
Vaccination of White-Tailed Deer with Mycobacterium bovis Bacillus Calmette–Guérin (BCG): Effect of Mycobacterium avium ssp. paratuberculosis Infection
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Deer, Vaccination, and Challenge
2.2. Postmortem Examination and Sample Collection
2.3. M. bovis BCG Vaccine and M. bovis Challenge Inoculum
2.4. Culture of Feces and Tissues for the Isolation of Map
2.5. Culture Confirmation Using PCR
2.6. Direct Fecal PCR Testing
2.7. Culture of Tissues for M. bovis
2.8. Statistics
3. Results
3.1. Bacteriologic Culture
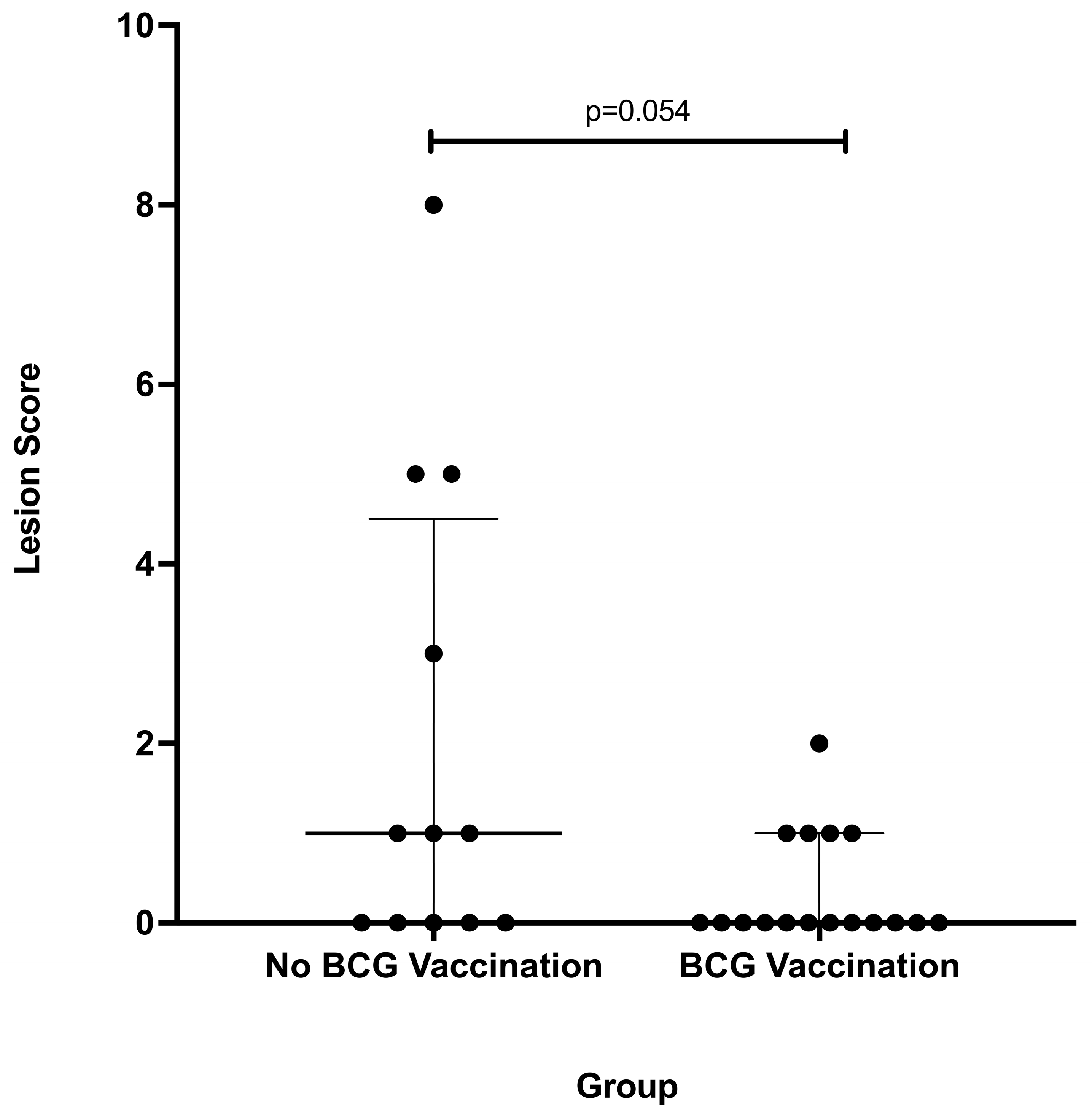
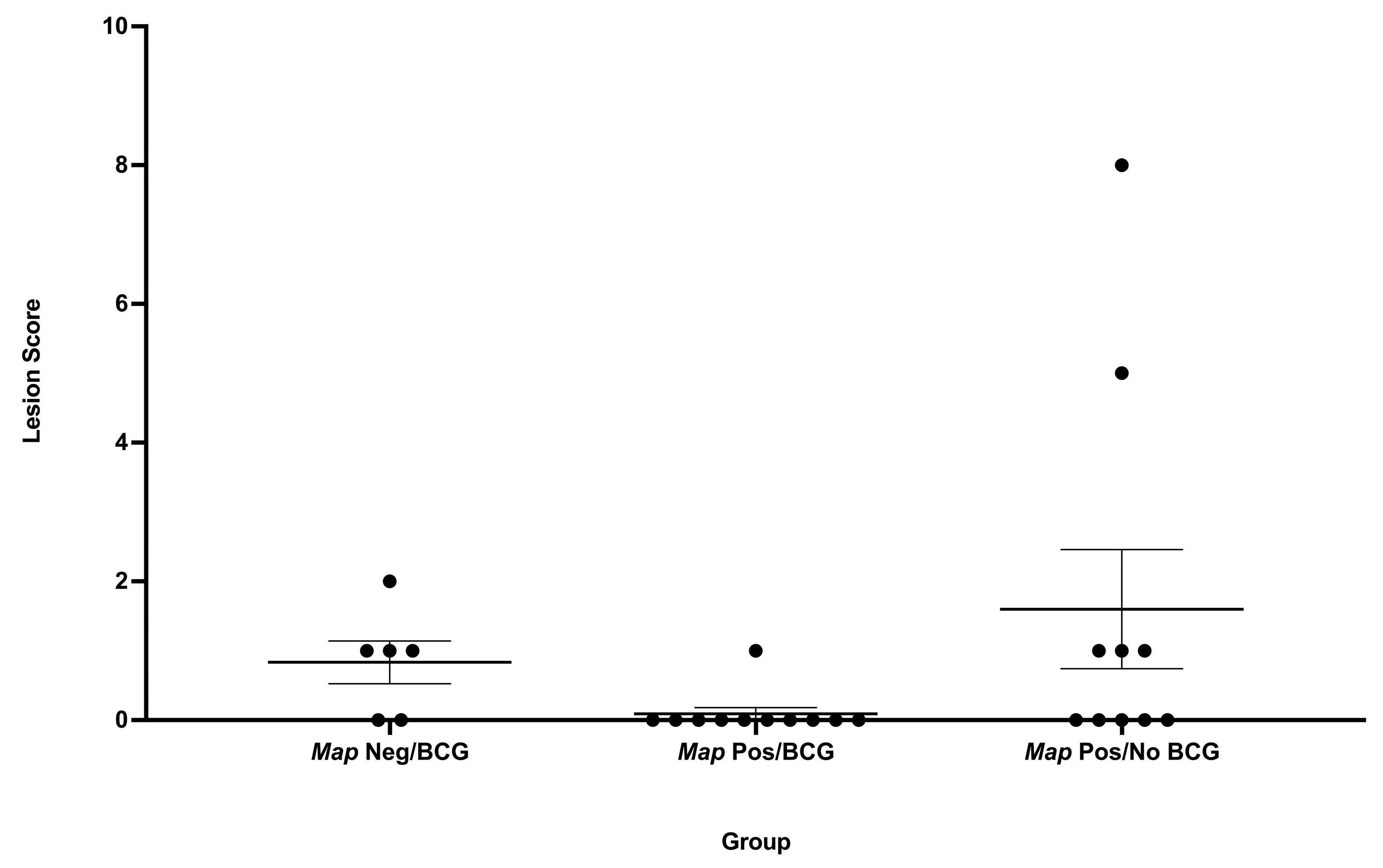
3.2. Gross Lesion Severity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gopinath, K.; Singh, S. Non-tuberculous mycobacteria in TB-endemic countries: Are we neglecting the danger? PLoS Negl. Trop. Dis. 2010, 4, e615. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Campos, S.; Smith, N.H.; Boniotti, M.B.; Aranaz, A. Overview and phylogeny of Mycobacterium tuberculosis complex organisms: Implications for diagnostics and legislation of bovine tuberculosis. Res. Vet. Sci. 2014, 97, S5–S19. [Google Scholar] [CrossRef] [PubMed]
- Coscolla, M.; Gagneux, S. Consequences of genomic diversity in Mycobacterium tuberculosis. Sem. Immunol. 2014, 26, 431–444. [Google Scholar] [CrossRef]
- Cousins, D.V.; Bastida, R.; Cataldi, A.; Quse, V.; Redrobe, S.; Dow, S.; Duignan, P.; Murray, A.; Dupont, C.; Ahmed, N.; et al. Tuberculosis in seals caused by a novel member of the Mycobacterium tuberculosis complex: Mycobacterium pinnipedii sp. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Cousins, D.V.; Francis, B.R.; Gow, B.L.; Collins, D.M.; McGlashan, C.H.; Gregory, A.; Mackenzie, R.M. Tuberculosis in captive seals: Bacteriological studies on an isolate belonging to the Mycobacterium tuberculosis complex. Res. Vet. Sci. 1990, 48, 196–200. [Google Scholar] [CrossRef]
- Cousins, D.V.; Peet, R.L.; Gaynor, W.T.; Williams, S.N.; Gow, B.L. Tuberculosis in imported hyrax (Procavia capensis) caused by an unusual variant belonging to the Mycobacterium tuberculosis complex. Vet. Microbiol. 1994, 42, 135–145. [Google Scholar] [CrossRef]
- Parsons, S.D.; Drewe, J.A.; Gey van Pittius, N.C.; Warren, R.M.; van Helden, P.D. Novel cause of tuberculosis in meerkats, South Africa. Emerg. Infect. Dis. 2013, 19, 2004–2007. [Google Scholar] [CrossRef]
- Alexander, K.A.; Laver, P.N.; Michel, A.L.; Williams, M.; van Helden, P.D.; Warren, R.M.; Gey van Pittius, N.C. Novel Mycobacterium tuberculosis complex pathogen, M. mungi. Emerg. Infect. Dis. 2010, 16, 1296–1299. [Google Scholar] [CrossRef]
