
Mechanisms of Lung and Intestinal Microbiota and Innate Immune Changes Caused by Pathogenic Enterococcus Faecalis Promoting the Development of Pediatric Pneumonia

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains Identification by 16S Full-Length Sequencing
2.2. Experimental Design and Animals
2.3. Hematoxylin-Eosin Tissue Staining
2.4. Immunohistochemistry
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Quantitative Real-Time Polymerase Chain Reaction
2.7. Western Blot
2.8. Immunofluorescence
2.9. 16SrRNA Sequence
2.10. Statistical Analysis
3. Results
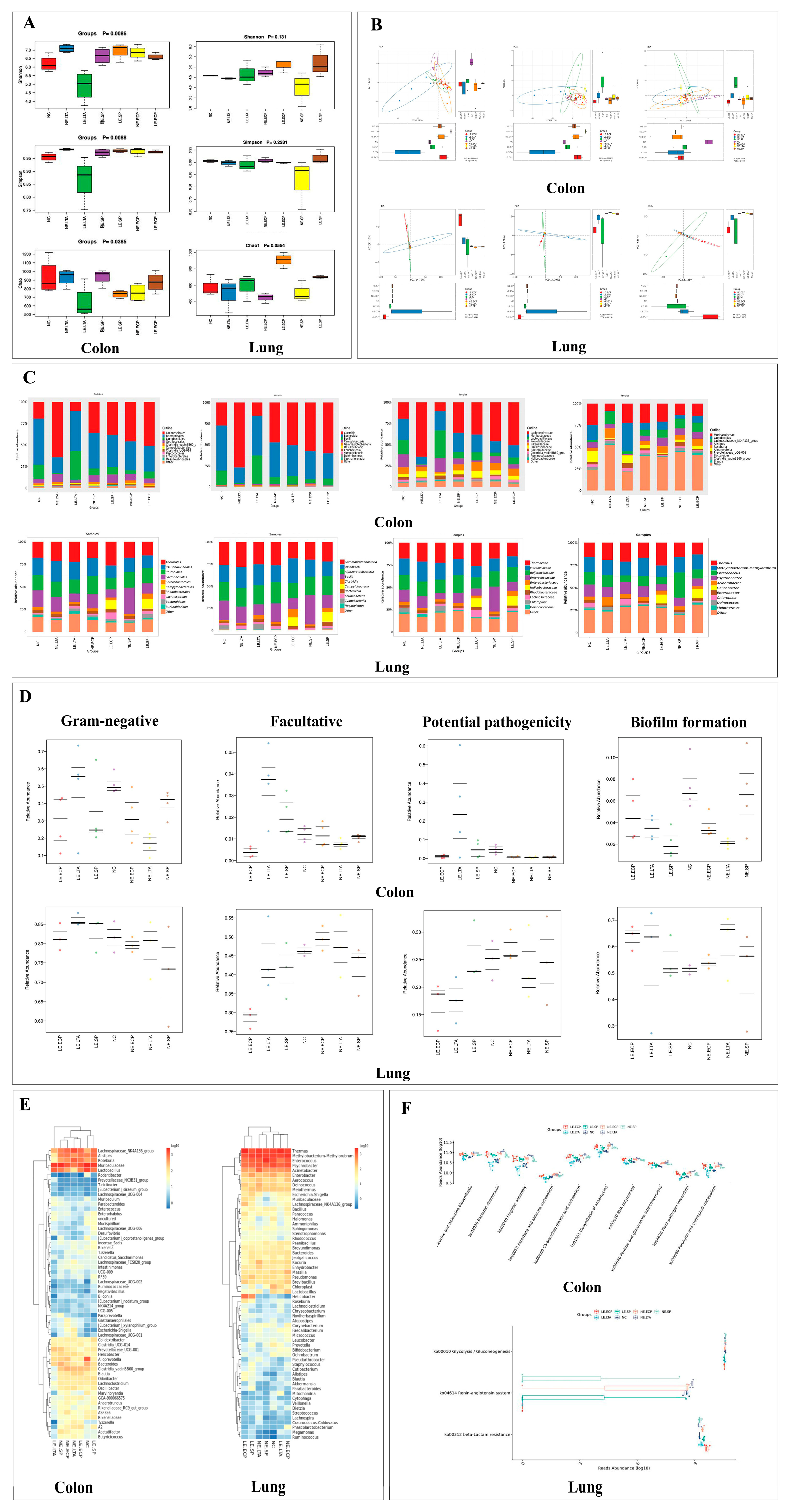
3.1. 16srRNA Sequence
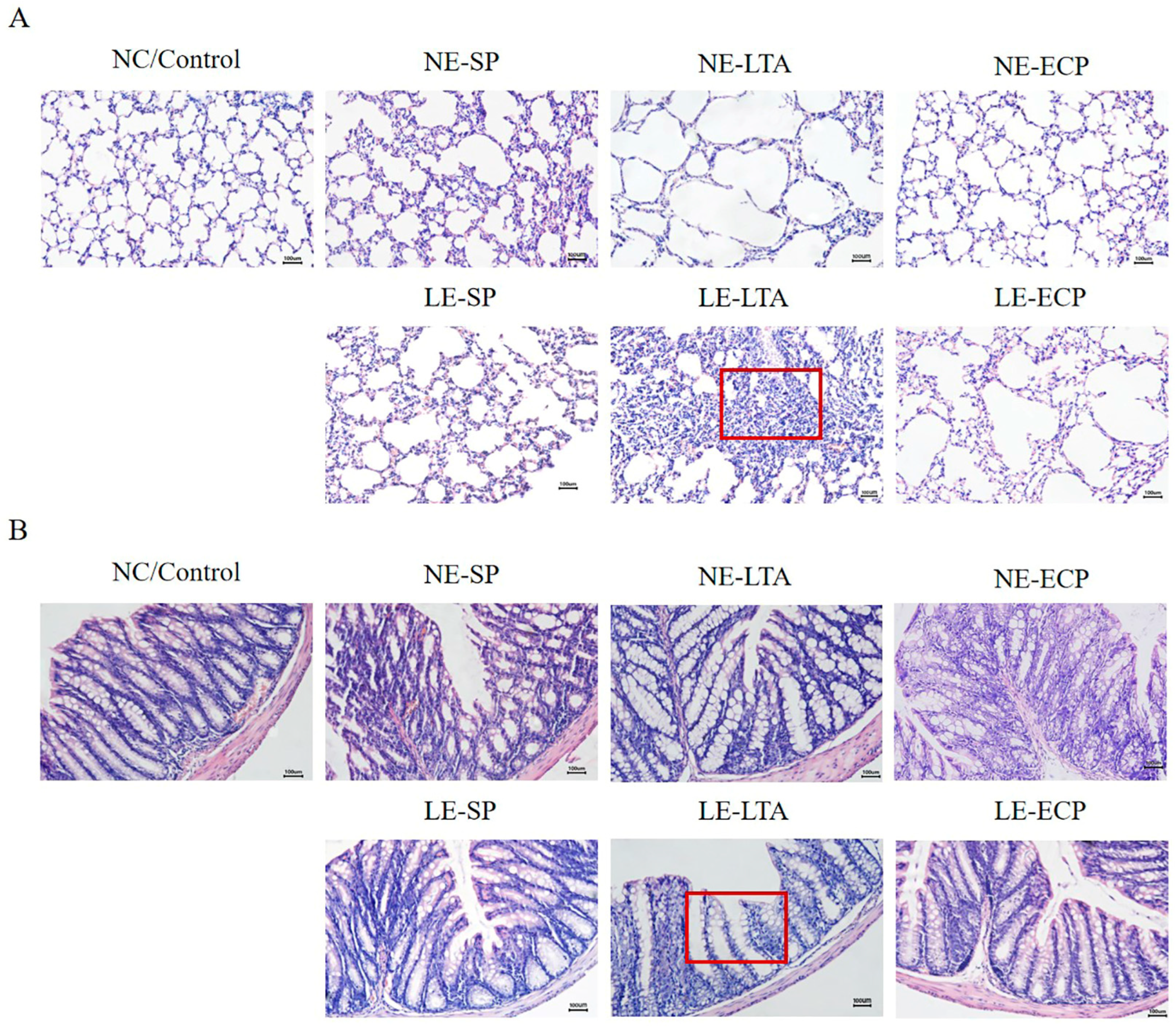
3.2. Pathological Findings of HE Staining of Mice Intestine and Lung
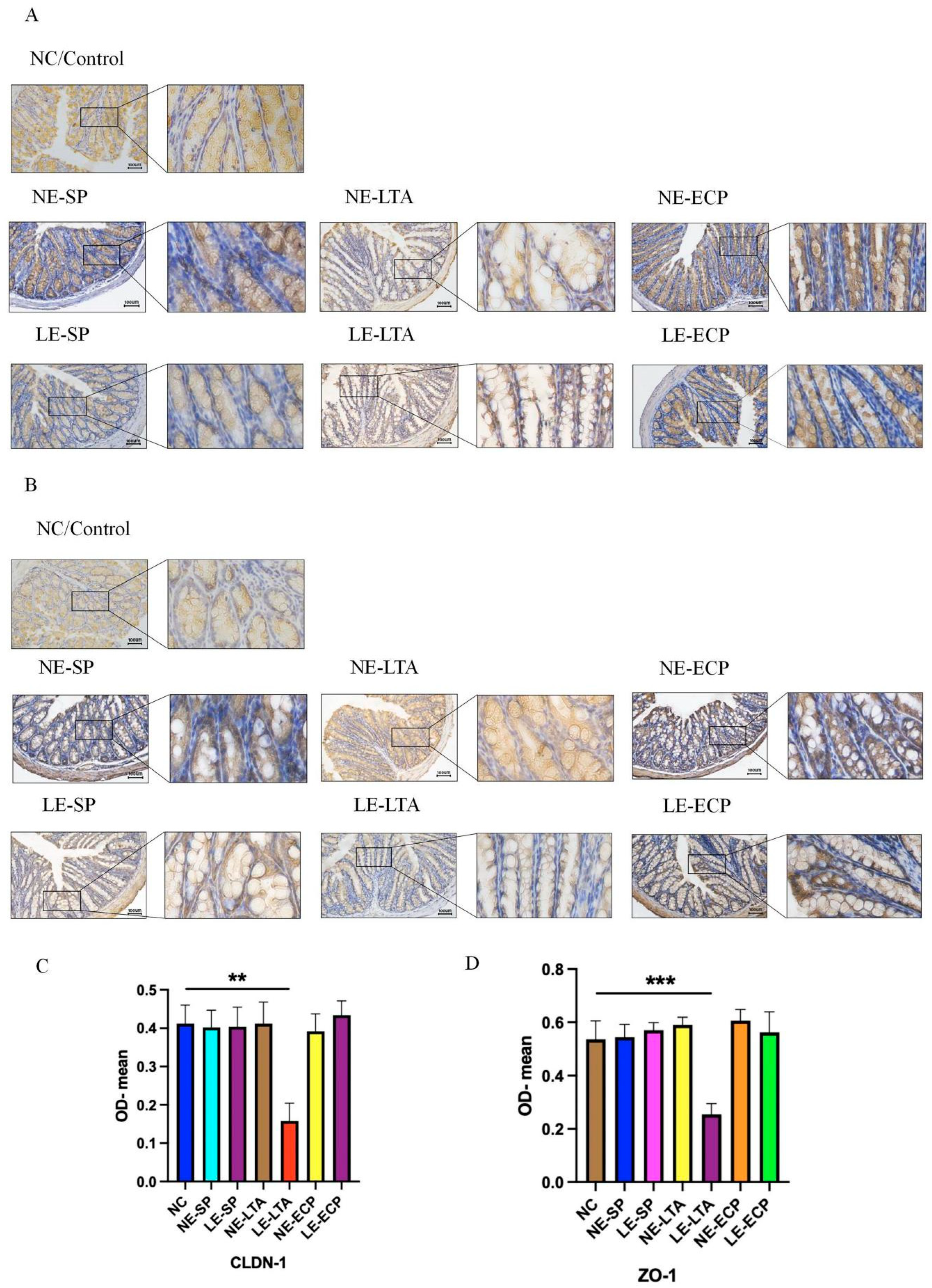
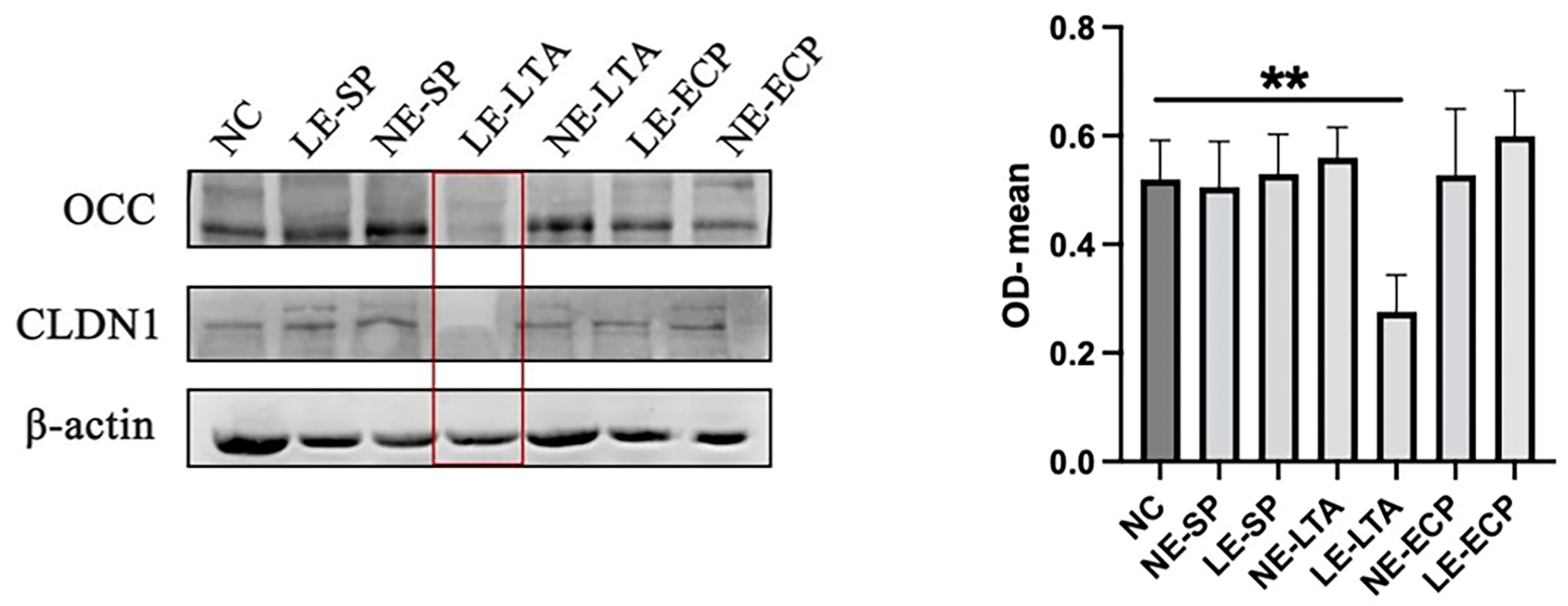
3.3. Mice Intestinal Closed Junction Protein Assay
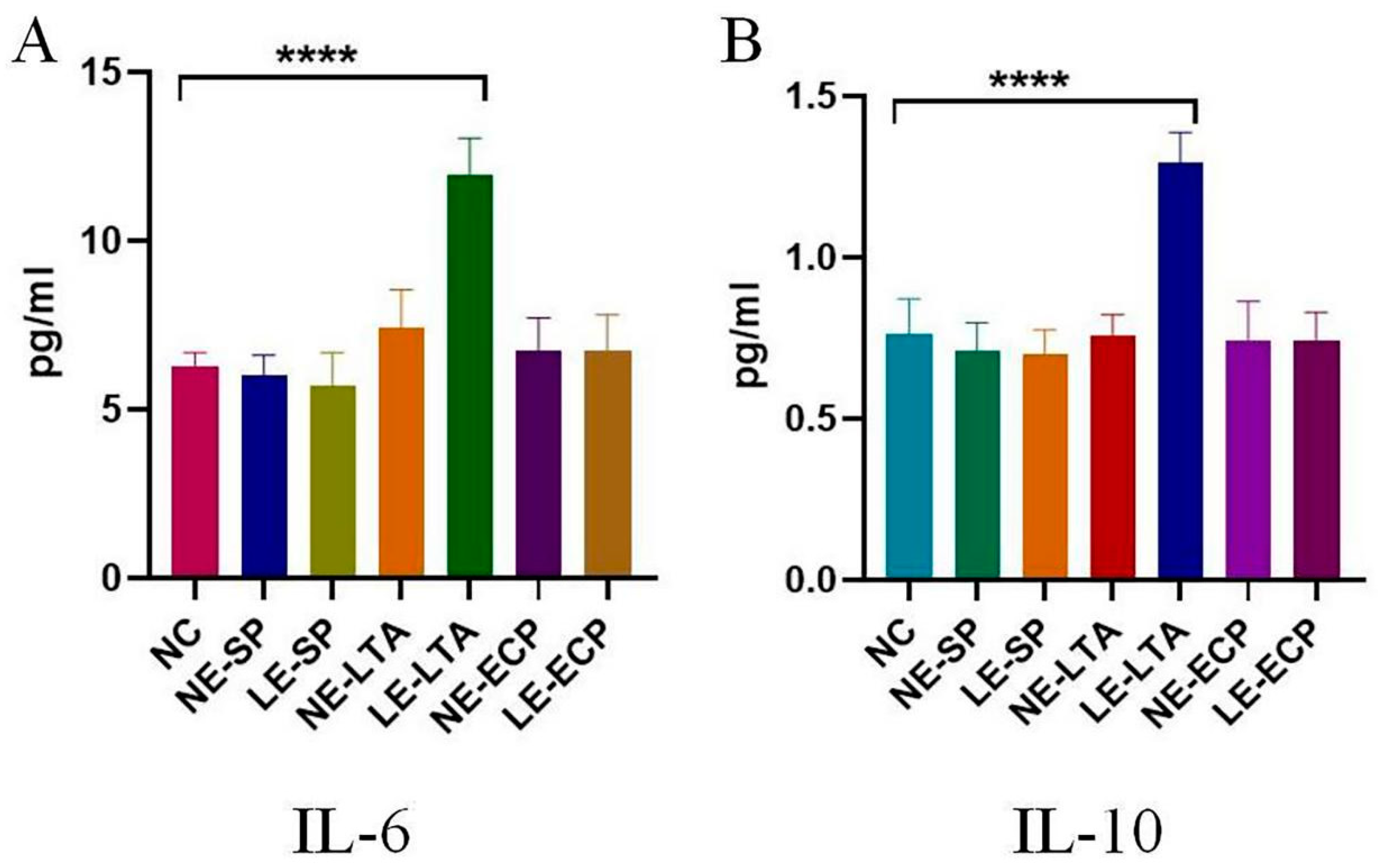
3.4. ELISA
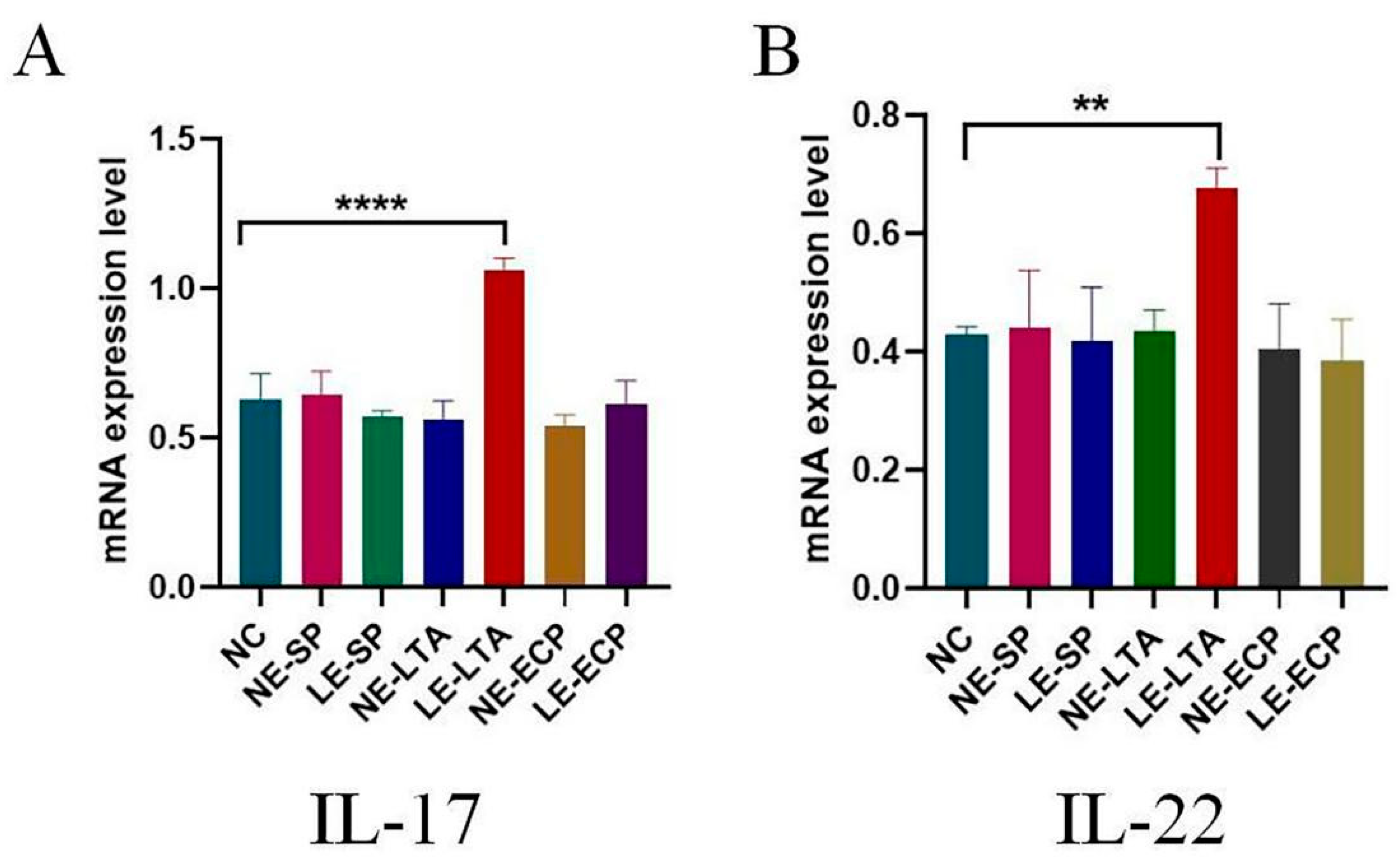
3.5. RT-PCR
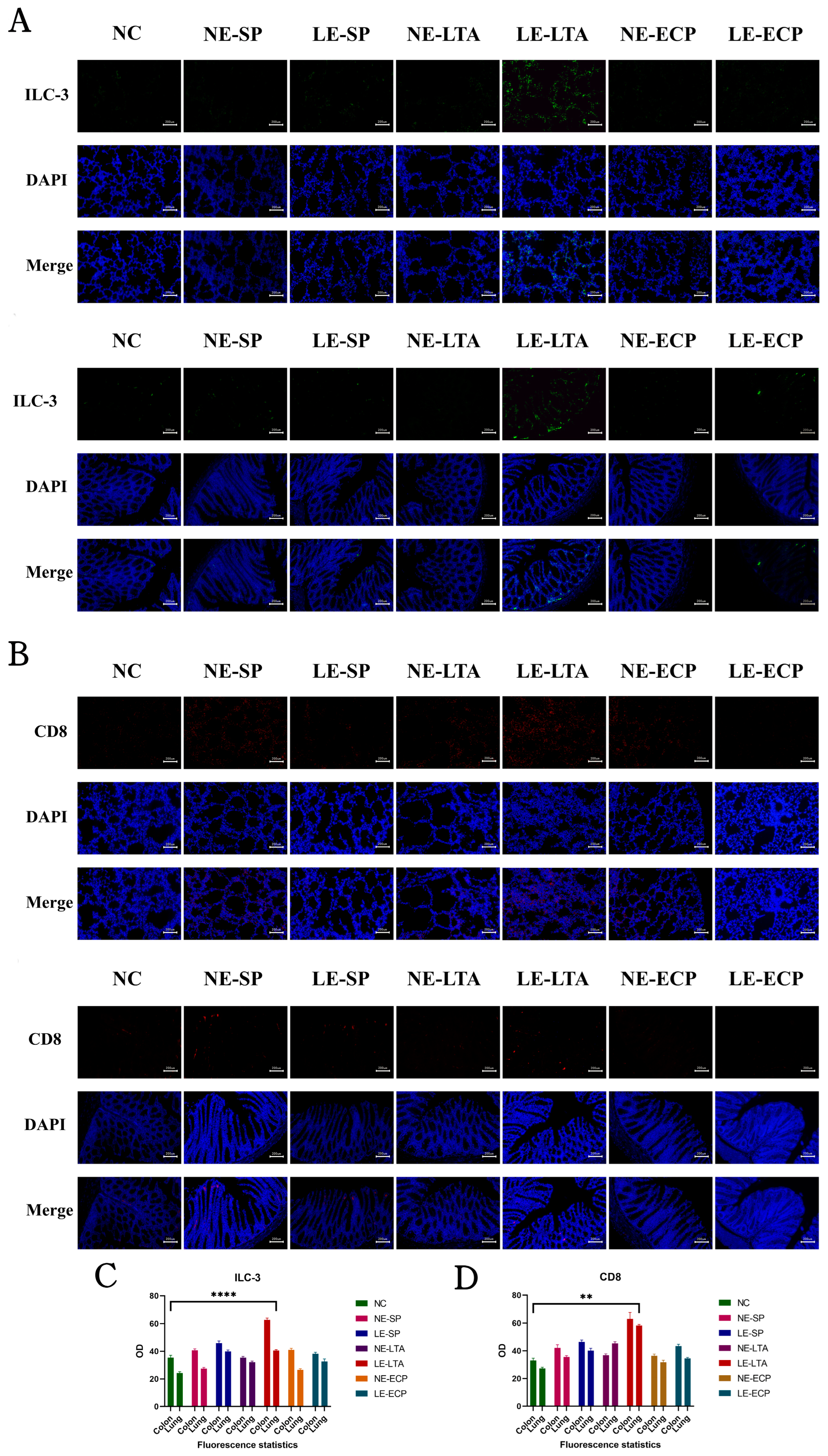
3.6. Immunofluorescence
3.7. Western Blot
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Langley, J.M.; Bradley, J.S. Defining pneumonia in critically ill infants and children. Pediatr. Crit. Care Med. 2005, 6, S9–S13. [Google Scholar] [CrossRef]
- Izadnegahdar, R.; Cohen, A.L.; Klugman, K.P.; Qazi, S. Childhood pneumonia in developing countries. Lancet Respir. Med. 2013, 1, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Obaro, S.K.; Madhi, S. Bacterial pneumonia vaccines and childhood pneumonia: Are we winning, refining, or redefining? Lancet Infect. Dis. 2006, 6, 150–161. [Google Scholar] [CrossRef]
- Zash, R.M.; Shapiro, R.L.; Leidner, J.; Wester, C.; McAdam, A.J.; Hodinka, R.L.; Thior, I.; Moffat, C.; Makhema, J.; McIntosh, K.; et al. The aetiology of diarrhoea, pneumonia and respiratory colonization of HIV-exposed infants randomized to breast- or formula-feeding. Paediatr. Int. Child Health 2016, 36, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Palmer, L.; Hall, C.B.; Katkin, J.P.; Shi, N.; Masaquel, A.S.; McLaurin, K.K.; Mahadevia, P.J. Respiratory outcomes, utilization and costs 12 months following a respiratory syncytial virus diagnosis among commercially insured late-preterm infants. Curr. Med. Res. Opin. 2011, 27, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Hernando, N.; Myakala, K.; Simona, F.; Knöpfel, T.; Thomas, L.; Murer, H.; Wagner, C.A.; Biber, J. Intestinal Depletion of NaPi-IIb/Slc34a2 in Mice: Renal and Hormonal Adaptation. J. Bone Miner. Res. 2015, 30, 1925–1937. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, L.; Walker, A.W.; Oliver, A.E.; Rogers, G.B.; Rivett, D.W.; Hampton, T.H.; Ashare, A.; Elborn, J.S.; De Soyza, A.; Carroll, M.P.; et al. Lung function and microbiota diversity in cystic fibrosis. Microbiome 2020, 8, 45. [Google Scholar] [CrossRef]
- Sivaprakasam, S.; Bhutia, Y.D.; Ramachandran, S.; Ganapathy, V. Cell-Surface and Nuclear Receptors in the Colon as Targets for Bacterial Metabolites and Its Relevance to Colon Health. Nutrients 2017, 9, 856. [Google Scholar] [CrossRef] [PubMed]
- Marincola Smith, P.; Choksi, Y.A.; Markham, N.O.; Hanna, D.N.; Zi, J.; Weaver, C.J.; Hamaamen, J.A.; Lewis, K.B.; Yang, J.; Liu, Q.; et al. Colon epithelial cell TGFβ signaling modulates the expression of tight junction proteins and barrier function in mice. Am. J. Physiol. Liver Physiol. 2021, 320, G936–G957. [Google Scholar] [CrossRef]
- Rittirsch, D.; Flierl, M.A.; Nadeau, B.A.; Day, D.E.; Huber-Lang, M.S.; Grailer, J.J.; Zetoune, F.S.; Andjelkovic, A.V.; Fasano, A.; Ward, P.A. Zonulin as pre-haptoglobin2 regulates lung permeability and activates the complement system. Am. J. Physiol. Lung Cell Mol. Physiol. 2013, 304, L863–L872. [Google Scholar] [CrossRef]
- Subramanian, S.; Geng, H.; Tan, X.-D. Cell death of intestinal epithelial cells in intestinal diseases. Sheng Li Xue Bao 2020, 72, 308–324. [Google Scholar] [PubMed]
