

16S and 18S rDNA Amplicon Sequencing Analysis of Aesthetically Problematic Microbial Mats on the Walls of the Petralona Cave: The Use of Essential Oils as a Cleaning Method

 and
and
Abstract
:1. Introduction
2. Materials and Methods
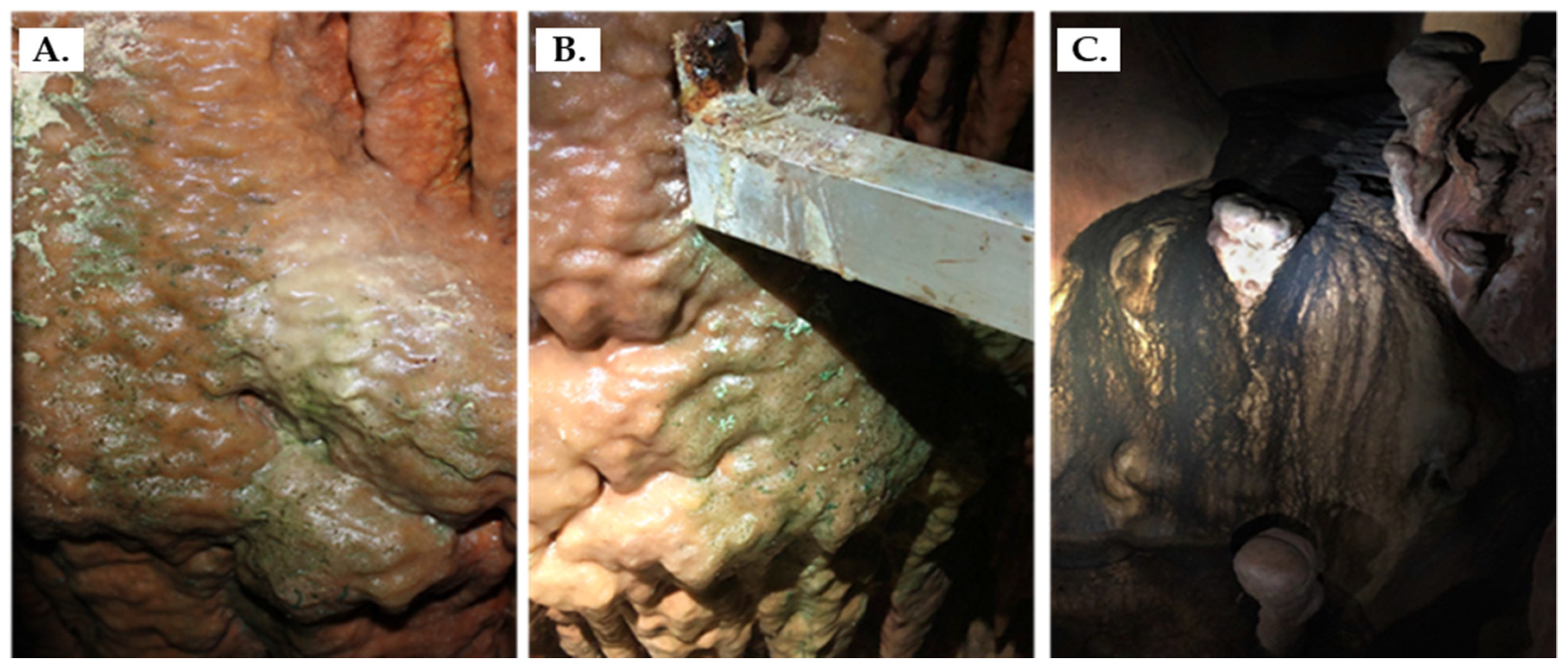
2.1. Sampling
2.2. DNA Extraction
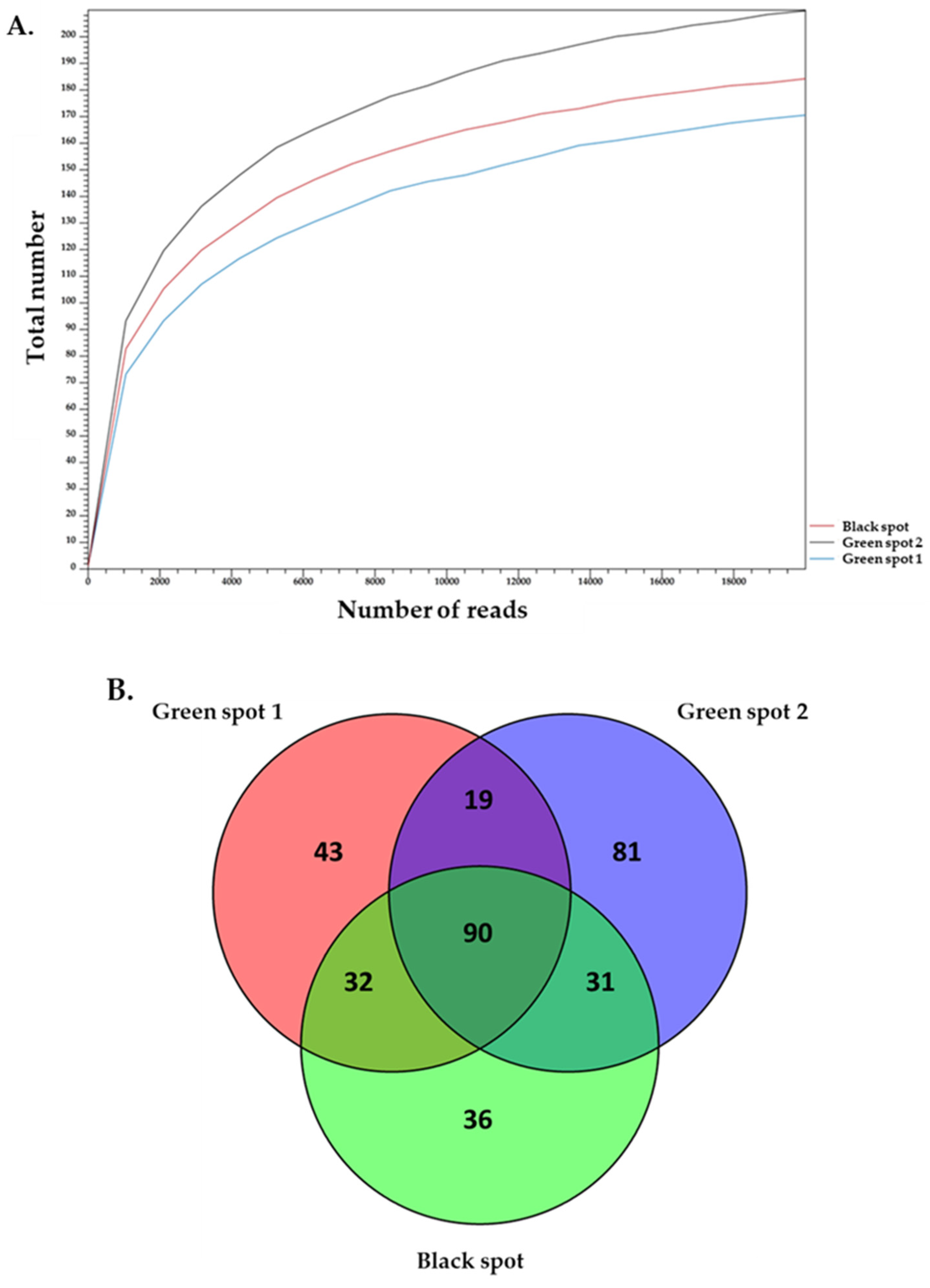
2.3. Sequencing and Bioinformatics Analysis—16S and 18S rDNA Analysis
2.4. Application of the Essential Oil Emulsions
2.5. Statistical Analysis
3. Results and Discussion
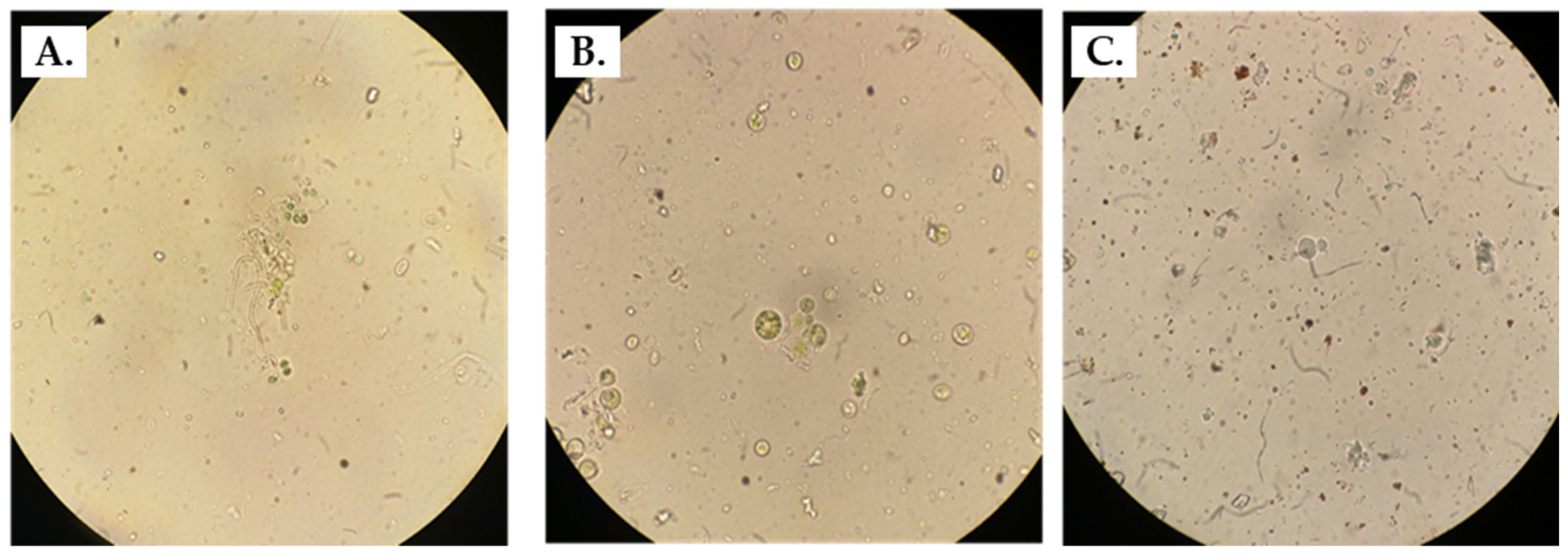
3.1. Observation through Microscopy
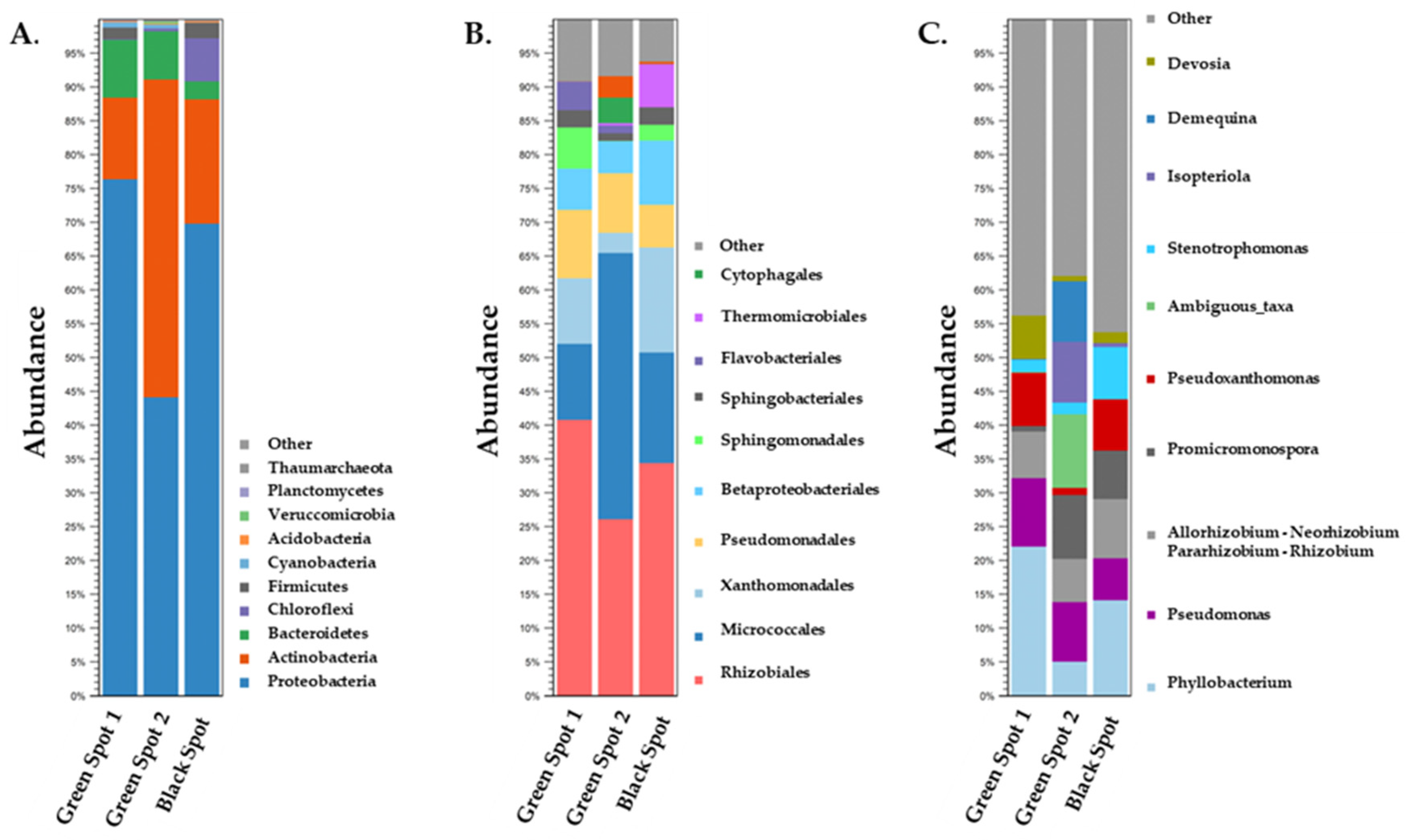
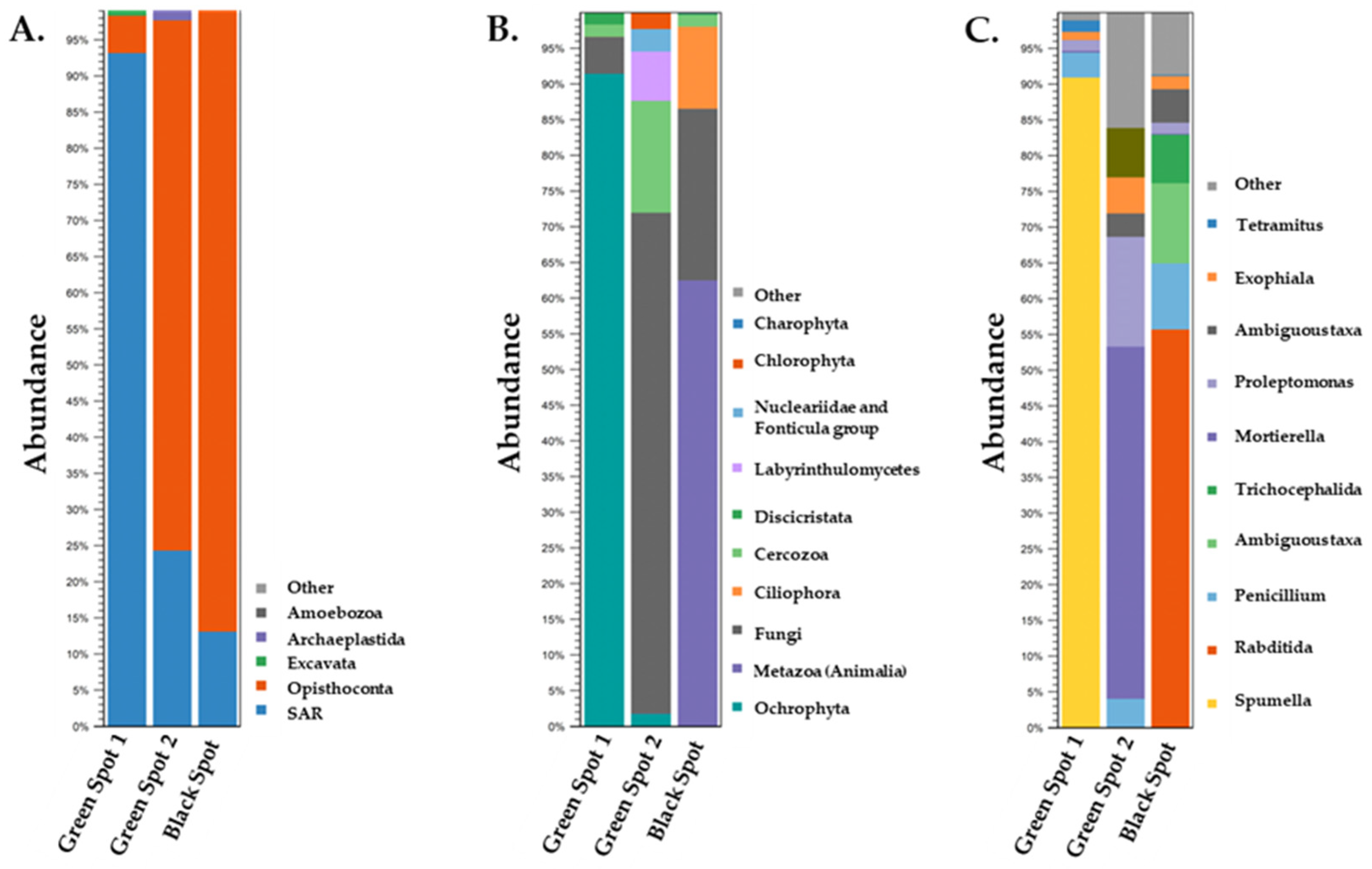
3.2. Description of the Biodiversity of Prokaryotic Microorganisms in the Petralona Cave Biofilms
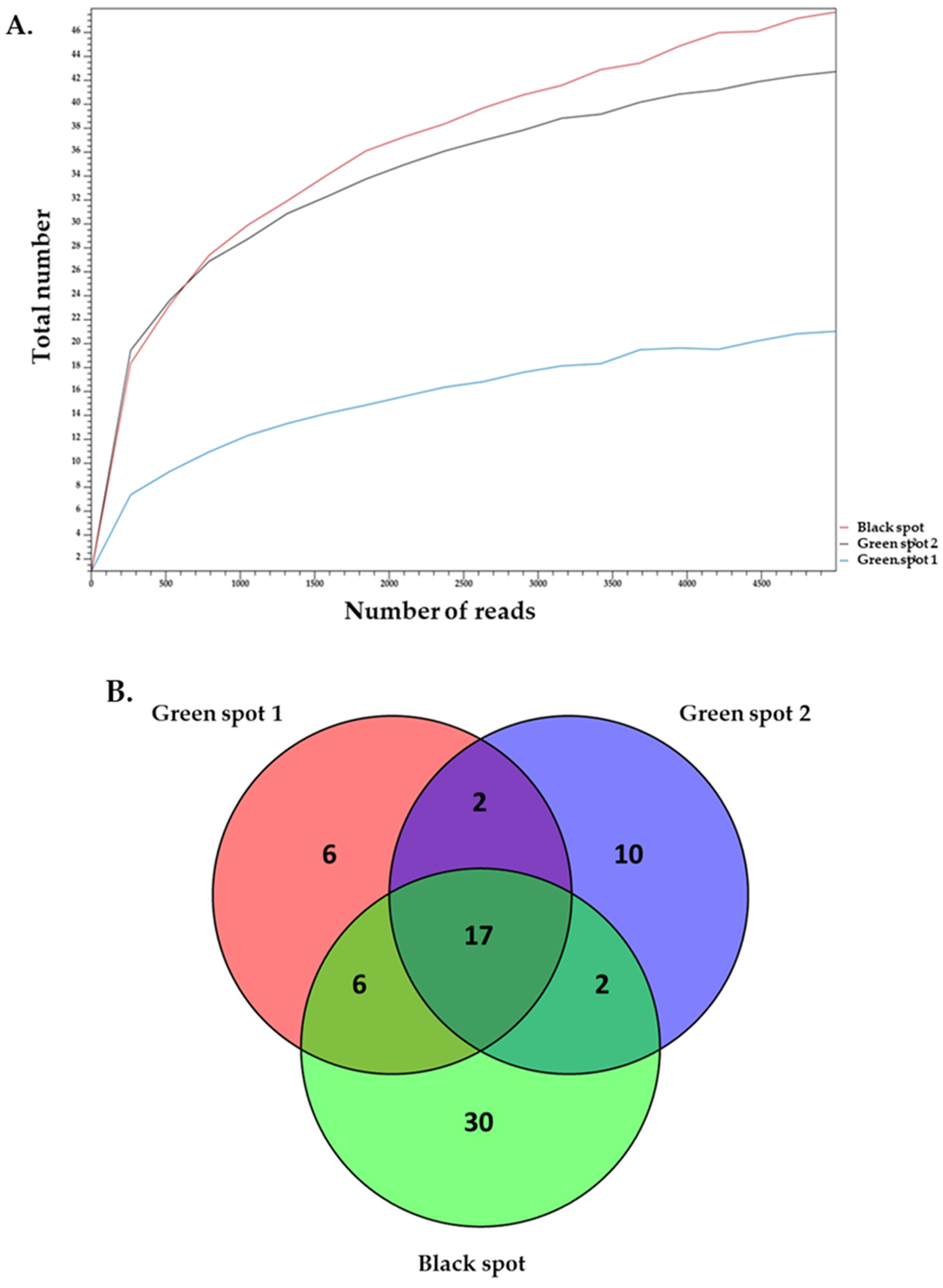
3.3. Description of the Biodiversity of Eukaryotic Microorganisms in the Petralona Cave Biofilms
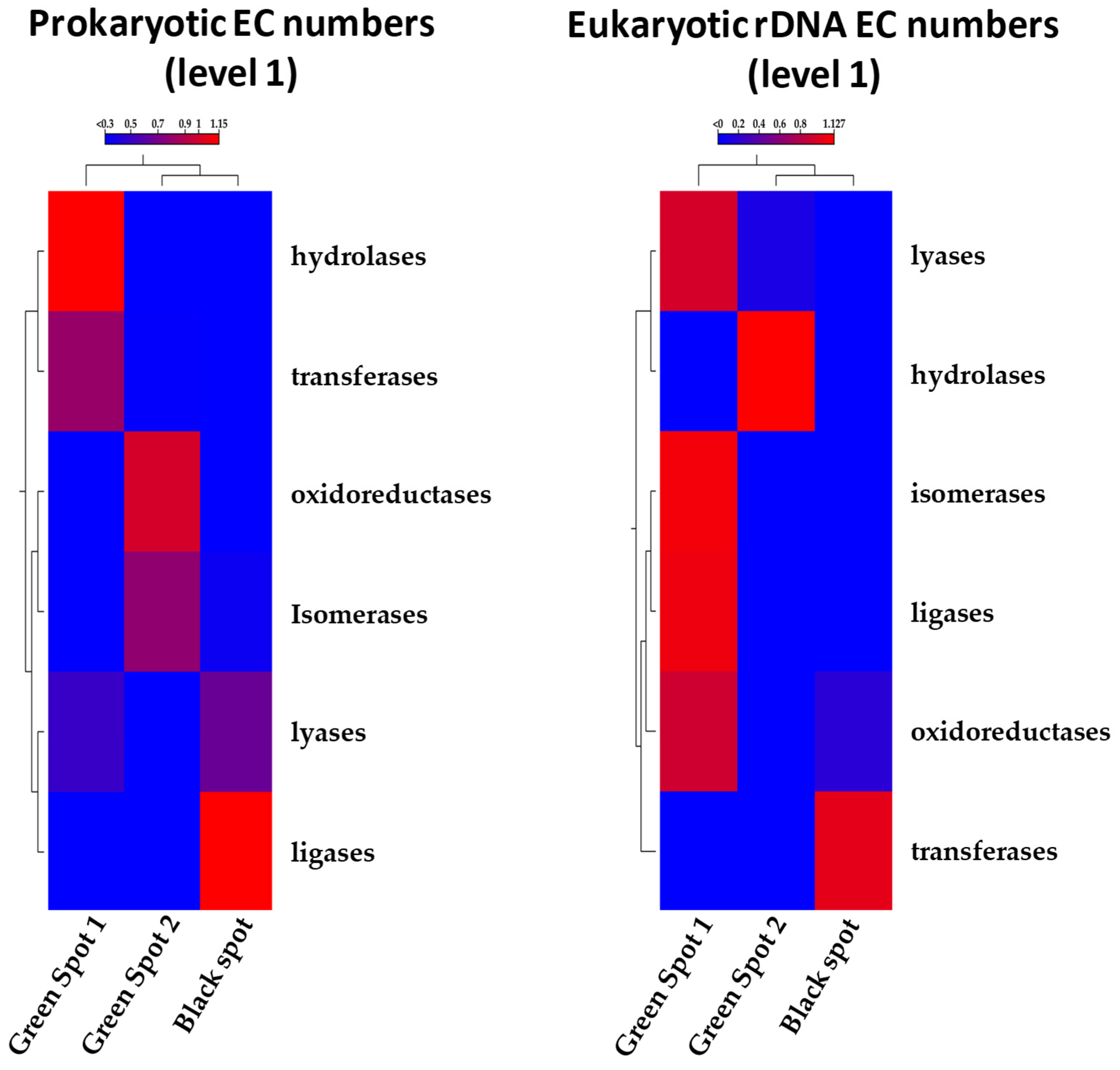
3.4. Functional Analysis Using PICRUST2
3.5. Antimicrobial Action of Essential Oils against the Petralona Cave Biofilms
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mulec, J.; Kosi, G. Lampenflora algae and methods of growth control. J. Cave Karst Stud. 2009, 71, 109–115. [Google Scholar]
- Pfendler, S.; Karimi, B.; Maron, P.A.; Ciadamidaro, L.; Valot, B.; Bousta, F.; Alaoui-Sosse, L.; Alaoui-Sosse, B.; Aleya, L. Biofilm biodiversity in French and Swiss show caves using the metabarcoding approach: First data. Sci. Total Environ. 2018, 615, 1207–1217. [Google Scholar] [CrossRef] [PubMed]
- Kurniawan, I.D.; Rahmadi, C.; Ardi, T.E.; Nasrullah, R.; Willyanto, M.I.; Setiabudi, A. The Impact of Lampenflora on Cave-dwelling Arthropods in Gunungsewu Karst, Java, Indonesia. Biosaintifika J. Biol. Biol. Educ. 2018, 10, 275–283. [Google Scholar] [CrossRef]
- Borderie, F.; Denis, M.; Barani, A.; Alaoui-Sosse, B.; Aleya, L. Microbial composition and ecological features of phototrophic biofilms proliferating in the Moidons Caves (France): Investigation at the single-cell level. Environ. Sci. Pollut. Res. Int. 2016, 23, 12039–12049. [Google Scholar] [CrossRef] [PubMed]
- Piano, E.; Bona, F.; Falasco, E.; La Morgia, V.; Badino, G.; Isaia, M. Environmental drivers of phototrophic biofilms in an Alpine show cave (SW-Italian Alps). Sci. Total Environ. 2015, 536, 1007–1018. [Google Scholar] [CrossRef]
- Cigna, A. The problem of lampenflora in show caves. In Proceedings of the 6th ISCA Congress, Liptovský Mikuláš, Slovakia, 18–23 October 2010; pp. 201–205. [Google Scholar]
- Lamprinou, V.; Danielidis, D.; Pantazidou, A.; Oikonomou, A.; Economou-Amilli, A. The show cave of Diros vs. wild caves of Peloponnese, Greece-distribution patterns of Cyanobacteria. Int. J. Speleol. 2014, 43, 335–342. [Google Scholar] [CrossRef]
- Mulec, J. Lampenflora. In Encyclopedia of Caves; Academic Press: Cambridge, MA, USA, 2019; pp. 635–641. [Google Scholar]
- Borderie, F.; Tete, N.; Cailhol, D.; Alaoui-Sehmer, L.; Bousta, F.; Rieffel, D.; Aleya, L.; Alaoui-Sosse, B. Factors driving epilithic algal colonization in show caves and new insights into combating biofilm development with UV-C treatments. Sci. Total Environ. 2014, 484, 43–52. [Google Scholar] [CrossRef]
- Nikolic, N.; Subakov-Simic, G.; Golic, I.; Popovic, S. The effects of biocides on the growth of aerophytic green algae (Chlorella sp.) isolated from a cave environment. Arch. Biol. Sci. 2021, 73, 341–351. [Google Scholar] [CrossRef]
- Meyer, E.; Seale, L.D.; Permar, B.; McClary, A. The effect of chemical treatments on Lampenflora and a Collembola indicator species at a popular tour cave in California, USA. Environ. Manag. 2017, 59, 1034–1042. [Google Scholar] [CrossRef]
- O’Bryan, C.A.; Pendleton, S.J.; Crandall, P.G.; Ricke, S.C. Potential of plant essential oils and their components in animal agriculture—In vitro studies on antibacterial mode of action. Front. Vet. Sci. 2015, 2, 35. [Google Scholar] [CrossRef]
