
Tackling Drug-Resistant Tuberculosis: New Challenges from the Old Pathogen Mycobacterium tuberculosis

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Epidemiology of Tuberculosis
3. Insights into Pathogenesis of Tuberculosis
4. Effects of SARS-CoV-2 Infection on Latent TB
5. Treatment of LTBI and TB Disease
6. The Complexity of Drug Resistance in M.tb
7. New Insights into the Treatment of MDR-TB
8. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shuaib, Y.A.; Utpatel, C.; Kohl, T.A.; Barilar, I.; Diricks, M.; Ashraf, N.; Wieler, L.H.; Kerubo, G.; Mesfin, E.A.; Diallo, A.B.; et al. Origin and Global Expansion of Mycobacterium tuberculosis Complex Lineage 3. Genes 2022, 13, 990. [Google Scholar] [CrossRef]
- Coscolla, M.; Gagneux, S.; Menardo, F.; Loiseau, C.; Ruiz-Rodriguez, P.; Borrell, S.; Otchere, I.D.; Asante-Poku, A.; Asare, P.; Sanchez-Buso, L.; et al. Phylogenomics of Mycobacterium africanum reveals a new lineage and a complex evolutionary history. Microb. Genom. 2021, 7, 000477. [Google Scholar] [CrossRef]
- Silva, M.L.; Ca, B.; Osorio, N.S.; Rodrigues, P.N.S.; Maceiras, A.R.; Saraiva, M. Tuberculosis caused by Mycobacterium africanum: Knowns and unknowns. PLoS Pathog. 2022, 18, e1010490. [Google Scholar] [CrossRef] [PubMed]
- Kanabalan, R.D.; Lee, L.J.; Lee, T.Y.; Chong, P.P.; Hassan, L.; Ismail, R.; Chin, V.K. Human tuberculosis and Mycobacterium tuberculosis complex: A review on genetic diversity, pathogenesis and omics approaches in host biomarkers discovery. Microbiol. Res. 2021, 246, 126674. [Google Scholar] [CrossRef]
- Alexander, K.A.; Laver, P.N.; Michel, A.L.; Williams, M.; van Helden, P.D.; Warren, R.M.; Gey van Pittius, N.C. Novel Mycobacterium tuberculosis complex pathogen, M. mungi. Emerg. Infect. Dis. 2010, 16, 1296–1299. [Google Scholar] [CrossRef] [PubMed]
- Van Ingen, J.; Rahim, Z.; Mulder, A.; Boeree, M.J.; Simeone, R.; Brosch, R.; van Soolingen, D. Characterization of Mycobacterium orygis as M. tuberculosis complex subspecies. Emerg. Infect. Dis. 2012, 18, 653–655. [Google Scholar] [CrossRef]
- Ghodbane, R.; Drancourt, M. Non-human sources of Mycobacterium tuberculosis. Tuberculosis 2013, 93, 589–595. [Google Scholar] [CrossRef]
- Grange, J.M. The biology of the genus Mycobacterium. Soc. Appl. Bacteriol. Symp. Ser. 1996, 25, 1S–9S. [Google Scholar] [CrossRef]
- Phillips, C.J.; Foster, C.R.; Morris, P.A.; Teverson, R. The transmission of Mycobacterium bovis infection to cattle. Res. Vet. Sci. 2003, 74, 1–15. [Google Scholar] [CrossRef]
- Somoskovi, A.; Dormandy, J.; Mayrer, A.R.; Carter, M.; Hooper, N.; Salfinger, M. “Mycobacterium canettii” isolated from a human immunodeficiency virus-positive patient: First case recognized in the United States. J. Clin. Microbiol. 2009, 47, 255–257. [Google Scholar] [CrossRef] [PubMed]
- Comas, I.; Gagneux, S. A role for systems epidemiology in tuberculosis research. Trends Microbiol. 2011, 19, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Abascal, E.; Genestet, C.; Valera, A.; Herranz, M.; Martinez-Lirola, M.; Munoz, P.; Dumitrescu, O.; Garcia de Viedma, D. Assessment of closely related Mycobacterium tuberculosis variants with different transmission success and in vitro infection dynamics. Sci. Rep. 2021, 11, 11041. [Google Scholar] [CrossRef] [PubMed]
- Saelens, J.W.; Viswanathan, G.; Tobin, D.M. Mycobacterial Evolution Intersects With Host Tolerance. Front. Immunol. 2019, 10, 528. [Google Scholar] [CrossRef]
