
Metagenomic Analysis of a Continuous-Flow Aerobic Granulation System for Wastewater Treatment
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Simulated PFR Characteristics and Local Domestic Wastewater
2.2. 16S rDNA Sequencing
2.3. Metagenomic Data Analysis
2.4. Diversity and Compositional Abundance Analysis
2.5. Predicted Functional Analysis
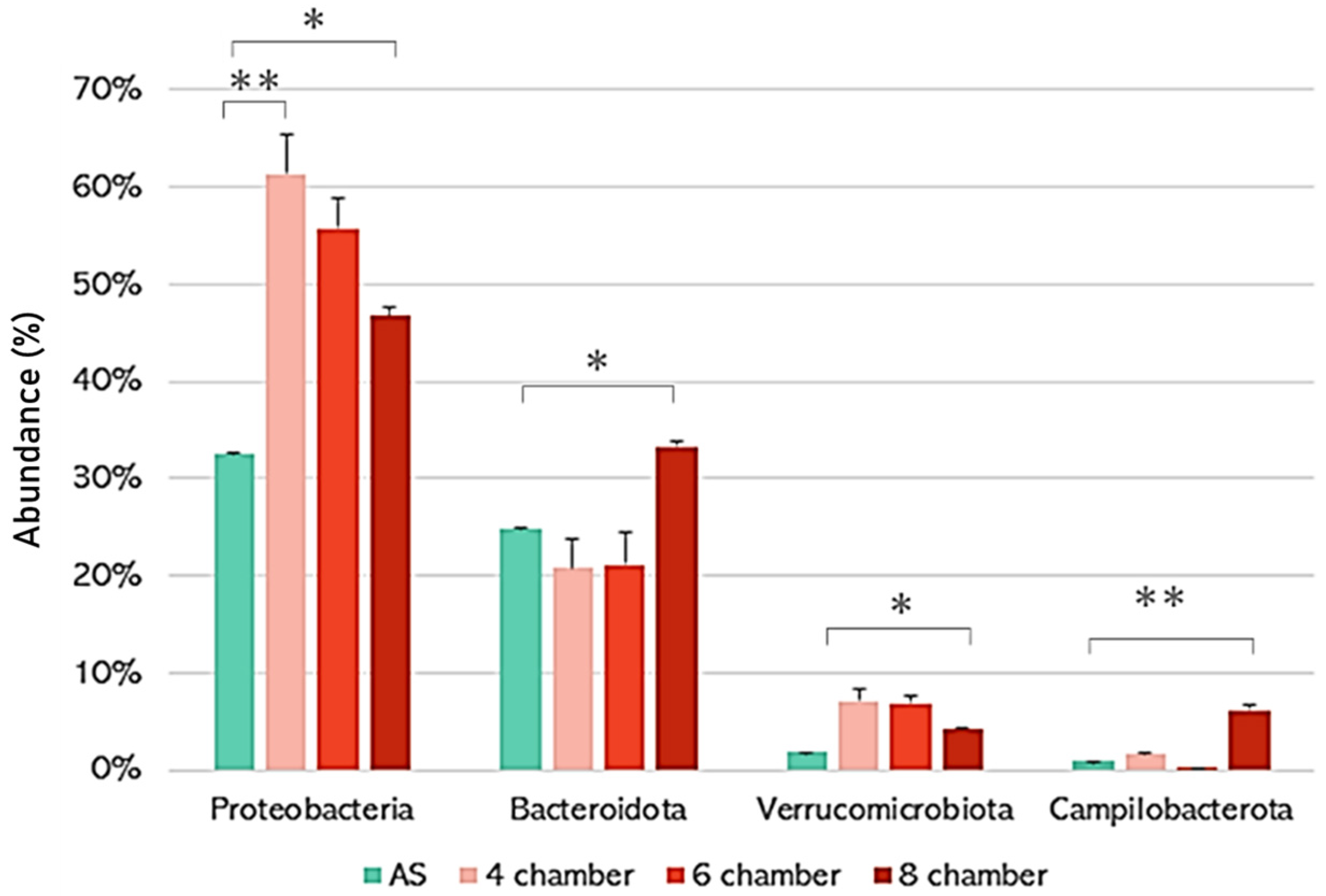
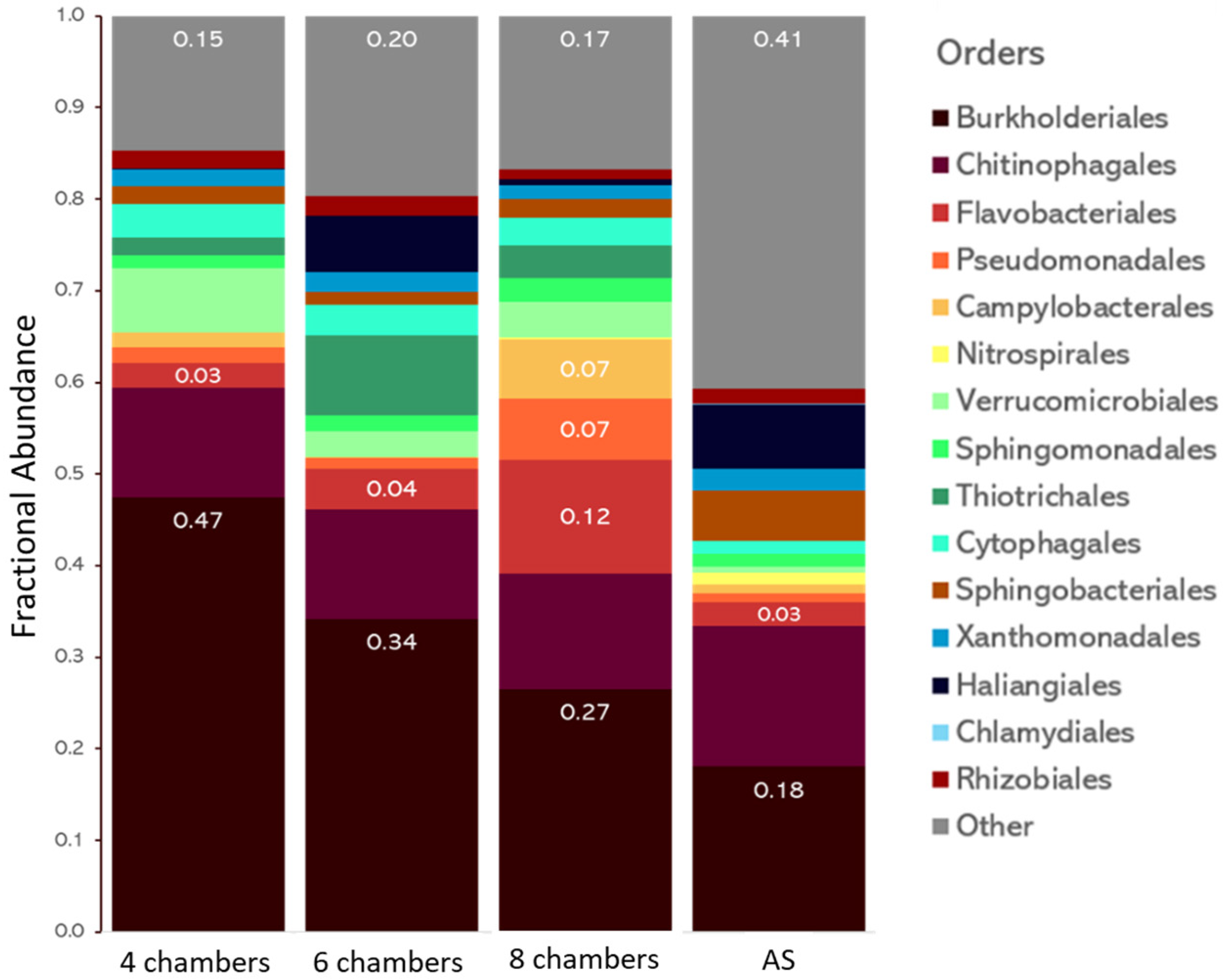
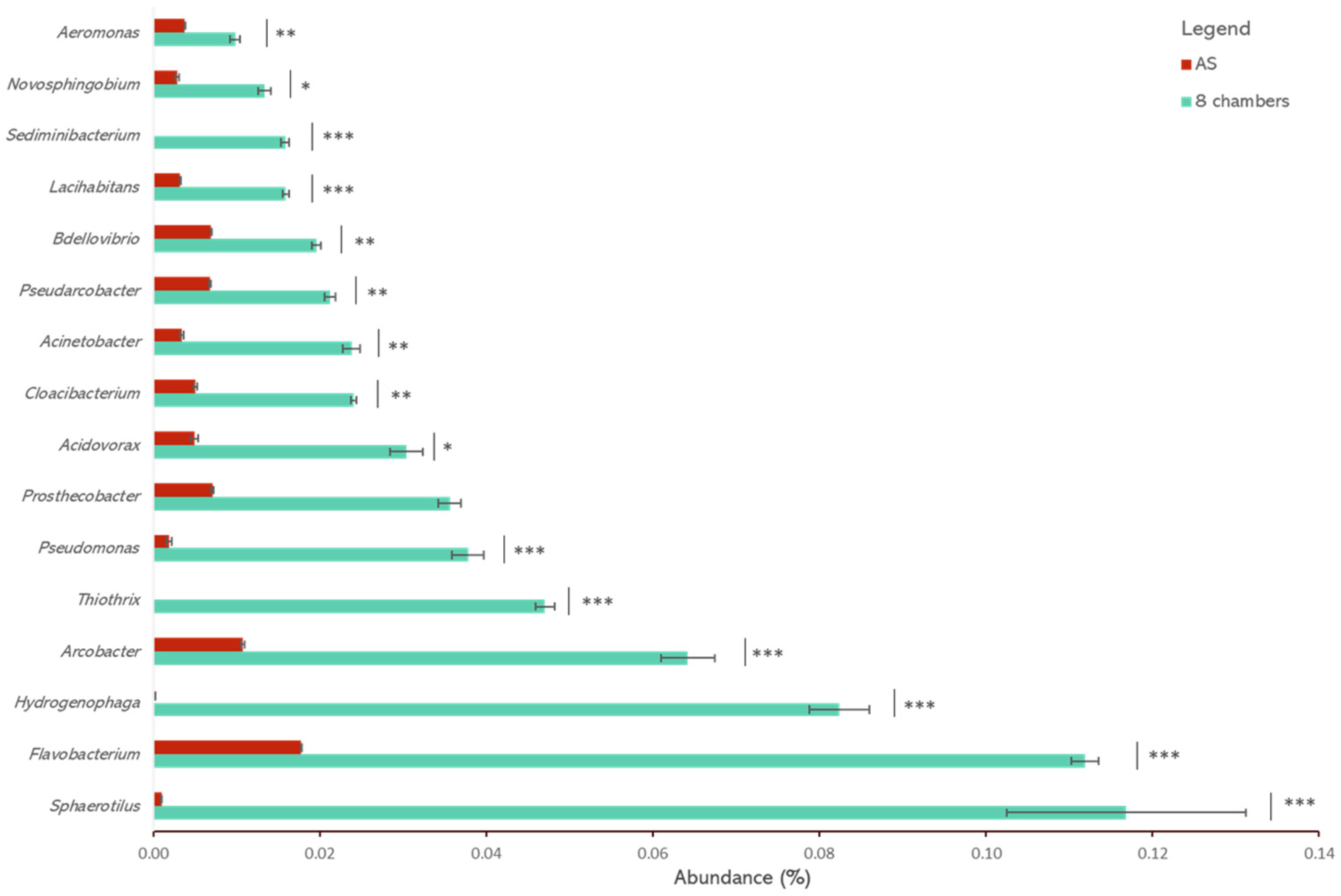
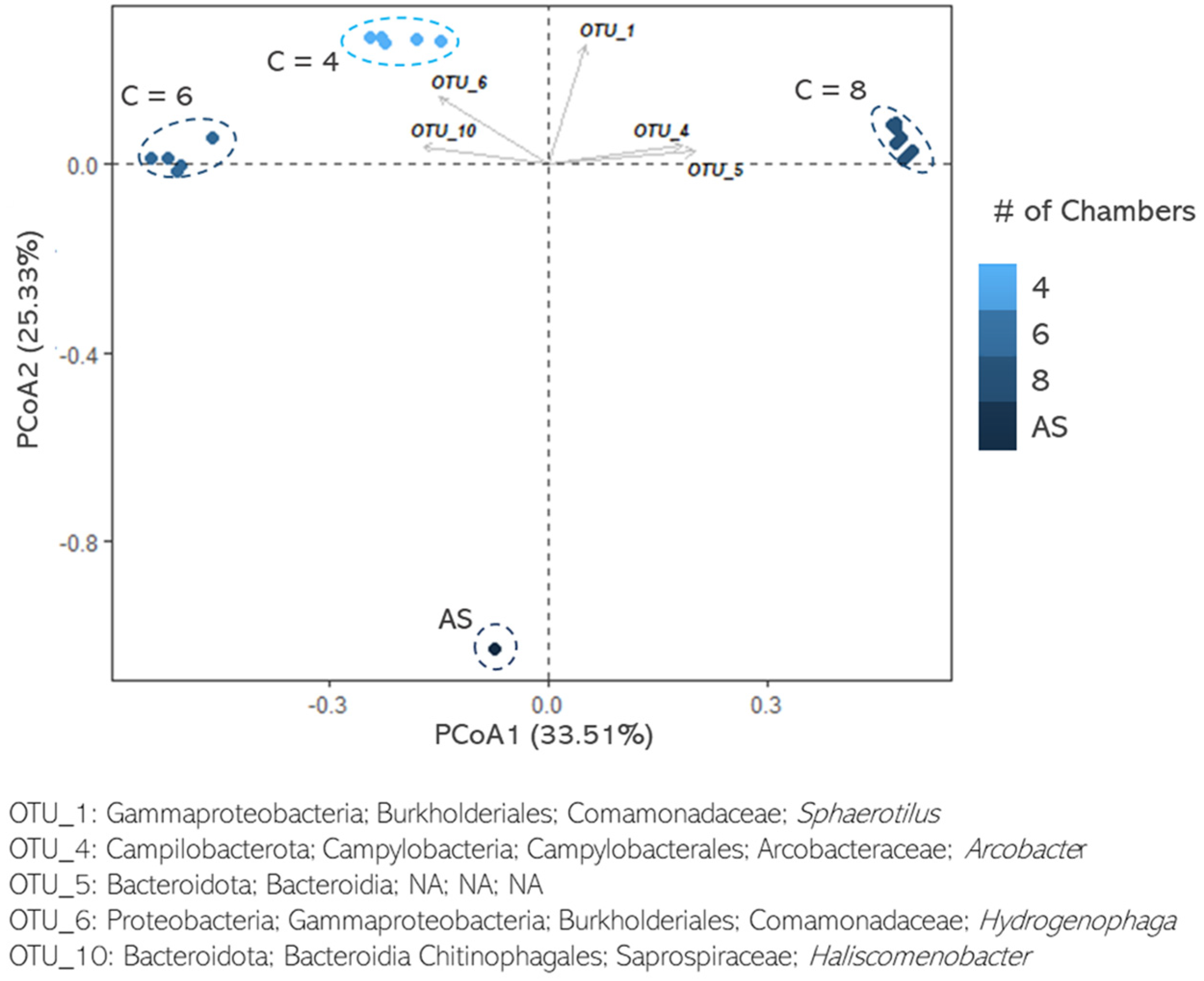
3. Results and Discussion
4. Conclusions, Limitations, and Future Prospects
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kuśnierz, M. Scale of Small Particle Population in Activated Sludge Flocs. Water Air Soil Pollut. 2018, 229, 327. [Google Scholar] [CrossRef] [PubMed]
- Hasebe, Y.; Meguro, H.; Kanai, Y.; Eguchi, M.; Osaka, T.; Tsuneda, S. High-Rate Nitrification of Electronic Industry Wastewater by Using Nitrifying Granules. Water Sci. Technol. 2017, 76, 3171–3180. [Google Scholar] [CrossRef]
- Toh, S.; Tay, J.; Moy, B.; Ivanov, V.; Tay, S. Size-Effect on the Physical Characteristics of the Aerobic Granule in a SBR. Appl. Microbiol. Biotechnol. 2003, 60, 687–695. [Google Scholar] [CrossRef]
- dos Santos, A.L.S.; Galdino, A.C.M.; de Mello, T.P.; de Souza Ramos, L.; Branquinha, M.H.; Bolognese, A.M.; Neto, C.J.; Roudbary, M. What Are the Advantages of Living in a Community? A Microbial Biofilm Perspective! Mem. Inst. Oswaldo Cruz 2018, 113, e180212. [Google Scholar] [CrossRef]
- Sengar, A.; Basheer, F.; Aziz, A.; Farooqi, I.H. Aerobic Granulation Technology: Laboratory Studies to Full Scale Practices. J. Clean. Prod. 2018, 197, 616–632. [Google Scholar] [CrossRef]
- Xia, J.; Ye, L.; Ren, H.; Zhang, X.-X. Microbial Community Structure and Function in Aerobic Granular Sludge. Appl. Microbiol. Biotechnol. 2018, 102, 3967–3979. [Google Scholar] [CrossRef]
