
Unraveling the Diverse Profile of N-Acyl Homoserine Lactone Signals and Their Role in the Regulation of Biofilm Formation in Porphyra haitanensis-Associated Pseudoalteromonas galatheae

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
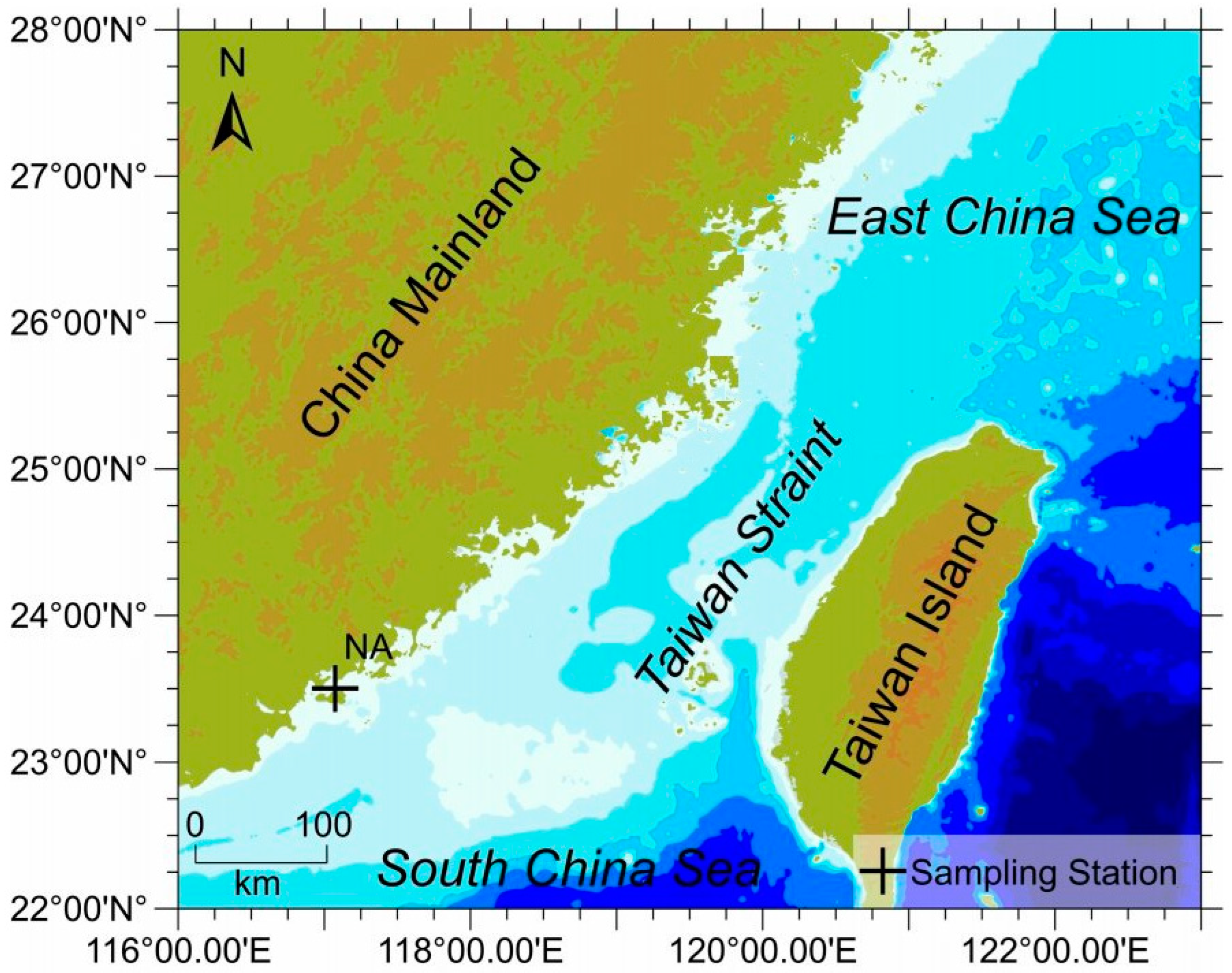
2.1. Sample Collection
2.2. Isolation of gDNA and 16S rRNA Gene Amplification
2.3. Bacterial Identification and Phylogenetical Analysis
2.4. Screening for AHL Production
2.5. Isolation and Characterization of AHLs
2.6. Identification and Characterization of AHLs by LC–MS
2.7. Biofilm formation Assay
2.7.1. Ninety-Six-Well Microtiter Plate Crystal Violet Biofilm Formation Assay
2.7.2. Scanning Electron Microscopy Assay
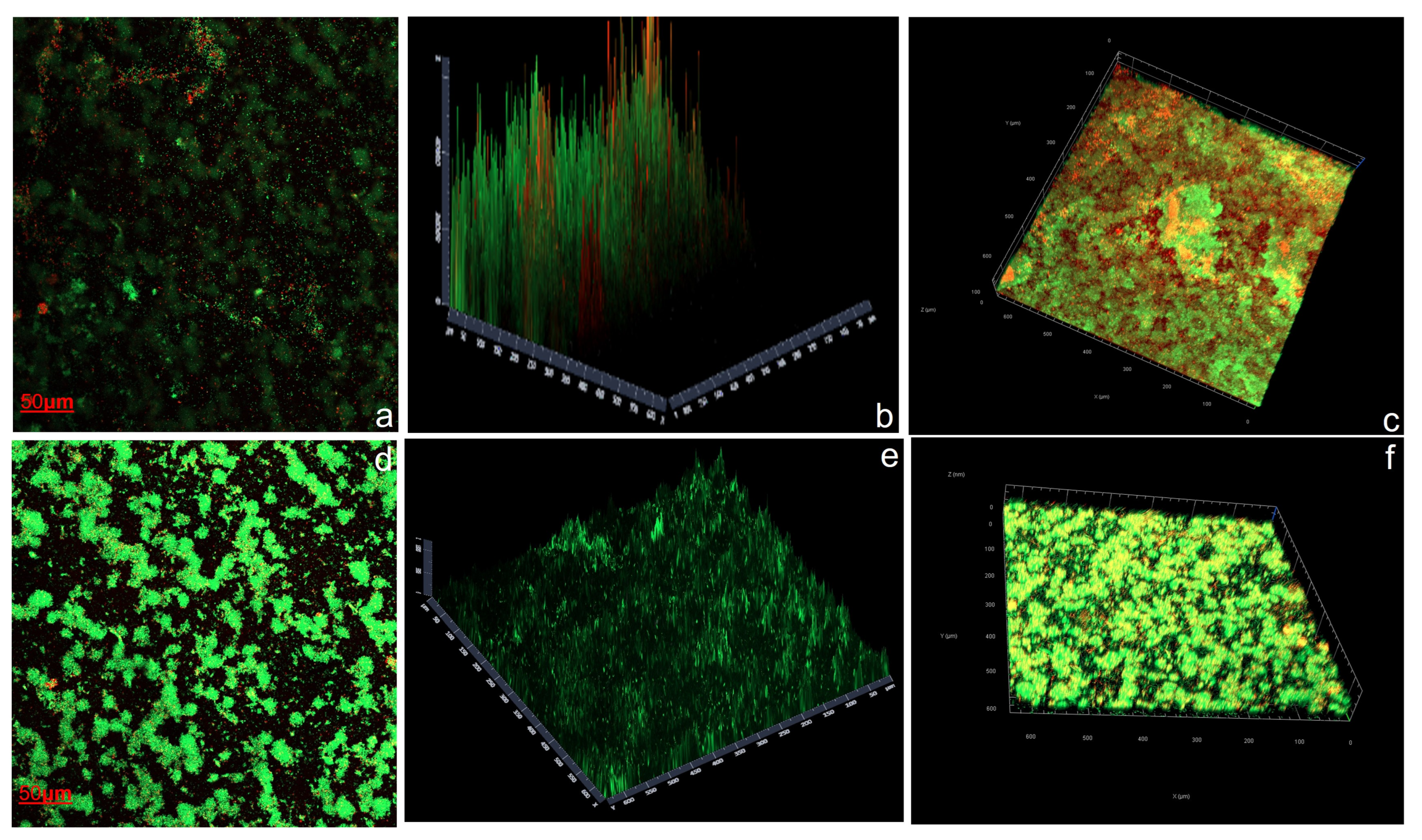
2.7.3. Confocal Laser Scanning Microscopy (CLSM) Assay
- Biomass (µm3/µm2): The volume of biomass per unit area, estimated as the volume of all voxels that contain biomass divided by the substratum area; COMSTAT 2 counts as biomass all voxels above a given threshold.
