
Reevaluation of Hemoparasites in the Black Spiny-Tailed Iguana (Ctenosaura similis) with the First Pathological and Molecular Characterizations of Lankesterella desseri n. sp. and Redescription of Hepatozoon gamezi
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Clinical Examination, and Sampling
2.2. Clinical Pathological Examination
2.3. Pathological Examination
2.4. Nucleic Acid Extraction
2.5. Polymerase Chain Reaction—Partial 18S rDNA and Partial mt COI of the Apicomplexan Parasites
2.6. Immunohistochemistry
2.7. Phylogenetic Analysis
2.7.1. Phylogenetic Analysis of Lankesterella n. sp.
2.7.2. Phylogenetic Analysis of Hepatozoon gamezi
3. Results
3.1. Clinical Examination
3.1.1. Physical Examination, Hematology, and Serum Biochemistry
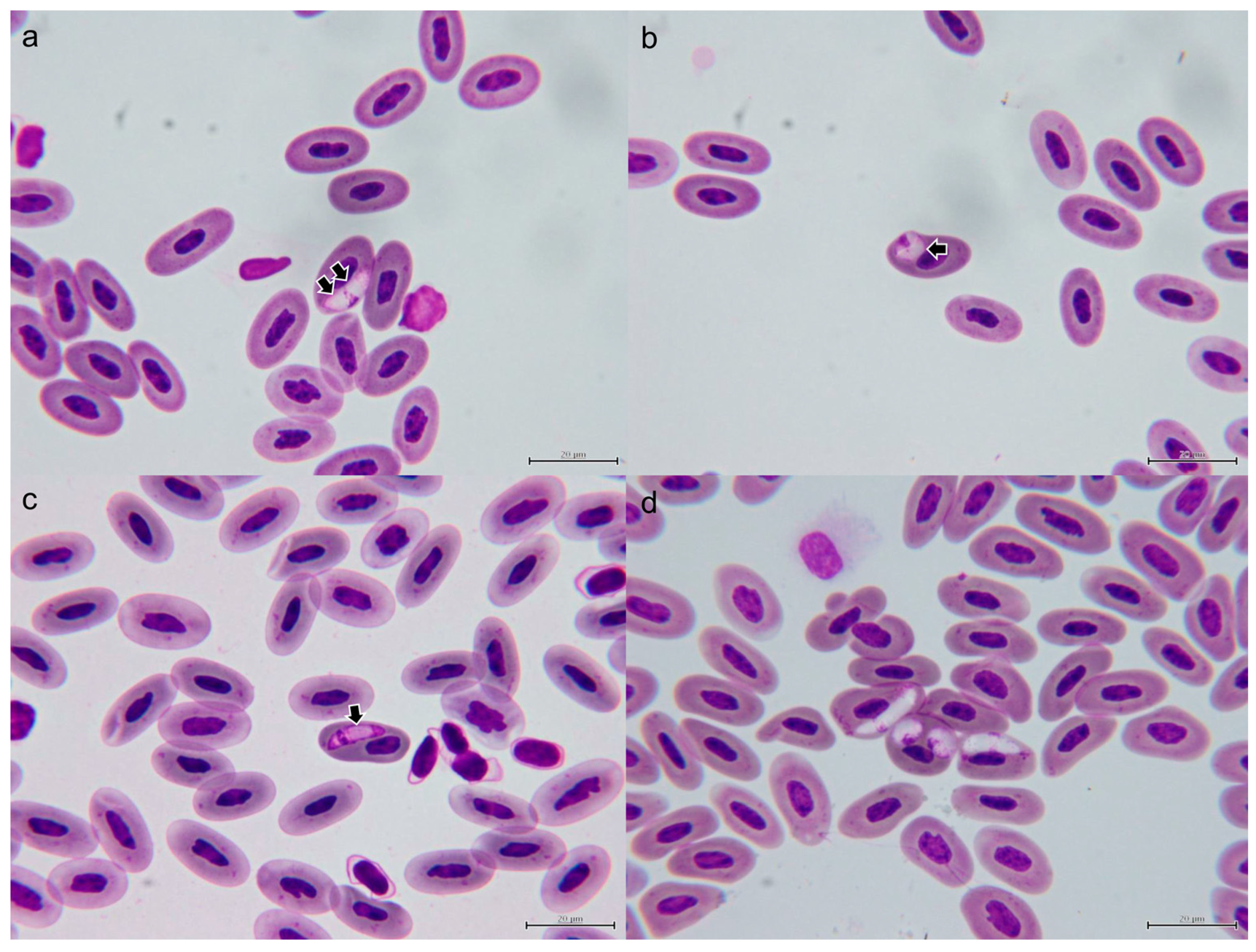
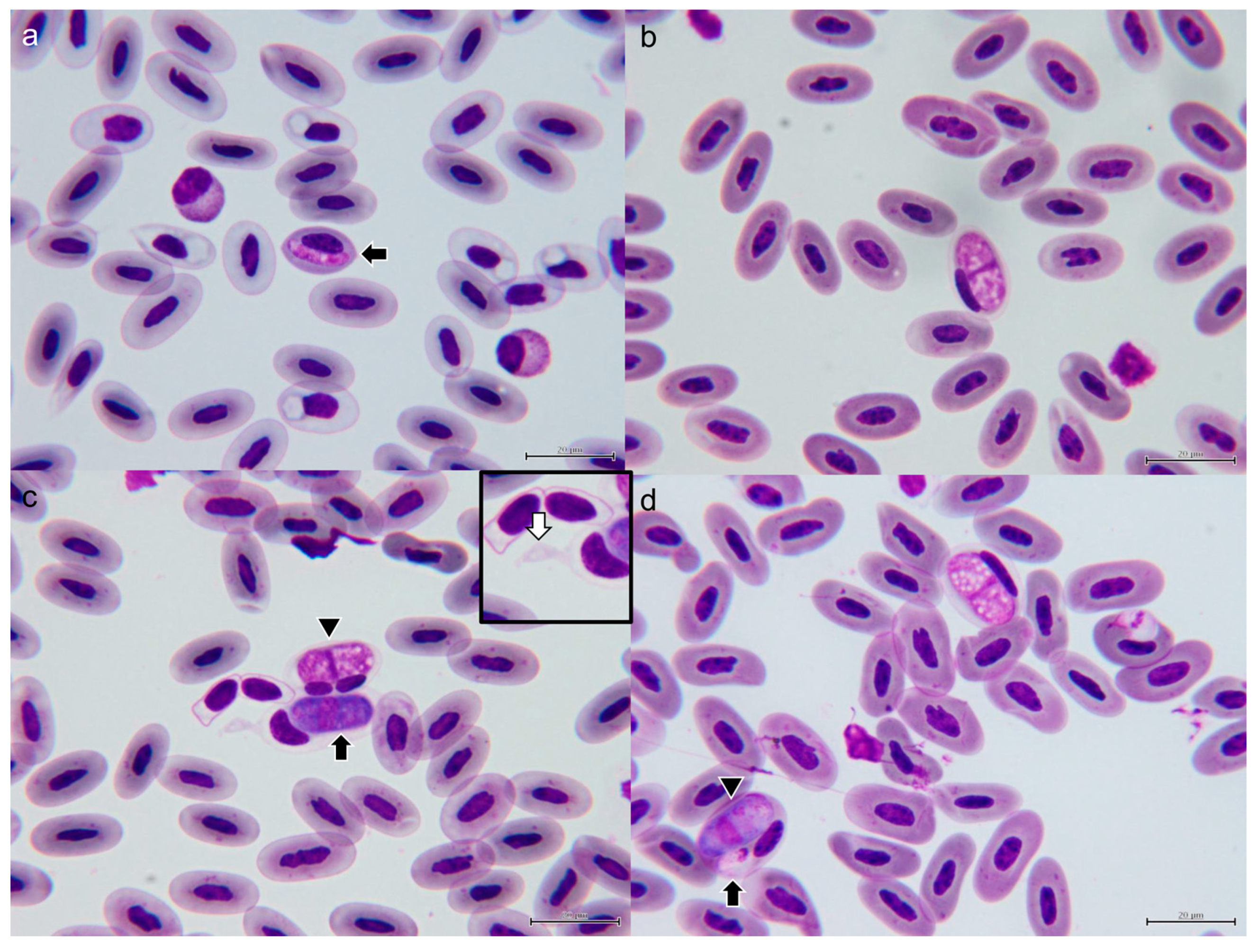
3.1.2. Morphological Characterization of Hemococcidia
3.1.3. Morphological Characterization of Hepatozoon sp.
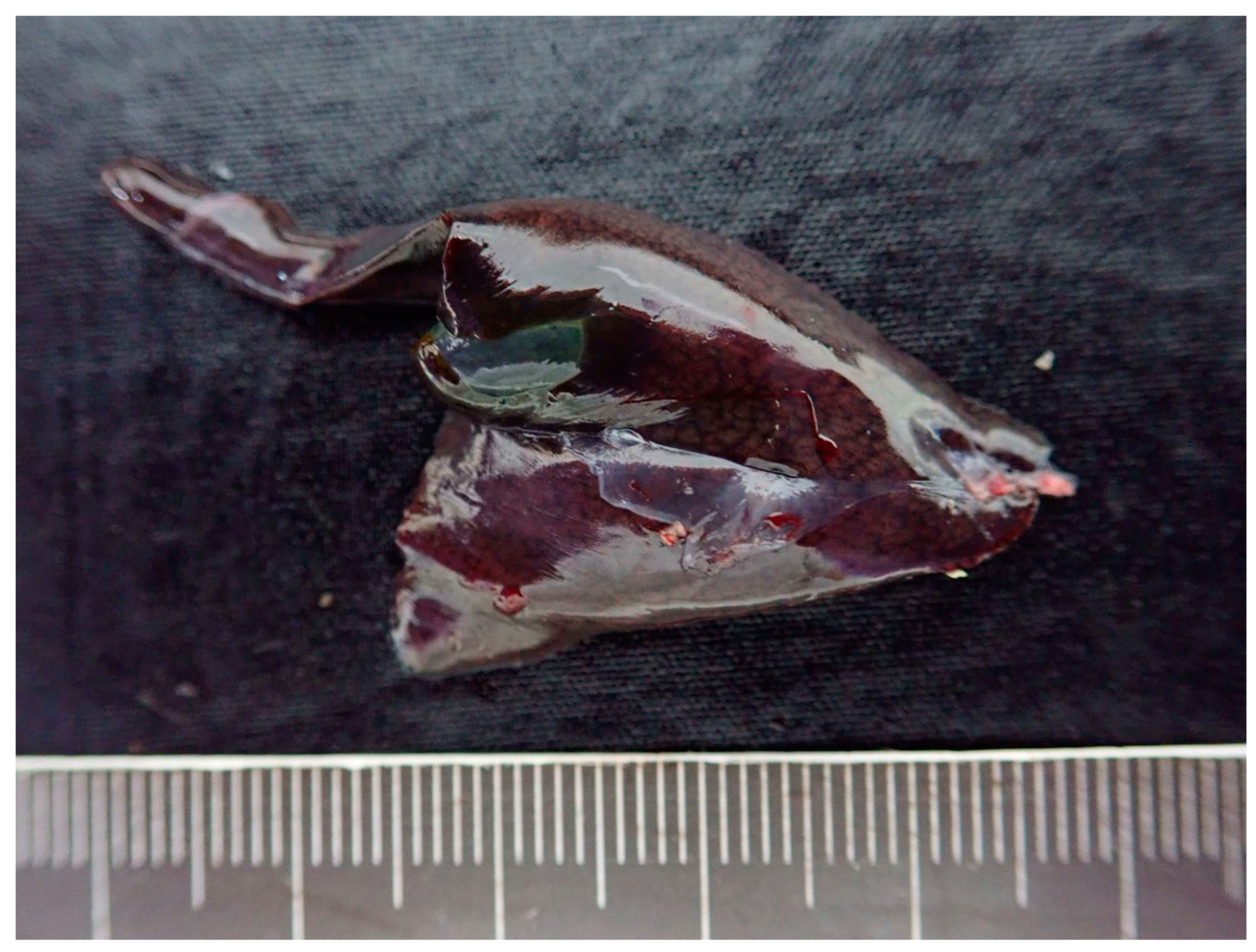
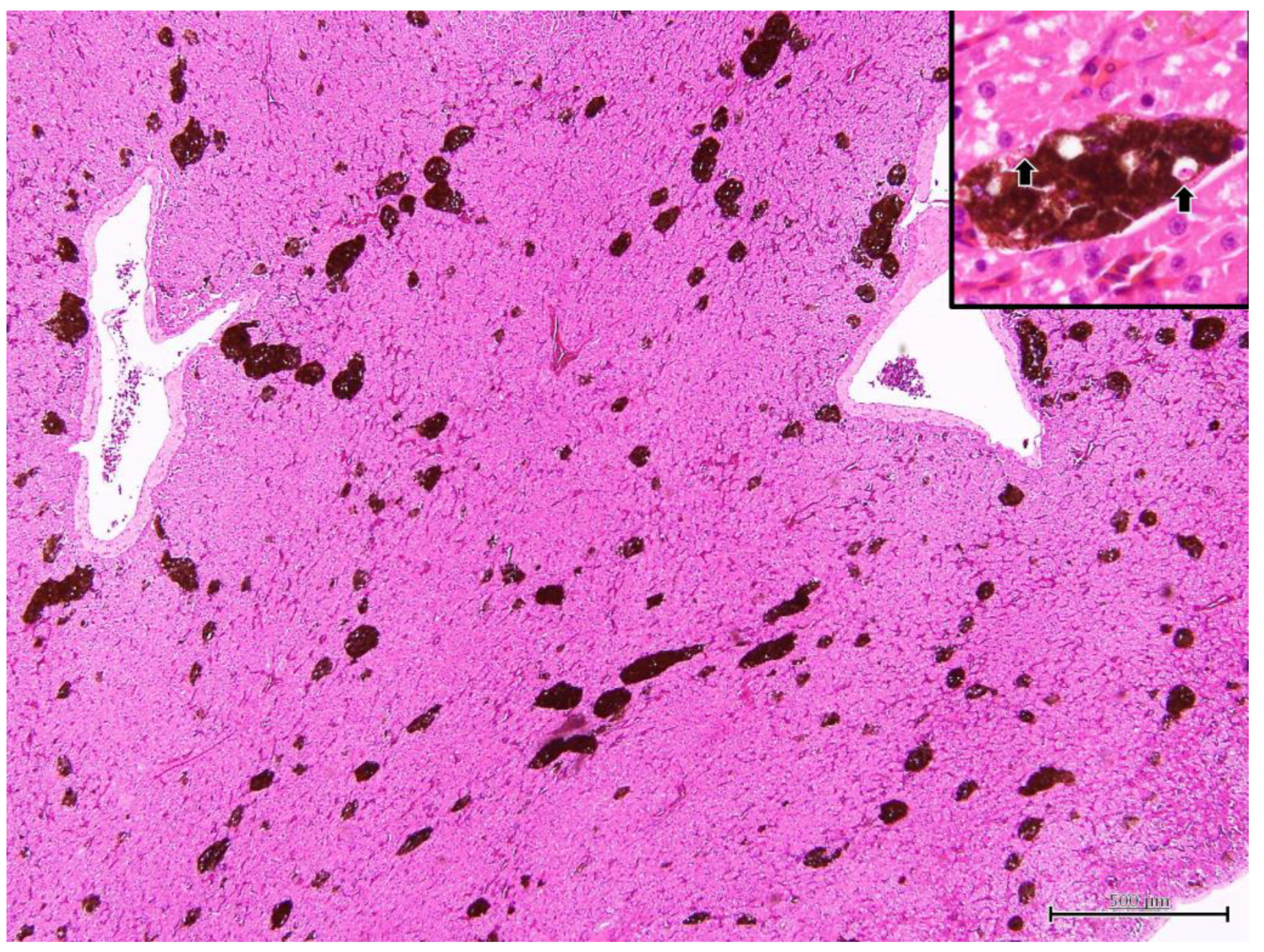
3.2. Postmortem Examination in Iguana 1
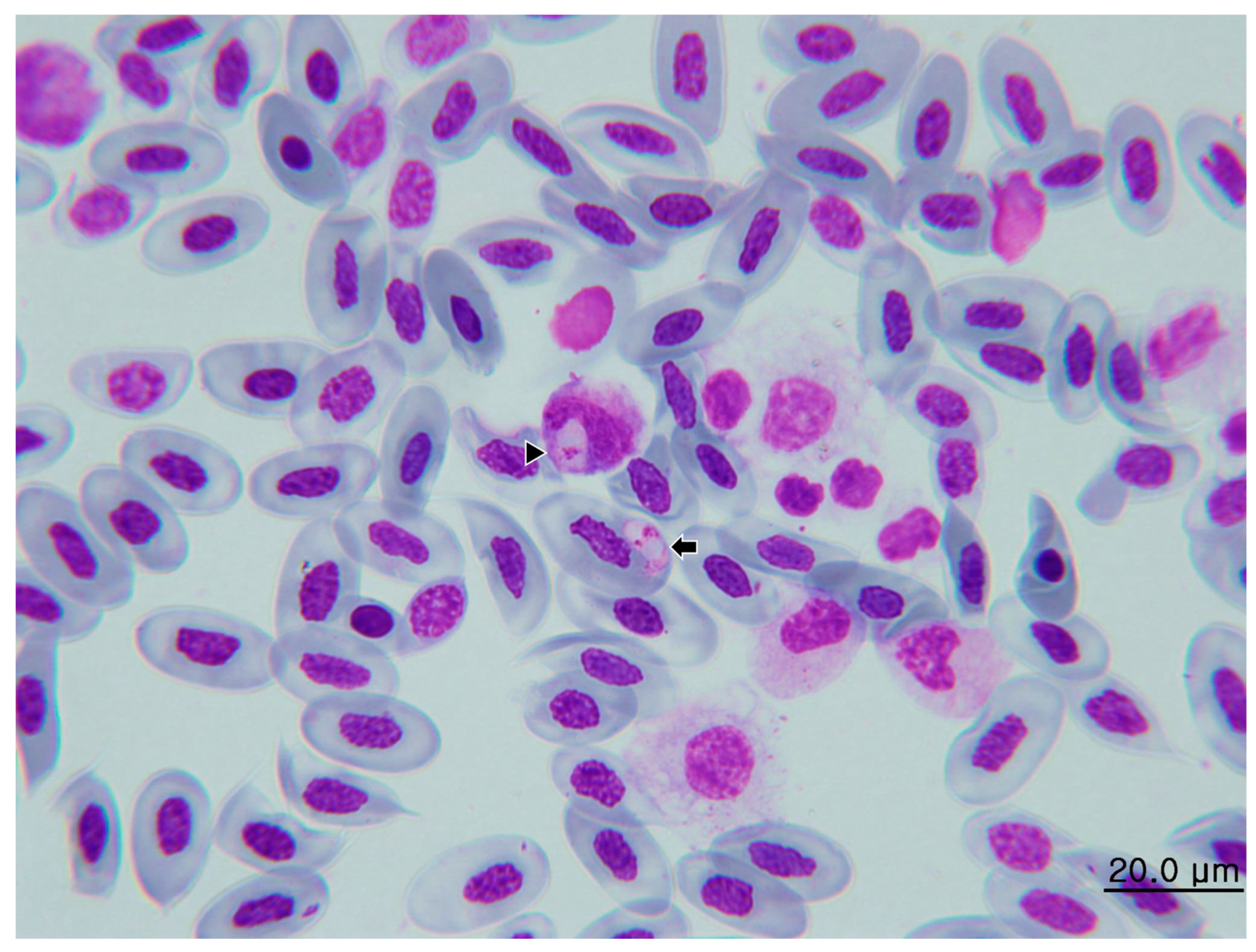
3.3. Immunohistochemistry
3.4. Polymerase Chain Reaction and Nucleotide Sequence Analysis
3.5. Phylogenetic Analysis
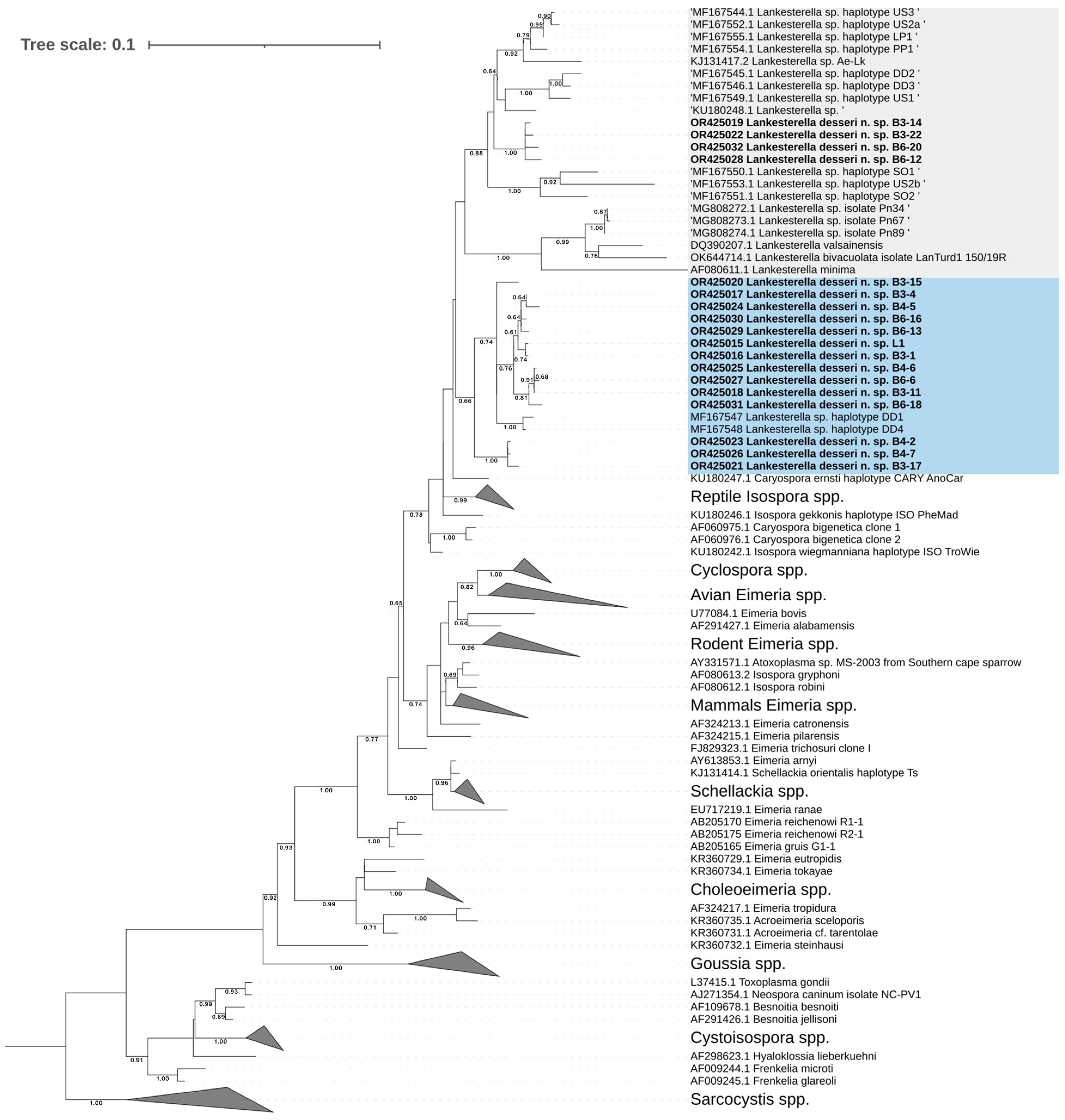
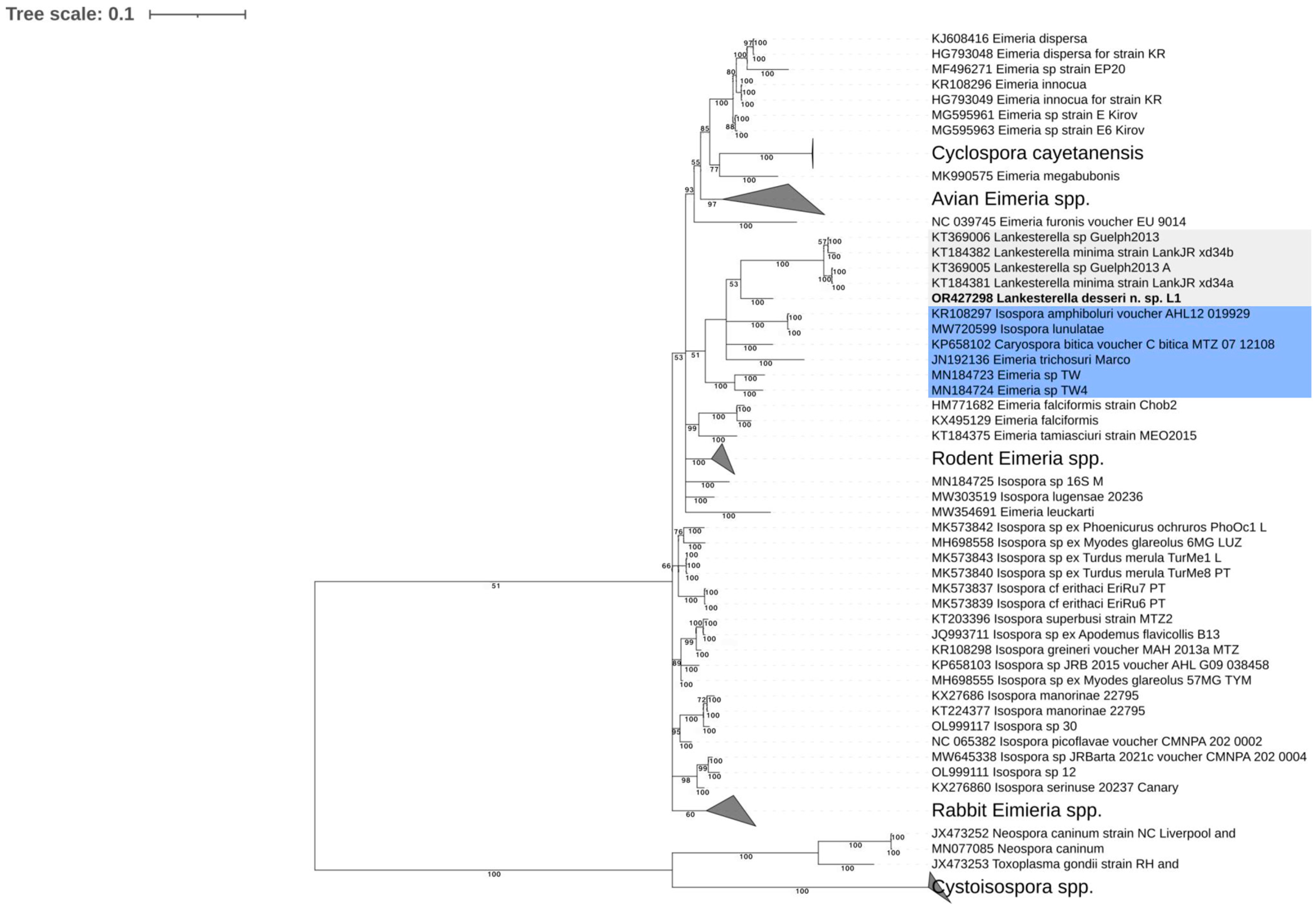
3.5.1. Phylogenetic Analysis of Lankesterella desseri n. sp.
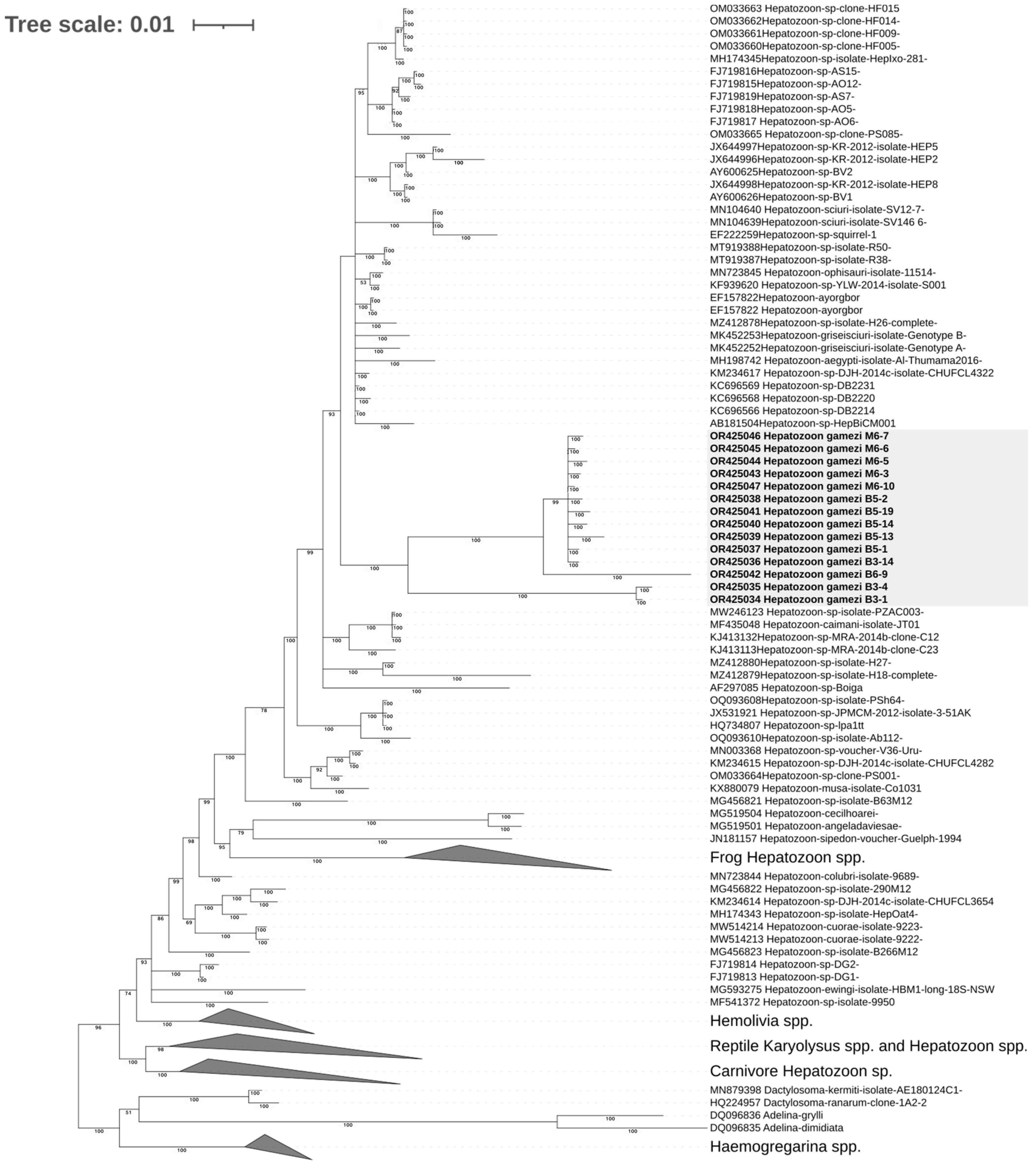
3.5.2. Phylogenetic Analysis of Hepatozoon gamezi
3.6. Description of New Lankesterella Species and Redescription of Hepatozoon gamezi
3.6.1. Lankesterella desseri n. sp.
3.6.2. Hepatozoon gamezi