- Sarma, S.J.; Tay, J.H. Aerobic Granulation for Future Wastewater Treatment Technology: Challenges Ahead. Environ. Sci. Water Res. Technol. 2018, 4, 9–15. [Google Scholar] [CrossRef]
- Kent, T.R.; Bott, C.B.; Wang, Z.-W. State of the Art of Aerobic Granulation in Continuous Flow Bioreactors. Biotechnol. Adv. 2018, 36, 1139–1166. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Q.-S. Causes and Control of Filamentous Growth in Aerobic Granular Sludge Sequencing Batch Reactors. Biotechnol. Adv. 2006, 24, 115–127. [Google Scholar] [CrossRef]
- López-Palau, S.; Pinto, A.; Basset, N.; Dosta, J.; Mata-Álvarez, J. ORP Slope and Feast–Famine Strategy as the Basis of the Control of a Granular Sequencing Batch Reactor Treating Winery Wastewater. Biochem. Eng. J. 2012, 68, 190–198. [Google Scholar] [CrossRef]
- López–Palau, S.; Dosta, J.; Mata-Álvarez, J. Start-up of an Aerobic Granular Sequencing Batch Reactor for the Treatment of Winery Wastewater. Water Sci. Technol. 2009, 60, 1049–1054. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Angelotti, B.; Brooks, M.; Wang, Z.-W. Feast/Famine Ratio Determined Continuous Flow Aerobic Granulation. Sci. Total Environ. 2021, 750, 141467. [Google Scholar] [CrossRef] [PubMed]
- Cha, L.; Liu, Y.-Q.; Duan, W.; Sternberg, C.E.W.; Yuan, Q.; Chen, F. Fluctuation and Re-Establishment of Aerobic Granules Properties during the Long-Term Operation Period with Low-Strength and Low C/N Ratio Wastewater. Processes 2021, 9, 1290. [Google Scholar] [CrossRef]
- Shrout, J.D.; Tolker-Nielsen, T.; Givskov, M.; Parsek, M.R. The Contribution of Cell-Cell Signaling and Motility to Bacterial Biofilm Formation. MRS Bull. 2011, 36, 367–373. [Google Scholar] [CrossRef]
- Świątczak, P.; Cydzik-Kwiatkowska, A. Performance and Microbial Characteristics of Biomass in a Full-Scale Aerobic Granular Sludge Wastewater Treatment Plant. Environ. Sci. Pollut. Res. 2018, 25, 1655–1669. [Google Scholar] [CrossRef]