- Average thickness (biomass) (µm): Measuring/considering only the area covered by the biomass.
- Average thickness (entire area) (µm): Measurement of the complete extent of the stack.
- Maximum thickness (µm): Measurement of the highest point of the biofilm relative to the substratum.
- Roughness coefficient, Ra (nondimensional): Quantifying the height variability of the biofilm.
- Surface area (µm2): The sum of the areas of all visible biomass voxel surfaces against the background, including the area occupied in each layer (µm2), considering biomass pixels in each layer (confocal slices).
2.8. Statistical Analysis
3. Results
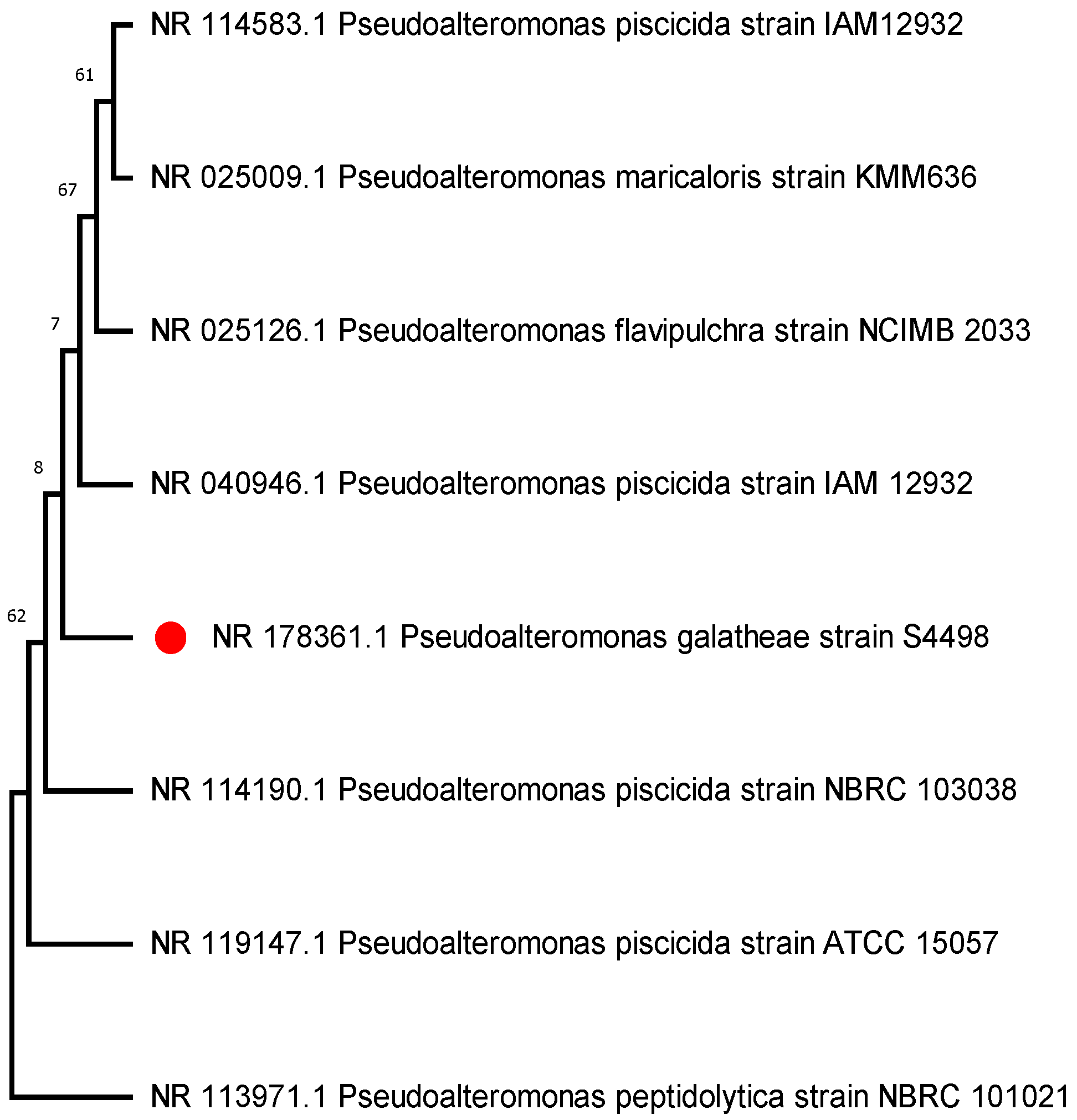
3.1. Identification and Phylogenetic Analysis
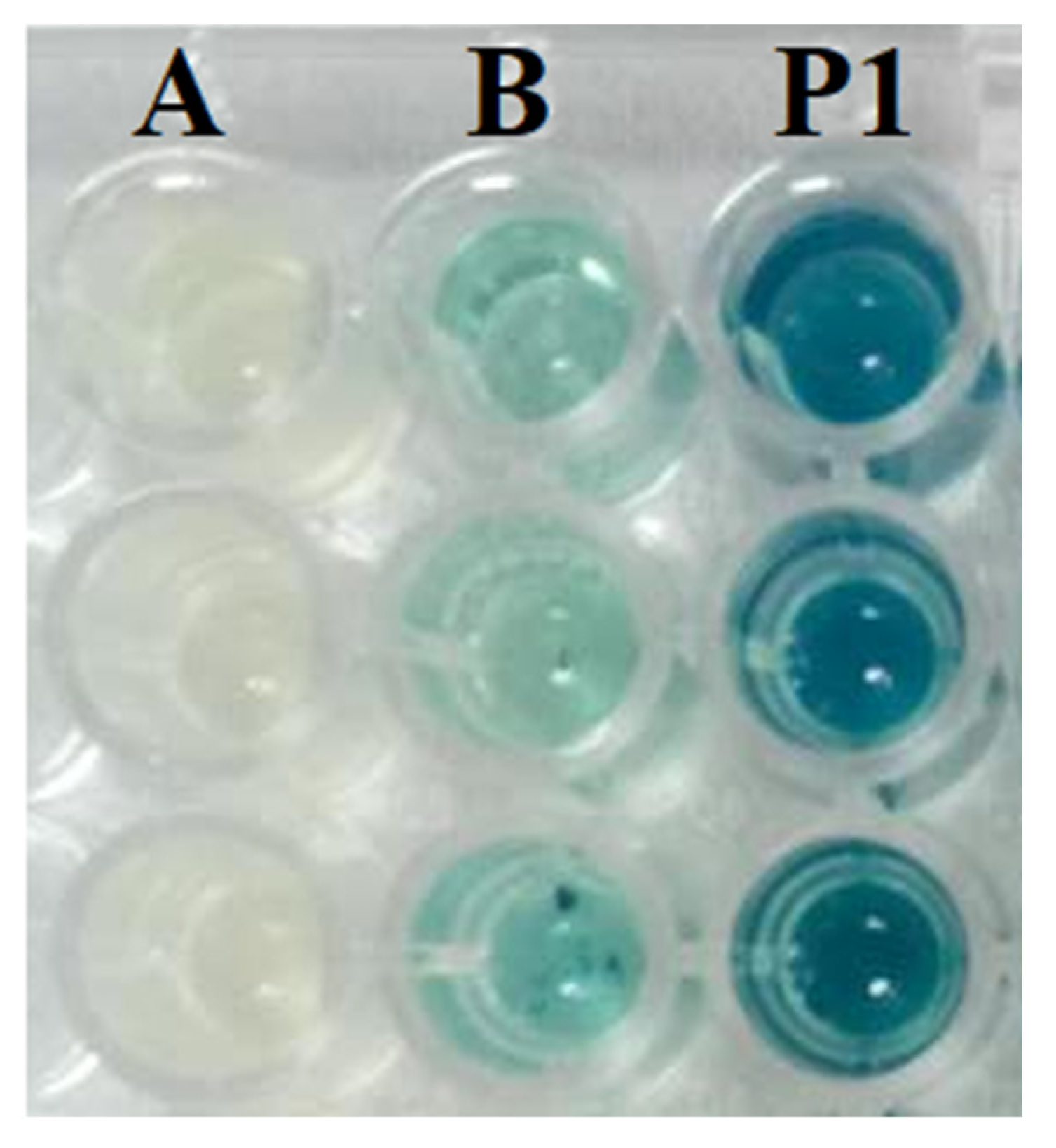
3.2. Screening for AHL Production
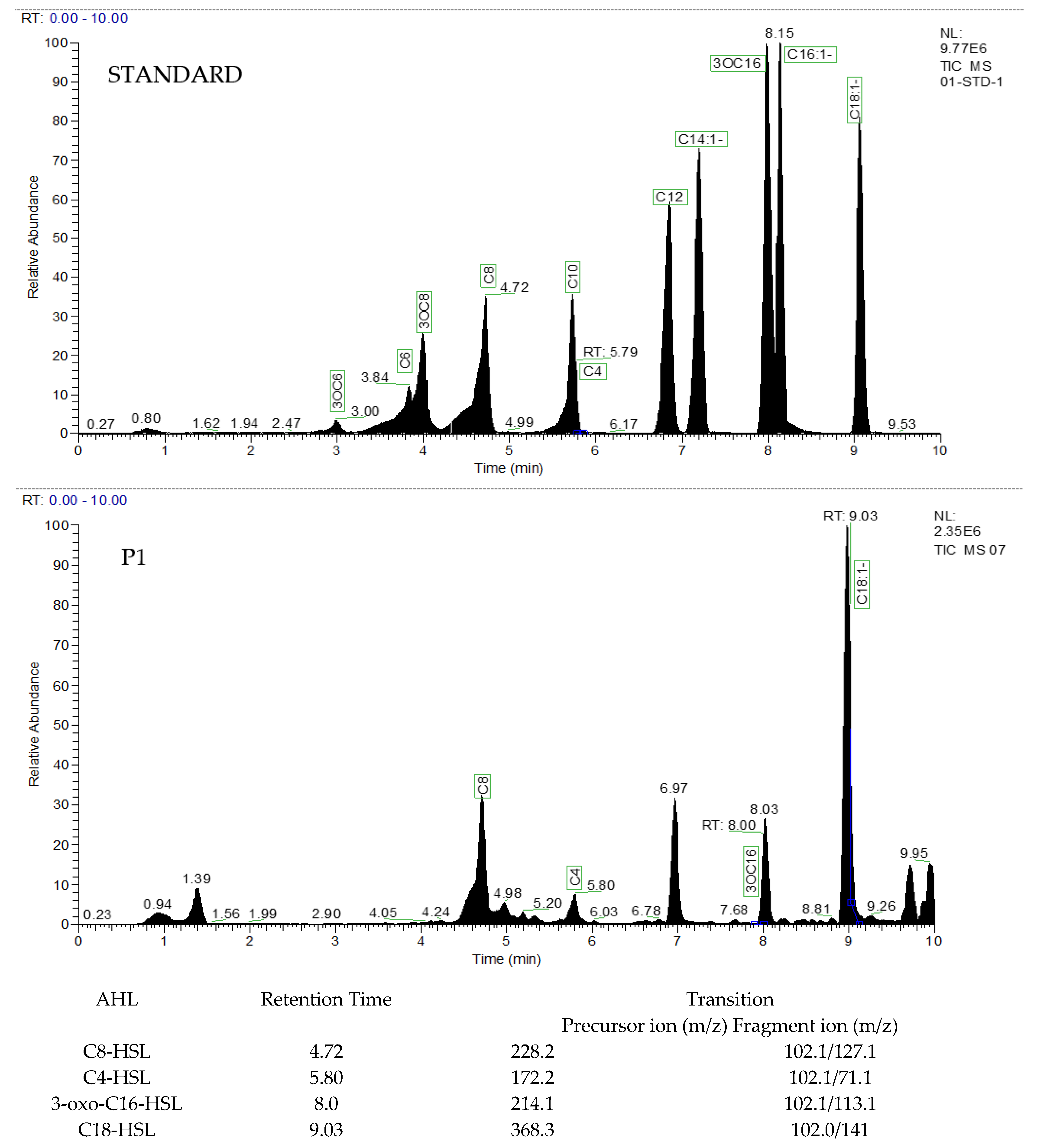
3.3. Extraction and Characterization of AHL
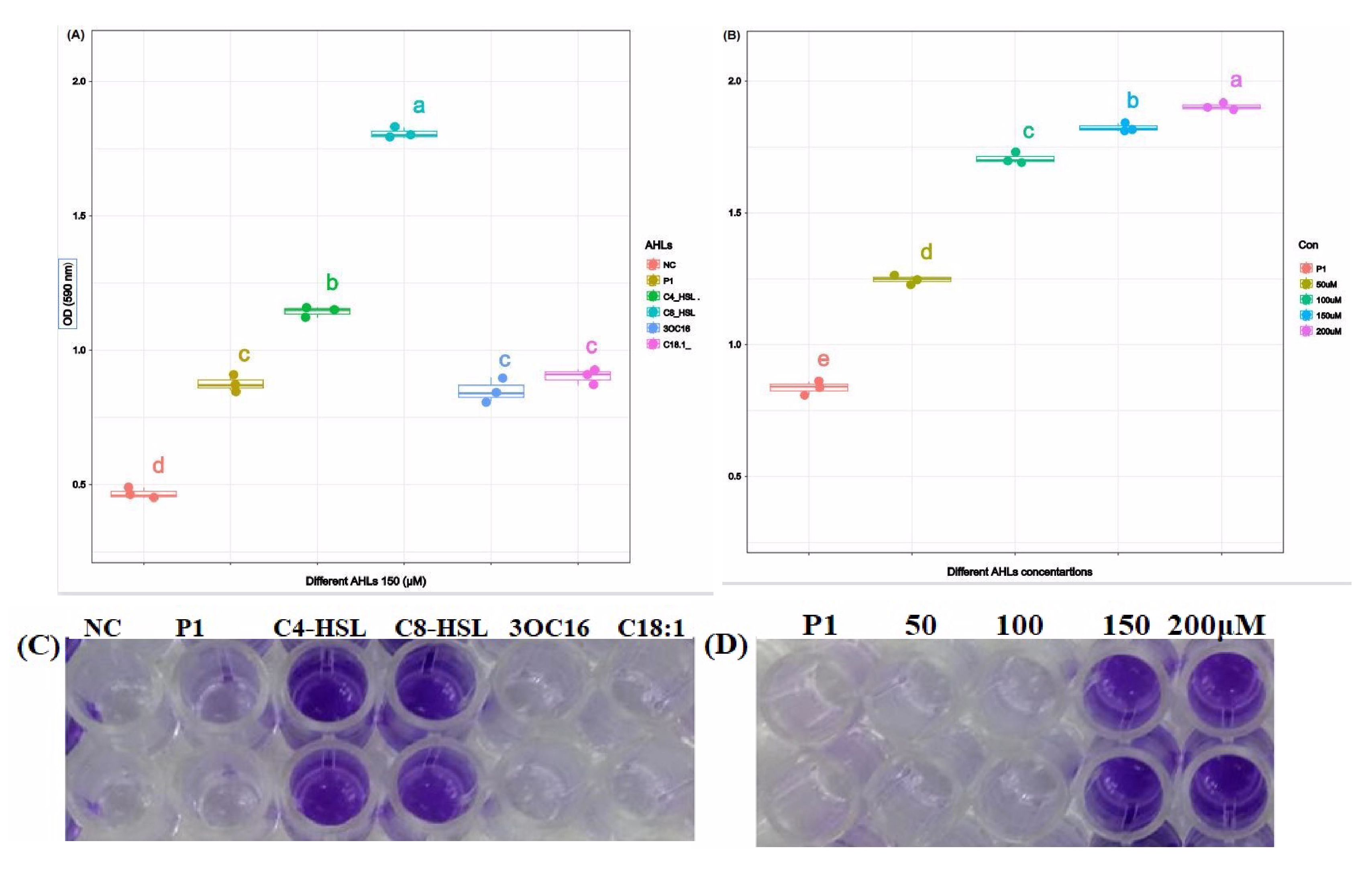
3.4. Biofilm Formation Assay
3.4.1. Crystal Violet Assay
3.4.2. Scanning Electron Microscopy (SEM) Assay
3.4.3. Confocal Laser Scanning Microscopy (CLSM) Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Egan, S.; Harder, T.; Burke, C.; Steinberg, P.; Kjelleberg, S.; Thomas, T. The seaweed holobiont: Understanding seaweed–bacteria interactions. FEMS Microbiol. Rev. 2013, 37, 462–476. [Google Scholar] [CrossRef] [PubMed]
- Duarte, C.M.; Middelburg, J.J.; Caraco, N. Major role of marine vegetation on the oceanic carbon cycle. Biogeosciences 2005, 2, 1–8. [Google Scholar] [CrossRef]
- D’Archino, R.; Piazzi, L. Macroalgal assemblages as indicators of the ecological status of marine coastal systems: A review. Ecol. Indic. 2021, 129, 107835. [Google Scholar] [CrossRef]
- Wahl, M.; Goecke, F.; Labes, A.; Dobretsov, S.; Weinberger, F. The Second Skin: Ecological Role of Epibiotic Biofilms on Marine Organisms. Front. Microbiol. 2012, 3, 292. [Google Scholar] [CrossRef] [PubMed]
- Hollants, J.; Leliaert, F.; De Clerck, O.; Willems, A. What we can learn from sushi: A review on seaweed–bacterial associations. FEMS Microbiol. Ecol. 2013, 83, 1–16. [Google Scholar] [CrossRef] [PubMed]
- van der Loos, L.M.; Eriksson, B.K.; Salles, J.F. The macroalgal holobiont in a changing sea. Trends Microbiol. 2019, 27, 635–650. [Google Scholar] [CrossRef]
- Thajuddin, N.; Dhanasekaran, D. Algae: Organisms for Imminent Biotechnology; BoD–Books on Demand: Norderstedt, Germany, 2016. [Google Scholar]
- Abisado, R.; Benomar, S.; Klaus, J.; Dandekar, A.; Chandler, J. Bacterial quorum sensing and microbial community interactions. MBio 2018, 9, e02331-17. [Google Scholar] [CrossRef]
- Borges, A.; Simões, M. Quorum sensing inhibition by marine bacteria. Mar. Drugs 2019, 17, 427. [Google Scholar] [CrossRef]
- Dobretsov, S.; Teplitski, M.; Paul, V. Mini-review: Quorum sensing in the marine environment and its relationship to biofouling. Biofouling 2009, 25, 413–427. [Google Scholar] [CrossRef]
- Turan, N.B.; Chormey, D.S.; Büyükpınar, Ç.; Engin, G.O.; Bakirdere, S. Quorum sensing: Little talks for an effective bacterial coordination. TrAC Trends Anal. Chem. 2017, 91, 1–11. [Google Scholar] [CrossRef]
- Armstrong, E.; Yan, L.; Boyd, K.G.; Wright, P.C.; Burgess, J.G. The symbiotic role of marine microbes on living surfaces. Hydrobiologia 2001, 461, 37–40. [Google Scholar] [CrossRef]
- Pei, P.; Aslam, M.; Du, H.; Liang, H.; Wang, H.; Liu, X.; Chen, W. Environmental factors shape the epiphytic bacterial communities of Gracilariopsis lemaneiformis. Sci. Rep. 2021, 11, 8671. [Google Scholar] [CrossRef] [PubMed]
