
Screening and Identification of a Streptomyces Strain with Quorum-Sensing Inhibitory Activity and Effect of the Crude Extracts on Virulence Factors of Pseudomonas aeruginosa
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Screening of QS-Inhibiting Active Actinomycetes
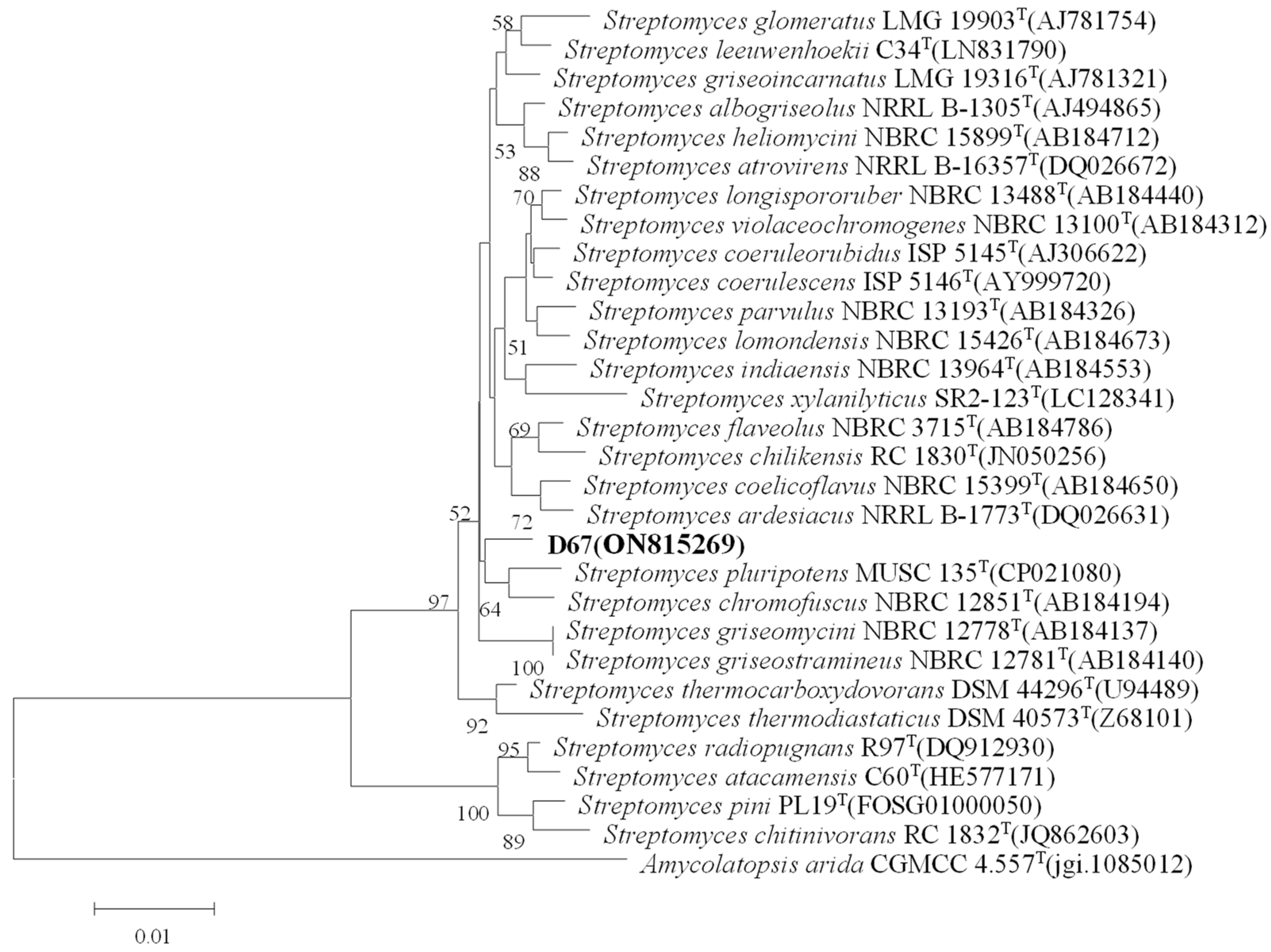
2.3. Morphological Observation and Molecular Identification of Strains
2.4. Preparation and Functional Validation of Crude Extract
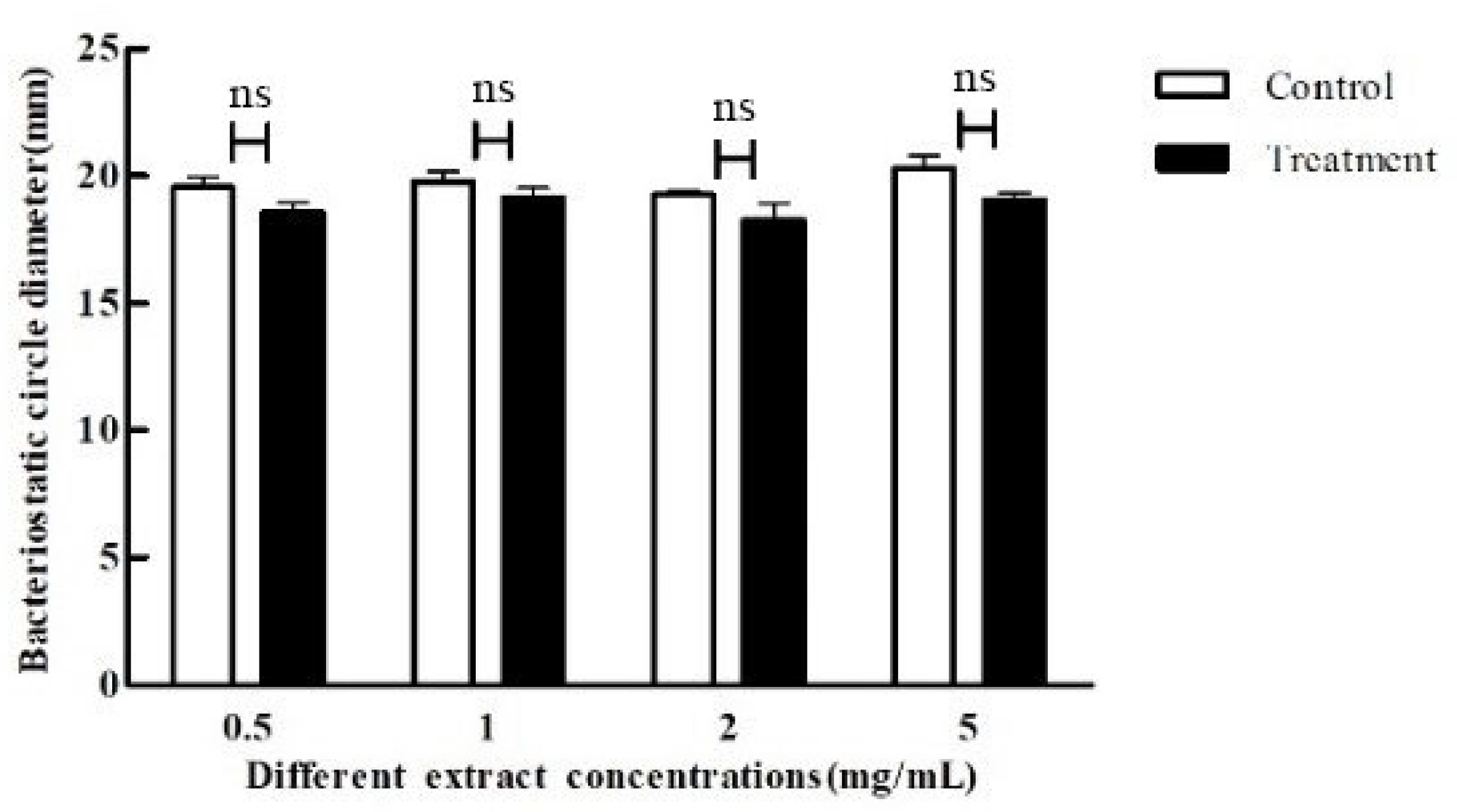
2.5. Effect of Crude Extract on Bacterial Inhibition
2.6. Effect of Crude Extract on the Production of Purple Pigment by C. violaceum
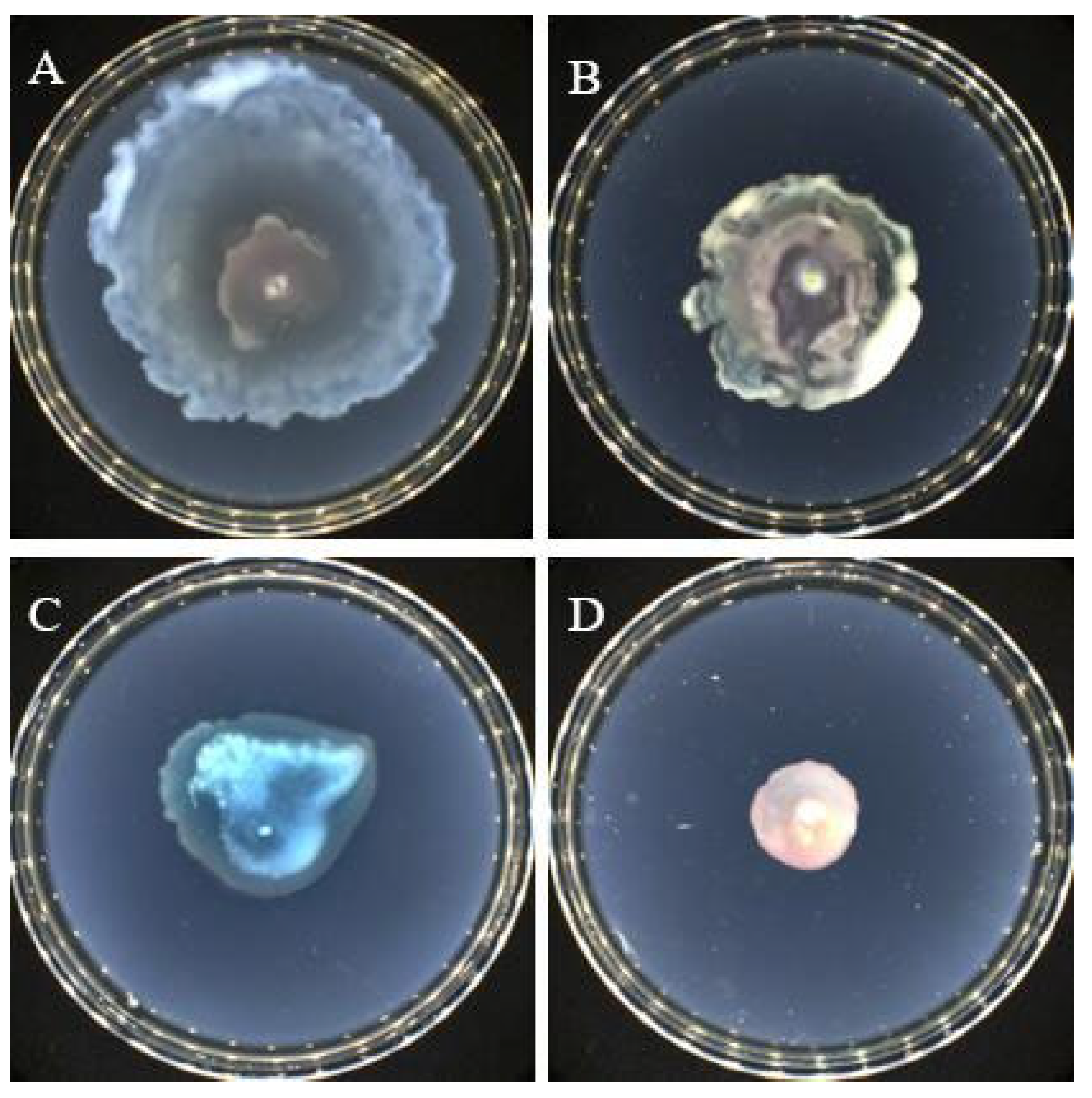
2.7. Effect of Crude Extracts on the Swarming and Swimming Properties of P. aeruginosa
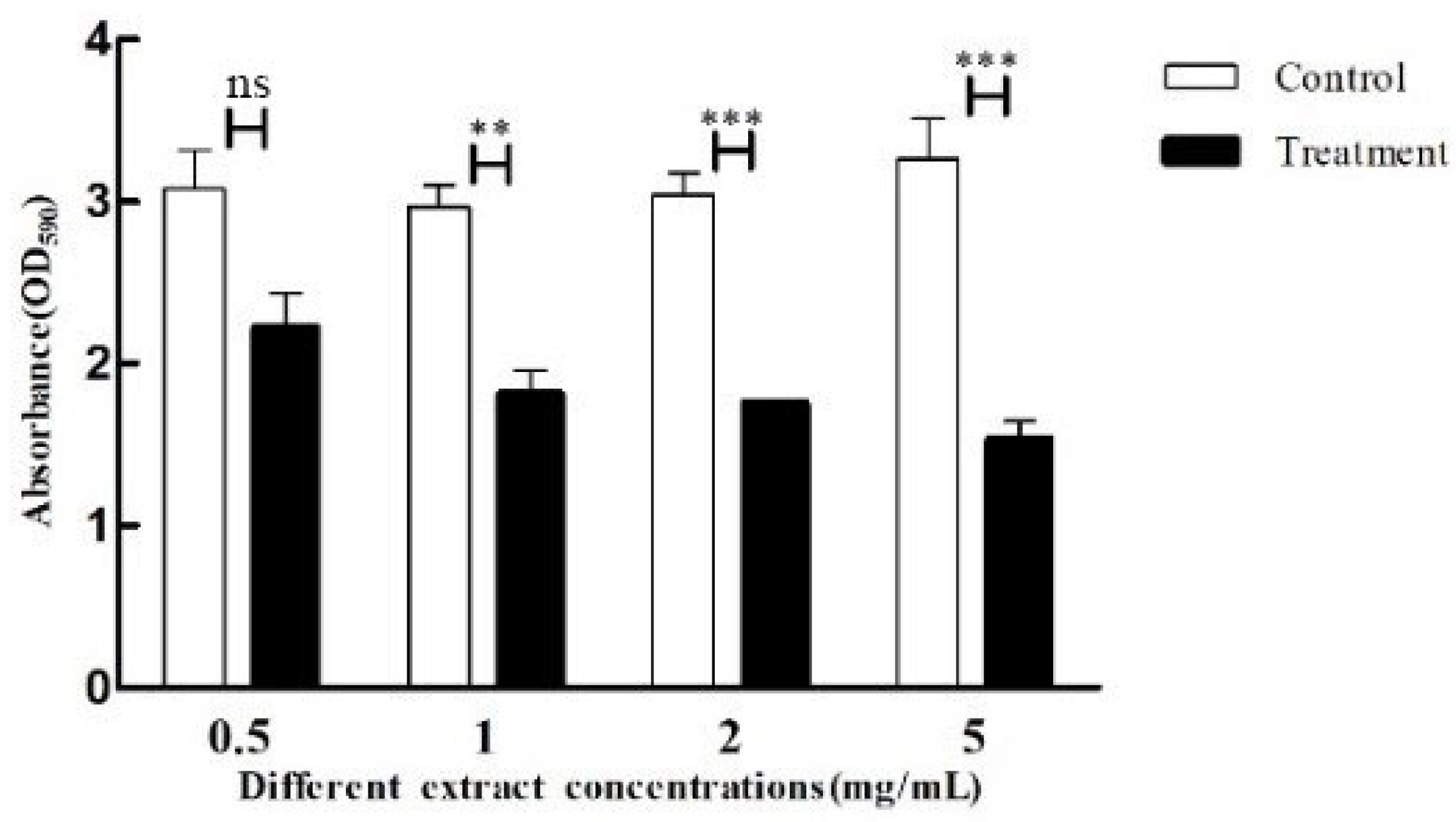
2.8. Effect of Crude Extract on the Biofilm of P. aeruginosa
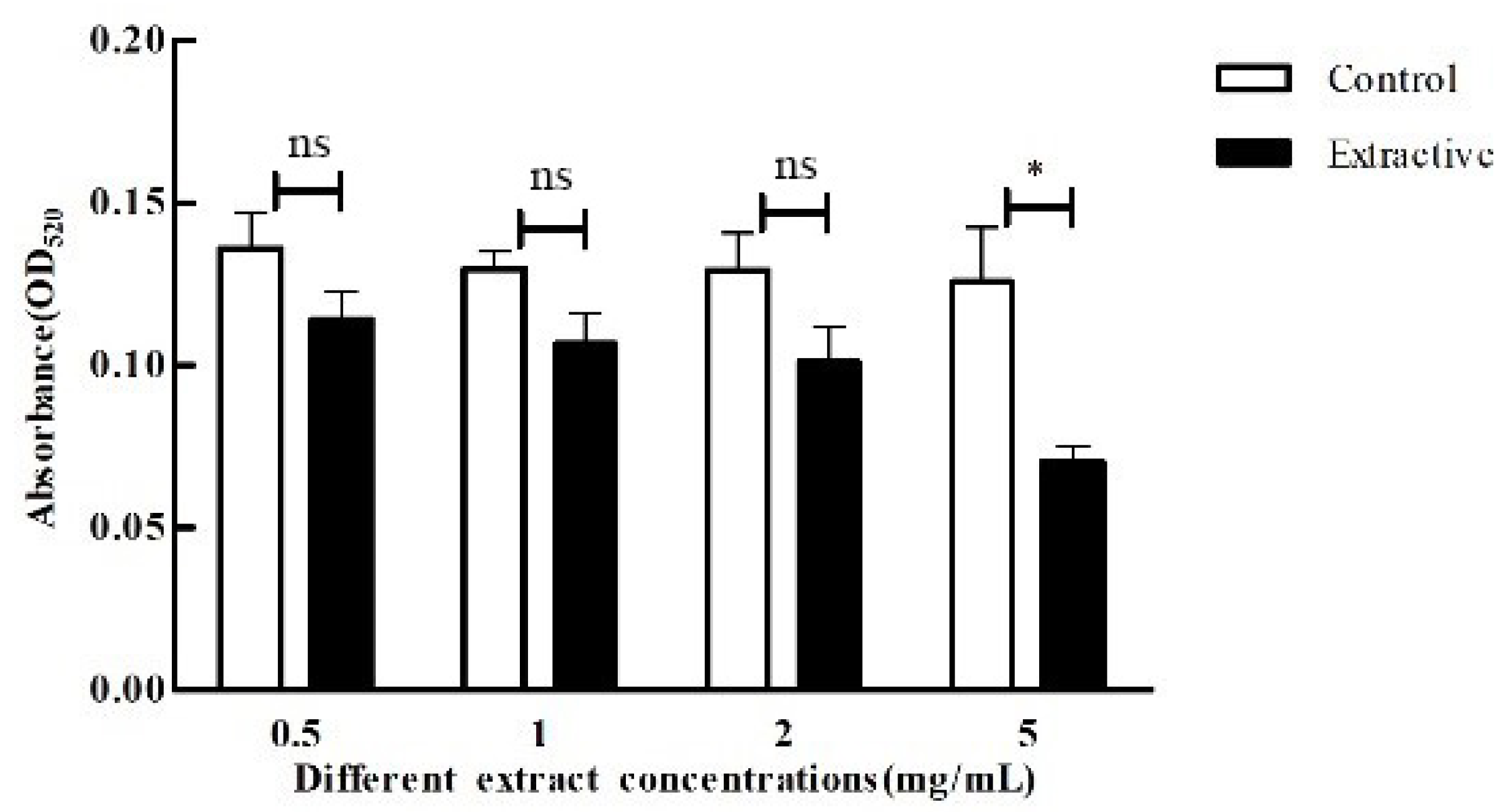
2.9. Effect of Crude Extracts on P. aeruginosa Pyocyanin
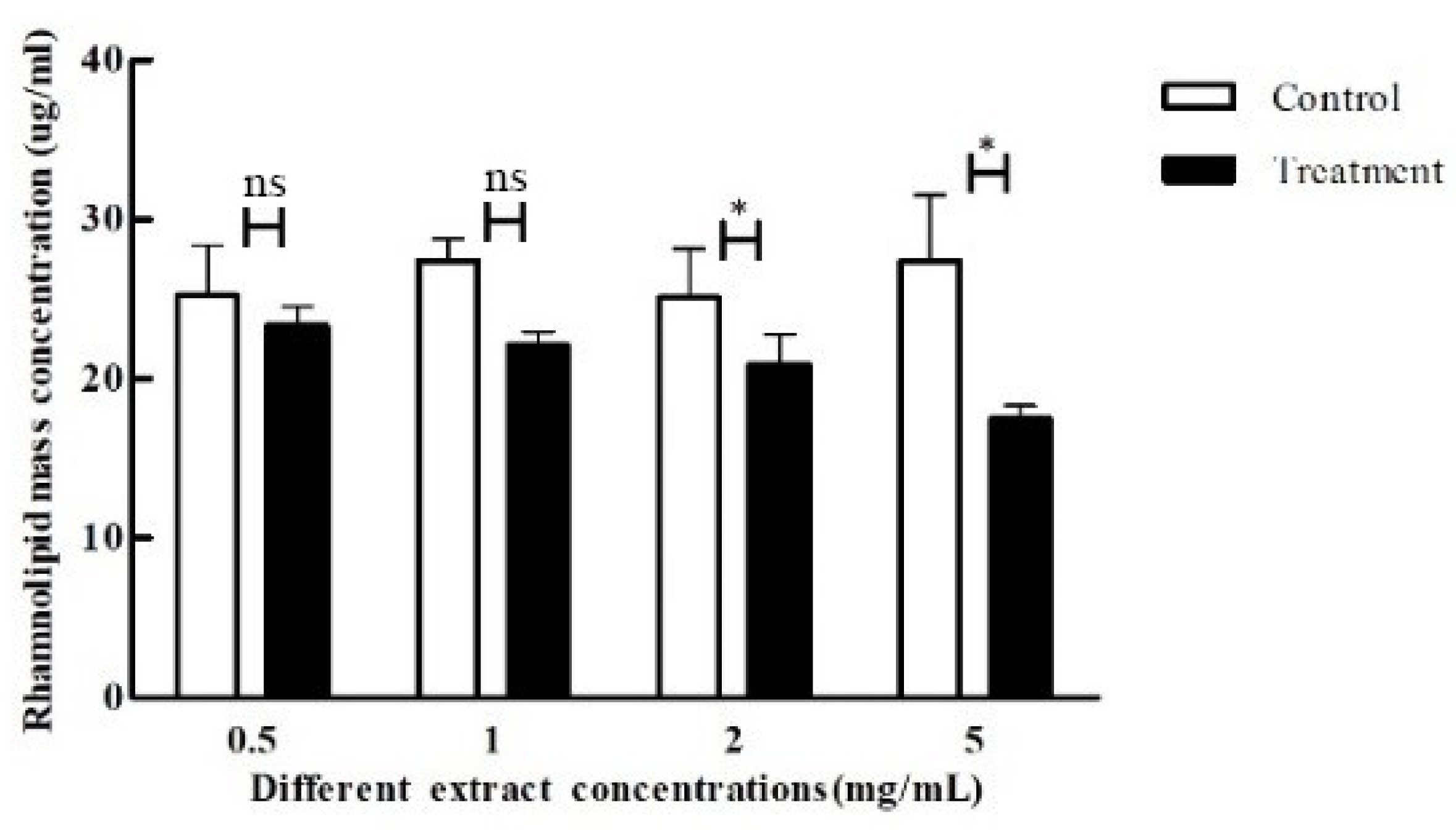
2.10. Determination of Crude Extracts on P. aeruginosa rhamnosus Lipids
2.11. Determination of Total Protease Production
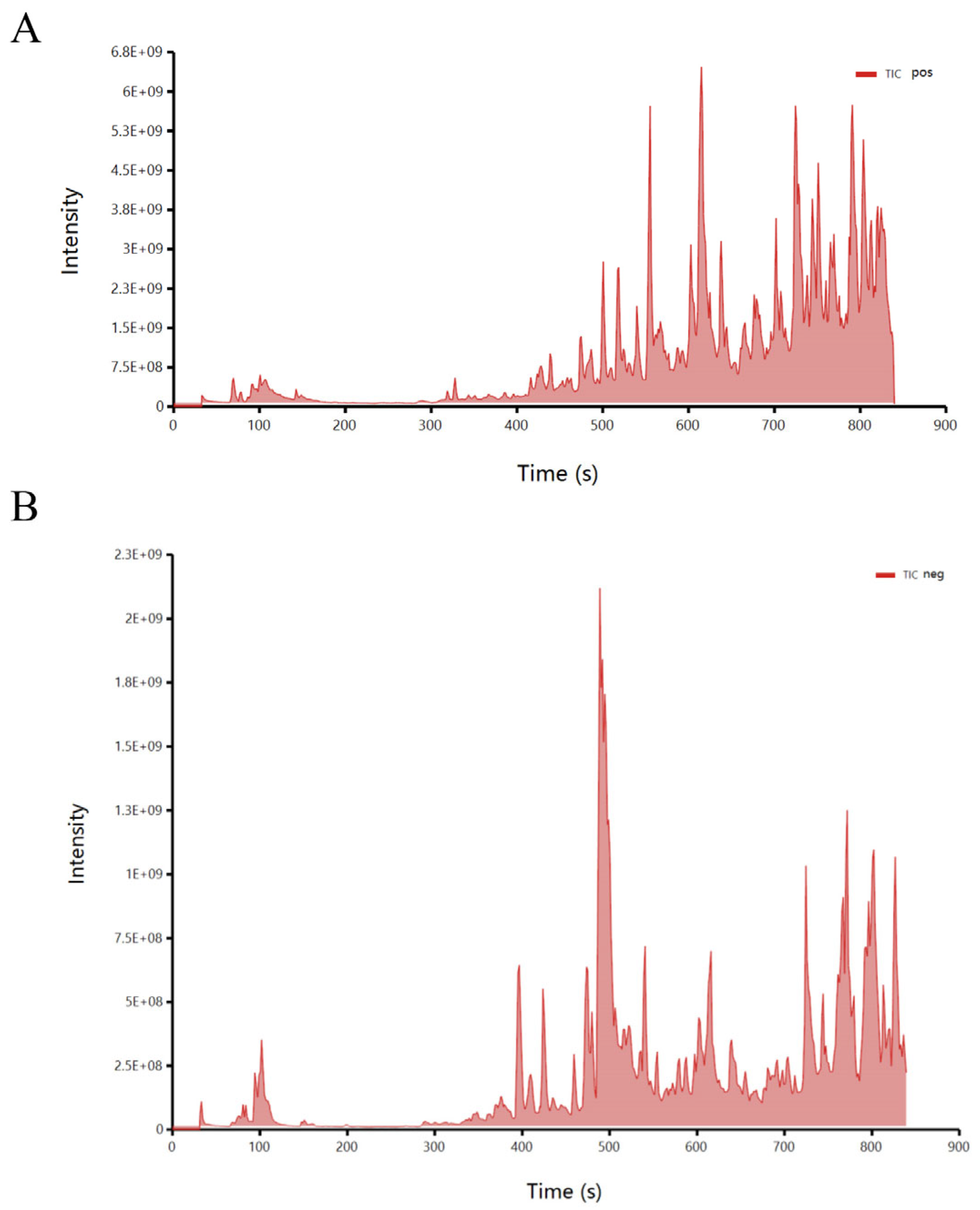
2.12. LC-MS Assay and Validation of Quorum Sensing Inhibitory Activity of Compounds
3. Results
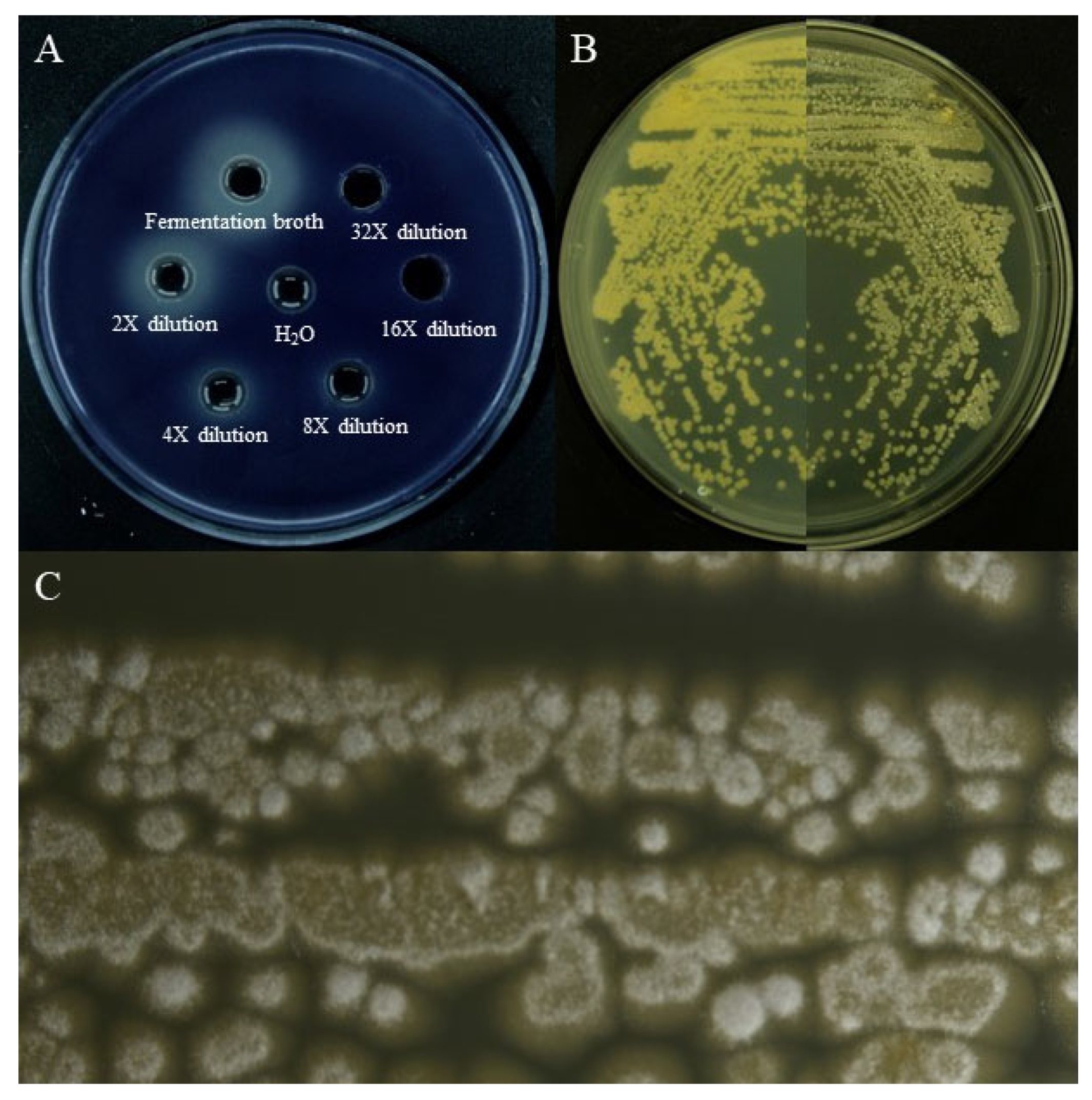
3.1. Screening and Identification of Quorum-Sensing Inhibitory Active Strains
3.2. Validation of the Effect of Crude Extract on Quorum Sensing
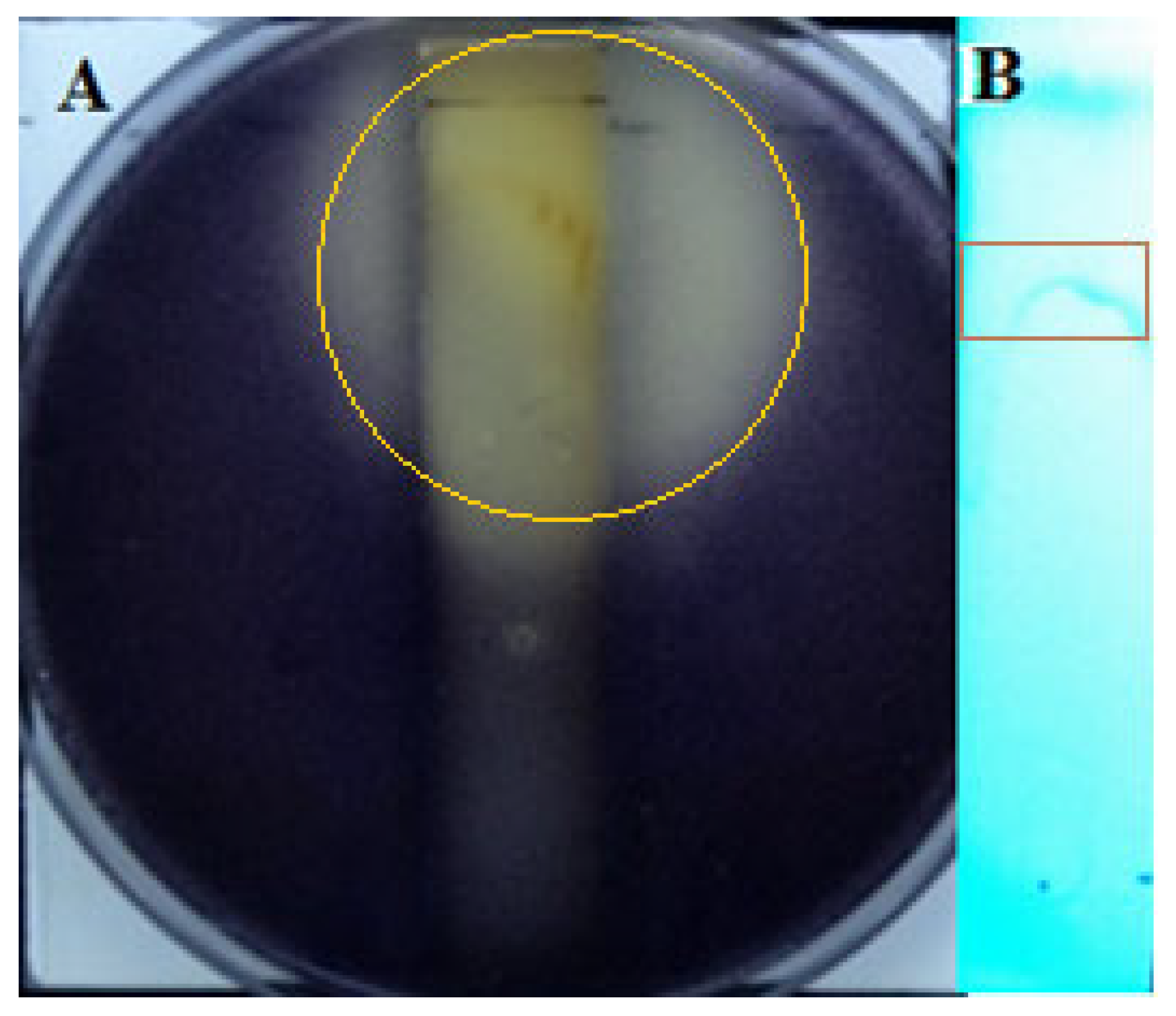
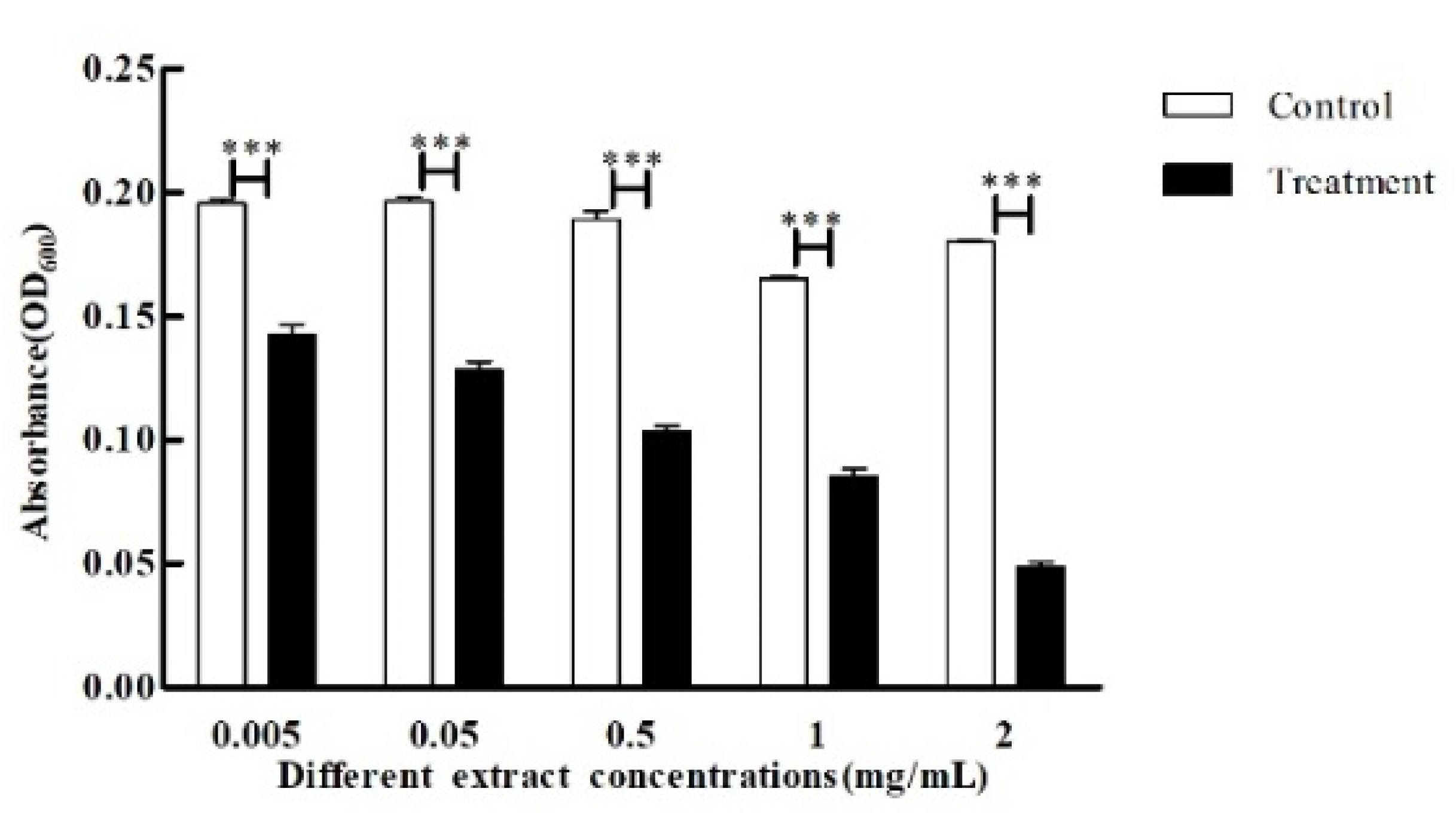
3.3. Effect of Crude Extract on P. aeruginosa Quorum Sensing Inhibition
3.4. Preliminary Analysis of Active Substances in Crude Extracts
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kalia, V.C. Quorum sensing inhibitors: An overview. Biotechnol. Adv. 2013, 31, 224–245. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Cui, F.; Wang, D.; Li, T.; Li, J. Quorum quenching enzyme (pf-1240) capable to degrade ahls as a candidate for inhibiting quorum sensing in food spoilage bacterium Hafnia alvei. Foods 2021, 10, 2700. [Google Scholar] [CrossRef]