- Ma, J.; Vongpradith, A.; Ledesma, J.R.; Novotney, A.; Yi, S.; Lim, K.; Hay, S.I.; Murray, C.J.L.; Kyu, H.H. Progress towards the 2020 milestones of the end TB strategy in Cambodia: Estimates of age and sex specific TB incidence and mortality from the Global Burden of Disease Study 2019. BMC Infect. Dis. 2022, 22, 904. [Google Scholar] [CrossRef]
- Fukunaga, R.; Glaziou, P.; Harris, J.B.; Date, A.; Floyd, K.; Kasaeva, T. Epidemiology of Tuberculosis and Progress Toward Meeting Global Targets—Worldwide, 2019. MMWR. Morb. Mortal. Wkly. Rep. 2021, 70, 427–430. [Google Scholar] [CrossRef]
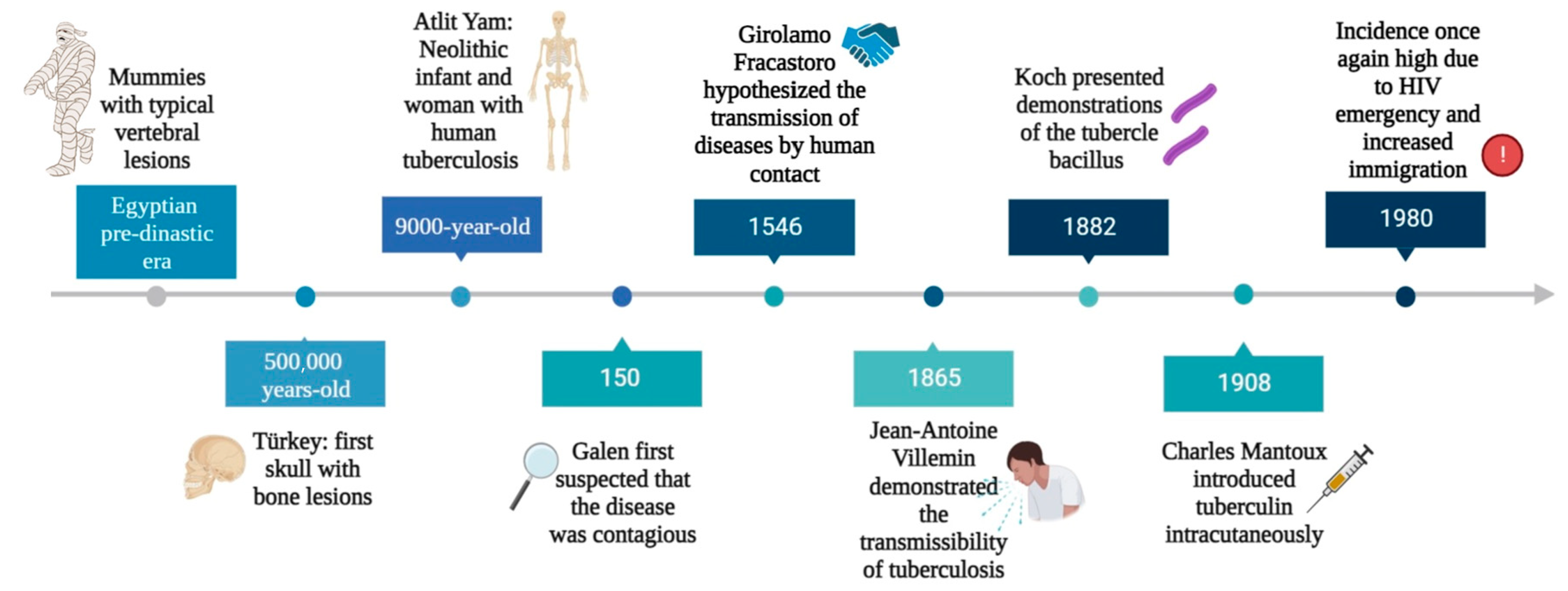
- Barberis, I.; Bragazzi, N.L.; Galluzzo, L.; Martini, M. The history of tuberculosis: From the first historical records to the isolation of Koch’s bacillus. J. Prev. Med. Hyg. 2017, 58, E9–E12. [Google Scholar]
- Buzic, I.; Giuffra, V. The paleopathological evidence on the origins of human tuberculosis: A review. J. Prev. Med. Hyg. 2020, 61, E3–E8. [Google Scholar] [CrossRef]
- Hershkovitz, I.; Donoghue, H.D.; Minnikin, D.E.; Besra, G.S.; Lee, O.Y.; Gernaey, A.M.; Galili, E.; Eshed, V.; Greenblatt, C.L.; Lemma, E.; et al. Detection and molecular characterization of 9000-year-old Mycobacterium tuberculosis from a Neolithic settlement in the Eastern Mediterranean. PLoS ONE 2008, 3, e3426. [Google Scholar] [CrossRef]
- Holloway, K.L.; Link, K.; Ruhli, F.; Henneberg, M. Skeletal lesions in human tuberculosis may sometimes heal: An aid to palaeopathological diagnoses. PLoS ONE 2013, 8, e62798. [Google Scholar] [CrossRef] [PubMed]
- McDonald, S.K.; Matisoo-Smith, E.A.; Buckley, H.R.; Walter, R.K.; Aung, H.L.; Collins, C.J.; Cook, G.M.; Kardailsky, O.; Krause, J.; Knapp, M. ‘TB or not TB’: The conundrum of pre-European contact tuberculosis in the Pacific. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2020, 375, 20190583. [Google Scholar] [CrossRef]
- Ojo, O.O.; Nadarajah, S.; Kebe, M. Integer time series models for tuberculosis in Africa. Sci. Rep. 2023, 13, 11443. [Google Scholar] [CrossRef]
- Cambau, E.; Drancourt, M. Steps towards the discovery of Mycobacterium tuberculosis by Robert Koch, 1882. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2014, 20, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Daniel, V.S.; Daniel, T.M. Old Testament biblical references to tuberculosis. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 1999, 29, 1557–1558. [Google Scholar] [CrossRef] [PubMed]
- Pesapane, F.; Marcelli, S.; Nazzaro, G. Hieronymi Fracastorii: The Italian scientist who described the “French disease”. An. Bras. De Dermatol. 2015, 90, 684–686. [Google Scholar] [CrossRef] [PubMed]
- Oren, E.; McDermid, J.M. Tuberculosis. In Nutrition and Health in a Developing World; de Pee, S., Taren, D., Bloem, M., Eds.; Humana Press: Cham, Switzerland, 2017. [Google Scholar]
- Sandhu, G.K. Tuberculosis: Current situation, challenges and overview of its control programs in India. J. Glob. Infect. Dis. 2011, 3, 143–150. [Google Scholar] [CrossRef]
- Riva, M.A. From milk to rifampicin and back again: History of failures and successes in the treatment for tuberculosis. J. Antibiot. 2014, 67, 661–665. [Google Scholar] [CrossRef]
- Kestler, B.; Tyler, S.K. Latent tuberculosis testing through the ages: The search for a sleeping killer. Am. J. Physiol. Lung Cell. Mol. Physiol. 2022, 322, L412–L419. [Google Scholar] [CrossRef]
- Yang, H.; Kruh-Garcia, N.A.; Dobos, K.M. Purified protein derivatives of tuberculin--past, present, and future. FEMS Immunol. Med. Microbiol. 2012, 66, 273–280. [Google Scholar] [CrossRef]