- Sotiriou, M.C.; Stryjewska, B.M.; Hill, C. Two Cases of Leprosy in Siblings Caused by Mycobacterium lepromatosis and Review of the Literature. Am. J. Trop. Med. Hyg. 2016, 95, 522–527. [Google Scholar] [CrossRef]
- Katoch, V.M. Infections due to non-tuberculous mycobacteria (NTM). Indian. J. Med. Res. 2004, 120, 290–304. [Google Scholar]
- Arrey, F.; Löwe, D.; Kuhlmann, S.; Kaiser, P.; Moura-Alves, P.; Krishnamoorthy, G.; Lozza, L.; Maertzdorf, J.; Skrahina, T.; Skrahina, A.; et al. Humanized mouse model mimicking pathology of human tuberculosis for in vivo evaluation of drug regimens. Front. Immunol. 2019, 10, 89. [Google Scholar] [CrossRef]
- van Ingen, J.; Turenne, C.Y.; Tortoli, E.; Wallace, R.J., Jr.; Brown-Elliott, B.A. A definition of the Mycobacterium avium complex for taxonomical and clinical purposes, a review. Int. J. Syst. Evol. Microbiol. 2018, 68, 3666–3677. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Waters, W.R. Bovine tuberculosis and the establishment of an eradication program in the United States: Role of veterinarians. Vet. Med. Int. 2011, 2011, 816345. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Thacker, T.C.; Waters, W.R.; Gortazar, C.; Corner, L.A. Mycobacterium bovis: A model pathogen at the interface of livestock, wildlife, and humans. Vet. Med. Int. 2012, 2012, 236205. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, D.J.; Schmitt, S.M.; Fitzgerald, S.D.; Berry, D.E. Management of bovine tuberculosis in Michigan wildlife: Current status and near term prospects. Vet. Microbiol. 2011, 151, 179–187. [Google Scholar] [CrossRef]
- O’Brien, D.J.; Schmitt, S.M.; Rudolph, B.A.; Nugent, G. Recent advances in the management of bovine tuberculosis in free-ranging wildlife. Vet. Microbiol. 2011, 151, 23–33. [Google Scholar] [CrossRef]
- Sunstrom, J.; Shoyinka, A.; Power, L.E.; Maxwell, D.; Stobierski, M.G.; Signs, K.; Sidge, J.L.; O’Brien, D.J.; Robbe-Austerman, S.; Davidson, P. Zoonotic Mycobacterium bovis disease in deer hunters—Michigan, 2002–2017. Morb. Mortal Wkly. Rep. 2019, 68, 807–808. [Google Scholar] [CrossRef]
- Abdallah, A.M.; Behr, M.A. Evolution and strain variation in BCG. Adv. Exp. Med. Biol. 2017, 1019, 155–169. [Google Scholar] [CrossRef]
- Nol, P.; Palmer, M.V.; Waters, W.R.; Aldwell, F.E.; Buddle, B.M.; Triantis, J.M.; Linke, L.M.; Phillips, G.E.; Thacker, T.C.; Rhyan, J.C.; et al. Efficacy of oral and parenteral routes of Mycobacterium bovis bacille Calmette-Guerin vaccination against experimental bovine tuberculosis in white-tailed deer (Odocoileus virginianus): A feasibility study. J. Wildl. Dis. 2008, 44, 247–259. [Google Scholar] [CrossRef]