- Argyri, A.A.; Doulgeraki, A.I.; Varla, E.G.; Bikouli, V.C.; Natskoulis, P.I.; Haroutounian, S.A.; Moulas, G.A.; Tassou, C.C.; Chorianopoulos, N.G. Evaluation of plant origin essential oils as herbal biocides for the protection of caves belonging to natural and cultural heritage sites. Microorganisms 2021, 9, 1836. [Google Scholar] [CrossRef] [PubMed]
- Papastefanou, C.; Manolopoulou, M.; Stoulos, S.; Ioannidou, A.; Gerasopoulos, E. Elevated radon concentrations in a Pleistocenic cave operating as a show cave. Int. Congr. Ser. 2005, 1276, 204–205. [Google Scholar] [CrossRef]
- Tzimtzimis, E.; Tzetzis, D.; Achillas, C.; Poulios, S.; Tzioumakis, P.; Darlas, A.; Athanasiou, A.; Aidonis, D.; Bochtis, D. Understanding Speleology and Paleoanthropology through Digital and Interactive Technologies. In Proceedings of the HCI International 2022 Posters, Cham, Switzerland, 26 June–1 July 2022; pp. 288–295. [Google Scholar]
- Papadimitriou, K.; Anastasiou, R.; Georgalaki, M.; Bounenni, R.; Paximadaki, A.; Charmpi, C.; Alexandraki, V.; Kazou, M.; Tsakalidou, E. Comparison of the Microbiome of Artisanal Homemade and Industrial Feta Cheese through Amplicon Sequencing and Shotgun Metagenomics. Microorganisms 2022, 10, 1073. [Google Scholar] [CrossRef] [PubMed]
- De Roos, J.; Vandamme, P.; De Vuyst, L. Wort Substrate Consumption and Metabolite Production during Lambic Beer Fermentation and Maturation Explain the Successive Growth of Specific Bacterial and Yeast Species. Front. Microbiol. 2018, 9, 2763. [Google Scholar] [CrossRef] [PubMed]
- Verce, M.; De Vuyst, L.; Weckx, S. Shotgun Metagenomics of a Water Kefir Fermentation Ecosystem Reveals a Novel Oenococcus Species. Front. Microbiol. 2019, 10, 479. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Chelius, M.; Moore, J.; Chelius, M.K.; Moore, J.C. Molecular phylogenetic analysis of Archaea and bacteria in Wind Cave, South Dakota. Geomicrobiol. J. 2004, 21, 123–134. [Google Scholar] [CrossRef]
- Macalady, J.L.; Lyon, E.H.; Koffman, B.; Albertson, L.K.; Meyer, K.; Galdenzi, S.; Mariani, S. Dominant microbial populations in limestone-corroding stream biofilms, Frasassi cave system, Italy. Appl. Environ. Microbiol. 2006, 72, 5596–5609. [Google Scholar] [CrossRef]
- Turrini, P.; Tescari, M.; Visaggio, D.; Pirolo, M.; Lugli, G.A.; Ventura, M.; Frangipani, E.; Visca, P. The microbial community of a biofilm lining the wall of a pristine cave in Western New Guinea. Microbiol. Res. 2020, 241, 126584. [Google Scholar] [CrossRef]
- Pasic, L.; Kovce, B.; Sket, B.; Herzog-Velikonja, B. Diversity of microbial communities colonizing the walls of a Karstic cave in Slovenia. FEMS Microbiol. Ecol. 2010, 71, 50–60. [Google Scholar] [CrossRef]
- Burgoyne, J.; Crepeau, R.; Jensen, J.; Smith, H.; Baker, G.; Leavitt, S.D. Lampenflora in a show cave in the Great Basin is distinct from communities on naturally lit rock surfaces in nearby wild Caves. Microorganisms 2021, 9, 1188. [Google Scholar] [CrossRef]
- Gonzalez-Pimentel, J.L.; Martin-Pozas, T.; Jurado, V.; Miller, A.Z.; Caldeira, A.T.; Fernandez-Lorenzo, O.; Sanchez-Moral, S.; Saiz-Jimenez, C. Prokaryotic communities from a lava tube cave in La Palma Island (Spain) are involved in the biogeochemical cycle of major elements. PeerJ 2021, 9, e11386. [Google Scholar] [CrossRef] [PubMed]
- Portillo, M.C.; Gonzalez, J.M.; Saiz-Jimenez, C. Metabolically active microbial communities of yellow and grey colonizations on the walls of Altamira Cave, Spain. J. Appl. Microbiol. 2008, 104, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, C.; Marshall Hathaway, J.J.; Enes Dapkevicius Mde, L.; Miller, A.Z.; Kooser, A.; Northup, D.E.; Jurado, V.; Fernandez, O.; Saiz-Jimenez, C.; Cheeptham, N. Actinobacterial diversity in volcanic caves and associated geomicrobiological interactions. Front. Microbiol. 2015, 6, 1342. [Google Scholar] [CrossRef] [PubMed]