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Telford, S.R., Jr. Hemoparasites of the Reptilia: Color Atlas and Text; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- O’Donoghue, P. Haemoprotozoa: Making biological sense of molecular phylogenies. Int. J. Parasitol. Parasites Wildl. 2017, 6, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Chagas, C.R.F.; Binkiene, R.; Valkiunas, G. Description and Molecular Characterization of Two Species of Avian Blood Parasites, with Remarks on Circadian Rhythms of Avian Haematozoa Infections (Lankesterella). Animals 2021, 11, 3490. [Google Scholar] [CrossRef] [PubMed]
- Megia-Palma, R.; Martinez, J.; Paranjpe, D.; D’Amico, V.; Aguilar, R.; Palacios, M.G.; Cooper, R.; Ferri-Yáñez, F.; Sinervo, B.; Merino, S. Phylogenetic analyses reveal that Schellackia parasites (Apicomplexa) detected in American lizards are closely related to the genus Lankesterella: Is the range of Schellackia restricted to the Old World? Parasit Vectors 2017, 10, 470. [Google Scholar] [CrossRef] [PubMed]
- Paperna, I.; Martin, C. The development and fine structure of Lankesterella cf. dicroglossi (Apicomplexa: Lankesterellidae) infecting frogs in Niger, West Africa. Folia Parasitol. 2001, 48, 178–186. [Google Scholar] [CrossRef]
- Desser, S.S.; Siddall, M.E.; Barta, J.R. Ultrastructural Observations on the Developmental Stages of Lankesterella Minima (Apicomplexa) in Experimentally Infected Rana Catesbeiana Tadpoles. J. Parasitol. 1990, 76, 97–103. [Google Scholar] [CrossRef]
- Paperna, I.; Ogara, W. Description and ultrastructure of Lankesterella species infecting frogs in Kenya. Parasite 1996, 3, 341–349. [Google Scholar] [CrossRef]
- Ostrovska, K.; Paperna, I. Fine-Structure of Gamont Stages of Schellackia cf. agamae (Lankesterellidae, Eucoccidia) from the Starred Lizard Agama stellio. Z. Fur Parasitenkd. Parasitol. Res. 1987, 73, 492–499. [Google Scholar] [CrossRef]
- Paperna, I.; Finkelman, S. Schellackia ptyodactyli sp. n. of the fan-footed gecko Ptyodactylus hasselquistii from the rift escarpment of the lower Jordan Valley. Folia Parasitol. 1996, 43, 161–172. [Google Scholar]