- Palmer, M.V.; Thacker, T.C.; Waters, W.R. Vaccination with Mycobacterium bovis BCG strains Danish and Pasteur in white-tailed deer (Odocoileus virginianus) experimentally challenged with Mycobacterium bovis. Zoonoses Public Health 2009, 56, 243–251. [Google Scholar] [CrossRef]
- Palmer, M.V.; Thacker, T.C.; Waters, W.R. Vaccination of white-tailed deer (Odocoileus virginianus) with Mycobacterium bovis bacillus Calmette Guérin. Vaccine 2007, 25, 6589–6597. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Thacker, T.C.; Waters, W.R.; Robbe-Austerman, S. Oral vaccination of white-tailed deer (Odocoileus virginianus) with Mycobacterium bovis Bacillus Calmette-Guérin (BCG). PLoS ONE 2014, 9, e97031. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Thacker, T.C. Use of the human vaccine, Mycobacterium bovis Bacillus Calmette-Guérin in deer. Front. Vet. Sci. 2018, 5, 244. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.E.; Long, M.W. Effects of infection with atypical mycobacteria on BCG vaccination and tuberculosis. Am. Rev. Respir. Dis. 1966, 94, 553–568. [Google Scholar]
- Colditz, G.A.; Berkey, C.S.; Mosteller, F.; Brewer, T.F.; Wilson, M.E.; Burdick, E.; Fineberg, H.V. The efficacy of bacillus Calmette-Guerin vaccination of newborns and infants in the prevention of tuberculosis: Meta-analysis of the published literature. Pediatrics 1995, 96, 29–35. [Google Scholar] [CrossRef]
- Buddle, B.M.; Vordermeier, H.M.; Chambers, M.A.; de Klerk-Lorist, L.-M. Efficacy and safety of BCG vaccine for control of tuberculosis in domestic livestock and wildlife. Front. Vet. Sci. 2018, 5, 259. [Google Scholar] [CrossRef]
- Edwards, M.L.; Goodrich, J.M.; Muller, D.; Pollack, A.; Ziegler, J.E.; Smith, D.W. Infection with Mycobacterium avium-intracellulare and the protective effects of Bacille Calmette-Guerin. J. Infect. Dis. 1982, 145, 733–741. [Google Scholar] [CrossRef]
- Orme, I.M.; Collins, F.M. Efficacy of Mycobacterium bovis BCG vaccination in mice undergoing prior pulmonary infection with atypical mycobacteria. Infect. Immun. 1984, 44, 28–32. [Google Scholar] [CrossRef]
- Orme, I.A.; Roberts, A.R.; Collins, F.M. Lack of evidence for a reduction in the efficacy of subcutaneous BCG vaccination in mice infected with nontuberculous mycobacteria. Tubercle 1986, 67, 41–46. [Google Scholar] [CrossRef]
- Hope, J.C.; Thom, M.L.; Villarreal-Ramos, B.; Vordermeier, H.M.; Hewinson, R.G.; Howard, C.J. Exposure to Mycobacterium avium induces low-level protection from Mycobacterium bovis infection but compromises diagnosis of disease in cattle. Clin. Exp. Immunol. 2005, 141, 432–439. [Google Scholar] [CrossRef]
- Buddle, B.M.; Wards, B.J.; Aldwell, F.E.; Collins, D.M.; de Lisle, G.W. Influence of sensitisation to environmental mycobacteria on subsequent vaccination against bovine tuberculosis. Vaccine 2002, 20, 1126–1133. [Google Scholar] [CrossRef] [PubMed]