- Fang, B.Z.; Salam, N.; Han, M.X.; Jiao, J.Y.; Cheng, J.; Wei, D.Q.; Xiao, M.; Li, W.J. Insights on the effects of heat pretreatment, pH, and calcium salts on isolation of rare Actinobacteria from karstic caves. Front. Microbiol. 2017, 8, 1535. [Google Scholar] [CrossRef]
- Ghezzi, D.; Sauro, F.; Columbu, A.; Carbone, C.; Hong, P.Y.; Vergara, F.; De Waele, J.; Cappelletti, M. Transition from unclassified Ktedonobacterales to Actinobacteria during amorphous silica precipitation in a quartzite cave environment. Sci. Rep. 2021, 11, 3921. [Google Scholar] [CrossRef]
- Alonso, L.; Creuze-des-Chatelliers, C.; Trabac, T.; Dubost, A.; Moenne-Loccoz, Y.; Pommier, T. Rock substrate rather than black stain alterations drives microbial community structure in the passage of Lascaux Cave. Microbiome 2018, 6, 216. [Google Scholar] [CrossRef]
- McKee, L.S.; La Rosa, S.L.; Westereng, B.; Eijsink, V.G.; Pope, P.B.; Larsbrink, J. Polysaccharide degradation by the Bacteroidetes: Mechanisms and nomenclature. Environ. Microbiol. Rep. 2021, 13, 559–581. [Google Scholar] [CrossRef]
- Jagannathan, B.; Golbeck, J.H. Photosynthesis: Microbial. In Encyclopedia of Microbiology, 3rd ed.; Schaechter, M., Ed.; Academic Press: Oxford, UK, 2009; pp. 325–341. [Google Scholar]
- Barton, H.A.; Giarrizzo, J.G.; Suarez, P.; Robertson, C.E.; Broering, M.J.; Banks, E.D.; Vaishampayan, P.A.; Venkateswaran, K. Microbial diversity in a Venezuelan orthoquartzite cave is dominated by the Chloroflexi (Class Ktedonobacterales) and Thaumarchaeota Group I.1c. Front. Microbiol. 2014, 5, 615. [Google Scholar] [CrossRef]
- Wieczorek, A.S.; Schmidt, O.; Chatzinotas, A.; von Bergen, M.; Gorissen, A.; Kolb, S. Ecological functions of agricultural soil Bacteria and Microeukaryotes in chitin degradation: A case study. Front. Microbiol. 2019, 10, 1293. [Google Scholar] [CrossRef]
- Dong, Y.; Gao, J.; Wu, Q.; Ai, Y.; Huang, Y.; Wei, W.; Sun, S.; Weng, Q. Co-occurrence pattern and function prediction of bacterial community in Karst cave. BMC Microbiol. 2020, 20, 137. [Google Scholar] [CrossRef] [PubMed]
- Schulz, H.N.; Brinkhoff, T.; Ferdelman, T.G.; Mariné, M.H.; Teske, A.; Jorgensen, B.B. Dense populations of a giant sulfur bacterium in Namibian shelf sediments. Science 1999, 284, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.P.; Gillespie, J.J.; Sobral, B.W.; Nordberg, E.K.; Snyder, E.E.; Shallom, J.M.; Dickerman, A.W. Phylogeny of Gammaproteobacteria. J. Bacteriol. 2010, 192, 2305–2314. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Paine, E.; Wall, R.; Kam, G.; Lauriente, T.; Sa-ngarmangkang, P.-C.; Horne, D.; Cheeptham, N. In situ cultured bacterial diversity from Iron Curtain Cave, Chilliwack, British Columbia, Canada. Diversity 2017, 9, 36. [Google Scholar] [CrossRef]
- Duncan, T.R.; Werner-Washburne, M.; Northup, D.E. Diversity of siderophore-producing bacterial cultures from Carlsbad caverns National Park (Ccnp) Caves, Carlsbad, New Mexico. J. Cave Karst Stud. 2021, 83, 29–43. [Google Scholar] [CrossRef] [PubMed]
- De Mandal, S.; Chatterjee, R.; Kumar, N.S. Dominant bacterial phyla in caves and their predicted functional roles in C and N cycle. BMC Microbiol. 2017, 17, 90. [Google Scholar] [CrossRef]
- D’Auria, G.; Artacho, A.; Rojas, R.A.; Bautista, J.S.; Mendez, R.; Gamboa, M.T.; Gamboa, J.R.; Gomez-Cruz, R. Metagenomics of bacterial diversity in Villa Luz Caves with sulfur water springs. Genes 2018, 9, 55. [Google Scholar] [CrossRef]
- Addesso, R.; Gonzalez-Pimentel, J.L.; D’Angeli, I.M.; De Waele, J.; Saiz-Jimenez, C.; Jurado, V.; Miller, A.Z.; Cubero, B.; Vigliotta, G.; Baldantoni, D. Microbial community characterizing vermiculations from Karst Caves and its role in their formation. Microb. Ecol. 2021, 81, 884–896. [Google Scholar] [CrossRef]