- Chagas, C.R.F.; Harl, J.; Preiksa, V.; Bukauskaite, D.; Ilgunas, M.; Weissenbock, H.; Valkiūnas, G. Lankesterella (Apicomplexa, Lankesterellidae) Blood Parasites of Passeriform Birds: Prevalence, Molecular and Morphological Characterization, with Notes on Sporozoite Persistence In Vivo and Development In Vitro. Animals 2021, 11, 1451. [Google Scholar] [CrossRef]
- Desser, S.S. Blood parasites of the iguanid lizard, Ctenosaura similis from Costa Rica, with a description of Hepatozoon gamezi n sp. J. Eukaryot. Microbiol. 1997, 44, 162–167. [Google Scholar] [CrossRef]
- Megia-Palma, R.; Martinez, J.; Merino, S. Phylogenetic analysis based on 18S rRNA gene sequences of Schellackia parasites (Apicomplexa: Lankesterellidae) reveals their close relationship to the genus Eimeria. Parasitology 2013, 140, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Megía-Palma, R.; Martínez, J.; Nasri, I.; Cuervo, J.J.; Martín, J.; Acevedo, I.; Belliure, J.; Ortega, J.; García-Roa, R.; Selmi, S.; et al. Phylogenetic relationships of Isospora, Lankesterella, and Caryospora species (Apicomplexa: Eimeriidae) infecting lizards. Org. Divers. Evol. 2015, 16, 275–288. [Google Scholar] [CrossRef]
- Maia, J.P.; Harris, D.J.; Carranza, S.; Gomez-Diaz, E. Assessing the diversity, host-specificity and infection patterns of apicomplexan parasites in reptiles from Oman, Arabia. Parasitology 2016, 143, 1730–1747. [Google Scholar] [CrossRef] [PubMed]
- Tome, B.; Maia, J.; Perera, A.; Carranza, S.; Vasconcelos, R. Parasites in a hotspot: Diversity and specificity patterns of apicomplexans infecting reptiles from the Socotra Archipelago. Parasitology 2021, 148, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Veith, Y.; Wende, A.L.; Matuschewski, K.; Schaer, J.; Muller, K.; Bannert, B. Molecular characterization of Schellackia parasites in an urban population of sand lizards (Lacerta agilis) from Berlin, Germany. Parasitol. Res. 2023, 122, 1759–1764. [Google Scholar] [CrossRef]