- Lan, H.; Zhang, L.Y.; He, W.; Li, W.Y.; Zeng, Z.; Qian, B.; Wang, C.; Song, J.L. Sinapic Acid Alleviated Inflammation-Induced Intestinal Epithelial Barrier Dysfunction in Lipopolysaccharide-(LPS-) Treated Caco-2 Cells. Mediat. Inflamm. 2021, 2021, 5514075. [Google Scholar] [CrossRef] [PubMed]
- Drakhlis, L.; Biswanath, S.; Farr, C.M.; Lupanow, V.; Teske, J.; Ritzenhoff, K.; Franke, A.; Manstein, F.; Bolesani, E.; Kempf, H.; et al. Human heart-forming organoids recapitulate early heart and foregut development. Nat. Biotechnol. 2021, 39, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Ghembaza, A.; Vautier, M.; Cacoub, P.; Pourcher, V.; Saadoun, D. Risk Factors and Prevention of Pneumocystis jirovecii Pneumonia in Patients with Autoimmune and Inflammatory Diseases. Chest 2020, 158, 2323–2332. [Google Scholar] [CrossRef]
- Han, H.; Ma, Q.; Li, C.; Liu, R.; Zhao, L.; Wang, W.; Zhang, P.; Liu, X.; Gao, G.; Liu, F.; et al. Profiling serum cytokines in COVID-19 patients reveals IL-6 and IL-10 are disease severity predictors. Emerg. Microbes Infect. 2020, 9, 1123–1130. [Google Scholar] [CrossRef]
- Oishi, Y.; Manabe, I. Macrophages in inflammation, repair and regeneration. Int. Immunol. 2018, 30, 511–528. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, R.; Shu, R.; Yu, J.; Li, H.; Long, H.; Jin, S.; Li, S.; Hu, Q.; Yao, F.; et al. Study of the Relationship between Microbiome and Colorectal Cancer Susceptibility Using 16SrRNA Sequencing. BioMed Res. Int. 2020, 2020, 7828392. [Google Scholar] [CrossRef]
- Naseribafrouei, A.; Hestad, K.; Avershina, E.; Sekelja, M.; Linløkken, A.; Wilson, R.; Rudi, K. Correlation between the human fecal microbiota and depression. Neurogastroenterol. Motil. 2014, 26, 1155–1162. [Google Scholar] [CrossRef]
- Sbaoui, Y.; Ezaouine, A.; Toumi, M.; Farkas, R.; Kbaich, M.A.; Habbane, M.; El Mouttaqui, S.; Kadiri, F.Z.; El Messal, M.; Tóth, E.; et al. Effect of Climate on Bacterial and Archaeal Diversity of Moroccan Marine Microbiota. Microorganisms 2022, 10, 1622. [Google Scholar] [CrossRef]