- Wilén, B.-M.; Liébana, R.; Persson, F.; Modin, O.; Hermansson, M. The Mechanisms of Granulation of Activated Sludge in Wastewater Treatment, Its Optimization, and Impact on Effluent Quality. Appl. Microbiol. Biotechnol. 2018, 102, 5005–5020. [Google Scholar] [CrossRef] [PubMed]
- Aslam, Z.; Im, W.-T.; Kim, M.K.; Lee, S.-T. Flavobacterium granuli Sp. Nov., Isolated from Granules Used in a Wastewater Treatment Plant. Int. J. Syst. Evol. Microbiol. 2005, 55, 747–751. [Google Scholar] [CrossRef]
- Kim, M.K.; Im, W.-T.; Ohta, H.; Lee, M.; Lee, S.-T. Sphingopyxis granuli Sp. Nov., a Beta-Glucosidase-Producing Bacterium in the Family Sphingomonadaceae in Alpha-4 Subclass of the Proteobacteria. J. Microbiol. 2005, 43, 152–157. [Google Scholar]
- Gómez-Acata, S.; Vital-Jácome, M.; Pérez-Sandoval, M.V.; Navarro-Noya, Y.E.; Thalasso, F.; Luna-Guido, M.; Conde-Barajas, E.; Dendooven, L. Microbial Community Structure in Aerobic and Fluffy Granules Formed in a Sequencing Batch Reactor Supplied with 4-Chlorophenol at Different Settling Times. J. Hazard. Mater. 2018, 342, 606–616. [Google Scholar] [CrossRef]
- Chung, B.S.; Ryu, S.H.; Park, M.; Jeon, Y.; Chung, Y.R.; Jeon, C.O. Hydrogenophaga caeni Sp. Nov., Isolated from Activated Sludge. Int. J. Syst. Evol. Microbiol. 2007, 57, 1126–1130. [Google Scholar] [CrossRef]
- Faizan Khan, M.; Yu, L.; Hollman, J.; Hwa Tay, J.; Achari, G. Integration of Aerobic Granulation and UV/H2O2 Processes in a Continuous Flow System for the Degradation of Sulfolane in Contaminated Water. Environ. Sci. Water Res. Technol. 2020, 6, 1711–1722. [Google Scholar] [CrossRef]
- Müller, E.; Schade, M.; Lemmer, H. Filamentous Scum Bacteria in Activated Sludge Plants: Detection and Identification Quality by Conventional Activated Sludge Microscopy versus Fluorescence in Situ Hybridization. Water Environ. Res. 2007, 79, 2274–2286. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Angelotti, B.; Wang, Z.-W. Continuous-Flow Aerobic Granulation in Plug-Flow Bioreactors Fed with Real Domestic Wastewater. Sci. Total Environ. 2019, 688, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Gomeiz, A.T.; Van Aken, B.; Angelotti, B.; Brooks, M.; Wang, Z.-W. Dynamic Response of Aerobic Granular Sludge to Feast and Famine Conditions in Plug Flow Reactors Fed with Real Domestic Wastewater. Sci. Total Environ. 2021, 758, 144155. [Google Scholar] [CrossRef]