- Zechmeisterova, K.; Javanbakht, H.; Kvicerova, J.; Siroky, P. Against growing synonymy: Identification pitfalls of Hepatozoon and Schellackia demonstrated on North Iranian reptiles. Eur. J. Protistol. 2021, 79, 125780. [Google Scholar] [CrossRef]
- Megia-Palma, R.; Martinez, J.; Cuervo, J.J.; Belliure, J.; Jimenez-Robles, O.; Gomes, V.; Cabido, C.; Pausas, J.G.; Fitze, P.S.; Martín, J.; et al. Molecular evidence for host-parasite co-speciation between lizards and Schellackia parasites. Int. J. Parasitol. 2018, 48, 709–718. [Google Scholar] [CrossRef]
- De Paula, F.R.; Picelli, A.M.; Perles, L.; Andre, M.R.; Viana, L.A. A 50-year-old redescription: Molecular and morphometric characterization of Hepatozoon carinicauda Pessoa and Cavalheiro, 1969 in the brown-banded water snake Helicops angulatus (Linnaeus, 1758). Parasitology 2022, 149, 1468–1478. [Google Scholar] [CrossRef]
- Maia, J.P.; Harris, D.J.; Perera, A. Molecular survey of Hepatozoon species in lizards from North Africa. J. Parasitol. 2011, 97, 513–517. [Google Scholar] [CrossRef]
- Gutierrez-Liberato, G.A.; Lotta-Arevalo, I.A.; Rodriguez-Almonacid, C.C.; Vargas-Ramirez, M.; Matta, N.E. Molecular and morphological description of the first Hepatozoon (Apicomplexa: Hepatozoidae) species infecting a neotropical turtle, with an approach to its phylogenetic relationships. Parasitology 2021, 148, 747–759. [Google Scholar] [CrossRef]
- Ogedengbe, M.E.; Ogedengbe, J.D.; Whale, J.C.; Elliot, K.; JuÁRez-Estrada, M.A.; Barta, J.R. Molecular phylogenetic analyses of tissue coccidia (sarcocystidae; apicomplexa) based on nuclear 18s RDNA and mitochondrial COI sequences confirms the paraphyly of the genus Hammondia. Parasitol. Open 2016, 2, e2. [Google Scholar] [CrossRef]