- Dutour, O.; Colombo, A.; Coqueugniot, H. Was the rise of TB contemporaneous with the industrial revolution? Epidemiological evolution of TB in France (17th–20th centuries) inferred from osteoarchaeological and historical archives. Int. J. Paleopathol. 2021, 34, 130–133. [Google Scholar] [CrossRef]
- Tripp, L.; Sawchuk, L.A. Insights into secular trends of respiratory tuberculosis: The 20th century Maltese experience. PLoS ONE 2017, 12, e0183296. [Google Scholar] [CrossRef]
- Al-Humadi, H.W.; Al-Saigh, R.J.; Al-Humadi, A.W. Addressing the Challenges of Tuberculosis: A Brief Historical Account. Front. Pharmacol. 2017, 8, 689. [Google Scholar] [CrossRef]
- Davies, P.D. The world-wide increase in tuberculosis: How demographic changes, HIV infection and increasing numbers in poverty are increasing tuberculosis. Ann. Med. 2003, 35, 235–243. [Google Scholar] [CrossRef]
- Cummings, K.J. Tuberculosis control: Challenges of an ancient and ongoing epidemic. Public Health Rep. 2007, 122, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Borgdorff, M.W.; van Soolingen, D. The re-emergence of tuberculosis: What have we learnt from molecular epidemiology? Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2013, 19, 889–901. [Google Scholar] [CrossRef]
- Porter, J.D.; McAdam, K.P. The re-emergence of tuberculosis. Annu. Rev. Public Health 1994, 15, 303–323. [Google Scholar] [CrossRef]
- Rossi, G. The re-emergence of tuberculosis: The endemia of multiple-antibiotic-resistant strains. Recent. Progress. Med. 1996, 87, 487–488. [Google Scholar]
- Vall Mayans, M.; Maguire, A.; Miret, M.; Alcaide, J.; Parron, I.; Casabona, J. The spread of AIDS and the re-emergence of tuberculosis in Catalonia, Spain. Aids 1997, 11, 499–505. [Google Scholar] [CrossRef]
- Soko, R.N.; Burke, R.M.; Feasey, H.R.A.; Sibande, W.; Nliwasa, M.; Henrion, M.Y.R.; Khundi, M.; Dodd, P.J.; Ku, C.C.; Kawalazira, G.; et al. Effects of Coronavirus Disease Pandemic on Tuberculosis Notifications, Malawi. Emerg. Infect. Dis. 2021, 27, 1831–1839. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, J.R.; Boccia, D.; Evans, C.A.; Adato, M.; Petticrew, M.; Porter, J.D. The social determinants of tuberculosis: From evidence to action. Am. J. Public Health 2011, 101, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Zwerling, A.; Hanrahan, C.; Dowdy, D.W. Ancient Disease, Modern Epidemiology: A Century of Progress in Understanding and Fighting Tuberculosis. Am. J. Epidemiol. 2016, 183, 407–414. [Google Scholar] [CrossRef]
- Barry, C.E. Lessons from seven decades of antituberculosis drug discovery. Curr. Top. Med. Chem. 2011, 11, 1216–1225. [Google Scholar] [CrossRef]
- Cegielski, J.P. Extensively drug-resistant tuberculosis: “There must be some kind of way out of here”. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2010, 50 (Suppl. 3), S195–S200. [Google Scholar] [CrossRef]
- Yang, Q.; Han, J.; Shen, J.; Peng, X.; Zhou, L.; Yin, X. Diagnosis and treatment of tuberculosis in adults with HIV. Medicine 2022, 101, e30405. [Google Scholar] [CrossRef]
- Glaziou, P.; Floyd, K.; Raviglione, M.C. Global Epidemiology of Tuberculosis. Semin. Respir. Crit. Care Med. 2018, 39, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Chakaya, J.; Petersen, E.; Nantanda, R.; Mungai, B.N.; Migliori, G.B.; Amanullah, F.; Lungu, P.; Ntoumi, F.; Kumarasamy, N.; Maeurer, M.; et al. The WHO Global Tuberculosis 2021 Report—Not so good news and turning the tide back to End TB. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2022, 124 (Suppl. 1), S26–S29. [Google Scholar] [CrossRef]