- Yang, B.; Wang, Y.; Qian, P.-Y. Sensitivity and Correlation of Hypervariable Regions in 16S rRNA Genes in Phylogenetic Analysis. BMC Bioinform. 2016, 17, 135. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A Comprehensive Online Resource for Quality Checked and Aligned Ribosomal RNA Sequence Data Compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef]
- Fernandes, A.D.; Macklaim, J.M.; Linn, T.G.; Reid, G.; Gloor, G.B. ANOVA-like Differential Expression (ALDEx) Analysis for Mixed Population RNA-Seq. PLoS ONE 2013, 8, e67019. [Google Scholar] [CrossRef]
- Xu, S.; Yu, G. MicrobiotaProcess: An R Package for Analysis, Visualization and Biomarker Discovery of Microbiome, Bioconductor version: Release (3.12). 2021.
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for Prediction of Metagenome Functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- McNally, C.P.; Eng, A.; Noecker, C.; Gagne-Maynard, W.C.; Borenstein, E. BURRITO: An Interactive Multi-Omic Tool for Visualizing Taxa–Function Relationships in Microbiome Data. Front. Microbiol. 2018, 9, 365. [Google Scholar] [CrossRef] [PubMed]
- de Kreuk, M.K.; van Loosdrecht, M.C.M. Selection of Slow Growing Organisms as a Means for Improving Aerobic Granular Sludge Stability. Water Sci. Technol. 2004, 49, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.Z.; Mondal, P.K.; Sabir, S. Aerobic Granulation for Wastewater Bioremediation: A Review. Can. J. Chem. Eng. 2013, 91, 1045–1058. [Google Scholar] [CrossRef]
- Guo, F.; Zhang, S.-H.; Yu, X.; Wei, B. Variations of Both Bacterial Community and Extracellular Polymers: The Inducements of Increase of Cell Hydrophobicity from Biofloc to Aerobic Granule Sludge. Bioresour. Technol. 2011, 102, 6421–6428. [Google Scholar] [CrossRef]