- Diaz-Herraiz, M.; Jurado, V.; Cuezva, S.; Laiz, L.; Pallecchi, P.; Tiano, P.; Sanchez-Moral, S.; Saiz-Jimenez, C. Deterioration of an Etruscan tomb by bacteria from the order Rhizobiales. Sci. Rep. 2014, 4, 3610. [Google Scholar] [CrossRef]
- Bogdan, D.F.; Baricz, A.I.; Chiciudean, I.; Bulzu, P.A.; Cristea, A.; Nastase-Bucur, R.; Levei, E.A.; Cadar, O.; Sitar, C.; Banciu, H.L.; et al. Diversity, distribution and organic substrates preferences of microbial communities of a low anthropic activity cave in North-Western Romania. Front. Microbiol. 2023, 14, 962452. [Google Scholar] [CrossRef]
- Mori, J.F.; Chen, L.X.; Jessen, G.L.; Rudderham, S.B.; McBeth, J.M.; Lindsay, M.B.J.; Slater, G.F.; Banfield, J.F.; Warren, L.A. Putative mixotrophic nitrifying-denitrifying Gammaproteobacteria implicated in nitrogen cycling within the ammonia/oxygen transition zone of an oil sands Pit lake. Front. Microbiol. 2019, 10, 2435. [Google Scholar] [CrossRef] [PubMed]
- Taubert, M.; Heinze, B.M.; Overholt, W.A.; Azemtsop, G.; Houhou, R.; Jehmlich, N.; Bergen, M.v.; Rösch, P.; Popp, J.; Küsel, K. Sulfur-fueled chemolithoautotrophs replenish organic carbon inventory in groundwater. bioRxiv 2021. [Google Scholar] [CrossRef]
- Marques, E.L.S.; Silva, G.S.; Dias, J.C.T.; Gross, E.; Costa, M.S.; Rezende, R.P. Cave drip water-related samples as a natural environment for aromatic hydrocarbon-degrading bacteria. Microorganisms 2019, 7, 33. [Google Scholar] [CrossRef]
- Dimkic, I.; Copic, M.; Petrovic, M.; Stupar, M.; Savkovic, Z.; Knezevic, A.; Subakov Simic, G.; Ljaljevic Grbic, M.; Unkovic, N. Bacteriobiota of the Cave Church of Sts. Peter and Paul in Serbia-culturable and non-culturable communities’ assessment in the bioconservation potential of a peculiar fresco painting. Int. J. Mol. Sci. 2023, 24, 1016. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Pan, J.; Zhang, R.; Wang, J.; Tian, D.; Niu, S. Environmental factors, bacterial interactions and plant traits jointly regulate epiphytic bacterial community composition of two alpine grassland species. Sci. Total Environ. 2022, 836, 155665. [Google Scholar] [CrossRef]
- Wiseschart, A.; Mhuantong, W.; Tangphatsornruang, S.; Chantasingh, D.; Pootanakit, K. Shotgun metagenomic sequencing from Manao-Pee cave, Thailand, reveals insight into the microbial community structure and its metabolic potential. BMC Microbiol. 2019, 19, 144. [Google Scholar] [CrossRef] [PubMed]
- Valverde, A.; Velazquez, E.; Fernandez-Santos, F.; Vizcaino, N.; Rivas, R.; Mateos, P.F.; Martinez-Molina, E.; Igual, J.M.; Willems, A. Phyllobacterium trifolii sp. nov., nodulating Trifolium and Lupinus in Spanish soils. Int. J. Syst. Evol. Microbiol. 2005, 55, 1985–1989. [Google Scholar] [CrossRef]
- Oger, P.M.; Mansouri, H.; Nesme, X.; Dessaux, Y. Engineering root exudation of Lotus toward the production of two novel carbon compounds leads to the selection of distinct microbial populations in the rhizosphere. Microb. Ecol. 2004, 47, 96–103. [Google Scholar] [CrossRef]
- Jurado, V.; Laiz, L.; Gonzalez, J.M.; Hernandez-Marine, M.; Valens, M.; Saiz-Jimenez, C. Phyllobacterium catacumbae sp. nov., a member of the order ‘Rhizobiales’ isolated from Roman catacombs. Int. J. Syst. Evol. Microbiol. 2005, 55, 1487–1490. [Google Scholar] [CrossRef]
- Martin-Sanchez, P.; Jurado, V.; Porca, E.; Bastian, F.; Lacanette, D.; Alabouvette, C.; Saiz-Jimenez, C. Airborne microorganisms in Lascaux Cave (France). Int. J. Speleol. 2014, 43, 295–303. [Google Scholar] [CrossRef]