- Hamada, Y.; Getahun, H.; Tadesse, B.T.; Ford, N. HIV-associated tuberculosis. Int. J. STD AIDS 2021, 32, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Hu, X.; Huang, Y.; Huang, X.; Tong, Y. Editorial: Updates on tuberculosis control and management. Front. Public Health 2022, 10, 1126429. [Google Scholar] [CrossRef]
- Bloom, B.R.; Atun, R.; Cohen, T.; Dye, C.; Fraser, H.; Gomez, G.B.; Knight, G.; Murray, M.; Nardell, E.; Rubin, E.; et al. Tuberculosis. In Major Infectious Diseases, 3rd ed.; Holmes, K.K., Bertozzi, S., Bloom, B.R., Jha, P., Eds.; The International Bank for Reconstruction and Development/The World Bank: Washington, DC, USA, 2017; p. 11. [Google Scholar]
- Chakaya, J.; Khan, M.; Ntoumi, F.; Aklillu, E.; Fatima, R.; Mwaba, P.; Kapata, N.; Mfinanga, S.; Hasnain, S.E.; Katoto, P.; et al. Global Tuberculosis Report 2020—Reflections on the Global TB burden, treatment and prevention efforts. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2021, 113 (Suppl. 1), S7–S12. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.G.; Chung, J.H. Diagnosis and treatment of multidrug-resistant tuberculosis. Yeungnam Univ. J. Med. 2020, 37, 277–285. [Google Scholar] [CrossRef]
- WHO. W.H.O. Tuberculosis; WHO: Geneva, Switzerland, 2023. [Google Scholar]
- Li, T.; Du, X.; Kang, J.; Luo, D.; Liu, X.; Zhao, Y. Patient, Diagnosis, and Treatment Delays Among Tuberculosis Patients Before and During COVID-19 Epidemic—China, 2018–2022. China CDC Wkly. 2023, 5, 259–265. [Google Scholar] [CrossRef]
- Dheda, K.; Perumal, T.; Moultrie, H.; Perumal, R.; Esmail, A.; Scott, A.J.; Udwadia, Z.; Chang, K.C.; Peter, J.; Pooran, A.; et al. The intersecting pandemics of tuberculosis and COVID-19: Population-level and patient-level impact, clinical presentation, and corrective interventions. Lancet. Respir. Med. 2022, 10, 603–622. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Tuberculosis Deaths and Disease Increase during the COVID-19 Pandemic; Global Tuberculosis Report 2022; WHO: Geneva, Switzerland, 2022. [Google Scholar]
- Dean, A.S.; Tosas Auguet, O.; Glaziou, P.; Zignol, M.; Ismail, N.; Kasaeva, T.; Floyd, K. 25 years of surveillance of drug-resistant tuberculosis: Achievements, challenges, and way forward. Lancet. Infect. Dis. 2022, 22, e191–e196. [Google Scholar] [CrossRef] [PubMed]
- Kostyukova, I.; Pasechnik, O.; Mokrousov, I. Epidemiology and Drug Resistance Patterns of Mycobacterium tuberculosis in High-Burden Area in Western Siberia, Russia. Microorganisms 2023, 11, 425. [Google Scholar] [CrossRef]
- Khabibullina, N.F.; Kutuzova, D.M.; Burmistrova, I.A.; Lyadova, I.V. The Biological and Clinical Aspects of a Latent Tuberculosis Infection. Trop. Med. Infect. Dis. 2022, 7, 48. [Google Scholar] [CrossRef]
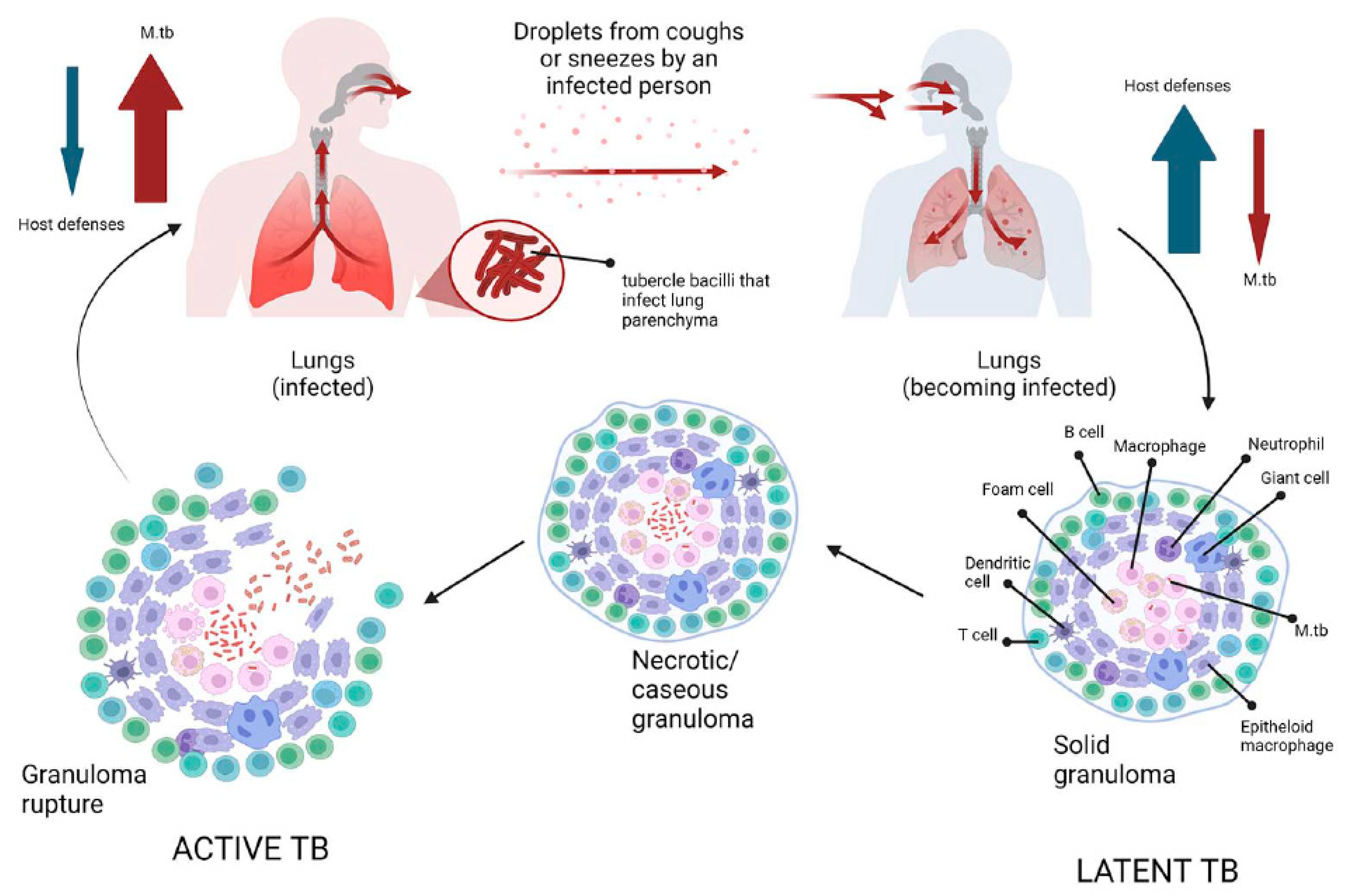
- Behr, M.A.; Edelstein, P.H.; Ramakrishnan, L. Revisiting the timetable of tuberculosis. Bmj 2018, 362, k2738. [Google Scholar] [CrossRef] [PubMed]
- Pawlowski, A.; Jansson, M.; Skold, M.; Rottenberg, M.E.; Kallenius, G. Tuberculosis and HIV co-infection. PLoS Pathog. 2012, 8, e1002464. [Google Scholar] [CrossRef]
- Patterson, B.; Wood, R. Is cough really necessary for TB transmission? Tuberculosis 2019, 117, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Ryndak, M.B.; Laal, S. Mycobacterium tuberculosis Primary Infection and Dissemination: A Critical Role for Alveolar Epithelial Cells. Front. Cell. Infect. Microbiol. 2019, 9, 299. [Google Scholar] [CrossRef]