- Liu, L.; Yan, Y.; Feng, L.; Zhu, J. Quorum sensing asai mutants affect spoilage phenotypes, motility, and biofilm formation in a marine fish isolate of Aeromonas salmonicida. Food Microbiol. 2018, 76, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Defoirdt, T. Quorum-sensing systems as targets for antivirulence therapy. Trends Microbiol. 2018, 26, 313–328. [Google Scholar] [CrossRef] [PubMed]
- Murray, E.J.; Dubern, J.F.; Chan, W.C.; Chhabra, S.R.; Williams, P. A Pseudomonas aeruginosa PQS quorum-sensing system inhibitor with anti-staphylococcal activity sensitizes polymicrobial biofilms to tobramycin. Cell Chem. Biol. 2022, 29, 1187–1199. [Google Scholar] [CrossRef]
- Mcgrath, S.; Wade, D.S.; Pesci, E.C. Dueling quorum sensing systems in Pseudomonas aeruginosa control the production of the Pseudomonas quinolone signal (PQS). FEMS Microbiol. Lett. 2004, 230, 27–34. [Google Scholar] [CrossRef]
- Girard, G.; Bloemberg, G.V. Central role of quorum sensing in regulating the production of pathogenicity factors in Pseudomonas aeruginosa. Future Microbiol. 2008, 3, 97–106. [Google Scholar] [CrossRef]
- Martinelli, D.; Grossmann, G.; Sequin, U.; Brandl, H.; Bachofen, R. Effects of natural and chemically synthesized furanones on quorum sensing in Chromobacterium violaceum. BMC Microbiol. 2004, 4, 25. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.K.; Mishra, A.; Jha, B. Anti-quorum sensing and anti-biofilm activity of Delftia tsuruhatensis extract by attenuating the quorum sensing-controlled virulence factor production in Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2017, 7, 337. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Liu, X.; Liang, H.; Che, Y.; Chen, C.; Dai, H.; Yu, K.; Liu, M.; Ma, L.; Yang, C.H.; et al. Effects of 14-alpha-lipoyl andrographolide on quorum sensing in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2012, 56, 6088–6094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijerathne, T.D.; Kim, J.H.; Kim, M.J.; Kim, C.Y.; Chae, M.R.; Lee, S.W.; Lee, K.P. Onion peel extract and its constituent, quercetin inhibits human slo3 in a ph and calcium dependent manner. Korean J. Physiol. Pharmacol. 2019, 23, 381–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2022, 39, 1122–1171. [Google Scholar] [CrossRef]
- Chen, J.; Wang, B.; Lu, Y.; Guo, Y.; Sun, J.; Wei, B.; Zhang, H.; Wang, H. Quorum sensing inhibitors from marine microorganisms and their synthetic derivatives. Mar. Drugs 2019, 17, 80. [Google Scholar] [CrossRef] [Green Version]
- Kachhadia, R.; Kapadia, C.; Singh, S.; Gandhi, K.; Jajda, H.; Alfarraj, S.; Ansari, M.J.; Danish, S.; Datta, R. Quorum sensing inhibitory and quenching activity of Bacillus cereus rc1 extracts on soft rot-causing bacteria Lelliottia amnigena. ACS Omega 2022, 7, 25291–25308. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.M.; Campos, C.R. Applications of quartering method in soils and foods. Int. J. Eng. Res. Appl. 2017, 7, 35–39. [Google Scholar] [CrossRef]
- Ding, X.; Zhao, Y.; Wang, W.; Bai, C.; Guan, P. Application of TLC-bioautography in active components screening. Pract. Pharm. Clin. Remedies 2017, 20, 1220–1224. [Google Scholar]
- Choo, J.H.; Rukayadi, Y.; Hwang, J.K. Inhibition of bacterial quorum sensing by vanilla extract. Lett. Appl. Microbiol. 2006, 42, 637–641. [Google Scholar] [CrossRef]
- Kapadia, C.; Kachhdia, R.; Singh, S.; Gandhi, K.; Poczai, P.; Alfarraj, S.; Ansari, M.J.; Gafur, A.; Sayyed, R.Z. Pseudomonas aeruginosa inhibits quorum-sensing mechanisms of soft rot pathogen Lelliottia amnigena RCE to regulate its virulence factors and biofilm formation. Front. Microbiol. 2022, 13, 977669. [Google Scholar] [CrossRef]