- Dominguez-Monino, I.; Jurado, V.; Rogerio-Candelera, M.A.; Hermosin, B.; Saiz-Jimenez, C. Airborne bacteria in show caves from Southern Spain. Microb. Cell 2021, 8, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Jurado, V.; Gonzalez-Pimentel, J.L.; Fernandez-Cortes, A.; Martin-Pozas, T.; Ontañon, R.; Palacio, E.; Hermosin, B.; Sanchez-Moral, S.; Saiz-Jimenez, C. Early Detection of Phototrophic Biofilms in the Polychrome Panel, El Castillo Cave, Spain. Appl. Biosci. 2022, 1, 40–63. [Google Scholar] [CrossRef]
- Mergaert, J.; Boley, A.; Cnockaert, M.C.; Muller, W.R.; Swings, J. Identity and potential functions of heterotrophic bacterial isolates from a continuous-upflow fixed-bed reactor for denitrification of drinking water with bacterial polyester as source of carbon and electron donor. Syst. Appl. Microbiol. 2001, 24, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Svec, P.; Kosina, M.; Zeman, M.; Holochova, P.; Kralova, S.; Nemcova, E.; Micenkova, L.; Urvashi; Gupta, V.; Sood, U.; et al. Pseudomonas karstica sp. nov. and Pseudomonas spelaei sp. nov., isolated from calcite moonmilk deposits from caves. Int. J. Syst. Evol. Microbiol. 2020, 70, 5131–5140. [Google Scholar] [CrossRef]
- Haynes, R.J. Nature of the belowground ecosystem and its development during pedogenesis. In Advances in Agronomy; Elsevier Science: Amsterdam, The Netherlands, 2014; pp. 43–109. [Google Scholar]
- Urzi, C.; De Leo, F.; Bruno, L.; Albertano, P. Microbial diversity in paleolithic caves: A study case on the phototrophic biofilms of the Cave of Bats (Zuheros, Spain). Microb. Ecol. 2010, 60, 116–129. [Google Scholar] [CrossRef]
- Yasir, M. Analysis of bacterial communities and characterization of antimicrobial strains from cave microbiota. Braz. J. Microbiol. 2018, 49, 248–257. [Google Scholar] [CrossRef]
- Hamedi, J.; Kafshnouchi, M.; Ranjbaran, M. A Study on actinobacterial diversity of Hampoeil cave and screening of their biological activities. Saudi J. Biol. Sci. 2019, 26, 1587–1595. [Google Scholar] [CrossRef]
- De Leo, F.; Iero, A.; Zammit, G.; Urzì, C. Chemoorganotrophic bacteria isolated from biodeteriorated surfaces in cave and catacombs. Int. J. Speleol. 2012, 41, 125–136. [Google Scholar] [CrossRef]
- Huang, Z.; Sheng, X.F.; Zhao, F.; He, L.Y.; Huang, J.; Wang, Q. Isoptericola nanjingensis sp. nov., a mineral-weathering bacterium. Int. J. Syst. Evol. Microbiol. 2012, 62, 971–976. [Google Scholar] [CrossRef]
- Wu, Y.; Li, W.J.; Tian, W.; Zhang, L.P.; Xu, L.; Shen, Q.R.; Shen, B. Isoptericola jiangsuensis sp. nov., a chitin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2010, 60, 904–908. [Google Scholar] [CrossRef]
- Ue, H.; Matsuo, Y.; Kasai, H.; Yokota, A. Demequina globuliformis sp. nov., Demequina oxidasica sp. nov. and Demequina aurantiaca sp. nov., actinobacteria isolated from marine environments, and proposal of Demequinaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2011, 61, 1322–1329. [Google Scholar] [CrossRef] [PubMed]
- Dettner, K. Chemical defense and toxins of lower terrestrial and freshwater animals. In Comprehensive Natural Products II; Liu, H.-W., Mander, L., Eds.; Elsevier: Oxford, UK, 2010; pp. 387–410. [Google Scholar]
- Simpson, A.G.B.; Eglit, Y. Protist Diversification. In Encyclopedia of Evolutionary Biology; Kliman, R.M., Ed.; Academic Press: Oxford, UK, 2016; pp. 344–360. [Google Scholar]
- Jurado, V.; del Rosal, Y.; Gonzalez-Pimentel, J.; Hermosin, B.; Saiz-Jimenez, C. Biological control of phototrophic biofilms in a show cave: The case of Nerja cave. Appl. Sci. 2020, 10, 3448. [Google Scholar] [CrossRef]
- Reboul, G.; Moreira, D.; Bertolino, P.; Hillebrand-Voiculescu, A.M.; Lopez-Garcia, P. Microbial eukaryotes in the suboxic chemosynthetic ecosystem of Movile Cave, Romania. Environ. Microbiol. Rep. 2019, 11, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Wiser, M.F. Protozoa. In Reference Module in Life Sciences; Elsevier: Amsterdam, The Netherlands, 2022. [Google Scholar]
