
New Pseudomonas Bacterial Strains: Biological Activity and Characteristic Properties of Metabolites
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial and Fungal Strains
2.2. Chemicals and Reagents
2.3. Growth Conditions
2.4. Antagonistic and Plant Growth Promotion Activity of Indigenous Pseudomonas Strains
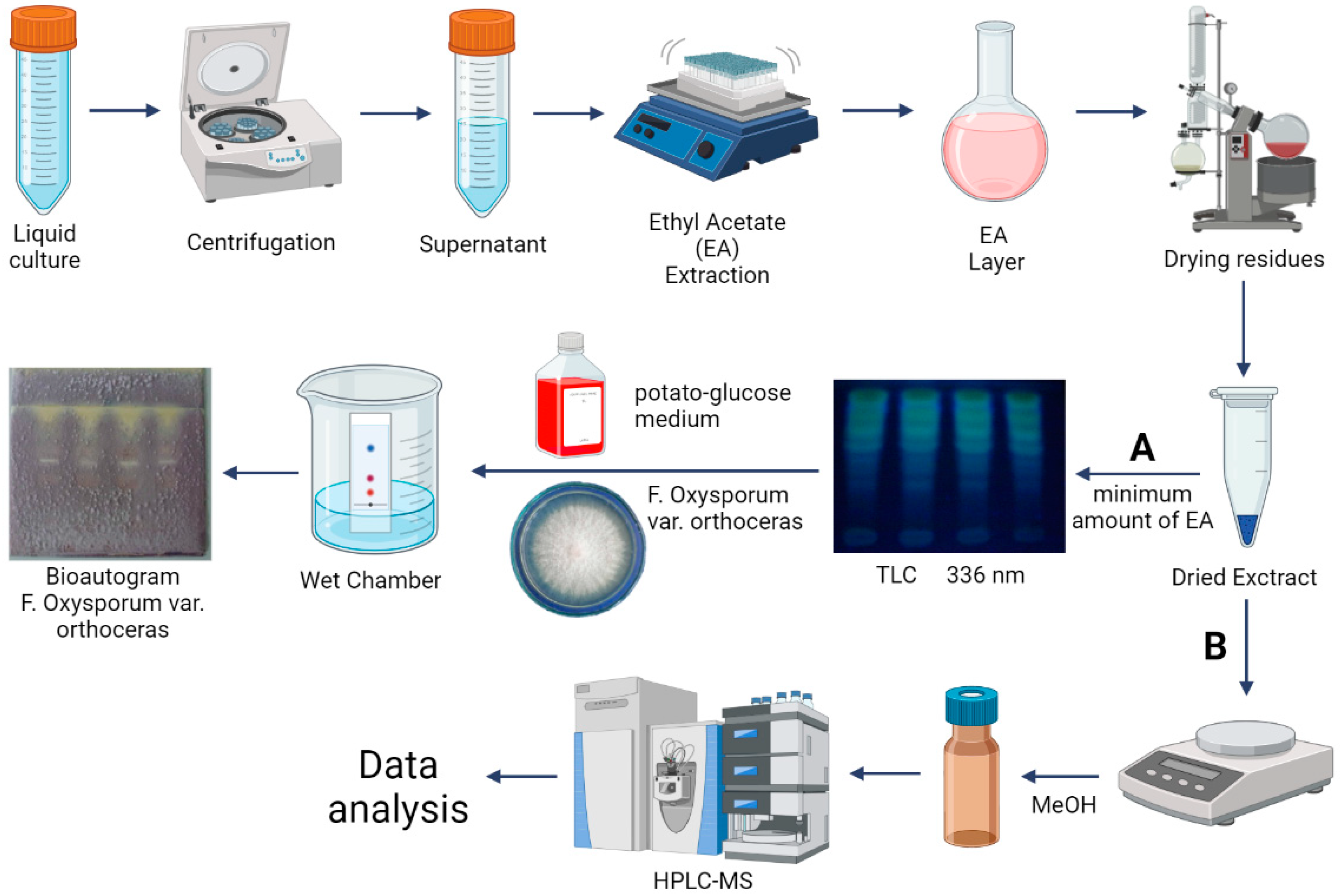
2.5. Sample Preparation
2.6. Isolation and Purification of Bacterial Exometabolites
2.7. Evaluation of the Antifungal Activity of Bacterial Metabolites by Bioautography
2.8. HPLC-MS Conditions
2.9. Quantitative Analysis of Phenazine Nature Compounds
2.10. HPLC-MS Data Analysis
3. Results
3.1. Antagonistic and Plant Growth Promotion Activity of Indigenous Pseudomonas Strains
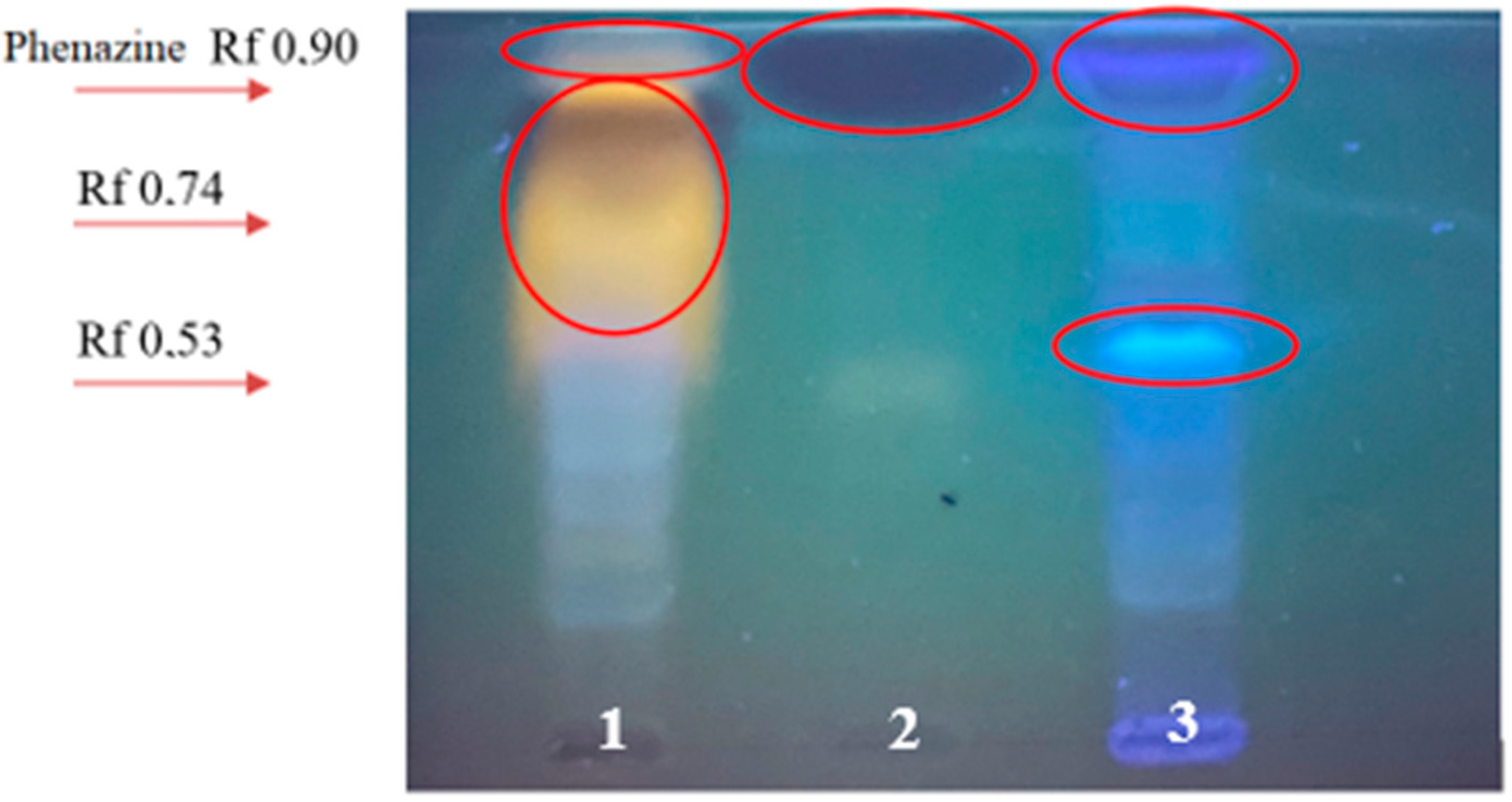
3.2. Isolation of Bacterial Metabolites by TLC
3.3. Preliminary Analysis of the Structure of Bacterial Metabolites