- Zou, Y.; Liang, N.; Zhang, X.; Han, C.; Nan, X. Functional differentiation related to decomposing complex carbohydrates of intestinal microbes between two wild zokor species based on 16SrRNA sequences. BMC Vet. Res. 2021, 17, 216. [Google Scholar] [CrossRef]
- Chen, S.; Gu, Z.; Zhang, W.; Jia, S.; Wu, Y.; Zheng, P.; Dai, Y.; Leng, J. Microbiome of the lower genital tract in Chinese women with endometriosis by 16s-rRNA sequencing technique: A pilot study. Ann. Transl. Med. 2020, 8, 1440. [Google Scholar] [CrossRef]
- West, C.E.; Rydén, P.; Lundin, D.; Engstrand, L.; Tulic, M.K.; Prescott, S.L. Gut microbiome and innate immune response patterns in IgE-associated eczema. Clin. Exp. Allergy 2015, 45, 1419–1429. [Google Scholar] [CrossRef] [PubMed]
- Remick, D.G. Old friends: Pneumonia and interleukin-6. Crit. Care Med. 2009, 37, 1809–1810. [Google Scholar] [CrossRef] [PubMed]
- Godkin, A.; Humphreys, I.R. Elevated interleukin-6, interleukin-10 and neutrophil: Lymphocyte ratio as identifiers of severe coronavirus disease 2019. Immunology 2020, 160, 221–222. [Google Scholar] [CrossRef] [PubMed]
- Trevejo-Nunez, G.; Elsegeiny, W.; Aggor, F.E.Y.; Tweedle, J.L.; Kaplan, Z.; Gandhi, P.; Castillo, P.; Ferguson, A.; Alcorn, J.F.; Chen, K.; et al. Interleukin-22 (IL-22) Binding Protein Constrains IL-22 Activity, Host Defense, and Oxidative Phosphorylation Genes during Pneumococcal Pneumonia. Infect. Immun. 2019, 87, e00550-19. [Google Scholar] [CrossRef]
- Steck, P.; Ritzmann, F.; Honecker, A.; Vella, G.; Herr, C.; Gaupp, R.; Bischoff, M.; Speer, T.; Tschernig, T.; Bals, R.; et al. Interleukin 17 Receptor E (IL-17RE) and IL-17C Mediate the Recruitment of Neutrophils during Acute Streptococcus pneumoniae Pneumonia. Infect. Immun. 2019, 87, e00329-19. [Google Scholar] [CrossRef]
- Nagashio, S.; Ajima, K.; Maejima, D.; Sanjo, H.; Kajihara, R.; Hayashi, M.; Watanabe-Asaka, T.; Kaidoh, M.; Yokoyama, Y.; Taki, S.; et al. Water intake increases mesenteric lymph flow and the total flux of albumin, long-chain fatty acids, and IL-22 in rats: New concept of absorption in jejunum. Am. J. Physiol. Liver Physiol. 2019, 316, G155–G165. [Google Scholar] [CrossRef]