- Nardell, E.A. Transmission and Institutional Infection Control of Tuberculosis. Cold Spring Harb. Perspect. Med. 2015, 6, a018192. [Google Scholar] [CrossRef]
- Delogu, G.; Sali, M.; Fadda, G. The biology of mycobacterium tuberculosis infection. Mediterr. J. Hematol. Infect. Dis. 2013, 5, e2013070. [Google Scholar] [CrossRef]
- Chandra, P.; Grigsby, S.J.; Philips, J.A. Immune evasion and provocation by Mycobacterium tuberculosis. Nat. Rev. Microbiol. 2022, 20, 750–766. [Google Scholar] [CrossRef]
- Smith, I. Mycobacterium tuberculosis pathogenesis and molecular determinants of virulence. Clin. Microbiol. Rev. 2003, 16, 463–496. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.; Rani, A.; Alam, A.; Zarin, S.; Pandey, S.; Singh, H.; Hasnain, S.E.; Ehtesham, N.Z. Macrophage: A Cell With Many Faces and Functions in Tuberculosis. Front. Immunol. 2022, 13, 747799. [Google Scholar] [CrossRef]
- Cohen, S.B.; Gern, B.H.; Delahaye, J.L.; Adams, K.N.; Plumlee, C.R.; Winkler, J.K.; Sherman, D.R.; Gerner, M.Y.; Urdahl, K.B. Alveolar Macrophages Provide an Early Mycobacterium tuberculosis Niche and Initiate Dissemination. Cell Host Microbe 2018, 24, 439–446.e434. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Singh, V.K.; Hunter, R.L.; Jagannath, C. Macrophage heterogeneity and plasticity in tuberculosis. J. Leukoc. Biol. 2019, 106, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Behar, S.M.; Martin, C.J.; Booty, M.G.; Nishimura, T.; Zhao, X.; Gan, H.X.; Divangahi, M.; Remold, H.G. Apoptosis is an innate defense function of macrophages against Mycobacterium tuberculosis. Mucosal Immunol. 2011, 4, 279–287. [Google Scholar] [CrossRef]
- Martin, C.J.; Booty, M.G.; Rosebrock, T.R.; Nunes-Alves, C.; Desjardins, D.M.; Keren, I.; Fortune, S.M.; Remold, H.G.; Behar, S.M. Efferocytosis is an innate antibacterial mechanism. Cell Host Microbe 2012, 12, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Bengoa, M.; Meurer, M.; Goethe, R.; Singh, M.; Reljic, R.; von Kockritz-Blickwede, M. Role of phagocyte extracellular traps during Mycobacterium tuberculosis infections and tuberculosis disease processes. Front. Microbiol. 2023, 14, 983299. [Google Scholar] [CrossRef]
- Ravesloot-Chavez, M.M.; Van Dis, E.; Stanley, S.A. The Innate Immune Response to Mycobacterium tuberculosis Infection. Annu. Rev. Immunol. 2021, 39, 611–637. [Google Scholar] [CrossRef]
- Cooper, A.M. Cell-mediated immune responses in tuberculosis. Annu. Rev. Immunol. 2009, 27, 393–422. [Google Scholar] [CrossRef]
- Esmail, H.; Barry, C.E., 3rd; Wilkinson, R.J. Understanding latent tuberculosis: The key to improved diagnostic and novel treatment strategies. Drug Discov. Today 2012, 17, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Gengenbacher, M.; Kaufmann, S.H. Mycobacterium tuberculosis: Success through dormancy. FEMS Microbiol. Rev. 2012, 36, 514–532. [Google Scholar] [CrossRef]
- Petersen, H.J.; Smith, A.M. The role of the innate immune system in granulomatous disorders. Front. Immunol. 2013, 4, 120. [Google Scholar] [CrossRef] [PubMed]
- Alsayed, S.S.R.; Gunosewoyo, H. Tuberculosis: Pathogenesis, Current Treatment Regimens and New Drug Targets. Int. J. Mol. Sci. 2023, 24, 5202. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Wainwright, H.C.; Locketz, M.; Bekker, L.G.; Walther, G.B.; Dittrich, C.; Visser, A.; Wang, W.; Hsu, F.F.; Wiehart, U.; et al. Caseation of human tuberculosis granulomas correlates with elevated host lipid metabolism. EMBO Mol. Med. 2010, 2, 258–274. [Google Scholar] [CrossRef]