- de Lisle, G.W.; Wards, B.J.; Buddle, B.M.; Collins, D.M. The efficacy of live tuberculosis vaccines after presensitization with Mycobacterium avium. Tuberculosis 2005, 85, 73. [Google Scholar] [CrossRef] [PubMed]
- de Lisle, G.W.; Cannon, M.C.; Yates, G.F.; Collins, D.M. Use of a polymerase chain reaction to subtype Mycobacterium avium subspecies paratuberculosis, an increasingly important pathogen from farmed deer in New Zealand. N. Z. Vet. J. 2006, 54, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Mackintosh, C.G.; de Lisle, G.W.; Collins, D.M.; Griffin, J.F. Mycobacterial diseases of deer. N. Z. Vet. J. 2004, 52, 163–174. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, R.; Hughes, A.; Liggett, S.; Griffin, F. Composite testing for ante-mortem diagnosis of Johne’s disease in farmed New Zealand deer; correlations between bacteriological culture, histopathology, serological reactivity and faecal shedding as determined by quantitative PCR. BMC Vet. Res. 2013, 9, 72. [Google Scholar] [CrossRef] [PubMed]
- Balseiro, A.; Garcia Marin, J.F.; Solano, P.; Garrido, J.M.; Prieto, J.M. Histopathological classification of lesions observed in natural cases of paratuberculosis in free-ranging fallow deer (Dama dama). J. Comp. Pathol. 2008, 138, 180–188. [Google Scholar] [CrossRef]
- Chiodini, R.J.; Van Kruiningen, H.J. Eastern white-tailed deer as a reservoir of ruminant paratuberculosis. J. Am. Vet. Med. Assoc. 1983, 182, 168–169. [Google Scholar]
- Clark, R.G.; Griffin, J.F.; Mackintosh, C.G. Johne’s disease caused by Mycobacterium avium subsp. paratuberculosis infection in red deer (Cervus elaphus): An histopathological grading system, and comparison of paucibacillary and multibacillary disease. N. Z. Vet. J. 2010, 58, 90–97. [Google Scholar] [CrossRef]
- Forde, T.; Orsel, K.; De Buck, J.; Cote, S.D.; Cuyler, C.; Davison, T.; Elkin, B.; Kelly, A.; Kienzler, M.; Popko, R.; et al. Detection of Mycobacterium avium subspecies paratuberculosis in several herds of Arctic Caribou (Rangifer tarandus ssp.). J. Wildl. Dis. 2012, 48, 918–924. [Google Scholar] [CrossRef]
- Murray, H.L.; Yabsley, M.J.; Keel, M.K.; Manning, E.J.; Wilmers, T.J.; Corn, J.L. Persistence of Mycobacterium avium subspecies paratuberculosis in endangered Florida Key deer and Key deer habitat. J. Wildl. Dis. 2014, 50, 349–353. [Google Scholar] [CrossRef]