- Bradley, J.S.; Byington, C.L.; Shah, S.S.; Alverson, B.; Carter, E.R.; Harrison, C.; Kaplan, S.L.; Mace, S.E.; McCracken, G.H., Jr.; Moore, M.R.; et al. The management of community-acquired pneumonia in infants and children older than 3 months of age: Clinical practice guidelines by the Pediatric Infectious Diseases Society and the Infectious Diseases Society of America. Clin. Infect. Dis. 2011, 53, e25–e76. [Google Scholar] [CrossRef] [PubMed]
- le Roux, D.M.; Zar, H.J. Community-acquired pneumonia in children—A changing spectrum of disease. Pediatr. Radiol. 2017, 47, 1392–1398. [Google Scholar] [CrossRef]
- Eslamy, H.K.; Newman, B. Pneumonia in Normal and Immunocompromised Children: An Overview and Update. Radiol. Clin. North Am. 2011, 49, 895–920. [Google Scholar] [CrossRef]
- Tang, J.; Xu, L.; Zeng, Y.; Gong, F. Effect of gut microbiota on LPS-induced acute lung injury by regu-lating the TLR4/NF-kB signaling pathway. Int. Immunopharmacol. 2021, 91, 107272. [Google Scholar] [CrossRef]
- Dejea, C.; Wick, E.; Sears, C.L. Bacterial oncogenesis in the colon. Futur. Microbiol. 2013, 8, 445–460. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.R.; Hoyles, L.; Flint, H.J.; Dumas, M.-E. Colonic bacterial metabolites and human health. Curr. Opin. Microbiol. 2013, 16, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Luis, A.S.; Jin, C.; Pereira, G.V.; Glowacki, R.W.P.; Gugel, S.R.; Singh, S.; Byrne, D.P.; Pudlo, N.A.; London, J.A.; Baslé, A.; et al. A single sulfatase is required to access colonic mucin by a gut bacterium. Nature 2021, 598, 332–337. [Google Scholar] [CrossRef]
- Momozawa, Y.; Deffontaine, V.; Louis, E.; Medrano, J.F. Characterization of Bacteria in Biopsies of Colon and Stools by High Throughput Sequencing of the V2 Region of Bacterial 16S rRNA Gene in Human. PLoS ONE 2011, 6, e16952. [Google Scholar] [CrossRef]