- Ulrichs, T.; Kosmiadi, G.A.; Trusov, V.; Jorg, S.; Pradl, L.; Titukhina, M.; Mishenko, V.; Gushina, N.; Kaufmann, S.H. Human tuberculous granulomas induce peripheral lymphoid follicle-like structures to orchestrate local host defence in the lung. J. Pathol. 2004, 204, 217–228. [Google Scholar] [CrossRef]
- Lee, O.Y.; Wu, H.H.; Donoghue, H.D.; Spigelman, M.; Greenblatt, C.L.; Bull, I.D.; Rothschild, B.M.; Martin, L.D.; Minnikin, D.E.; Besra, G.S. Mycobacterium tuberculosis complex lipid virulence factors preserved in the 17,000-year-old skeleton of an extinct bison, Bison antiquus. PLoS ONE 2012, 7, e41923. [Google Scholar] [CrossRef]
- Forrellad, M.A.; Klepp, L.I.; Gioffre, A.; Sabio y Garcia, J.; Morbidoni, H.R.; de la Paz Santangelo, M.; Cataldi, A.A.; Bigi, F. Virulence factors of the Mycobacterium tuberculosis complex. Virulence 2013, 4, 3–66. [Google Scholar] [CrossRef]
- Biondo, C.; Midiri, A.; Gerace, E.; Zummo, S.; Mancuso, G. SARS-CoV-2 Infection in Patients with Cystic Fibrosis: What We Know So Far. Life 2022, 12, 2087. [Google Scholar] [CrossRef]
- Biondo, C.; Ponzo, E.; Midiri, A.; Ostone, G.B.; Mancuso, G. The Dark Side of Nosocomial Infections in Critically Ill COVID-19 Patients. Life 2023, 13, 1408. [Google Scholar] [CrossRef]
- Pathak, L.; Gayan, S.; Pal, B.; Talukdar, J.; Bhuyan, S.; Sandhya, S.; Yeger, H.; Baishya, D.; Das, B. Coronavirus Activates an Altruistic Stem Cell-Mediated Defense Mechanism that Reactivates Dormant Tuberculosis: Implications in Coronavirus Disease 2019 Pandemic. Am. J. Pathol. 2021, 191, 1255–1268. [Google Scholar] [CrossRef]
- Hildebrand, R.E.; Chandrasekar, S.S.; Riel, M.; Touray, B.J.B.; Aschenbroich, S.A.; Talaat, A.M. Superinfection with SARS-CoV-2 Has Deleterious Effects on Mycobacterium bovis BCG Immunity and Promotes Dissemination of Mycobacterium tuberculosis. Microbiol. Spectr. 2022, 10, e0307522. [Google Scholar] [CrossRef]
- Group, T.C.-G.S. Tuberculosis and COVID-19 co-infection: Description of the global cohort. Eur. Respir. J. 2022, 59, 2102538. [Google Scholar] [CrossRef]
- Luke, E.; Swafford, K.; Shirazi, G.; Venketaraman, V. TB and COVID-19: An Exploration of the Characteristics and Resulting Complications of Co-infection. Front. Biosci. 2022, 14, 6. [Google Scholar] [CrossRef]
- Oh, C.E.; Menzies, D. Four months of rifampicin monotherapy for latent tuberculosis infection in children. Clin. Exp. Pediatr. 2022, 65, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Sterling, T.R.; Njie, G.; Zenner, D.; Cohn, D.L.; Reves, R.; Ahmed, A.; Menzies, D.; Horsburgh, C.R., Jr.; Crane, C.M.; Burgos, M.; et al. Guidelines for the Treatment of Latent Tuberculosis Infection: Recommendations from the National Tuberculosis Controllers Association and CDC, 2020. MMWR. Recomm. Rep. Morb. Mortal. Wkly. Rep. Recomm. Rep. 2020, 69, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Assefa, D.G.; Bedru, A.; Zeleke, E.D.; Negash, S.E.; Debela, D.T.; Molla, W.; Mengistu, N.; Woldesenbet, T.T.; Bedane, N.F.; Kajogoo, V.D.; et al. Efficacy and safety of different regimens in the treatment of patients with latent tuberculosis infection: A systematic review and network meta-analysis of randomized controlled trials. Arch. Public Health = Arch. Belg. De Sante Publique 2023, 81, 82. [Google Scholar] [CrossRef]
- WHO Consolidated Guidelines on Tuberculosis: Module 4: Treatment—Drug-Resistant Tuberculosis Treatment, 2022 Update; WHO Guidelines Approved by the Guidelines Review Committee: Geneva, Switzerland, 2022.