- Case, R.J.; Longford, S.R.; Campbell, A.H.; Low, A.; Tujula, N.; Steinberg, P.D.; Kjelleberg, S. Temperature induced bacterial virulence and bleaching disease in a chemically defended marine macroalga. Environ. Microbiol. 2011, 13, 529–537. [Google Scholar] [CrossRef]
- Croft, M.T.; Lawrence, A.D.; Raux-Deery, E.; Warren, M.J.; Smith, A.G. Algae acquire vitamin B 12 through a symbiotic relationship with bacteria. Nature 2005, 438, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.H. Observations on the pathology of seaweeds in the Pacific Northwest. Can. J. Bot. 1977, 55, 1019–1027. [Google Scholar] [CrossRef]
- Sawabe, T.; Makino, H.; Tatsumi, M.; Nakano, K.; Tajima, K.; Iqbal, M.M.; Yumoto, I.; Ezura, Y.; Christen, R. Pseudoalteromonas bacteriolytica sp. nov., a marine bacterium that is the causative agent of red spot disease of Laminaria japonica. Int. J. Syst. Evol. Microbiol. 1998, 48, 769–774. [Google Scholar] [CrossRef]
- Huang, L.; Yan, X. Study on the red-rotting disease of Porphyra blades. J. Shanghai Ocean Univ. 2010, 19, 226–231. [Google Scholar]
- Takahashi, M. Pythium porphyrae Takahashi et Sasaki, sp. nov. causing red rot of marine red algae Porphyra spp. Trans. Mycol. Soc. Jpn. 1977, 18, 279–285. [Google Scholar]
- Wang, J.; Stanford, K.; McAllister, T.A.; Johnson, R.P.; Chen, J.; Hou, H.; Zhang, G.; Niu, Y.D. Biofilm formation, virulence gene profiles, and antimicrobial resistance of nine serogroups of non-O157 Shiga toxin–producing Escherichia coli. Foodborne Pathog. Dis. 2016, 13, 316–324. [Google Scholar] [CrossRef]
- Gachon, C.M.; Sime-Ngando, T.; Strittmatter, M.; Chambouvet, A.; Kim, G.H. Algal diseases: Spotlight on a black box. Trends Plant Sci. 2010, 15, 633–640. [Google Scholar] [CrossRef]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef] [PubMed]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as complex differentiated communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef]
- Novick, R.P.; Geisinger, E. Quorum sensing in staphylococci. Annu. Rev. Genet. 2008, 42, 541–564. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.-L.; Bassler, B.L. Bacterial quorum-sensing network architectures. Annu. Rev. Genet. 2009, 43, 197–222. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef]
- Kalia, V.C.; Patel, S.K.; Kang, Y.C.; Lee, J.-K. Quorum sensing inhibitors as antipathogens: Biotechnological applications. Biotechnol. Adv. 2019, 37, 68–90. [Google Scholar] [CrossRef]
- Schuster, M.; Joseph Sexton, D.; Diggle, S.P.; Peter Greenberg, E. Acyl-homoserine lactone quorum sensing: From evolution to application. Annu. Rev. Microbiol. 2013, 67, 43–63. [Google Scholar] [CrossRef]
- Monnet, V.; Juillard, V.; Gardan, R. Peptide conversations in Gram-positive bacteria. Crit. Rev. Microbiol. 2016, 42, 339–351. [Google Scholar] [CrossRef]
- Williams, P. Quorum sensing, communication and cross-kingdom signalling in the bacterial world. Microbiology 2007, 153, 3923–3938. [Google Scholar] [CrossRef]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal–response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef]
- Huang, X.; Gao, Y.; Ma, Z.; Lin, G.; Cai, Z.; Zhou, J. Profile of Citrobacter freundii ST2, a Multi-acyl-homoserine Lactone Producer Associated with Marine Dinoflagellates. Curr. Microbiol. 2017, 74, 68–76. [Google Scholar] [CrossRef]
- Decho, A.W.; Visscher, P.T.; Ferry, J.; Kawaguchi, T.; He, L.; Przekop, K.M.; Norman, R.S.; Reid, R.P. Autoinducers extracted from microbial mats reveal a surprising diversity of N-acylhomoserine lactones (AHLs) and abundance changes that may relate to diel pH. Environ. Microbiol. 2009, 11, 409–420. [Google Scholar] [CrossRef]