- Esseku, F.; Adeyeye, M.C. Bacteria and pH-sensitive polysaccharide-polymer films for colon targeted delivery. Crit. Rev. Ther. Drug Carr. Syst. 2011, 28, 395–445. [Google Scholar] [CrossRef]
- Arnoldini, M.; Cremer, J.; Hwa, T. Bacterial growth, flow, and mixing shape human gut microbiota density and composition. Gut Microbes 2018, 9, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Loman, B.R.; Jordan, K.R.; Haynes, B.; Bailey, M.T.; Pyter, L.M. Chemotherapy-induced neuroinflamma-tion is associated with disrupted colonic and bacterial homeostasis in female mice. Sci. Rep. 2019, 9, 16490. [Google Scholar] [CrossRef]
- Han, Y.; Duan, X.; Yang, L.; Nilsson-Payant, B.E.; Wang, P.; Duan, F.; Tang, X.; Yaron, T.M.; Zhang, T.; Uhl, S.; et al. Identification of SARS-CoV-2 inhibitors using lung and colonic organoids. Nature 2021, 589, 270–275. [Google Scholar] [CrossRef]
- Sharifianjazi, F.; Rad, A.J.; Bakhtiari, A.; Niazvand, F.; Esmaeilkhanian, A.; Bazli, L.; Abniki, M.; Irani, M.; Moghanian, A. Biosensors and nanotechnology for cancer diagnosis (lung and bronchus, breast, prostate, and colon): A systematic review. Biomed. Mater. 2021, 17, 012002. [Google Scholar] [CrossRef]
- Kriegsmann, M.; Longuespée, R.; Wandernoth, P.; Mohanu, C.; Lisenko, K.; Weichert, W.; Warth, A.; Dienemann, H.; De Pauw, E.; Katzenberger, T.; et al. Typing of colon and lung adenocarcinoma by high throughput imaging mass spectrometry. Biochim. Biophys. Acta Proteins Proteom. 2017, 1865, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Moshensky, A.; Brand, C.S.; Alhaddad, H.; Shin, J.; Masso-Silva, J.A.; Advani, I.; Gunge, D.; Sharma, A.; Mehta, S.; Jahan, A.; et al. Effects of mango and mint pod-based e-cigarette aerosol inhalation on inflammatory states of the brain, lung, heart, and colon in mice. eLife 2022, 11, e67621. [Google Scholar] [CrossRef] [PubMed]
- Valitutti, F.; Fasano, A. Breaking Down Barriers: How Understanding Celiac Disease Pathogenesis Informed the Development of Novel Treatments. Dig. Dis. Sci. 2019, 64, 1748–1758. [Google Scholar] [CrossRef] [PubMed]
