acts as an Avirulence Effector toward Shallot (Allium cepa L. Aggregatum Group). Microorganisms. 2023; 11(12):2861. https://doi.org/10.3390/microorganisms11122861
Chicago/Turabian StyleSakane, Kosei, Masaaki Kunimoto, Kazuki Furumoto, Masayoshi Shigyo, Kazunori Sasaki, and Shin-ichi Ito. 2023. "The SIX5 Protein in Fusarium oxysporum f. sp. cepae Acts as an Avirulence Effector toward Shallot (Allium cepa L. Aggregatum Group)" Microorganisms 11, no. 12: 2861. https://doi.org/10.3390/microorganisms11122861