- Liu, D.; Keesing, J.K.; Dong, Z.; Zhen, Y.; Di, B.; Shi, Y.; Fearns, P.; Shi, P. Recurrence of the world’s largest green-tide in 2009 in Yellow Sea, China: Porphyra yezoensis aquaculture rafts confirmed as nursery for macroalgal blooms. Mar. Pollut. Bull. 2010, 60, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Venkatraman, K.L.; Mehta, A. Health benefits and pharmacological effects of Porphyra species. Plant Foods Hum. Nutr. 2019, 74, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xie, C.; Chen, C.; Ji, D.; Zhou, W. Physiological responses of gametophytic blades of Porphyra haitanensis to rising temperature stresses. J. Fish. China 2011, 35, 379–386. [Google Scholar]
- Pan, C.; Ma, J.; Tao, F.; Ji, C.; Zhao, Y.; Chen, S.; Yang, X. Novel insight into the antioxidant proteins derived from laver (Porphyra haitanensis) by proteomics analysis and protein based bioinformatics. Food Biosci. 2021, 42, 101134. [Google Scholar] [CrossRef]
- Ye, Y.; Zhang, L.; Yang, R.; Luo, Q.; Chen, H.; Yan, X.; Tang, H. Metabolic phenotypes associated with high-temperature tolerance of Porphyra haitanensis strains. J. Agric. Food Chem. 2013, 61, 8356–8363. [Google Scholar] [CrossRef]
- Xiong, Y.; Yang, R.; Sun, X.; Yang, H.; Chen, H. Effect of the epiphytic bacterium Bacillus sp. WPySW2 on the metabolism of Pyropia haitanensis. J. Appl. Phycol. 2018, 30, 1225–1237. [Google Scholar] [CrossRef]
- Singh, R.P.; Bijo, A.; Baghel, R.S.; Reddy, C.; Jha, B. Role of bacterial isolates in enhancing the bud induction in the industrially important red alga Gracilaria dura. FEMS Microbiol. Ecol. 2011, 76, 381–392. [Google Scholar] [CrossRef]
- Lane, D. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; John Wiley and Sons: New York, NY, USA, 1991. [Google Scholar]
- Bauer, R.; Bekker, J.P.; van Wyk, N.; du Toit, C.; Dicks, L.M.; Kossmann, J. Exopolysaccharide production by lactose-hydrolyzing bacteria isolated from traditionally fermented milk. Int. J. Food Microbiol. 2009, 131, 260–264. [Google Scholar] [CrossRef]
- Saitoh, N. The neighbour-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 10, 471–483. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- McLean, R.J.; Whiteley, M.; Stickler, D.J.; Fuqua, W.C. Evidence of autoinducer activity in naturally occurring biofilms. FEMS Microbiol. Lett. 1997, 154, 259–263. [Google Scholar] [CrossRef]
- Torres, M.; Dessaux, Y.; Llamas, I. Saline environments as a source of potential quorum sensing disruptors to control bacterial infections: A review. Mar. Drugs 2019, 17, 191. [Google Scholar] [CrossRef]
- Yin, F.; Gao, D.; Yue, L.; Zhang, Y.; Liu, J.; Zhang, X.-H.; Yu, M. Diversity of Bacteria with Quorum Sensing and Quenching Activities from Hydrothermal Vents in the Okinawa Trough. Microorganisms 2023, 11, 748. [Google Scholar] [CrossRef]
- Blanchet, E.; Prado, S.; Stien, D.; Oliveira da Silva, J.; Ferandin, Y.; Batailler, N.; Intertaglia, L.; Escargueil, A.; Lami, R. Quorum sensing and quorum quenching in the Mediterranean seagrass Posidonia oceanica microbiota. Front. Mar. Sci. 2017, 4, 218. [Google Scholar] [CrossRef]
- Chu, W.; Lu, F.; Zhu, W.; Kang, C. Isolation and characterization of new potential probiotic bacteria based on quorum-sensing system. J. Appl. Microbiol. 2011, 110, 202–208. [Google Scholar] [CrossRef]
- Huang, Y.L.; Ki, J.S.; Case, R.J.; Qian, P. Diversity and acyl-homoserine lactone production among subtidal biofilm-forming bacteria. Aquat. Microb. Ecol. 2008, 52, 185–193. [Google Scholar] [CrossRef]