- Galiero, A.; Leo, S.; Garbarino, C.; Arrigoni, N.; Russo, S.; Giacomelli, S.; Bianchi, A.; Trevisiol, K.; Idrizi, I.; Daka, G.; et al. Mycobacterium avium subsp. paratuberculosis isolated from wild red deer (Cervus elaphus) in Northern Italy. Vet. Microbiol. 2018, 217, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Whittington, R.; Donat, K.; Weber, M.F.; Kelton, D.; Nielsen, S.S.; Eisenberg, S.; Arrigoni, N.; Juste, R.; Saez, J.L.; Dhand, N.; et al. Control of paratuberculosis: Who, why and how. A review of 48 countries. BMC Vet. Res. 2019, 15, 198. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.C.; Figueira, L.; Martins, M.H.; Matos, M.; Alvares, S.; Mendes, A.; Pinto, M.L.; Coelho, A.C. Detection of Mycobacterium avium subsp. paratuberculosis in kidney samples of red deer (Cervus elaphus) in Portugal: Evaluation of different methods. J. Vet. Med. Sci. 2017, 79, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Kopecna, M.; Lamka, J.; Parmova, I.; Trcka, I.; Svastova, P.; Bartos, M.; Pavlik, I. Paratuberculosis in wild ruminants in teh Czech Republic from 1997–2004. Vet. Arh. 2006, 76, S19–S26. [Google Scholar]
- Nebbia, P.; Robino, P.; Ferroglio, E.; Rossi, L.; Meneguz, G.; Rosati, S. Paratuberculosis in red deer (Cervus elaphus hippelaphus) in the Western Alps. Vet. Res. Commun. 2000, 24, 435–443. [Google Scholar] [CrossRef]
- Pavlik, I.; Bartl, J.; Dvorska, L.; Svastova, P.; du Maine, R.; Machackova, M.; Ayele, W.Y.; Horvatho, A. Epidemiology of paratuberculosis in wild ruminants studied by restriction fragment length polymorphism in the Czech Republic during the period 1995–1998. Vet. Microbiol. 2000, 77, 231–251. [Google Scholar] [CrossRef]
- Schmitt, S.M.; Fitzgerald, S.D.; Cooley, T.M.; Bruning-Fann, C.S.; Sullivan, L.; Berry, D.; Carlson, T.; Minnis, R.B.; Payeur, J.B.; Sikarskie, J. Bovine tuberculosis in free-ranging white-tailed deer from Michigan. J. Wildl. Dis. 1997, 33, 749–758. [Google Scholar] [CrossRef]
- Palmer, M.V.; Whipple, D.L.; Payeur, J.B.; Alt, D.P.; Esch, K.J.; Bruning-Fann, C.S.; Kaneene, J.B. Naturally occurring tuberculosis in white-tailed deer. J. Am. Vet. Med. Assoc. 2000, 216, 1921–1924. [Google Scholar] [CrossRef]
- Vordermeier, H.M.; Chambers, M.A.; Cockle, P.J.; Whelan, A.O.; Simmons, J.; Hewinson, R.G. Correlation of ESAT-6-specific gamma interferon production with pathology in cattle following Mycobacterium bovis BCG vaccination against experimental bovine tuberculosis. Infect. Immun. 2002, 70, 3026–3032. [Google Scholar] [CrossRef]
- Wangoo, A.; Johnson, L.; Gough, J.; Ackbar, R.; Inglut, S.; Hicks, D.; Spencer, Y.; Hewinson, G.; Vordermeier, M. Advanced granulomatous lesions in Mycobacterium bovis-infected cattle are associated with increased expression of type I procollagen, gamma delta (WC1+) T cells and CD 68+ cells. J. Comp. Pathol. 2005, 133, 223–234. [Google Scholar] [CrossRef]