- Hamza, R.A.S.E. Development of Upflow Aerobic Granular Sludge Bioreactor (UAGSBR) for Treatment of High-Strength Organic Wastewater. Ph.D. Thesis, University of Calgargy, Calgary, AB, Canada, 2019; p. 242. [Google Scholar]
- Lee, D.-J.; Chen, Y.-Y.; Show, K.-Y.; Whiteley, C.G.; Tay, J.-H. Advances in Aerobic Granule Formation and Granule Stability in the Course of Storage and Reactor Operation. Biotechnol. Adv. 2010, 28, 919–934. [Google Scholar] [CrossRef]
- Ji, B.; Zhang, M.; Gu, J.; Ma, Y.; Liu, Y. A Self-Sustaining Synergetic Microalgal-Bacterial Granular Sludge Process towards Energy-Efficient and Environmentally Sustainable Municipal Wastewater Treatment. Water Res. 2020, 179, 115884. [Google Scholar] [CrossRef]
- Richard, M.; Hao, O.; Jenkins, D. Growth Kinetics of Sphaerotilus Species and Their Significance in Activated Sludge Bulking. Water Pollut. Control Fed. 1985, 57, 68–81. [Google Scholar]
- Weissbrodt, D.G.; Lochmatter, S.; Ebrahimi, S.; Rossi, P.; Maillard, J.; Holliger, C. Bacterial Selection during the Formation of Early-Stage Aerobic Granules in Wastewater Treatment Systems Operated Under Wash-Out Dynamics. Front. Microbiol. 2012, 3, 332. [Google Scholar] [CrossRef]
- Krasowska, A.; Sigler, K. How Microorganisms Use Hydrophobicity and What Does This Mean for Human Needs? Front. Cell Infect. Microbiol. 2014, 4, 112. [Google Scholar] [CrossRef]
- Vu, B.; Chen, M.; Crawford, R.J.; Ivanova, E.P. Bacterial Extracellular Polysaccharides Involved in Biofilm Formation. Molecules 2009, 14, 2535–2554. [Google Scholar] [CrossRef] [PubMed]
- de Graaff, D.R.; van Loosdrecht, M.C.M.; Pronk, M. Stable Granulation of Seawater-Adapted Aerobic Granular Sludge with Filamentous Thiothrix Bacteria. Water Res. 2020, 175, 115683. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Choi, E.; Yun, Z.; Park, Y.K. Microbial Structure and Community of RBC Biofilm Removing Nitrate and Phosphorus from Domestic Wastewater. J. Microbiol. Biotechnol. 2008, 18, 1459–1469. [Google Scholar]