- Shaw, P.D.; Ping, G.; Daly, S.L.; Cha, C.; Cronan Jr, J.E.; Rinehart, K.L.; Farrand, S.K. Detecting and characterizing N-acyl-homoserine lactone signal molecules by thin-layer chromatography. Proc. Natl. Acad. Sci. USA 1997, 94, 6036–6041. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhao, A.; Feng, L.; Gao, H. Quorum sensing signals affect spoilage of refrigerated large yellow croaker (Pseudosciaena crocea) by Shewanella baltica. Int. J. Food Microbiol. 2016, 217, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Morin, D.; Grasland, B.; Vallée-Réhel, K.; Dufau, C.; Haras, D. On-line high-performance liquid chromatography–mass spectrometric detection and quantification of N-acylhomoserine lactones, quorum sensing signal molecules, in the presence of biological matrices. J. Chromatogr. A 2003, 1002, 79–92. [Google Scholar] [CrossRef]
- Hou, H.-M.; Zhu, Y.-L.; Wang, J.-Y.; Jiang, F.; Qu, W.-Y.; Zhang, G.-L.; Hao, H.-S. Characteristics of N-acylhomoserine lactones produced by Hafnia alvei H4 isolated from spoiled instant sea cucumber. Sensors 2017, 17, 772. [Google Scholar] [CrossRef]
- Nithya, C.; Begum, M.F.; Pandian, S.K. Marine bacterial isolates inhibit biofilm formation and disrupt mature biofilms of Pseudomonas aeruginosa PAO1. Appl. Microbiol. Biotechnol. 2010, 88, 341–358. [Google Scholar] [CrossRef]
- Heydorn, A.; Nielsen, A.T.; Hentzer, M.; Sternberg, C.; Givskov, M.; Ersbøll, B.K.; Molin, S. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology 2000, 146 Pt 10, 2395–2407. [Google Scholar] [CrossRef]
- Marra, D.; Perna, I.; Pota, G.; Vitiello, G.; Pezzella, A.; Toscano, G.; Luciani, G.; Caserta, S. Nanoparticle Coatings on Glass Surfaces to Prevent Pseudomonas fluorescens AR 11 Biofilm Formation. Microorganisms 2023, 11, 621. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, N.M.; Cicirelli, E.M.; Kan, J.; Chen, F.; Fuqua, C.; Hill, R.T. Diversity and quorum-sensing signal production of Proteobacteria associated with marine sponges. Environ. Microbiol. 2008, 10, 75–86. [Google Scholar] [CrossRef]
- Fuqua, C.; Parsek, M.R.; Greenberg, E.P. Regulation of gene expression by cell-to-cell communication: Acyl-homoserine lactone quorum sensing. Annu. Rev. Genet. 2001, 35, 439–468. [Google Scholar] [CrossRef]
- Hughes, D.T.; Sperandio, V. Inter-kingdom signalling: Communication between bacteria and their hosts. Nat. Rev. Microbiol. 2008, 6, 111–120. [Google Scholar] [CrossRef]
- Burke, C.; Thomas, T.; Lewis, M.; Steinberg, P.; Kjelleberg, S. Composition, uniqueness and variability of the epiphytic bacterial community of the green alga Ulva australis. ISME J. 2011, 5, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Lachnit, T.; Meske, D.; Wahl, M.; Harder, T.; Schmitz, R. Epibacterial community patterns on marine macroalgae are host-specific but temporally variable. Environ. Microbiol. 2011, 13, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Jatt, A.N.; Tang, K.; Liu, J.; Zhang, Z.; Xiao-Hua, Z. Quorum sensing in marine snow and its possible influence on production of extracellular hydrolytic enzymes in marine snow bacterium Pantoea ananatis B9. FEMS Microbiol. Ecol. 2015, 91, 1–13. [Google Scholar] [CrossRef]
- Urvoy, M.; Labry, C.; L’helguen, S.; Lami, R. Quorum sensing regulates bacterial processes that play a major role in marine biogeochemical cycles. Front. Mar. Sci. 2022, 9, 834337. [Google Scholar] [CrossRef]
- Cuadrado-Silva, C.T.; Castellanos, L.; Arévalo-Ferro, C.; Osorno, O.E. Detection of quorum sensing systems of bacteria isolated from fouled marine organisms. Biochem. Syst. Ecol. 2013, 46, 101–107. [Google Scholar] [CrossRef]
- Freckelton, M.L.; Høj, L.; Bowden, B.F. Quorum sensing interference and structural variation of quorum sensing mimics in Australian soft coral. Front. Mar. Sci. 2018, 5, 198. [Google Scholar] [CrossRef]