- Pasachnik, S. Ctenosaura similis. The IUCN Red List of Threatened Species 2015. Available online: https://www.iucnredlist.org/es/species/174480/73611567 (accessed on 30 August 2023).
- Divers, S.J. 43—Diagnostic Techniques and Sample Collection. In Mader’s Reptile and Amphibian Medicine and Surgery, 3rd ed.; Divers, S.J., Stahl, S.J., Eds.; W.B. Saunders: St. Louis, MO, USA, 2019; pp. 405–421.e1. [Google Scholar]
- Merino, S.; Potti, J. High Prevalence of Hematozoa in Nestlings of a Passerine Species, the Pied Flycatcher (Ficedula hypoleuca). Auk 1995, 112, 1041–1043. [Google Scholar] [CrossRef]
- Ogedengbe, J.D.; Hanner, R.H.; Barta, J.R. DNA barcoding identifies Eimeria species and contributes to the phylogenetics of coccidian parasites (Eimeriorina, Apicomplexa, Alveolata). Int. J. Parasitol. 2011, 41, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Gericota, B.; Garner, M.M.; Barr, B.; Nordhausen, R.; Larsen, R.S.; Lowenstine, L.J.; Murphy, B.G. Morphologic, immunohistochemical, and molecular characterization of a novel Lankesterella protozoan in two White’s tree frogs (Litoria caerulea). J. Zoo Wildl. Med. 2010, 41, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Burland, T.G. DNASTAR’s Lasergene Sequence Analysis Software. In Bioinformatics Methods and Protocols; Misener, S., Krawetz, S.A., Eds.; Humana Press: Totowa, NJ, USA, 1999; pp. 71–91. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S.; Gascuel, O. NGPhylogeny.fr: New generation phylogenetic services for non-specialists. Nucleic Acids Res. 2019, 47, W260–W265. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Klaphake, E.; Gibbons, P.M.; Sladky, K.K.; Carpenter, J.W. Chapter 4—Reptiles. In Exotic Animal Formulary, 5th ed.; Carpenter, J.W., Marion, C.J., Eds.; W.B. Saunders: St. Louis, MO, USA, 2018; pp. 81–166. [Google Scholar]