- Tang, C.-J.; Zheng, P.; Wang, C.-H.; Mahmood, Q.; Zhang, J.-Q.; Chen, X.-G.; Zhang, L.; Chen, J.-W. Performance of High-Loaded ANAMMOX UASB Reactors Containing Granular Sludge. Water Res. 2011, 45, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ding, L.-B.; Cai, A.; Huang, G.-X.; Horn, H. Aerobic Sludge Granulation in a Full-Scale Sequencing Batch Reactor. BioMed Res. Int. 2014, 2014, 268789. [Google Scholar] [CrossRef]
- Nouha, K.; Kumar, R.S.; Tyagi, R.D. Heavy Metals Removal from Wastewater Using Extracellular Polymeric Substances Produced by Cloacibacterium Normanense in Wastewater Sludge Supplemented with Crude Glycerol and Study of Extracellular Polymeric Substances Extraction by Different Methods. Bioresour. Technol. 2016, 212, 120–129. [Google Scholar] [CrossRef]
- Williams, H.N.; Baer, M.L.; Tudor, J.J. Bdellovibrio. In Bergey’s Manual of Systematics of Archaea and Bacteria; American Cancer Society: Hoboken, NJ, USA, 2015; pp. 1–22. ISBN 978-1-118-96060-8. [Google Scholar]
- Kämpfer, P.; Witzenberger, R.; Denner, E.B.M.; Busse, H.-J.; Neef, A. Novosphingobium hassiacum Sp. Nov., a New Species Isolated from an Aerated Sewage Pond. Syst. Appl. Microbiol. 2002, 25, 37–45. [Google Scholar] [CrossRef]
- Wongwongsee, W.; Chareanpat, P.; Pinyakong, O. Abilities and Genes for PAH Biodegradation of Bacteria Isolated from Mangrove Sediments from the Central of Thailand. Mar. Pollut. Bull. 2013, 74, 95–104. [Google Scholar] [CrossRef]
- Winkler, M.-K.H.; Kleerebezem, R.; de Bruin, L.M.M.; Verheijen, P.J.T.; Abbas, B.; Habermacher, J.; van Loosdrecht, M.C.M. Microbial Diversity Differences within Aerobic Granular Sludge and Activated Sludge Flocs. Appl. Microbiol. Biotechnol. 2013, 97, 7447–7458. [Google Scholar] [CrossRef]