- Charlesworth, J.C.; Watters, C.; Wong, H.L.; Visscher, P.T.; Burns, B.P. Isolation of novel quorum-sensing active bacteria from microbial mats in Shark Bay Australia. FEMS Microbiol. Ecol. 2019, 95, fiz035. [Google Scholar] [CrossRef]
- Reen, F.J.; Gutiérrez-Barranquero, J.A.; McCarthy, R.R.; Woods, D.F.; Scarciglia, S.; Adams, C.; Fog Nielsen, K.; Gram, L.; O’Gara, F. Quorum sensing signaling alters virulence potential and population dynamics in complex microbiome-host interactomes. Front. Microbiol. 2019, 10, 2131. [Google Scholar] [CrossRef]
- Su, Y.; Tang, K.; Liu, J.; Wang, Y.; Zheng, Y.; Zhang, X.-H. Quorum sensing system of Ruegeria mobilis Rm01 controls lipase and biofilm formation. Front. Microbiol. 2019, 9, 3304. [Google Scholar] [CrossRef]
- Urvoy, M.; Lami, R.; Dreanno, C.; Daudé, D.; Rodrigues, A.M.; Gourmelon, M.; L’Helguen, S.; Labry, C. Quorum sensing disruption regulates hydrolytic enzyme and biofilm production in estuarine bacteria. Environ. Microbiol. 2021, 23, 7183–7200. [Google Scholar] [CrossRef]
- Doberva, M.; Sanchez-Ferandin, S.; Toulza, E.; Lebaron, P.; Lami, R. Diversity of quorum sensing autoinducer synthases in the Global Ocean Sampling metagenomic database. Aquat. Microb. Ecol. 2015, 74, 107–119. [Google Scholar] [CrossRef]
- Su, Y.; Yang, Y.; Zhu, X.-Y.; Zhang, X.-H.; Yu, M. Metagenomic insights into the microbial assemblage capable of quorum sensing and quorum quenching in particulate organic matter in the Yellow Sea. Front. Microbiol. 2021, 11, 602010. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhu, J.; Cai, Z.; Lao, Y.; Jin, H.; Yu, K.; Zhang, B.; Zhou, J. Profiles of quorum sensing (QS)-related sequences in phycospheric microorganisms during a marine dinoflagellate bloom, as determined by a metagenomic approach. Microbiol. Res. 2018, 217, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Alviz-Gazitua, P.; González, A.; Lee, M.R.; Aranda, C.P. Molecular Relationships in Biofilm Formation and the Biosynthesis of Exoproducts in Pseudoalteromonas spp. Mar. Biotechnol. 2022, 24, 431–447. [Google Scholar] [CrossRef] [PubMed]
- Holmström, C.; Kjelleberg, S. Marine Pseudoalteromonas species are associated with higher organisms and produce biologically active extracellular agents. FEMS Microbiol. Ecol. 1999, 30, 285–293. [Google Scholar] [CrossRef]
- Zhu, J.; Beaber, J.W.; Moré, M.I.; Fuqua, C.; Eberhard, A.; Winans, S.C. Analogs of the autoinducer 3-oxooctanoyl-homoserine lactone strongly inhibit activity of the TraR protein of Agrobacterium tumefaciens. J. Bacteriol. 1998, 180, 5398–5405. [Google Scholar] [CrossRef]
- Zhu, J.; Chai, Y.; Zhong, Z.; Li, S.; Winans, S.C. Agrobacterium bioassay strain for ultrasensitive detection of N-acylhomoserine lactone-type quorum-sensing molecules: Detection of autoinducers in Mesorhizobium huakuii. Appl. Environ. Microbiol. 2003, 69, 6949–6953. [Google Scholar] [CrossRef]
- Llamas, I.; Quesada, E.; Martínez-Cánovas, M.J.; Gronquist, M.; Eberhard, A.; Gonzalez, J.E. Quorum sensing in halophilic bacteria: Detection of N-acyl-homoserine lactones in the exopolysaccharide-producing species of Halomonas. Extremophiles 2005, 9, 333–341. [Google Scholar] [CrossRef]
- Yu, Z.; Yu, D.; Mao, Y.; Zhang, M.; Ding, M.; Zhang, J.; Wu, S.; Qiu, J.; Yin, J. Identification and characterization of a LuxI/R-type quorum sensing system in Pseudoalteromonas. Res. Microbiol. 2019, 170, 243–255. [Google Scholar] [CrossRef]
- Dixit, S.; Dubey, R.C.; Maheshwari, D.K.; Seth, P.K.; Bajpai, V.K. Roles of quorum sensing molecules from Rhizobium etli RT1 in bacterial motility and biofilm formation. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2017, 48, 815–821. [Google Scholar] [CrossRef]