- Palmer, M.V.; Waters, W.R.; Thacker, T.C. Lesion development and immunohistochemical changes in granulomas from cattle experimentally infected with Mycobacterium bovis. Vet. Pathol. 2007, 44, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, E.R.; Frank, A.A.; Orme, I.M. Progression of chronic pulmonary tuberculosis in mice aerogenically infected with virulent Mycobacterium tuberculosis. Tuber. Lung Dis. 1997, 78, 57–66. [Google Scholar] [CrossRef] [PubMed]
- FASS. Guide for the Care and Use of Agricultural Animals in Research and Teaching, 3rd ed.; Federation of Animal Science Societies: Champaign, IL, USA, 2010; p. 164. [Google Scholar]
- Garber, J.C. Guide for the Care and Use of Laboratory Animals, 8th ed.; The National Academies Press: Washington, DC, USA, 2011; p. 220. [Google Scholar]
- Bolin, C.A.; Whipple, D.L.; Khanna, K.V.; Risdahl, J.M.; Peterson, P.K.; Molitor, T.W. Infection of swine with Mycobacterium bovis as a model of human tuberculosis. J. Infect. Dis. 1997, 176, 1559–1566. [Google Scholar] [CrossRef]
- Larsen, M.H.; Biermann, K.; Jacobs, W.R., Jr. Laboratory maintenance of Mycobacterium tuberculosis. Curr. Protoc. Microbiol. 2007, 6, 10A.11.11–10A.11.18. [Google Scholar] [CrossRef] [PubMed]
- Robbe-Austerman, S.; Bravo, D.M.; Harris, B. Comparison of the MGIT 960, BACTEC 460 TB and solid media for isolation of Mycobacterium bovis in United States veterinary specimens. BMC Vet. Res. 2013, 9, 74. [Google Scholar] [CrossRef]
- Dykema, P.E.; Stokes, K.D.; Beckwith, N.R.; Mungin, J.W.; Xu, L.; Vickers, D.J.; Reising, M.M.; Bravo, D.M.; Thomsen, B.V.; Robbe-Austerman, S. Development and validation of a direct real-time PCR assay for Mycobacterium bovis and implementation in to the United States national surveillance program. PeerJ Prepr. 2016, 4, e1703v1701. [Google Scholar] [CrossRef]
- Garrido, J.M.; Sevilla, I.A.; Beltran-Beck, B.; Minguijon, E.; Ballesteros, C.; Galindo, R.C.; Boadella, M.; Lyashchenko, K.P.; Romero, B.; Geijo, M.; et al. Protection against tuberculosis in Eurasian wild boar vaccinated with heat-inactivated Mycobacterium bovis. PLoS ONE 2011, 6, e24905. [Google Scholar] [CrossRef]
- Verma, D.; Chan, E.D.; Ordway, D.J. Non-Tuberculous Mycobacteria Interference with BCG-Current Controversies and Future Directions. Vaccines 2020, 8, 688. [Google Scholar] [CrossRef]
- Palmer, M.V.; Thacker, T.C.; Waters, W.R.; Robbe-Austerman, S.; Aldwell, F.E. Persistence of Mycobacterium bovis bacillus Calmette-Guerin (BCG) Danish in white-tailed deer (Odocoileus virginianus) vaccinated with a lipid-formulated oral vaccine. Transbound. Emerg. Dis. 2012, 61, 266–272. [Google Scholar] [CrossRef]
- Thacker, T.C.; Robbe-Austerman, S.; Harris, B.; Palmer, M.V.; Waters, W.R. Isolation of mycobacteria from clinical samples collected in the United States from 2004 to 2011. BMC Vet Res 2013, 9, 100. [Google Scholar] [CrossRef]