- Lemgruber, L.; Lupetti, P. Crystalloid body, refractile body and virus-like particles in Apicomplexa: What is in there? Parasitology 2012, 139, 285–293. [Google Scholar] [CrossRef]
- Davis, A.K.; Benz, A.C.; Ruyle, L.E.; Kistler, W.M.; Shock, B.C.; Yabsley, M.J. Searching Before It Is Too Late: A Survey of Blood Parasites in Ctenosaura melanosterna, a Critically Endangered Reptile of Honduras (Lankesterella). ISRN Parasitol. 2013, 2013, 495304. [Google Scholar] [CrossRef]
- Paperna, I.; Bastien, P.; Chavatte, J.-M.; Landau, I. Lankesterella poeppigii n. sp. (Apicomplexa, Lankesterellidae) de Bufo poeppigii (Tschudi, 1845) del Perú. Rev. Peru. De Biol. 2009, 16, 165–168. [Google Scholar] [CrossRef]
- Francesco, C.; Origgi, M.T. Immunology of Reptiles. In Infectious Diseases and Pathology of Reptiles; Elliott, R., Jacobson, M.M.G., Eds.; CRC Press: Boca Raton, FL, USA, 2020; Volume 1, pp. 215–266. [Google Scholar]
- Stenberg, P.L.; Bowerman, W.J. Hemoparasites in Oregon spotted frogs (Rana pretiosa) from central Oregon, USA. J. Wildl. Dis. 2008, 44, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Lainson, R.; Shaw, J.J.; Ward, R.D. Schellackia landauae sp.-Nov (Eimeriorina-Lankesterellidae) in Brazilian Lizard Polychrus marmoratus (Iguanidae)—Experimental Transmission by Culex pipiens-Fatigans. Parasitology 1976, 72, 225–243. [Google Scholar] [CrossRef]
- Bristovetzky, M.; Paperna, I. Life-Cycle and Transmission of Schellackia Cf Agamae, a Parasite of the Starred Lizard Agama-Stellio. Int. J. Parasitol. 1990, 20, 883–892. [Google Scholar] [CrossRef]
- Ogedengbe, M.E.; El-Sherry, S.; Ogedengbe, J.D.; Chapman, H.D.; Barta, J.R. Phylogenies based on combined mitochondrial and nuclear sequences conflict with morphologically defined genera in the eimeriid coccidia (Apicomplexa). Int. J. Parasitol. 2018, 48, 59–69. [Google Scholar] [CrossRef]