- Los Huertos, M. The stage: Typologies of aquatic systems. In Ecology and Management of Inland Waters; Elsevier Science: Amsterdam, The Netherlands, 2020; pp. 225–256. [Google Scholar]
- King, N. Choanoflagellates. Curr. Biol. 2005, 15, 113–114. [Google Scholar] [CrossRef]
- Ogórek, R. Fungal communities on rock surfaces in Demänovská Ice Cave and Demänovská Cave of Liberty (Slovakia). Geomicrobiol. J. 2018, 35, 266–276. [Google Scholar] [CrossRef]
- Hampl, V.; Hug, L.; Leigh, J.W.; Dacks, J.B.; Lang, B.F.; Simpson, A.G.; Roger, A.J. Phylogenomic analyses support the monophyly of Excavata and resolve relationships among eukaryotic “supergroups”. Proc. Natl. Acad. Sci. USA 2009, 106, 3859–3864. [Google Scholar] [CrossRef]
- Dorrell, R.G.; Smith, A.G. Do red and green make brown?: Perspectives on plastid acquisitions within chromalveolates. Eukaryot. Cell 2011, 10, 856–868. [Google Scholar] [CrossRef]
- Tikhonenkov, D.V. Predatory flagellates-the new recently discovered deep branches of the eukaryotic tree and their evolutionary and ecological significance. Protistology 2020, 14, 15–22. [Google Scholar] [CrossRef]
- Cahoon, A.B.; VanGundy, R.D. Alveolates (dinoflagellates, ciliates and apicomplexans) and Rhizarians are the most common microbial eukaryotes in temperate Appalachian karst caves. Environ. Microbiol. Rep. 2022, 14, 538–548. [Google Scholar] [CrossRef]
- Schilde, C. Amoebozoa. In Reference Module in Life Sciences; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Landolt, J.; Stephenson, S.; Slay, M. Dictyostelid cellular slime molds from caves. J. Cave Karst Stud. 2006, 68, 22–26. [Google Scholar]
- Cavalier-Smith, T. Kingdom Chromista and its eight phyla: A new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences. Protoplasma 2018, 255, 297–357. [Google Scholar] [CrossRef] [PubMed]
- Rioux, L.-E.; Turgeon, S.L. Seaweed carbohydrates. In Seaweed Sustainability; Academic Press: San Diego, CA, USA, 2015; pp. 141–192. [Google Scholar]
- Biagioli, F.; Coleine, C.; Piano, E.; Nicolosi, G.; Poli, A.; Prigione, V.; Zanellati, A.; Varese, C.; Isaia, M.; Selbmann, L. Microbial diversity and proxy species for human impact in Italian karst caves. Sci. Rep. 2023, 13, 689. [Google Scholar] [CrossRef] [PubMed]
- van Vuuren, S.J.; du Preez, G.; Levanets, A.; Maree, L. Epilythic cyanobacteria and algae in two geologically distinct caves in South Africa. J. Cave Karst Stud. 2019, 81, 254–263. [Google Scholar] [CrossRef]
- Havlena, Z.; Kieft, T.L.; Veni, G.; Horrocks, R.D.; Jones, D.S. Lighting effects on the development and diversity of photosynthetic biofilm communities in Carlsbad Cavern, New Mexico. Appl. Environ. Microbiol. 2021, 87, e02695-20. [Google Scholar] [CrossRef]
- Abolafia, J.; Peña-Santiago, R. Description and SEM observations of Stegelletina coprophila sp.n. (Nematoda: Rhabditida) from Caves of Andalucía Oriental, Spain. J. Nematol. 2006, 38, 411–417. [Google Scholar]
- Traunspurger, W. Nematoda. In Encyclopedia of Inland Waters; Likens, G.E., Ed.; Academic Press: Oxford, UK, 2009; pp. 372–383. [Google Scholar]
- Kulaš, A.; Gulin, V.; Matoničkin Kepčija, R.; Žutinić, P.; Sertić Perić, M.; Orlić, S.; Kajan, K.; Stoeck, T.; Lentendu, G.; Čanjevac, I.; et al. Ciliates (Alveolata, Ciliophora) as bioindicators of environmental pressure: A karstic river case. Ecol. Indic 2021, 124, 107430. [Google Scholar] [CrossRef]
- Bass, D.; Cavalier-Smith, T. Phylum-specific environmental DNA analysis reveals remarkably high global biodiversity of Cercozoa (Protozoa). Int. J. Syst. Evol. Microbiol. 2004, 54, 2393–2404. [Google Scholar] [CrossRef]
- Sanders, R.W. Protists. In Encyclopedia of Inland Waters; Likens, G.E., Ed.; Academic Press: Oxford, UK, 2009; pp. 252–260. [Google Scholar]
- Cennamo, P.; Marzano, C.; Ciniglia, C.; Pinto, G.; Cappelletti, P.; Caputo, P.; Pollio, A. A survey of the algal flora of anthropogenic caves of Campi Flegrei (Naples, Italy) archeological district. J. Cave Karst Stud. 2012, 74, 243–250. [Google Scholar] [CrossRef]