- Chinh, T.T.; Hieu, P.D.; Cuong, B.V.; Linh, N.N.; Lan, N.N.; Nguyen, N.S.; Hung, N.Q.; Hien, L.T.T. Sequencing Batch Reactor and Bacterial Community in Aerobic Granular Sludge for Wastewater Treatment of Noodle-Manufacturing Sector. Appl. Sci. 2018, 8, 509. [Google Scholar] [CrossRef]
- Zhang, D.; Li, W.; Hou, C.; Shen, J.; Jiang, X.; Sun, X.; Li, J.; Han, W.; Wang, L.; Liu, X. Aerobic Granulation Accelerated by Biochar for the Treatment of Refractory Wastewater. Chem. Eng. J. 2017, 314, 88–97. [Google Scholar] [CrossRef]
- Liu, C.; Sun, D.; Zhu, J.; Liu, W. Two-Component Signal Transduction Systems: A Major Strategy for Connecting Input Stimuli to Biofilm Formation. Front. Microbiol. 2019, 9, 3279. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.K. Precedence for the Structural Role of Flagella in Biofilms. mBio 2013, 4, e00225-13. [Google Scholar] [CrossRef]
- Hao, T.; Mackey, H.R.; Guo, G.; Liu, R.; Chen, G. Resilience of Sulfate-Reducing Granular Sludge against Temperature, pH, Oxygen, Nitrite, and Free Nitrous Acid. Appl. Microbiol. Biotechnol. 2016, 100, 8563–8572. [Google Scholar] [CrossRef]
- Feng, Y.; Zhao, Y.; Guo, Y.; Liu, S. Microbial Transcript and Metabolome Analysis Uncover Discrepant Metabolic Pathways in Autotrophic and Mixotrophic Anammox Consortia. Water Res. 2018, 128, 402–411. [Google Scholar] [CrossRef]
- Li, W.-M.; Liao, X.-W.; Guo, J.-S.; Zhang, Y.-X.; Chen, Y.-P.; Fang, F.; Yan, P. New Insights into Filamentous Sludge Bulking: The Potential Role of Extracellular Polymeric Substances in Sludge Bulking in the Activated Sludge Process. Chemosphere 2020, 248, 126012. [Google Scholar] [CrossRef]
- Liang, J.; Wang, Q.; Li, J.; Guo, S.; Ke, M.; Gamal El-Din, M.; Chen, C. Effects of Anaerobic Granular Sludge towards the Treatment of Flowback Water in an Up-Flow Anaerobic Sludge Blanket Bioreactor: Comparison between Mesophilic and Thermophilic Conditions. Bioresour. Technol. 2021, 326, 124784. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Maratukulam, P.D.; Lum, A.M.; Clark, D.S.; Keasling, J.D. Metabolic Engineering of an Aerobic Sulfate Reduction Pathway and Its Application to Precipitation of Cadmium on the Cell Surface. Appl. Environ. Microbiol. 2000, 66, 4497–4502. [Google Scholar] [CrossRef]