- JirkŮ, M.; JirkŮ, M.; ObornÍK, M.; LukeŠ, J.; ModrÝ, D. A Model for Taxonomic Work on Homoxenous Coccidia: Redescription, Host Specificity, and Molecular Phylogeny of Eimeria ranae Dobell, 1909, with a Review of Anuran-Host Eimeria (Apicomplexa: Eimeriorina). J. Eukaryot. Microbiol. 2009, 56, 39–51. [Google Scholar] [CrossRef]
- Wozniak, E.J.; Kazacos, K.R.; Telford, S.R.; McLaughlin, G.L. Characterization of the clinical and anatomical pathological changes associated with Hepatozoon mocassini infections in unnatural reptilian hosts. Int. J. Parasitol. 1996, 26, 141–146. [Google Scholar] [CrossRef]
- Baneth, G.; Samish, M.; Shkap, V. Life cycle of Hepatozoon canis (Apicomplexa: Adeleorina: Hepatozoidae) in the tick Rhipicephalus sanguineus and domestic dog (Canis familiaris). J. Parasitol. 2007, 93, 283–299. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items or Status | 1 a | 2 a | 3 a | 4 a | 5 a | 6 a | Reference Interval b |
|---|---|---|---|---|---|---|---|
| Weight (grams) | - | 224.5 | 87.5 | 131.3 | 128.2 | 88.0 | |
| Dehydration | 10–15% | 0–5% | 5–10% | 5–10% | 5–10% | 5–10% | |
| Hemococcidia intensity (/15,000 erythrocytes) | 8 | 6 | 9 | 48 | 0 | 22 | |
| Hepatozoon intensity (/15,000 erythrocytes) | 0 | 2 | 18 | 11 | 10 | 3 | |
| Aspartate aminotransferase | - | 20 | 45 | 60 | 20 | 18 | 0–97 U/L |
| Bile acid | - | <35 | <35 | <35 | 35 | <35 | 2.6–30.3 µmol/L |
| Creatine kinase | - | 2854 | 2790 | - | 613 | 1421 | 174–8768 U/L |
| Uric acid | - | 2.3 | 1 | 1.2 | 1.5 | 0.9 | 0–8.2 mg/dL |
| Glucose | - | 157 | 182 | 374 | 258 | 221 | 169–288 mg/dL |
| Calcium | - | 10.6 | 11.8 | 13.5 | 10.4 | 10.1 | 6–18 mg/dL |
| Phosphate | - | 8.8 | 7.7 | 14.8 | 11.1 | 9.2 | 2.5–21 mg/dL |
| Total protein | - | 7.2 | 8.5 | 9.7 | 7.3 | 8.1 | 4.1–7.4 g/dL |