- Zhang, J.; Rui, X.; Wang, L.; Guan, Y.; Sun, X.; Dong, M. Polyphenolic extract from rosa rugosa tea inhibits bacterial quorum sensing and biofilm formation. Food Control 2014, 42, 125–131. [Google Scholar] [CrossRef]
- Miao, J.; Chi, X.; Wang, Y.; Feng, Z.; Xue, W.; Huang, R.; Zhang, H.; Tian, L.; Zhang, H.; Zhai, J.; et al. Regulation of pyocyanin biosynthesis by transcriptional factor sigma38 in Pseudomonas aeruginosa pao1. Acta Microbiol. Sin. 2017, 57, 229–239. [Google Scholar]
- Lou, Z.; Wang, H.; Tang, Y.; Chen, X. The effect of burdock leaf fraction on adhesion, biofilm formation, quorum sensing and virulence factors of Pseudomonas aeruginosa. J. Appl. Microbiol. 2017, 122, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Zelena, E.; Dunn, W.B.; Broadhurst, D.; Francis-Mcintyre, S.; Carroll, K.M.; Begley, P.; O’Hagan, S.; Knowles, J.D.; Halsall, A.; Wilson, I.D.; et al. Development of a robust and repeatable UPLC-MS method for the long-term metabolomic study of human serum. Anal. Chem. 2009, 81, 1357–1364. [Google Scholar] [CrossRef]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, I.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J.K. Global metabolic profiling of animal and human tissues via UPLC-MS. Nat. Protoc. 2013, 8, 17–32. [Google Scholar] [CrossRef]
- Nouioui, I.; Carro, L.; Garcia-Lopez, M.; Meier-Kolthoff, J.P.; Woyke, T.; Kyrpides, N.C.; Pukall, R.; Klenk, H.P.; Goodfellow, M.; Goker, M. Genome-based taxonomic classification of the phylum Actinobacteria. Front. Microbiol. 2018, 9, 2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Shi, Y.L.; Wang, H.; Zhang, T.; Yu, L.Y.; Sun, H.; Zhang, Y.Q. Diversity of bacteria and the characteristics of Actinobacteria community structure in Badain Jaran desert and Tengger desert of China. Front. Microbiol. 2018, 9, 1068. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16s rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Zhang, F.; Zhou, L.; Kong, F.; Ma, Q.; Xie, Q.; Li, J.; Dai, H.; Guo, L.; Zhao, Y. Altertoxins with quorum sensing inhibitory activities from the marine-derived fungus Cladosporium sp. Kfd33. Mar. Drugs 2020, 18, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.E.; Han, J.W.; Jeon, B.J.; Kim, B.S. Efficacies of quorum sensing inhibitors, piericidin A and glucopiericidin A, produced by Streptomyces xanthocidicus kpp01532 for the control of potato soft rot caused by Erwinia carotovora subsp. atroseptica. Microbiol. Res. 2016, 184, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Shaaban, M.I.; Abdel, B.F.; El-Mahdy, A.M.; Shokralla, S. Quorum sensing inhibiting activity of Streptomyces coelicoflavus isolated from soil. Front. Microbiol. 2016, 7, 659. [Google Scholar] [CrossRef] [Green Version]
- Ooka, K.; Fukumoto, A.; Yamanaka, T.; Shimada, K.; Ishihara, R.; Anzai, Y.; Kato, F. Piericidins, novel quorum-sensing inhibitors against Chromobacterium violaceum cv026, from Streptomyces sp. TOHO-Y209 and TOHO-O348. Open J. Med. Chem. 2013, 3, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Jiang, Y.; Xu, Y.; Qian, S.; Miao, L. Research on quorum sensing inhibitory activity and culture condition of a marine Streptomyces parvulus. Biotechnol. Bull. 2019, 35, 137–143. [Google Scholar]
- Ishaque, N.M.; Burgsdorf, I.; Limlingan, M.J.; Saha, S.; Teta, R.; Ewe, D.; Kannabiran, K.; Hrouzek, P.; Steindler, L.; Costantino, V.; et al. Isolation, genomic and metabolomic characterization of Streptomyces tendae VITAKN with quorum sensing inhibitory activity from southern India. Microorganisms 2020, 8, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velasco-Bucheli, R.; Hormigo, D.; Fernández-Lucas, J.; Torres-Ayuso, P.; Alfaro-Ureña, Y.; Saborido, A.I.; Serrano-Aguirre, L.; García, J.L.; Ramón, F.; Acebal, C.; et al. Penicillin acylase from Streptomyces lavendulae and aculeacin A acylase from Actinoplanes utahensis: Two versatile enzymes as useful tools for quorum quenching processes. Catalysts 2020, 10, 730. [Google Scholar] [CrossRef]