- Mulec, J.; Kosi, G.; Danijel, A.; Ek, V. Characterization of cave aerophytic algal communities and effects of irradiance levels on production of pigments. J. Cave Karst Stud 2008, 70, 3–12. [Google Scholar]
- Mondini, A.; Anwar, M.Z.; Ellegaard-Jensen, L.; Lavin, P.; Jacobsen, C.S.; Purcarea, C. Heat shock response of the active microbiome from Perennial Cave Ice. Front. Microbiol. 2021, 12, 809076. [Google Scholar] [CrossRef]
- Jeong, M.; Kim, J.I.; Nam, S.W.; Shin, W. Molecular phylogeny and taxonomy of the genus Spumella (Chrysophyceae) based on morphological and molecular evidence. Front. Plant Sci. 2021, 12, 758067. [Google Scholar] [CrossRef] [PubMed]
- Ozimek, E.; Hanaka, A. Mortierella species as the plant growth-promoting fungi present in the agricultural soils. Agriculture 2020, 11, 7. [Google Scholar] [CrossRef]
- Vickerman, K.; Le Ray, D.; Hoef-Emden, K.; De Jonckheere, J. The soil flagellate Proleptomonas faecicola: Cell organisation and phylogeny suggest that the only described free-living trypanosomatid is not a kinetoplastid but has cercomonad affinities. Protist 2002, 153, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Martin-Pozas, T.; Novakova, A.; Jurado, V.; Fernandez-Cortes, A.; Cuezva, S.; Saiz-Jimenez, C.; Sanchez-Moral, S. Diversity of microfungi in a high radon cave ecosystem. Front. Microbiol. 2022, 13, 869661. [Google Scholar] [CrossRef] [PubMed]
- Curcic, B.P.; Sudhaus, W.; Dimitrijevic, R.N.; Makarov, S.E.; Tomic, V.T. Rhabditophanes schneideri (Rhabditida) phoretic on a cave pseudoscorpion. J. Invertebr. Pathol. 2008, 99, 254–256. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, M.G.; Yang, J.; Ai, Y.; Xu, R. Community characteristics of soil Ciliates at Baiyun Mountain, Guangzhou, China. Zool. Stud. 2010, 49, 713–723. [Google Scholar]
- Bharti, D.; Kumar, S.; Buonanno, F.; Ortenzi, C.; Montanari, A.; Quintela-Alonso, P.; La Terza, A. Free living ciliated protists from the chemoautotrophic cave ecosystem of Frasassi (Italy). Subterr. Biol. 2022, 44, 167–198. [Google Scholar] [CrossRef]
- Bare, J.; Sabbe, K.; Van Wichelen, J.; van Gremberghe, I.; D’Hondt, S.; Houf, K. Diversity and habitat specificity of free-living protozoa in commercial poultry houses. Appl. Environ. Microbiol. 2009, 75, 1417–1426. [Google Scholar] [CrossRef]
- Russo, R.; Palla, F. Plant Essential Oils as Biocides in Sustainable Strategies for the Conservation of Cultural Heritage. Sustainability 2023, 15, 8522. [Google Scholar] [CrossRef]
- Chouhan, S.; Sharma, K.; Guleria, S. Antimicrobial activity of some essential oils-present status and future perspectives. Medicines 2017, 4, 58. [Google Scholar] [CrossRef]
- Nguefack, J.; Budde, B.B.; Jakobsen, M. Five essential oils from aromatic plants of Cameroon: Their antibacterial activity and ability to permeabilize the cytoplasmic membrane of Listeria innocua examined by flow cytometry. Lett. Appl. Microbiol. 2004, 39, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhou, F.; Ji, B.P.; Pei, R.S.; Xu, N. The antibacterial mechanism of carvacrol and thymol against Escherichia coli. Lett. Appl. Microbiol. 2008, 47, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Helander, I.M.; Alakomi, H.-L.; Latva-Kala, K.; Mattila-Sandholm, T.; Pol, I.; Smid, E.J.; Gorris, L.G.M.; von Wright, A. Characterization of the action of selected essential oil components on Gram-negative bacteria. J. Agric. Food Chem. 1998, 46, 3590–3595. [Google Scholar] [CrossRef]