- Kuo, W.-T.; Zuo, L.; Odenwald, M.A.; Madha, S.; Singh, G.; Gurniak, C.B.; Abraham, C.; Turner, J.R. The Tight Junction Protein ZO-1 Is Dispensable for Barrier Function but Critical for Effective Mucosal Repair. Gastroenterology 2021, 161, 1924–1939. [Google Scholar] [CrossRef] [PubMed]
- Liu, W. Retraction Note: Long non-coding RNA VPS9D1-AS1 promotes growth of colon adeno-carcinoma by sponging miR-1301-3p and CLDN1. Hum Cell. 2022, 35, 2030. [Google Scholar] [CrossRef]
- Chou, S.C.; Ko, H.W.; Lin, Y.C. CRP/IL-6/IL-10 Single-Nucleotide Polymorphisms Correlate with the Susceptibility and Severity of Community-Acquired Pneumonia. Genet. Test Mol. Biomark. 2016, 20, 732–740. [Google Scholar] [CrossRef]
- Xiang, X.; Feng, D.; Hwang, S.; Ren, T.; Wang, X.; Trojnar, E.; Matyas, C.; Mo, R.; Shang, D.; He, Y.; et al. Interleukin-22 ameliorates acute-on-chronic liver failure by reprogramming impaired regeneration pathways in mice. J. Hepatol. 2020, 72, 736–745. [Google Scholar] [CrossRef]
- Lei, L.; Zhao, C.; Qin, F.; He, Z.-Y.; Wang, X.; Zhong, X.-N. Th17 cells and IL-17 promote the skin and lung inflammation and fibrosis process in a bleomycin-induced murine model of systemic sclerosis. Clin. Exp. Rheumatol. 2016, 34, 14–22. [Google Scholar]
- Leoney, A.; Karthigeyan, S.; Asharaf, A.S.; Felix, A.J.W. Detection and categorization of biofilm-forming Staphylococcus aureus, Viridans streptococcus, Klebsiella pneumoniae, and Escherichia coli isolated from complete denture patients and visualization using scanning electron microscopy. J. Int. Soc. Prev. Community Dent. 2020, 10, 627–633. [Google Scholar] [CrossRef]
- Baum, B.; Liebler-Tenorio, E.M.; Enß, M.-L.; Pohlenz, J.F.; Breves, G. Saccharomyces boulardii and Bacillus cereus Var. Toyoi Influence the Morphology and the Mucins of the Intestine of Pigs. Z. Gastroenterol. 2002, 40, 277–284. [Google Scholar] [CrossRef]
- Pengpeng, W.; Tan, Z. Ammonia Assimilation in Rumen Bacteria: A Review. Anim. Biotechnol. 2013, 24, 107–128. [Google Scholar] [CrossRef] [PubMed]