- Ransome, E.; Munn, C.B.; Halliday, N.; Cámara, M.; Tait, K. Diverse profiles of N-acyl-homoserine lactone molecules found in cnidarians. FEMS Microbiol. Ecol. 2014, 87, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ikawa, A.; Okaue, S.; Taniguchi, S.; Osaka, I.; Yoshimoto, A.; Kishida, Y.; Arakawa, R.; Enomoto, K. Quorum sensing signaling molecules involved in the production of violacein by Pseudoalteromonas. Biosci. Biotechnol. Biochem. 2008, 72, 1958–1961. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zheng, L.; Zhou, W.; Cui, Z.; Han, P.; Tian, L.; Wang, X. A case study on chemical defense based on quorum sensing: Antibacterial activity of sponge-associated bacterium Pseudoalteromonas sp. NJ6-3-1 induced by quorum sensing mechanisms. Ann. Microbiol. 2011, 61, 247–255. [Google Scholar] [CrossRef]
- Ziesche, L.; Bruns, H.; Dogs, M.; Wolter, L.; Mann, F.; Wagner-Döbler, I.; Brinkhoff, T.; Schulz, S. Homoserine lactones, methyl oligohydroxybutyrates, and other extracellular metabolites of macroalgae-associated bacteria of the Roseobacter clade: Identification and functions. ChemBioChem 2015, 16, 2094–2107. [Google Scholar] [CrossRef]
- Morohoshi, T.; Nakamura, Y.; Yamazaki, G.; Ishida, A.; Kato, N.; Ikeda, T. The plant pathogen Pantoea ananatis produces N-acylhomoserine lactone and causes center rot disease of onion by quorum sensing. J. Bacteriol. 2007, 189, 8333–8338. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Zhu, J.; Feng, L.; Li, J.; Liu, X. Characterization of LuxI/LuxR and their regulation involved in biofilm formation and stress resistance in fish spoilers Pseudomonas fluorescens. Int. J. Food Microbiol. 2019, 297, 60–71. [Google Scholar] [CrossRef]
- Singh, R.P.; Baghel, R.S.; Reddy, C.; Jha, B. Effect of quorum sensing signals produced by seaweed-associated bacteria on carpospore liberation from Gracilaria dura. Front. Plant Sci. 2015, 6, 117. [Google Scholar] [CrossRef]
- Whiteley, M.; Diggle, S.P.; Greenberg, E.P. Progress in and promise of bacterial quorum sensing research. Nature 2017, 551, 313–320. [Google Scholar] [CrossRef]
- Abdul Malik, S.A.; Bazire, A.; Gamboa-Muñoz, A.; Bedoux, G.; Robledo, D.; García-Maldonado, J.Q.; Bourgougnon, N. Screening of Surface-associated Bacteria from the Mexican Red Alga Halymenia floresii for Quorum Sensing Activity. Microbiology 2020, 89, 778–788. [Google Scholar] [CrossRef]
- Hu, H.; He, J.; Liu, J.; Yu, H.; Tang, J.; Zhang, J. Role of N-acyl-homoserine lactone (AHL) based quorum sensing on biofilm formation on packing media in wastewater treatment process. RSC Adv. 2016, 6, 11128–11139. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Q.; Ma, S.; Hu, H.; Wu, B.; Zhang, X.-X.; Ren, H. Distribution characteristics of N-acyl homoserine lactones during the moving bed biofilm reactor biofilm development process: Effect of carbon/nitrogen ratio and exogenous quorum sensing signals. Bioresour. Technol. 2019, 289, 121591. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Zhu, X.; Zhou, J.; Cai, Z. Biofilm inhibition and pathogenicity attenuation in bacteria by Proteus mirabilis. R. Soc. Open Sci. 2018, 5, 170702. [Google Scholar] [CrossRef] [PubMed]
- Fazli, M.; Almblad, H.; Rybtke, M.L.; Givskov, M.; Eberl, L.; Tolker-Nielsen, T. Regulation of biofilm formation in P seudomonas and B urkholderia species. Environ. Microbiol. 2014, 16, 1961–1981. [Google Scholar] [CrossRef] [PubMed]