| Albumin | - | 3.3 | 3 | 3.8 | 3.2 | 3 | 2.1–2.8 g/dL |
| Globulin | - | 3.9 | 5.5 | 6 | 4.1 | 5.1 | 2.5–4.3 g/dL |
| Potassium | - | 3.8 | 2.6 | 4.9 | 5.1 | 3.7 | 1.3–3 mmol/L |
| Sodium | - | 161 | 153 | 167 | 161 | 152 | 158–183 mmol/L |
| Parasite Haplotype | Number | Length a | Width a |
|---|---|---|---|
| Lankesterella desseri n. sp. | 59 | 7.3–15.9 (12.25 ± 1.62) | 1.87–5.49 (3.71 ± 0.84) |
| Hepatozoon gamezi vacuolar gamonts | 28 | 14.40–18.61 (16.55 ± 0.93) | 5.65–9.45 (7.61 ± 0.90) |
| Hepatozoon gamezi basophilic gamonts | 20 | 15.37–18.48 (17.00 ± 1.06) | 5.60–9.95 (7.54 ± 1.79) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, Y.-C.; Lin, T.-S.; Huang, W.-W.; Lee, H.-Y.; Shih, C.-H.; Wu, Y.-C.; Huang, C.-C.; Chen, T.-H. Reevaluation of Hemoparasites in the Black Spiny-Tailed Iguana (Ctenosaura similis) with the First Pathological and Molecular Characterizations of Lankesterella desseri n. sp. and Redescription of Hepatozoon gamezi. Microorganisms 2023, 11, 2374. https://doi.org/10.3390/microorganisms11102374
Chang Y-C, Lin T-S, Huang W-W, Lee H-Y, Shih C-H, Wu Y-C, Huang C-C, Chen T-H. Reevaluation of Hemoparasites in the Black Spiny-Tailed Iguana (Ctenosaura similis) with the First Pathological and Molecular Characterizations of Lankesterella desseri n. sp. and Redescription of Hepatozoon gamezi. Microorganisms. 2023; 11(10):2374. https://doi.org/10.3390/microorganisms11102374
Chicago/Turabian StyleChang, Yen-Chi, Tai-Shen Lin, Wei-Wen Huang, Hung-Yi Lee, Cheng-Hsin Shih, Ying-Chen Wu, Chiu-Chen Huang, and Ter-Hsin Chen. 2023. "Reevaluation of Hemoparasites in the Black Spiny-Tailed Iguana (Ctenosaura similis) with the First Pathological and Molecular Characterizations of Lankesterella desseri n. sp. and Redescription of Hepatozoon gamezi" Microorganisms 11, no. 10: 2374. https://doi.org/10.3390/microorganisms11102374