3.4. Evaluation of the Antifungal Activity of Bacterial Metabolites by Bioautography
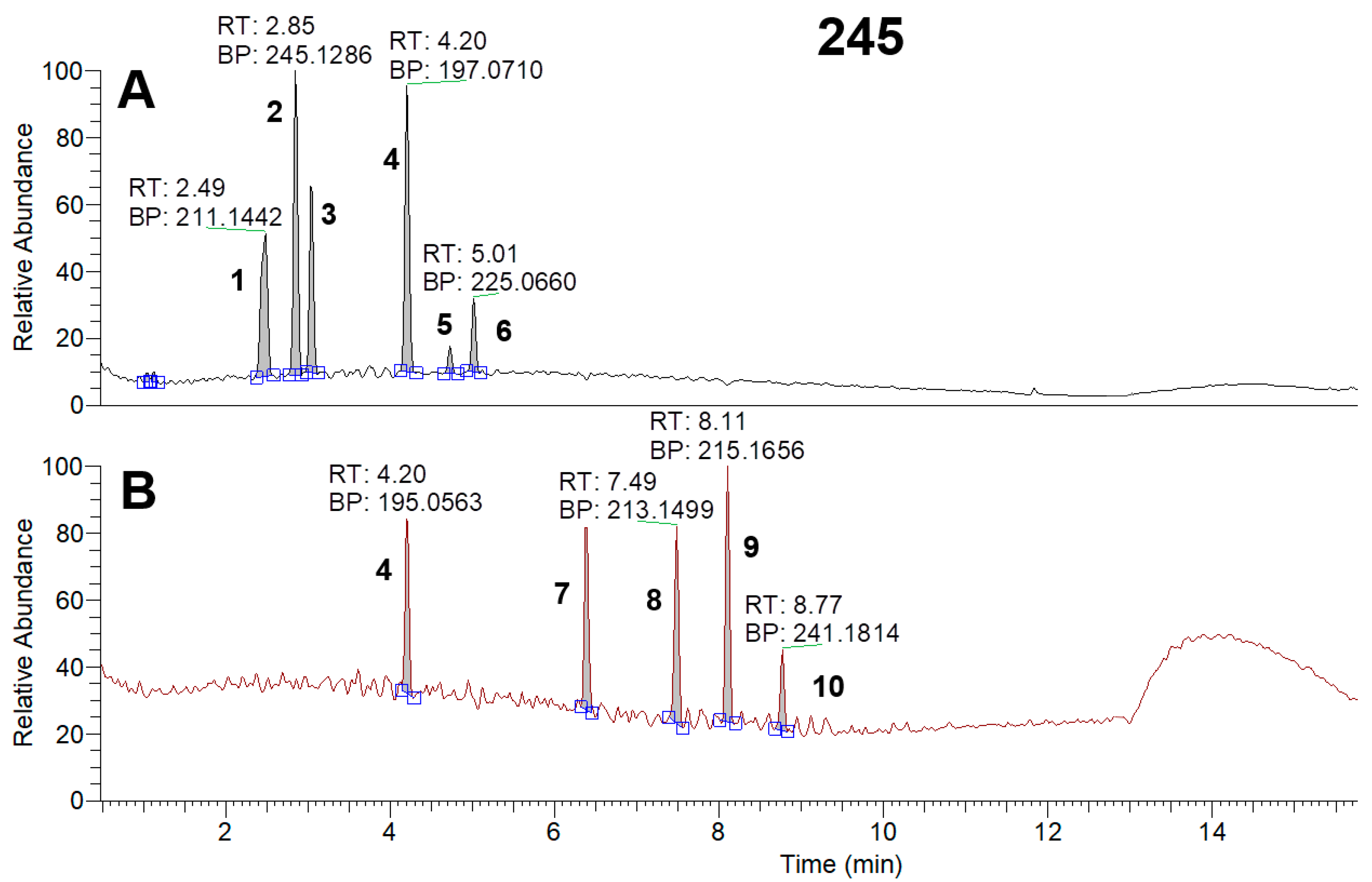
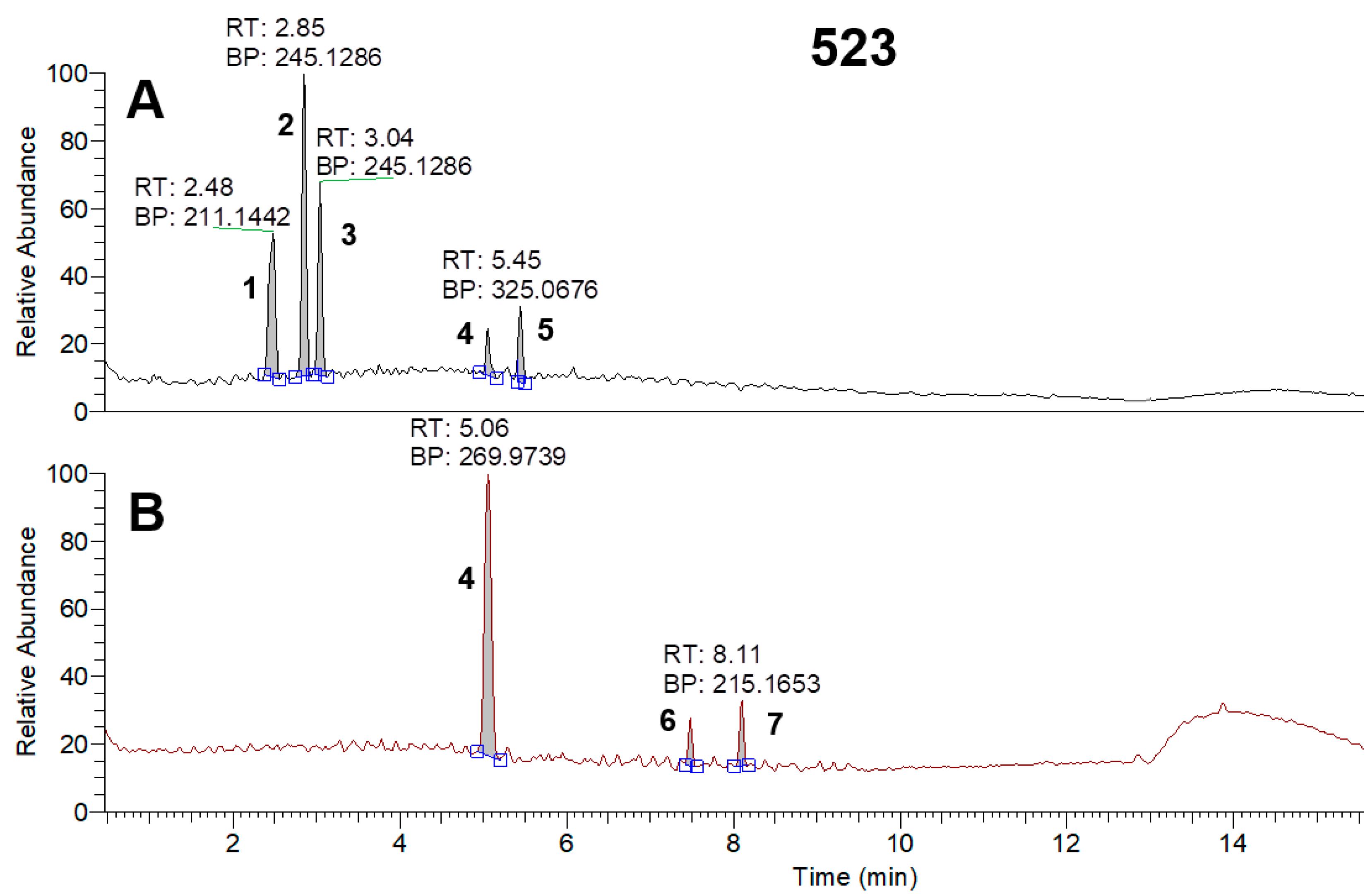
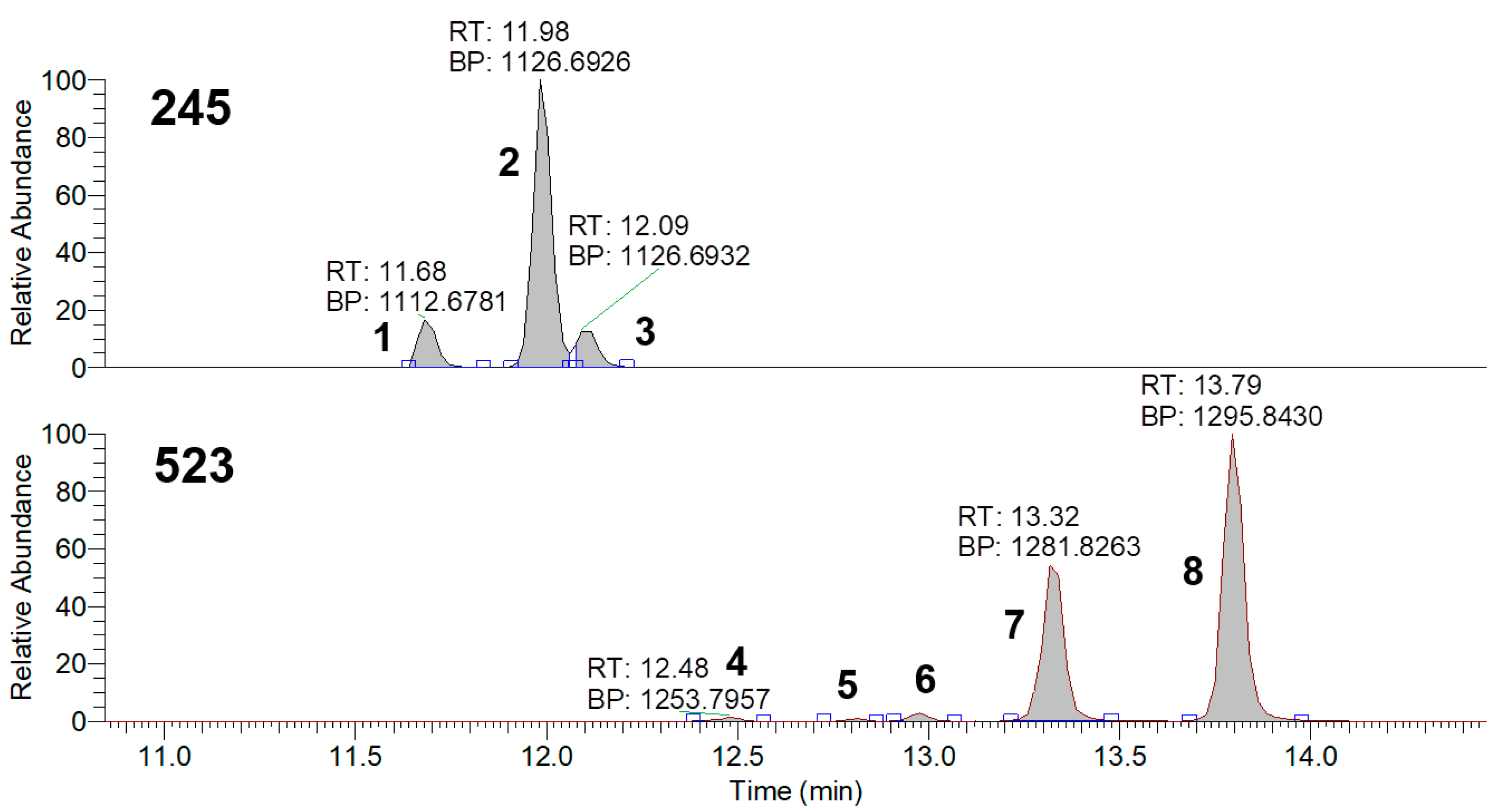
3.5. HPLC-MS Analysis of Metabolites of Strain Samples
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miftakhov, A.K.; Diabankana, R.G.; Frolov, M.A.; Yusupov, M.M.; Validov, S.Z.; Afordoanyi, D.M. Persistence as a Constituent of a Biocontrol Mechanism (Competition for Nutrients and Niches) in Pseudomonas putida PCL1760. Microorganisms 2023, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Singh, P.; Singh, R.; Zhou, Y.; Wang, J.; Jiang, Y.; Shen, N.; Wang, Y.; Yang, L.; Jiang, M. Unlocking the strength of plant growth promoting Pseudomonas in improving crop productivity in normal and challenging environments: A review. J. Plant Interact. 2022, 17, 220–238. [Google Scholar] [CrossRef]