- Pate, M.; Zolnir-Dovc, M.; Kusar, D.; Krt, B.; Spicic, S.; Cvetnic, Z.; Ocepek, M. The first report of Mycobacterium celatum isolation from domestic pig (Sus scrofa domestica) and Roe deer (Capreolus capreolus) and an overview of human infections in Slovenia. Vet. Med. Int. 2011, 2011, 432954. [Google Scholar] [CrossRef] [PubMed]
- Ronai, Z.; Eszterbauer, E.; Csivincsik, A.; Guti, C.F.; Dencso, L.; Janosi, S.; Dan, A. Detection of wide genetic diversity and several novel strains among non-avium nontuberculous mycobacteria isolated from farmed and wild animals in Hungary. J. Appl. Microbiol. 2016, 121, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Kaneene, J.B.; Bruning-Fann, C.S.; Granger, L.M.; Miller, R.; Porter-Spalding, B.A. Environmental and farm management factors associated with tuberculosis on cattle farms in northeastern Michigan. J. Am. Vet. Med. Assoc. 2002, 221, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Okafor, C.C.; Grooms, D.L.; Bruning-Fann, C.S.; Averill, J.J.; Kaneene, J.B. Descriptive epidemiology of bovine tuberculosis in Michigan (1975–2010): Lessons learned. Vet. Med. Int. 2011, 2011, 874924. [Google Scholar] [CrossRef] [PubMed]
- VerCauteren, K.C.; Lavelle, M.J.; Campa, H., 3rd. Persistent spillback of bovine tuberculosis from white-tailed deer to cattle in Michigan, USA: Status, strategies, and needs. Front. Vet. Sci. 2018, 5, 301. [Google Scholar] [CrossRef]
- Riemann, H.; Zaman, M.R.; Ruppanner, R.; Aalund, O.; Jorgensen, J.B.; Worsaae, H.; Behymer, D. Paratuberculosis in cattle and free-living exotic deer. J. Am. Vet. Med. Assoc. 1979, 174, 841–843. [Google Scholar]
- Fritsch, I.; Luyven, G.; Kohler, H.; Lutz, W.; Mobius, P. Suspicion of Mycobacterium avium subsp. paratuberculosis transmission between cattle and wild-living red deer (Cervus elaphus) by multitarget genotyping. Appl. Environ. Microbiol. 2012, 78, 1132–1139. [Google Scholar] [CrossRef]
- Raizman, E.A.; Wells, S.J.; Jordan, P.A.; DelGiudice, G.D.; Bey, R.R. Mycobacterium avium subsp. paratuberculosis from free-ranging deer and rabbits surrounding Minnesota dairy herds. Can. J. Vet. Res. 2005, 69, 32–38. [Google Scholar]
- de Lisle, G.W.; Yates, G.F.; Montgomery, H. The emergence of Mycobacterium paratuberculosis in farmed deer in New Zealand—A review of 619 cases. N. Z. Vet. J. 2003, 51, 58–62. [Google Scholar] [CrossRef]
- O’Brien, D.J.; Fierke, J.S.; Carlson, E.S.; Schmitt, S.M. An Evaluation of Liberalized Distribution of Disease Control Permits for White-Tailed Deer, 2008–2010; Report #35653565; Michigan Department of Natural Resources Wildlife Division: Lansing, MI, USA, 2013.