- Longman, R.S.; Littman, D.R. The functional impact of the intestinal microbiome on mucosal immunity and systemic autoimmunity. Curr. Opin. Rheumatol. 2015, 27, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Sánchez, B.P.; Cruz-Zárate, D.; Estrada-García, I.; Wong-Baeza, I. Innate lymphoid cells and their role in immune response regulation. Rev. Alerg. Mex. 2017, 64, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Ronan, V.; Yeasin, R.; Claud, E.C. Childhood Development and the Microbiome-The Intestinal Microbiota in Maintenance of Health and Development of Disease During Childhood Development. Gastroenterology 2021, 160, 495–506. [Google Scholar] [CrossRef]
- Yamanaka, K.; Yamamoto, O.; Honda, T. Pathophysiology of psoriasis: A review. J. Dermatol. 2021, 48, 722–731. [Google Scholar] [CrossRef]
- Raftery, A.L.; Tsantikos, E.; Harris, N.L.; Hibbs, M.L. Links between Inflammatory Bowel Disease and Chronic Obstructive Pulmonary Disease. Front. Immunol. 2020, 11, 2144. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Z.; Deng, T.; Gui, X.; Wang, L.; Yan, Q.; Wang, L. Mechanisms of Lung and Intestinal Microbiota and Innate Immune Changes Caused by Pathogenic Enterococcus Faecalis Promoting the Development of Pediatric Pneumonia. Microorganisms 2023, 11, 2203. https://doi.org/10.3390/microorganisms11092203
Tian Z, Deng T, Gui X, Wang L, Yan Q, Wang L. Mechanisms of Lung and Intestinal Microbiota and Innate Immune Changes Caused by Pathogenic Enterococcus Faecalis Promoting the Development of Pediatric Pneumonia. Microorganisms. 2023; 11(9):2203. https://doi.org/10.3390/microorganisms11092203
Chicago/Turabian StyleTian, Zhiying, Ting Deng, Xuwen Gui, Leilei Wang, Qiulong Yan, and Liang Wang. 2023. "Mechanisms of Lung and Intestinal Microbiota and Innate Immune Changes Caused by Pathogenic Enterococcus Faecalis Promoting the Development of Pediatric Pneumonia" Microorganisms 11, no. 9: 2203. https://doi.org/10.3390/microorganisms11092203