- Anderson, A.J.; Kim, Y.C. Biopesticides produced by plant-probiotic Pseudomonas chlororaphis isolates. Crop Prot. 2018, 105, 62–69. [Google Scholar] [CrossRef]
- Platel, R.; Chaveriat, L.; Le Guenic, S.; Pipeleers, R.; Magnin-Robert, M.; Randoux, B.; Trapet, P.; Lequart, V.; Joly, N.; Halama, P.; et al. Importance of the C12 Carbon Chain in the Biological Activity of Rhamnolipids Conferring Protection in Wheat against Zymoseptoria tritici. Molecules 2021, 26, 40. [Google Scholar] [CrossRef] [PubMed]
- Chin-A-Woeng, T.F.C.; Bloemberg, G.V.; Lugtenberg, B.J.J. Phenazines and their role in biocontrol by Pseudomonas bacteria. New Phytol. 2003, 137, 503–523. [Google Scholar] [CrossRef]
- Wang, S.Y.; Shi, X.C.; Chen, X.A.; Laborda, P.P.; Zhao, Y.Y.; Liu, F.Q.; Laborda, P. Biocontrol ability of phenazine-producing strains for the management of fungal plant pathogens. A Rev. Biol. Control. 2021, 155, 104548. [Google Scholar] [CrossRef]