- O’Loughlin, C.T.; Miller, L.C.; Siryaporn, A.; Drescher, K.; Semmelhack, M.F.; Bassler, B.L. A quorum-sensing inhibitor blocks Pseudomonas aeruginosa virulence and biofilm formation. Proc. Natl. Acad. Sci. USA 2013, 110, 17981–17986. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Zhou, J.; Zhu, X.; Yu, S.; Che, L.; Jin, H.; Cai, Z. Strain identification and quorum sensing inhibition characterization of marine-derived Rhizobium sp. Nao1. R. Soc. Open Sci. 2017, 4, 170025. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Kang, H.O.; Jang, H.S.; Lee, J.K.; Koo, B.T.; Yum, D.Y. Identification of extracellular n-acylhomoserine lactone acylase from a Streptomyces sp. and its application to quorum quenching. Appl. Environ. Microbiol. 2005, 71, 2632–2641. [Google Scholar] [CrossRef] [Green Version]
- Nithya, C.; Aravindraja, C.; Pandian, S.K. Bacillus pumilus of Palk bay origin inhibits quorum-sensing-mediated virulence factors in gram-negative bacteria. Res. Microbiol. 2010, 161, 293–304. [Google Scholar] [CrossRef]
- Jakobsen, T.H.; van Gennip, M.; Phipps, R.K.; Shanmugham, M.S.; Christensen, L.D.; Alhede, M.; Skindersoe, M.E.; Rasmussen, T.B.; Friedrich, K.; Uthe, F.; et al. Ajoene, a sulfur-rich molecule from garlic, inhibits genes controlled by quorum sensing. Antimicrob. Agents Chemother. 2012, 56, 2314–2325. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Deng, Y.; Wang, H.; Liu, W.; Zhuang, X.; Chu, W. Corrigendum: Tea polyphenols as an antivirulence compound disrupt quorum-sensing regulated pathogenicity of Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 17987. [Google Scholar] [CrossRef]