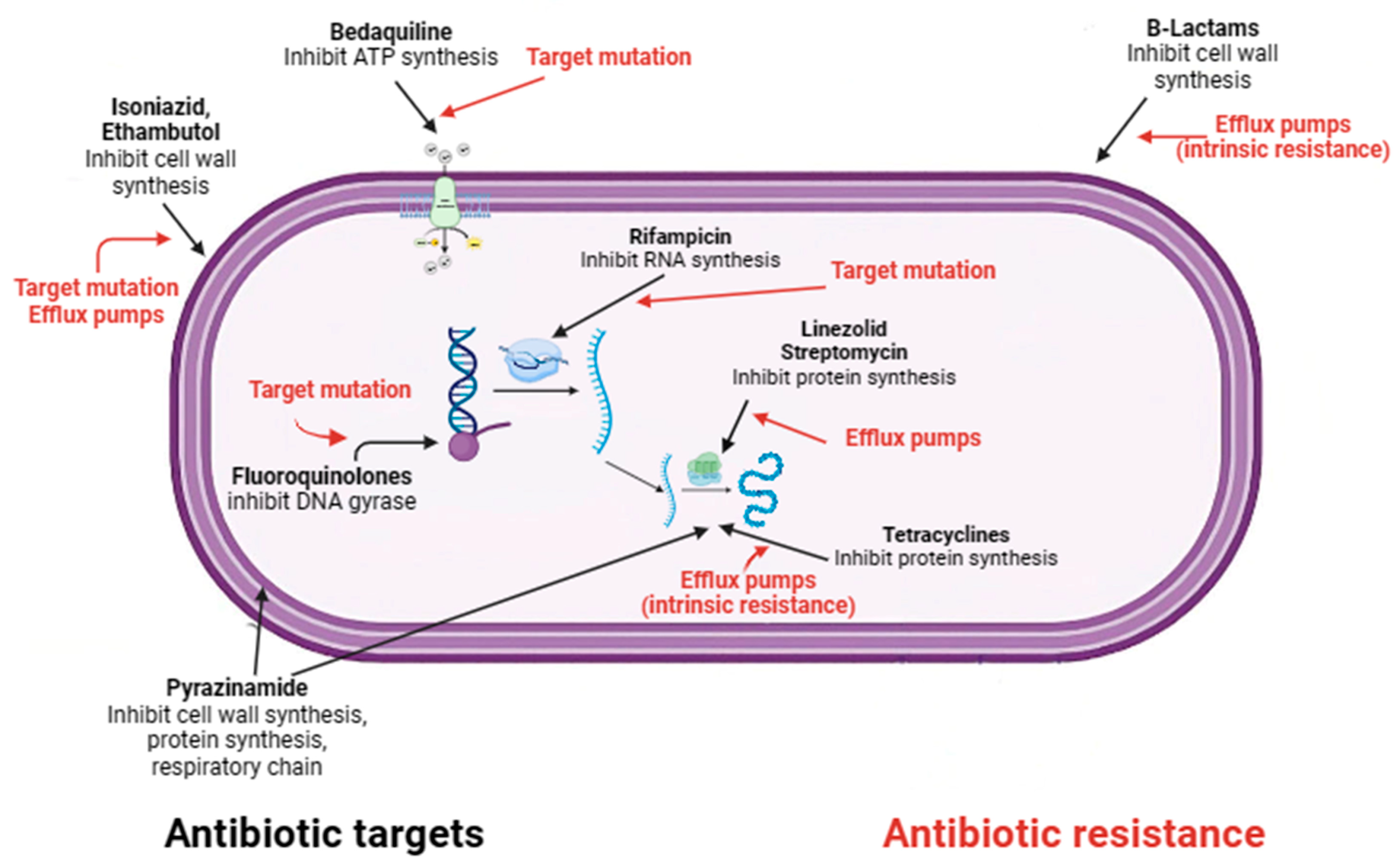
- Dartois, V.A.; Rubin, E.J. Anti-tuberculosis treatment strategies and drug development: Challenges and priorities. Nat. Rev.. Microbiol. 2022, 20, 685–701. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.R.; Manialawy, Y.; Siraki, A.G. Isoniazid and host immune system interactions: A proposal for a novel comprehensive mode of action. Br. J. Pharmacol. 2019, 176, 4599–4608. [Google Scholar] [CrossRef]
- Campbell, E.A.; Korzheva, N.; Mustaev, A.; Murakami, K.; Nair, S.; Goldfarb, A.; Darst, S.A. Structural mechanism for rifampicin inhibition of bacterial rna polymerase. Cell 2001, 104, 901–912. [Google Scholar] [CrossRef]
- Gopal, P.; Gruber, G.; Dartois, V.; Dick, T. Pharmacological and Molecular Mechanisms Behind the Sterilizing Activity of Pyrazinamide. Trends Pharmacol. Sci. 2019, 40, 930–940. [Google Scholar] [CrossRef]
- Batt, S.M.; Burke, C.E.; Moorey, A.R.; Besra, G.S. Antibiotics and resistance: The two-sided coin of the mycobacterial cell wall. Cell Surf. 2020, 6, 100044. [Google Scholar] [CrossRef]
- Occhineri, S.; Matucci, T.; Rindi, L.; Tiseo, G.; Falcone, M.; Riccardi, N.; Besozzi, G. Pretomanid for tuberculosis treatment: An update for clinical purposes. Curr. Res. Pharmacol. Drug Discov. 2022, 3, 100128. [Google Scholar] [CrossRef] [PubMed]
- Bagcchi, S. WHO’s Global Tuberculosis Report 2022. Lancet. Microbe 2023, 4, e20. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, K.; Ahmad, R.; Sinha, S.; Dutta, S.; Haque, M. Multidrug-Resistant TB (MDR-TB) and Extensively Drug-Resistant TB (XDR-TB) Among Children: Where We Stand Now. Cureus 2023, 15, e35154. [Google Scholar] [CrossRef]
- Espinosa-Pereiro, J.; Sanchez-Montalva, A.; Aznar, M.L.; Espiau, M. MDR Tuberculosis Treatment. Medicina 2022, 58, 188. [Google Scholar] [CrossRef] [PubMed]
- Kherabi, Y.; Frechet-Jachym, M.; Rioux, C.; Yazdanpanah, Y.; Mechai, F.; Pourcher, V.; Robert, J.; Guglielmetti, L.; Group, M.-T.M. Revised Definitions of Tuberculosis Resistance and Treatment Outcomes, France, 2006-2019. Emerg. Infect. Dis. 2022, 28, 1796–1804. [Google Scholar] [CrossRef]
- Swain, S.S.; Sharma, D.; Hussain, T.; Pati, S. Molecular mechanisms of underlying genetic factors and associated mutations for drug resistance in Mycobacterium tuberculosis. Emerg. Microbes Infect. 2020, 9, 1651–1663. [Google Scholar] [CrossRef]
- Gygli, S.M.; Borrell, S.; Trauner, A.; Gagneux, S. Antimicrobial resistance in Mycobacterium tuberculosis: Mechanistic and evolutionary perspectives. FEMS Microbiol. Rev. 2017, 41, 354–373. [Google Scholar] [CrossRef] [PubMed]
- Allue-Guardia, A.; Garcia, J.I.; Torrelles, J.B. Evolution of Drug-Resistant Mycobacterium tuberculosis Strains and Their Adaptation to the Human Lung Environment. Front. Microbiol. 2021, 12, 612675. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Guo, H.; Chen, T.; Yu, L.; Chen, Y.; Zhao, J.; Yan, H.; Chen, M.; Sun, Q.; Zhang, C.; et al. Genome-wide SNP and InDel mutations in Mycobacterium tuberculosis associated with rifampicin and isoniazid resistance. Int. J. Clin. Exp. Pathol. 2018, 11, 3903–3914. [Google Scholar]
- Biondo, C. Bacterial Antibiotic Resistance: The Most Critical Pathogens. Pathogens 2023, 12, 116. [Google Scholar] [CrossRef]
- Bakhtiyariniya, P.; Khosravi, A.D.; Hashemzadeh, M.; Savari, M. Detection and characterization of mutations in genes related to isoniazid resistance in Mycobacterium tuberculosis clinical isolates from Iran. Mol. Biol. Rep. 2022, 49, 6135–6143. [Google Scholar] [CrossRef]