- Forde, T.; Pruvot, M.; DeBuck, J.; Orsel, K. A high-morbidity outbreak of Johne’s disease in game-ranched elk. Can. Vet. J. 2015, 56, 479–483. [Google Scholar]
- Manning, E.J.; Steinberg, H.; Rossow, K.; Ruth, G.R.; Collins, M.T. Epizootic of paratuberculosis in farmed elk. J. Am. Vet. Med. Assoc. 1998, 213, 1280–1281. [Google Scholar]
- Godfroid, J.; Boelaert, F.; Heier, A.; Clavareau, C.; Wellemans, V.; Desmecht, M.; Roels, S.; Walravens, K. First evidence of Johne’s disease in farmed red deer (Cervus elaphus) in Belgium. Vet. Microbiol. 2000, 77, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Machackova, M.; Svastova, P.; Lamka, J.; Parmova, I.; Liska, V.; Smolik, J.; Fischer, O.A.; Pavlik, I. Paratuberculosis in farmed and free-living wild ruminants in the Czech Republic (1999–2001). Vet. Microbiol. 2004, 101, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Whipple, D.L.; Butler, K.L.; Fitzgerald, S.D.; Bruning-Fann, C.S.; Schmitt, S.M. Tonsillar lesions in white-tailed deer (Odocoileus virginianus) naturally infected with Mycobacterium bovis. Vet. Rec. 2002, 151, 149–150. [Google Scholar] [CrossRef] [PubMed]
- Prieto, J.M.; Balseiro, A.; Casais, R.; Abendano, N.; Fitzgerald, L.E.; Garrido, J.M.; Juste, R.A.; Alonso-Hearn, M. Sensitive and specific enzyme-linked immunosorbent assay for detecting serum antibodies against Mycobacterium avium subsp. paratuberculosis in fallow deer. Clin. Vaccine Immunol. 2014, 21, 1077–1085. [Google Scholar] [CrossRef]
- Palmer, M.V.; Kanipe, C.; Cox, R.; Robbe-Austerman, S.; Thacker, T.C. Characteristics of subclinical Mycobacterium avium ssp. paratuberculosis infection in a captive white-tailed deer herd. J. Vet. Diagn. Investig. 2019, 31, 844–851. [Google Scholar] [CrossRef]
- Davidson, W.R.; Manning, E.J.; Nettles, V.F. Culture and serologic survey for Mycobacterium avium subsp. paratuberculosis infection among southeastern white-tailed deer (Odocoileus virginianus). J. Wildl. Dis. 2004, 40, 301–306. [Google Scholar] [CrossRef]
- Quist, C.F.; Nettles, V.F.; Manning, E.J.; Hall, D.G.; Gaydos, J.K.; Wilmers, T.J.; Lopez, R.R. Paratuberculosis in key deer (Odocoileus virginianus clavium). J. Wildl. Dis. 2002, 38, 729–737. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Assumed Status | BCG/Map POS | No BCG/Map POS | BCG/Map NEG | No BCG/Map NEG | Total |
| n | 8 | 7 | 9 | 5 | 29 |
| True infection status | BCG/Map POS | No BCG/Map POS | BCG/Map NEG | No BCG/Map NEG | |
| n | 11 | 10 | 6 | 2 | 29 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palmer, M.V.; Kanipe, C.; Lehman, K.A.; Thacker, T.C.; Putz, E.J.; Boggiatto, P.M. Vaccination of White-Tailed Deer with Mycobacterium bovis Bacillus Calmette–Guérin (BCG): Effect of Mycobacterium avium ssp. paratuberculosis Infection. Microorganisms 2023, 11, 2488. https://doi.org/10.3390/microorganisms11102488
Palmer MV, Kanipe C, Lehman KA, Thacker TC, Putz EJ, Boggiatto PM. Vaccination of White-Tailed Deer with Mycobacterium bovis Bacillus Calmette–Guérin (BCG): Effect of Mycobacterium avium ssp. paratuberculosis Infection. Microorganisms. 2023; 11(10):2488. https://doi.org/10.3390/microorganisms11102488
Chicago/Turabian StylePalmer, Mitchell V., Carly Kanipe, Kimberly A. Lehman, Tyler C. Thacker, Ellie J. Putz, and Paola M. Boggiatto. 2023. "Vaccination of White-Tailed Deer with Mycobacterium bovis Bacillus Calmette–Guérin (BCG): Effect of Mycobacterium avium ssp. paratuberculosis Infection" Microorganisms 11, no. 10: 2488. https://doi.org/10.3390/microorganisms11102488