- Dimkić, I.; Janakiev, T.; Petrović, M.; Degrassi, G.; Fira, D. Plant-associated Bacillus and Pseudomonas antimicrobial activities in plant disease suppression via biological control mechanisms—A review. Physiol. Mol. Plant Pathol. 2022, 117, 101754. [Google Scholar] [CrossRef]
- Soesanto, L.; Pradiptha, N.C.; Mugiastuti, E. Raw Secondary Metabolites of Chitosan-enriched Pseudomonas fluorescens P60 to Control Corn Sheath Blight. Biosaintifika. J. Biol. Biol. Educ. 2021, 13, 113–120. [Google Scholar] [CrossRef]
- Biessy, A.; Filion, M. Phenazines in plant-beneficial Pseudomonas spp.: Biosynthesis, regulation, function and genomics. Environ. Microbiol. 2018, 20, 3905–3917. [Google Scholar] [CrossRef] [Green Version]
- Ony, F.E.; Esmacel, Q.; Onyeka, J.T.; Adeleka, R.; Jacquard, C.; Gross, H.; Barka, E.A.; Höfte, M. Pseudomonas lipopeptide—Mediated biocontrol: Chemotaxonomy and biological activity. Molecules 2022, 27, 372. [Google Scholar] [CrossRef] [PubMed]
- Roo, V.D.; Verleysen, Y.; Kovács, B.; Matthias, V.; Girard, L.; Höfte, M.; De Mot, R.; Madder, A.; Geudens, N.; Martins, J.C. An NMR fingerprint matching approach for the identification and structural re-evaluation of Pseudomonas lipopeptides. bioRxiv 2022, 47, 54–72. [Google Scholar]
- Certificate of the Database State Registration «Antagonistic Bacteria from the Bioresource Collection “State Collection of Entomoacariphages and Microorganisms” Federal State Budgetary Scientific Institution “Federal Research Center of Biological Plant Protection”». Available online: https://www.fips.ru/iiss/document.xhtml?faces-redirect=true&id=95ebd47a99325aab84df687df0ff936d (accessed on 13 July 2023).
- Syrmolot, O.V. Formation of the productivity of soybean plants depending on the use of biological products in Primorye. Far East. Agrar. Bull. 2018, 4, 115–119. [Google Scholar]
- Syrmolot, O.V.; Asaturova, A.M.; Khomyak, A.I. The effect of biological products on soybean productivity in Primorye. Agrochem. Bull. 2016, 6, 52–54. [Google Scholar]
- Baydelyuk, E.S. The effect of drugs based on bacterial strains Bacillus subtilis and Pseudomonas sp. when growing tomatoes in Primorsky Krai. Far East. Agrar. Bull. 2019, 4, 5–9. [Google Scholar]
- Asaturova, A.M.; Nadykta, V.D.; Dubyaga, V.M.; Tomashevich, N.S.; Zharnikova, M.D.; Zhevnova, N.A. Study of the effect of seed bacterization on the growth and development of winter wheat. Sci. J. KUBGAU 2013, 85. Available online: http://ej.kubagro.ru/2013/01/pdf/66.pdf (accessed on 28 July 2023).