- Packiavathy, I.A.; Priya, S.; Pandian, S.K.; Ravi, A.V. Inhibition of biofilm development of uropathogens by curcumin—An anti-quorum sensing agent from Curcuma longa. Food Chem. 2014, 148, 453–460. [Google Scholar] [CrossRef]
- Kwan, J.C.; Meickle, T.; Ladwa, D.; Teplitski, M.; Paul, V.; Luesch, H. Lyngbyoic acid, a “tagged” fatty acid from a marine cyanobacterium, disrupts quorum sensing in Pseudomonas aeruginosa. Mol. Biosyst. 2011, 7, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Abd-Alla, M.H.; Bashandy, S.R. Production of quorum sensing inhibitors in growing onion bulbs infected with Pseudomonas aeruginosa E (hq324110). ISRN Microbiol. 2012, 2012, 161890. [Google Scholar] [CrossRef] [Green Version]
- Lequette, Y.; Lee, J.H.; Ledgham, F.; Lazdunski, A.; Greenberg, E.P. A distinct QSCR regulon in the Pseudomonas aeruginosa quorum-sensing circuit. J. Bacteriol. 2006, 188, 3365–3370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.J.; Zhou, J.W.; Zhang, P.P.; Luo, H.Z.; Tang, S.; Li, J.J.; Deng, S.M.; Jia, A.Q. Quorum sensing inhibition and tobramycin acceleration in Chromobacterium violaceum by two natural cinnamic acid derivatives. Appl. Microbiol. Biotechnol. 2020, 104, 5025–5037. [Google Scholar] [CrossRef]
- He, Z.; Jiang, W.; Jiang, Y.; Dong, J.; Song, Z.; Xu, J.; Zhou, W. Anti-biofilm activities of coumarin as quorum sensing inhibitor for Porphyromonas gingivalis. J. Oral Microbiol. 2022, 14, 2055523. [Google Scholar] [CrossRef]
- Wang, J.; Jiao, H.; Meng, J.; Qiao, M.; Du, H.; He, M.; Ming, K.; Liu, J.; Wang, D.; Wu, Y. Baicalin inhibits biofilm formation and the quorum-sensing system by regulating the MsrA drug efflux pump in Staphylococcus saprophyticus. Front. Microbiol. 2019, 10, 2800. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration of Crude Extracts (mg/mL) | C. violaceum CV026 | P. aeruginosa |
|---|---|---|
| 20 | − | − |
| 10 | − | W |
| 5 | W | + |