- Jian, Z.; Zeng, L.; Xu, T.; Sun, S.; Yan, S.; Yang, L.; Huang, Y.; Jia, J.; Dou, T. Antibiotic resistance genes in bacteria: Occurrence, spread, and control. J. Basic Microbiol. 2021, 61, 1049–1070. [Google Scholar] [CrossRef] [PubMed]
- Huemer, M.; Mairpady Shambat, S.; Brugger, S.D.; Zinkernagel, A.S. Antibiotic resistance and persistence-Implications for human health and treatment perspectives. EMBO Rep. 2020, 21, e51034. [Google Scholar] [CrossRef] [PubMed]
- Urbaniec, J.; Xu, Y.; Hu, Y.; Hingley-Wilson, S.; McFadden, J. Phenotypic heterogeneity in persisters: A novel ‘hunker’ theory of persistence. FEMS Microbiol. Rev. 2022, 46, fuab042. [Google Scholar] [CrossRef] [PubMed]
- Lanni, A.; Iacobino, A.; Fattorini, L.; Giannoni, F. Eradication of Drug-Tolerant Mycobacterium tuberculosis 2022: Where We Stand. Microorganisms 2023, 11, 1511. [Google Scholar] [CrossRef] [PubMed]
- Al-Saeedi, M.; Al-Hajoj, S. Diversity and evolution of drug resistance mechanisms in Mycobacterium tuberculosis. Infect. Drug Resist. 2017, 10, 333–342. [Google Scholar] [CrossRef]
- Jones, R.M.; Adams, K.N.; Eldesouky, H.E.; Sherman, D.R. The evolving biology of Mycobacterium tuberculosis drug resistance. Front. Cell. Infect. Microbiol. 2022, 12, 1027394. [Google Scholar] [CrossRef]
- Hassan, S.T.S.; Sudomova, M.; Berchova-Bimova, K.; Gowrishankar, S.; Rengasamy, K.R.R. Antimycobacterial, Enzyme Inhibition, and Molecular Interaction Studies of Psoromic Acid in Mycobacterium tuberculosis: Efficacy and Safety Investigations. J. Clin. Med. 2018, 7, 226. [Google Scholar] [CrossRef]
- Sudomova, M.; Shariati, M.A.; Echeverria, J.; Berindan-Neagoe, I.; Nabavi, S.M.; Hassan, S.T.S. A Microbiological, Toxicological, and Biochemical Study of the Effects of Fucoxanthin, a Marine Carotenoid, on Mycobacterium tuberculosis and the Enzymes Implicated in Its Cell Wall: A Link Between Mycobacterial Infection and Autoimmune Diseases. Mar. Drugs 2019, 17, 641. [Google Scholar] [CrossRef] [PubMed]
- Vanino, E.; Granozzi, B.; Akkerman, O.W.; Munoz-Torrico, M.; Palmieri, F.; Seaworth, B.; Tiberi, S.; Tadolini, M. Update of drug-resistant tuberculosis treatment guidelines: A turning point. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2023, 130 (Suppl. 1), S12–S15. [Google Scholar] [CrossRef] [PubMed]
- Conradie, F.; Bagdasaryan, T.R.; Borisov, S.; Howell, P.; Mikiashvili, L.; Ngubane, N.; Samoilova, A.; Skornykova, S.; Tudor, E.; Variava, E.; et al. Bedaquiline-Pretomanid-Linezolid Regimens for Drug-Resistant Tuberculosis. N. Engl. J. Med. 2022, 387, 810–823. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mancuso, G.; Midiri, A.; De Gaetano, S.; Ponzo, E.; Biondo, C. Tackling Drug-Resistant Tuberculosis: New Challenges from the Old Pathogen Mycobacterium tuberculosis. Microorganisms 2023, 11, 2277. https://doi.org/10.3390/microorganisms11092277
Mancuso G, Midiri A, De Gaetano S, Ponzo E, Biondo C. Tackling Drug-Resistant Tuberculosis: New Challenges from the Old Pathogen Mycobacterium tuberculosis. Microorganisms. 2023; 11(9):2277. https://doi.org/10.3390/microorganisms11092277
Chicago/Turabian StyleMancuso, Giuseppe, Angelina Midiri, Silvia De Gaetano, Elena Ponzo, and Carmelo Biondo. 2023. "Tackling Drug-Resistant Tuberculosis: New Challenges from the Old Pathogen Mycobacterium tuberculosis" Microorganisms 11, no. 9: 2277. https://doi.org/10.3390/microorganisms11092277