- Liu, C.; Huang, H.; Duan, X.; Chen, Y. Integrated Metagenomic and Metaproteomic Analyses Unravel Ammonia Toxicity to Active Methanogens and Syntrophs, Enzyme Synthesis, and Key Enzymes in Anaerobic Digestion. Environ. Sci. Technol. 2021, 55, 14817–14827. [Google Scholar] [CrossRef]
- Rubio Gomez, M.A.; Ibba, M. Aminoacyl-tRNA Synthetases. RNA 2020, 26, 910–936. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Shannon Diversity Index (H) | Evenness (E) | Good’s Coverage | Simpson Index (D) | Chao 1 | Number of Phylotypes or OTUs | |
|---|---|---|---|---|---|---|
| 4-chamber PFR (n = 6) a | 4.74 ± 0.09 | 0.78 ± 0.01 | 0.9961 | 0.972 | 456 | 436 |
| 6-chamber PFR (n = 8) | 4.52 ± 0.27 | 0.77 ± 0.02 | 0.9970 | 0.965 | 377 | 362 |
| 8-chamber PFR (n = 8) | 5.43 ± 0.20 | 0.80 ± 0.02 | 0.9996 | 0.985 | 906 | 896 |
| Activated Sludge (n = 2) | 6.28 ± 0.03 | 0.87 ± 0.00 | 0.9996 | 0.996 | 1324 | 1313 |
| Nitrifying (n = 1) | 3.45 | 0.60 | 1.0000 | 0.899 | 320 | 319 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomeiz, A.T.; Sun, Y.; Newborn, A.; Wang, Z.-W.; Angelotti, B.; Van Aken, B. Metagenomic Analysis of a Continuous-Flow Aerobic Granulation System for Wastewater Treatment. Microorganisms 2023, 11, 2328. https://doi.org/10.3390/microorganisms11092328
Gomeiz AT, Sun Y, Newborn A, Wang Z-W, Angelotti B, Van Aken B. Metagenomic Analysis of a Continuous-Flow Aerobic Granulation System for Wastewater Treatment. Microorganisms. 2023; 11(9):2328. https://doi.org/10.3390/microorganisms11092328
Chicago/Turabian StyleGomeiz, Alison T., Yewei Sun, Aaron Newborn, Zhi-Wu Wang, Bob Angelotti, and Benoit Van Aken. 2023. "Metagenomic Analysis of a Continuous-Flow Aerobic Granulation System for Wastewater Treatment" Microorganisms 11, no. 9: 2328. https://doi.org/10.3390/microorganisms11092328