- Hayek, M.; Baraquet, C.; Lami, R.; Blache, Y.; Molmeret, M. The Marine Bacterium Shewanella woodyi Produces C 8-HSL to Regulate Bioluminescence. Microb. Ecol. 2020, 79, 865–881. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Song, G.; Son, J.; Howard, L.; Yu, X.-Y. Revealing the Bacterial Quorum-Sensing Effect on the Biofilm Formation of Diatom Cylindrotheca sp. Using Multimodal Imaging. Microorganisms 2023, 11, 1841. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location (Nan’ao) | ||||
|---|---|---|---|---|
| Month | Temperature (°C) | pH | Salinity (ppt) | DO |
| November | 13.8 | 7.8 | 30.5 | 8.0 |
| March | 21.2 | 8.1 | 31.7 | 8.1 |
| Lab Code | Identified as | Accession No. | Similarity (%) | Genera |
|---|---|---|---|---|
| P1 | Pseudoalteromonas galatheae | NR_178361.1 | 99.59 | Proteobacteria |
| P2 | Staphylococcus saprophyticus strain UTI-045 | CP054831.1 | 100.00 | Firmicutes |
| P3 | Vibrio alginolyticus strain NBRC 15630 | NR_122050.1 | 99.55 | Proteobacteria |
| P4 | Vibrio sp. strain 201709CJKOP-32 | MH093789.1 | 99.48 | Proteobacteria |
| P5 | Vibrio antiquarius | NC_013456.1 | 98.82 | Proteobacteria |
| P6 | Vibrio sp. strain 201709CJKOP-38 | MH093795.1 | 99.93 | Proteobacteria |
| P8 | Zobellia russellii strain KMM 3677 | NR_024828.1 | 100 | Bacteroidetes |
| P9 | Paraglaciecola mesophila KMM 241 | NZ_BAEP01000046.1 | 99.70 | Proteobacteria |
| P10 | Vibrio diabolicus strain HE800 | NR_036811.1 | 98.82 | Proteobacteria |
| P12 | Vibrio sp. VibC-Oc-059 | KF577068.1 | 100.00 | Proteobacteria |
| P15 | Pseudoalteromonas sp. strain 5315 | ON026012.1 | 100 | Proteobacteria |
| P17 | Pseudoalteromonas distincta strain KMM 3548 | NR_025654.1 | 99.7 | Proteobacteria |
| P18 | Pseudoalteromonas sp. ZJHD1-34 | JN107745.1 | 99.77 | Proteobacteria |
| P19 | Vibrio sp. strain 201709CJKOP-60 | MH093817.1 | 99.92 | Proteobacteria |
| P20 | Vibrio alginolyticus strain 2014V-1011 | CP046772.1 | 100.00 | Proteobacteria |
| P21 | Halomonas venusta strain MA-ZP17-13 | CP034367.1 | 99.80 | Proteobacteria |
| P. galatheae | Without AHL | With AHL | ||
|---|---|---|---|---|
| Dead | Live | Dead | Live | |
| Biomass (µm3/µm2) | 1.14 ± 0.01 | 2.63 ± 0.37 | 5.91 ± 0.78 | 9.99 ± 1.30 |
| Mean thickness (Biomass) (µm) | 4.36 ± 0.60 | 6.50 ± 1.25 | 7.42 ± 1.20 | 9.60 ± 0.94 |
| Mean thickness (Area) (µm) | 1.24 ± 0.03 | 4.36 ± 1.05 | 5.05 ± 1.31 | 8.10 ± 1.37 |
| Maximum thickness (µm) | 5.54 ± 0.52 | 8.61 ± 1.88 | 21.01 ± 4.18 | 24.05 ± 6.24 |
| Roughness (–) | 1.85 ± 0.03 | 0.95 ± 0.22 | 0.76 ± 0.29 | 0.19 ± 0.18 |
| Surface Area (105 m2) | 0.22 ± 0.02 | 1.81 ± 0.44 | 2.90 ± 0.71 | 3.76 ± 0.33 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aslam, M.; Pei, P.; Ye, P.; Li, T.; Liang, H.; Zhang, Z.; Ke, X.; Chen, W.; Du, H. Unraveling the Diverse Profile of N-Acyl Homoserine Lactone Signals and Their Role in the Regulation of Biofilm Formation in Porphyra haitanensis-Associated Pseudoalteromonas galatheae. Microorganisms 2023, 11, 2228. https://doi.org/10.3390/microorganisms11092228
Aslam M, Pei P, Ye P, Li T, Liang H, Zhang Z, Ke X, Chen W, Du H. Unraveling the Diverse Profile of N-Acyl Homoserine Lactone Signals and Their Role in the Regulation of Biofilm Formation in Porphyra haitanensis-Associated Pseudoalteromonas galatheae. Microorganisms. 2023; 11(9):2228. https://doi.org/10.3390/microorganisms11092228
Chicago/Turabian StyleAslam, Muhammad, Pengbing Pei, Peilin Ye, Tangcheng Li, Honghao Liang, Zezhi Zhang, Xiao Ke, Weizhou Chen, and Hong Du. 2023. "Unraveling the Diverse Profile of N-Acyl Homoserine Lactone Signals and Their Role in the Regulation of Biofilm Formation in Porphyra haitanensis-Associated Pseudoalteromonas galatheae" Microorganisms 11, no. 9: 2228. https://doi.org/10.3390/microorganisms11092228