| 2 | + | + |
| 1 | + | + |
| 0.5 | + | + |
| 0.05 | + | + |
| 0.005 | + | + |
| Compound | Inhibition Area (cm2) | Violacein Inhibition Area (cm2) |
|---|---|---|
| Eugenol | 2.27 | 16.59 |
| 4-Allylphenol | 2.27 | 10.93 |
| Isoeugenol | 1.54 | 6.5 |
| Cinnamaldehyde | 2.84 | 6.24 |
| Methyl 3-formylbenzoate | - | 4.52 |
| Caffeine | - | 4.15 |
| Cinnamic acid | - | 4.15 |
| 1,2,3,4-Tetrahydroa cridin-9-amine | 2.54 | 3.62 |
| trans-o-Coumaric acid | - | 3.46 |
| Baicalein | - | 3.14 |
| Vanillic acid | - | 3.14 |
| Coniferyl alcohol | 2.54 | 2.77 |
| 3-Indolylacetic acid | - | 2.54 |
| (4-Ethoxyphenyl)urea | - | 2.54 |
| 6-Gingerol | - | 1.77 |
| Pyrrole-2-carboxylic acid | 0.2 | 1.57 |
| 3,4-Dihydroxyhydro cinnamic acid | 0.2 | 1.57 |
| Coumarin | - | 1.54 |
| DL-4-Hydroxyphenyllactic acid | - | 0.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Sun, Y.; Yi, Y.; Bai, X.; Zhu, L.; Zhu, J.; Gu, M.; Zhu, Y.; Jiang, L. Screening and Identification of a Streptomyces Strain with Quorum-Sensing Inhibitory Activity and Effect of the Crude Extracts on Virulence Factors of Pseudomonas aeruginosa. Microorganisms 2023, 11, 2079. https://doi.org/10.3390/microorganisms11082079
Zhang Z, Sun Y, Yi Y, Bai X, Zhu L, Zhu J, Gu M, Zhu Y, Jiang L. Screening and Identification of a Streptomyces Strain with Quorum-Sensing Inhibitory Activity and Effect of the Crude Extracts on Virulence Factors of Pseudomonas aeruginosa. Microorganisms. 2023; 11(8):2079. https://doi.org/10.3390/microorganisms11082079
Chicago/Turabian StyleZhang, Zhidong, Yang Sun, Yuanyang Yi, Xiaoyu Bai, Liying Zhu, Jing Zhu, Meiying Gu, Yanlei Zhu, and Ling Jiang. 2023. "Screening and Identification of a Streptomyces Strain with Quorum-Sensing Inhibitory Activity and Effect of the Crude Extracts on Virulence Factors of Pseudomonas aeruginosa" Microorganisms 11, no. 8: 2079. https://doi.org/10.3390/microorganisms11082079