- Lambert, R.J.; Skandamis, P.N.; Coote, P.J.; Nychas, G.J. A study of the minimum inhibitory concentration and mode of action of oregano essential oil, thymol and carvacrol. J. Appl. Microbiol. 2001, 91, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Oussalah, M.; Caillet, S.; Lacroix, M. Mechanism of action of Spanish oregano, Chinese cinnamon, and savory essential oils against cell membranes and walls of Escherichia coli O157:H7 and Listeria monocytogenes. J. Food Prot. 2006, 69, 1046–1055. [Google Scholar] [CrossRef]
- Fisher, K.; Phillips, C. The mechanism of action of a citrus oil blend against Enterococcus faecium and Enterococcus faecalis. J. Appl. Microbiol. 2009, 106, 1343–1349. [Google Scholar] [CrossRef]
- Fitzgerald, D.J.; Stratford, M.; Gasson, M.J.; Ueckert, J.; Bos, A.; Narbad, A. Mode of antimicrobial action of vanillin against Escherichia coli, Lactobacillus plantarum and Listeria innocua. J. Appl. Microbiol. 2004, 97, 104–113. [Google Scholar] [CrossRef]
- Gatti, L.; Troiano, F.; Vacchini, V.; Cappitelli, F.; Balloi, A. An In Vitro Evaluation of the Biocidal Effect of Oregano and Cloves’ Volatile Compounds against Microorganisms Colonizing an Oil Painting—A Pioneer Study. Appl. Sci. 2020, 11, 78. [Google Scholar] [CrossRef]
- Mulec, J.; Oarga-Mulec, A. ATP luminescence assay as a bioburden estimator of biomass accumulation in caves. Int. J. Speleol. 2016, 45, 207–218. [Google Scholar] [CrossRef]
- Caillet, S.; Ursachi, L.; Shareck, F.; Lacroix, M. Effect of gamma radiation and oregano essential oil on murein and ATP concentration of Staphylococcus aureus. J. Food Sci. 2009, 74, M499–M508. [Google Scholar] [CrossRef]
- Gill, A.O.; Holley, R.A. Mechanisms of bactericidal action of cinnamaldehyde against Listeria monocytogenes and of eugenol against L. monocytogenes and Lactobacillus sakei. Appl. Environ. Microbiol. 2004, 70, 5750–5755. [Google Scholar] [CrossRef] [PubMed]
- Stupar, M.; Grbić, M.L.; Džamić, A.; Unković, N.; Ristić, M.; Jelikić, A.; Vukojević, J. Antifungal activity of selected essential oils and biocide benzalkonium chloride against the fungi isolated from cultural heritage objects. S. Afr. J. Bot. 2014, 93, 118–124. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 16S rDNA Amplicon | 18S rDNA Amplicon | |||
|---|---|---|---|---|
| Samples | Number of Reads * | Number of OTUs | Number of Reads * | Number of OTUs |
| Green Spot 1 | 32,466 | 184 | 50,601 | 35 |
| Green Spot 2 | 31,439 | 221 | 5761 | 35 |
| Black Spot | 24,411 | 189 | 44,977 | 59 |
| AΤP (RLU) | Before EO Treatment | After Oregano Oil Treatment | After Cinnamon Oil Treatment | After Application of the Mixed Solution (Oregano and Cinnamon Oil) |
|---|---|---|---|---|
| Area 1 (green spot) | 867.200 | 481.929 (44%) * | 103.450 (88%) | 45.163 (95%) |
| Area 2 (green spot) | 148.226 | 20.561 (86%) | 9.689 (93%) | 5.719 (96%) |
| Area 3 (black spot) | 299.099 | 70.206 (77%) | 33.453 (89%) | 16.230 (95%) |
| Mean % average | 0 ± 0% a | 69.0 ± 22.1% b | 90.0 ± 2.6%c | 95.3 ± 0.6% c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsouggou, N.; Oikonomou, A.; Papadimitriou, K.; Skandamis, P.N. 16S and 18S rDNA Amplicon Sequencing Analysis of Aesthetically Problematic Microbial Mats on the Walls of the Petralona Cave: The Use of Essential Oils as a Cleaning Method. Microorganisms 2023, 11, 2681. https://doi.org/10.3390/microorganisms11112681
Tsouggou N, Oikonomou A, Papadimitriou K, Skandamis PN. 16S and 18S rDNA Amplicon Sequencing Analysis of Aesthetically Problematic Microbial Mats on the Walls of the Petralona Cave: The Use of Essential Oils as a Cleaning Method. Microorganisms. 2023; 11(11):2681. https://doi.org/10.3390/microorganisms11112681
Chicago/Turabian StyleTsouggou, Natalia, Alexandra Oikonomou, Konstantinos Papadimitriou, and Panagiotis N. Skandamis. 2023. "16S and 18S rDNA Amplicon Sequencing Analysis of Aesthetically Problematic Microbial Mats on the Walls of the Petralona Cave: The Use of Essential Oils as a Cleaning Method" Microorganisms 11, no. 11: 2681. https://doi.org/10.3390/microorganisms11112681