- Asaturova, A.M.; Khomyak, A.I.; Tomashevich, N.S.; Pavlova, M.D.; Zhevnova, N.A.; Dubyaga, V.M.; Kozitsin, A.Y.; Sidorova, T.M.; Nadykta, V.D.; Ismailov, V.Y.A. Conditions for the cultivation of new Bacillus bacteria being micro bioproduct producers. J. Pur. Appl. Microbiol. 2015, 4, 2797–2804. [Google Scholar]
- Montealegre, J.R.; Reyes, R.; Perez, L.M.; Herrera, R.; Silva, P.; Besoain, X. Selection of bioantagonistic bacteria to be used in biological control of Rhizoctonia solani in tomato. Electron. J. Biotechnol. 2003, 6, 116–127. [Google Scholar] [CrossRef]
- Sidorova, T.M.; Asaturova, A.M.; Homyak, A.I.; Tomashevich, N.S. Isolation and characterization of antifungal metabolites of Bacillus subtilis strains BZR 336 g and BZR 517 using the modified bioauthography method. Agric. Boil. 2019, 1, 178–185. [Google Scholar]
- Hosh, A.; Cimpoiu, C. Evaluation of various biological activities of natural compounds by TLC/HPTLC. J. Liq. Chromatogr. Relat. Technol. 2020, 43, 305–318. [Google Scholar] [CrossRef]
- El-Shouny, W.A.; Ali, S.S.; Sun, J.; Samy, S.M.; Ali, A. Drug resistance profile and molecular characterization of extended spectrum beta-lactamase (esβl)-producing Pseudomonas aeruginosa isolated from burn wound infections. Essential oils and their potential for utilization. Microb. Pathog. 2018, 116, 301–312. [Google Scholar] [CrossRef]
- Thacharodi, A.; Priyadharshini, R.; Karthikeyan, G.; Jeganathan, C.; Reghu, A.P.; Pugazhendhi, A. Extraction, purification and characterization of phenazine from Pseudomonas aeruginosa isolate of wastewater sources: A panacea towards clinical pathogens. Appl. Nanosci. 2023, 13, 2365–2378. [Google Scholar] [CrossRef]
- Chauhan, V.; Mazumdar, S.; Pandey, A.; Kanwar, S.S. Pseudomonas Lipopeptide: An Excellent Biomedical Agent. MedComm. Biomater. Appl. 2023, 2, e27. [Google Scholar] [CrossRef]
- Serafim, B.; Bernardino, A.R.; Freitas, F.; Torres, C.A.V. Recent Developments in the Biological Activities, Bioproduction, and Applications of Pseudomonas spp. Phenazines. Molecules 2023, 28, 1368. [Google Scholar] [CrossRef] [PubMed]
- Muratova, A.A.; Buklyarevich, A.A.; Evdokimova, O.V.; Mandrik–Litvinkovich, M.N.; Valentovich, L.N.; Titok, M.A.; Kolomiets, E.I. Influence of the groEL genes encoding the synthesis of chaperone proteins on the production of 2,4-diacetylphloroglucinol in Pseudomonas brassicacearum S-1 bacteria. Bull. Fund Basic Res. 2013, 3, 195–206. [Google Scholar]
- Kong, S.; Shen, C.; Li, Y.; Meng, Q. Rhamnolipids Sustain Unchanged Surface Activities during Decomposition in Alkaline Solutions. ACS Omega 2021, 6, 15750–15755. [Google Scholar] [CrossRef]
- Romero, E.M.; Brenner, R.R. Fatty Acids Synthesized from Hexadecane by Pseudomonas aeruginosa. J. Bacteriol. 1966, 91, 183–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.M.; Wang, D.; Pierson, L.S.; Pierson, E.A. Effect of producing different phenozines on bacterial fitness and biological control in Pseudomonas chlororaphis 30–84. J. Plant Pathol. 2018, 34, 44–58. [Google Scholar] [CrossRef]
- Yu, J.M.; Wang, D.; Ries, T.R.; Pierson, L.S.; Pierson, E.A. An upstream sequence modulates phenazine production at the level of transcription and translation in the biological control strain Pseudomonas chlororaphis 30–84. PLoS ONE 2018, 13, e0193063. [Google Scholar] [CrossRef]
- Götze, S.; Stallforth, P. Structure, properties, and biological functions of nonribosomal lipopeptides from pseudomonads. Nat. Prod. Rep. 2020, 37, 29–54. [Google Scholar] [CrossRef]
- Guo, H.; Roman, D.; Beemelmanns, C. Tropolone natural products. Nat. Prod. Rep. 2019, 36, 137–1155. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yu, J.M.; Dorosky, R.J.; Pierson, L.S.; Pierson, E.A. The phenazine 2-hydroxy-phenazine-1-carboxylic acid promotes extracellular DNA release and has broad transcriptomic consequences in Pseudomonas chlororaphis 30–84. PLoS ONE 2016, 11, e0148003. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, T.R. Bacterially Mediated Water Stress Tolerance in Wheat Conferred by Phenazine-Producing Rhizobacteria. Master’s Thesis, Texas A&M University, College Station, TX, USA, 2017. [Google Scholar]
- Anderson, A.J.; Kim, Y.C. Insighs into plant-benefical traits of probiotic Pseudomonas chlororaphis isolates. J. Med. Microbiol. 2020, 69, e01569820. [Google Scholar] [CrossRef]
- Andrić, S.; Meyer, T.; Rigolet, A.; Prigent-Combaret, C.; Höfte, M.; Hoff, G.; De Mot, R.; McCann, A.; De Pauw, E.; Arias, A.A.; et al. Lipopeptide interplay mediates molecular interactions between soil Bacilli and Pseudomonads. Microbiol. Spectr. 2021, 9, e0203821. [Google Scholar] [CrossRef]
- Jang, J.Y.; Yang, S.Y.; Kim, Y.C.; Lee, C.W.; Park, M.S.; Kim, J.C.; Kim, I.S. Identification of Orfamide A as an Insecticidal Metabolite Produced by Pseudomonas protegens F6. J. Agric. Food Chem. 2013, 61, 6786–6791. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antagonistic Bacterial Strain | Antagonistic Activity against F. graminearum BZR F-4, % | Antagonistic Activity against F. oxysporum var. orthoceras BZR F-6, % | ||||||
|---|---|---|---|---|---|---|---|---|
| 5 Days | 10 Days | 5 Days | 10 Days | |||||
| Growth of Fungal Strain | % Inhibition | Growth of Fungal Strain | % Inhibition | Growth of Fungal Strain | % Inhibition | Growth of Fungal Strain | % Inhibition | |
| Negative control (without bacterial strains) | 50.0 ± 0.6 a | – | 75.0 ± 0 a | – | 46.7 ± 3.0 a | – | 75.0 ± 0 a | – |
| P. chlororaphis BZR 245-F | 38.7 ± 3.5 b | 22.7 | 30.3 ± 1.5 c | 59.6 | 33.7 ± 1.5 b | 27.9 | 37.3 ± 4.7 c | 50.2 |
| Pseudomonas sp. BZR 523-2 | 43.7 ± 1.2 b | 12.7 | 63.7 ± 2.9 b | 15.1 | 35.7 ± 1.2 b | 23.6 | 68.3 ± 0.6 b | 8.9 |
| Treatment | Plant Height, mm | Root Length, mm | Weight of Dry Biomass, g | |
|---|---|---|---|---|
| Shoots | Roots | |||
| Control | 23.3 ± 0.3 a | 12.9 ± 0.6 a | 0.0161 ± 0.002 a | 0.0083 ± 0.0005 a |
| P. chlororaphis BZR 245-F | 24.1 ± 0.2 b | 11.9 ± 0.4 a | 0.0166 ± 0.002 a | 0.0089 ± 0.0002 a |
| Pseudomonas sp. BZR 523-2 | 25.7 ± 0.5 c | 15.3 ± 0.5 b | 0.0197 ± 0.003 b | 0.0094 ± 0.0001 b |
| Compound | Structure and Gross Formula | Content, mg/g | |
|---|---|---|---|
| P. chlororaphis BZR 245-F | Pseudomonas sp. BZR 523-2 | ||
| Phenazine |  C12H8N2 | 0.07 | 2.77 |
| Pyocyanin |  C13H10N2O | 0.16 | 0.14 |
| Phenazine-1-carboxylic acid |  C13H8N2O2 | 3.22 | – |
| 1-Phenazinol |  C12H8N2O | 11.3 | – |
| 2-Hydroxyphenazine-1-carboxylic acid |  C13H8N2O3 | 2.26 | – |
| Phenazine-1-carboxamide |  C13H9N3O | 0.01 | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sidorova, T.M.; Tomashevich, N.S.; Allahverdyan, V.V.; Tupertsev, B.S.; Kostyukevich, Y.I.; Asaturova, A.M. New Pseudomonas Bacterial Strains: Biological Activity and Characteristic Properties of Metabolites. Microorganisms 2023, 11, 1943. https://doi.org/10.3390/microorganisms11081943
Sidorova TM, Tomashevich NS, Allahverdyan VV, Tupertsev BS, Kostyukevich YI, Asaturova AM. New Pseudomonas Bacterial Strains: Biological Activity and Characteristic Properties of Metabolites. Microorganisms. 2023; 11(8):1943. https://doi.org/10.3390/microorganisms11081943
Chicago/Turabian StyleSidorova, Tatiana M., Natalia S. Tomashevich, Valeria V. Allahverdyan, Boris S. Tupertsev, Yuri I. Kostyukevich, and Anzhela M. Asaturova. 2023. "New Pseudomonas Bacterial Strains: Biological Activity and Characteristic Properties of Metabolites" Microorganisms 11, no. 8: 1943. https://doi.org/10.3390/microorganisms11081943