
Investigation of the Critical Biomass of Acclimated Microbial Communities to High Ammonia Concentrations for a Successful Bioaugmentation of Biogas Anaerobic Reactors with Ammonia Inhibition
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Inoculum and Feedstock
2.2. Bioaugmentation Culture
2.3. Experimental Setup
2.4. Analytical Methods
2.5. Microbial Community and Bioinformatics Analysis
2.6. Statistical Analyses
2.7. Calculations
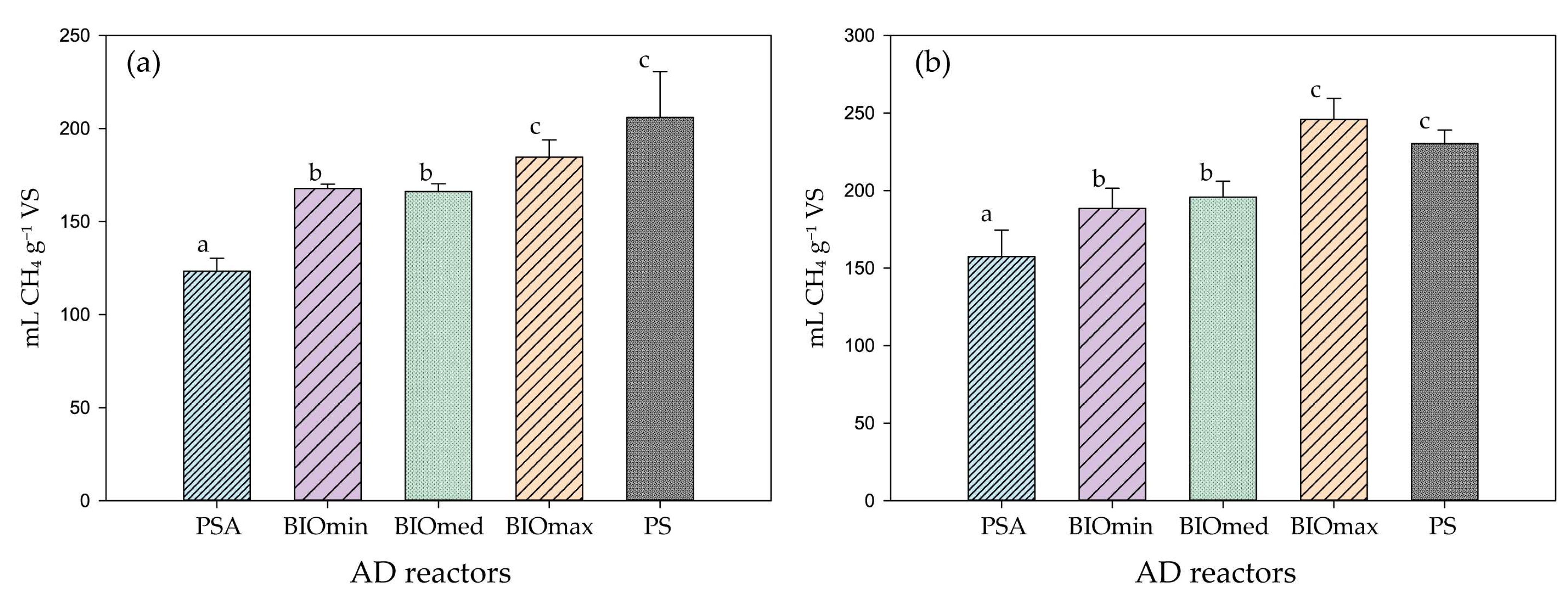
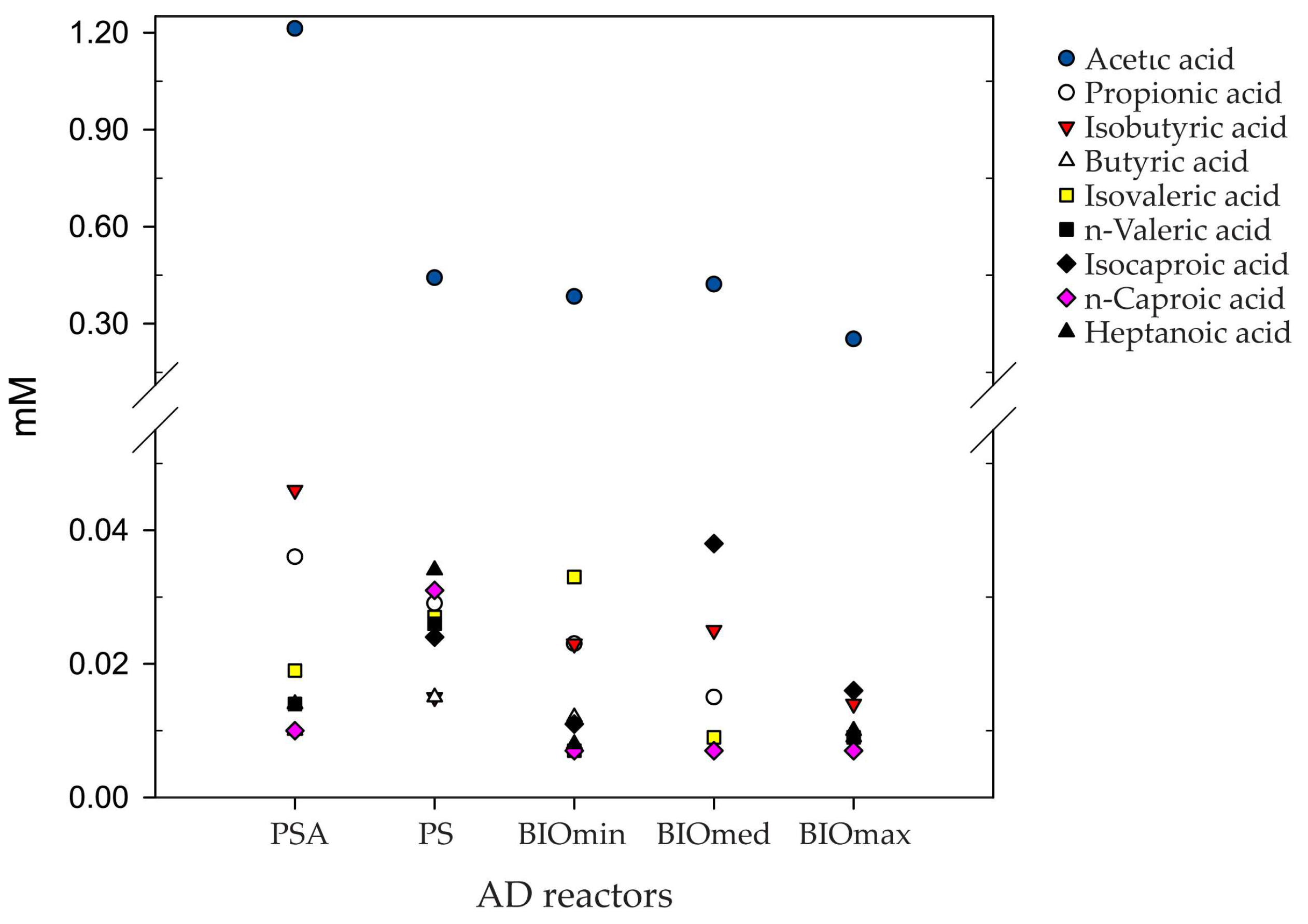
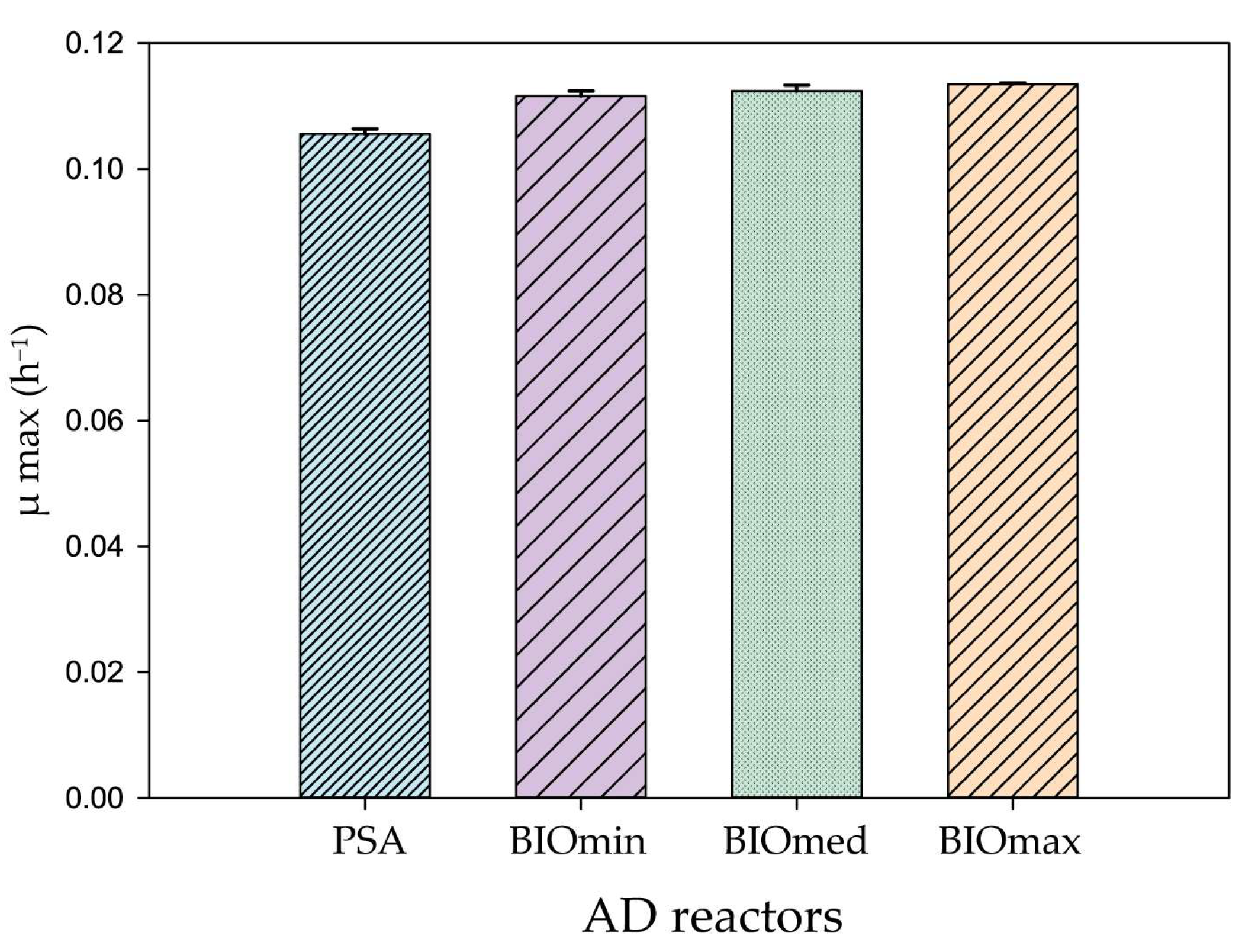
3. Results & Discussion
3.1. Anaerobic Digestion
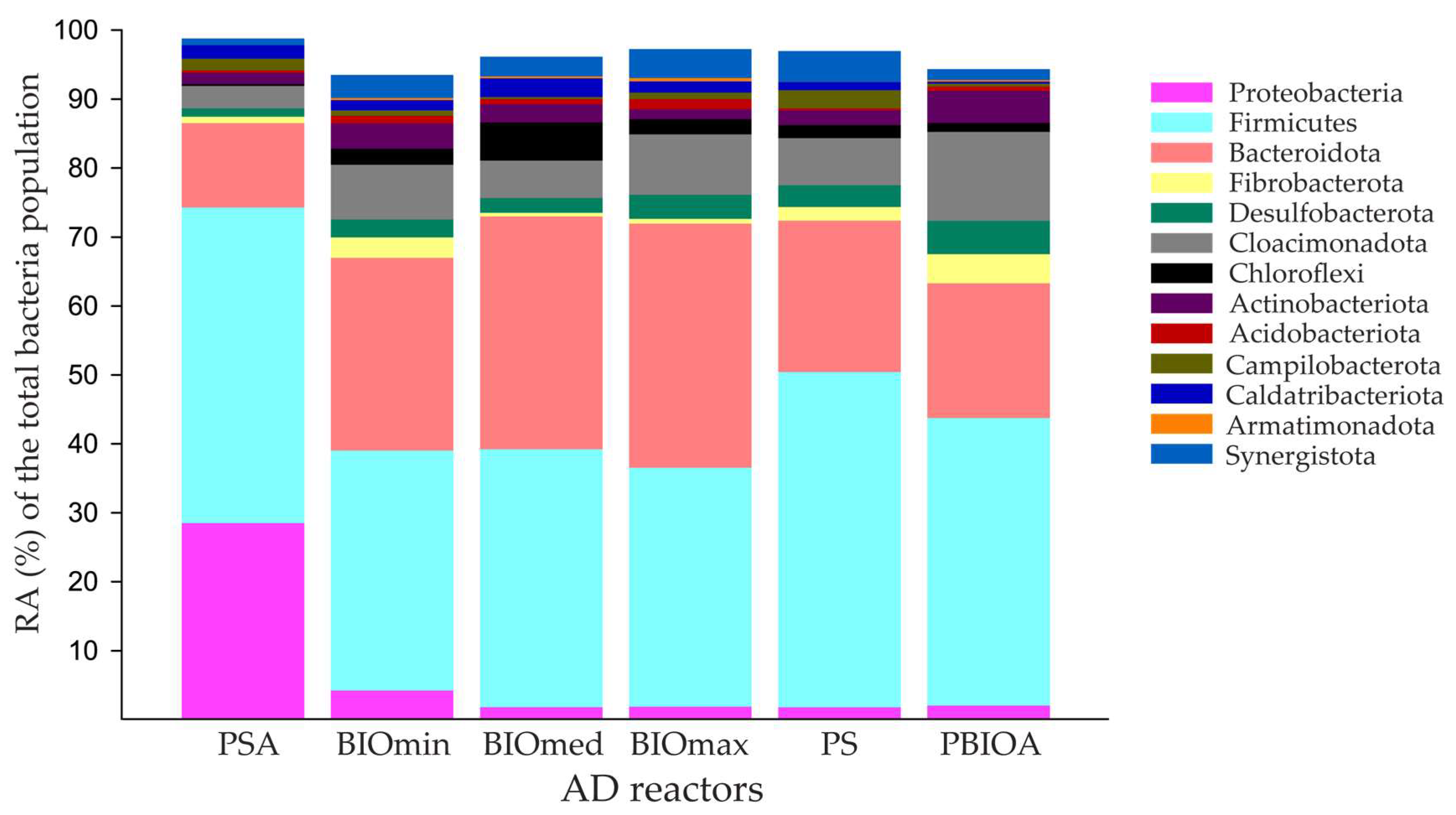
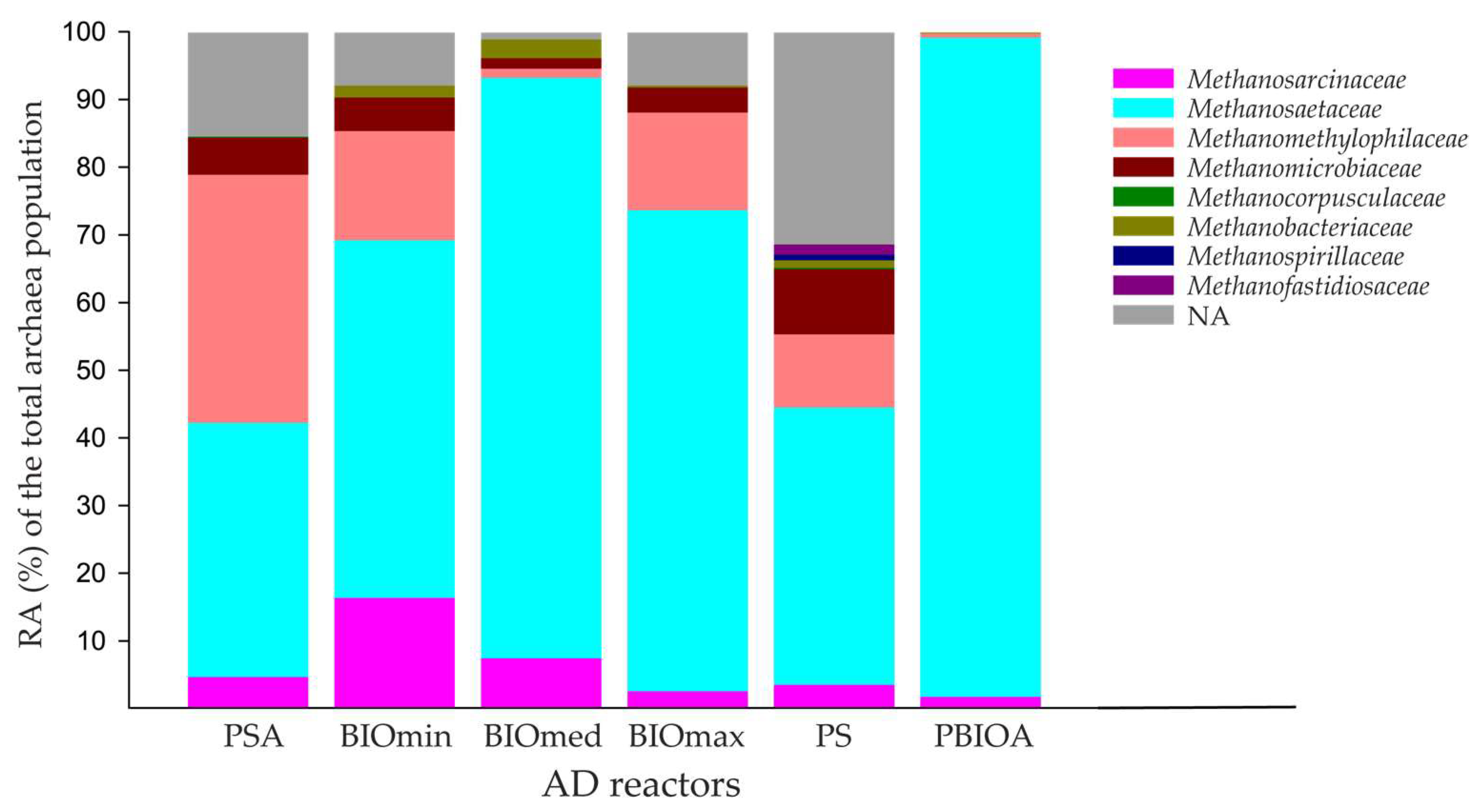
3.2. Effect of Ammonia and Bioaugmentation on the Microbial Community of Anaerobic Reactors
3.2.1. Bacterial Community Composition
3.2.2. Archaeal Community Composition
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zupančič, M.; Možic, V.; Može, M.; Cimerman, F.; Golobič, I. Current Status and Review of Waste-to-Biogas Conversion for Selected European Countries and Worldwide. Sustainability 2022, 14, 1823. [Google Scholar] [CrossRef]
- Huang, W.; Zhao, Z.; Yuan, T.; Yu, Y.; Huang, W.; Lei, Z.; Zhang, Z. Enhanced dry anaerobic digestion of swine excreta after organic nitrogen being recovered as soluble proteins and amino acids using hydrothermal technology. Biomass Bioenergy 2018, 108, 120–125. [Google Scholar] [CrossRef]
- Kotsopoulos, T.A.; Karamanlis, X.; Dotas, D.; Martzopoulos, G.G. The impact of different natural zeolite concentrations on the methane production in thermophilic anaerobic digestion of pig waste. Biosyst. Eng. 2008, 99, 105–111. [Google Scholar] [CrossRef]
- Wang, H.; Fotidis, I.A.; Angelidaki, I. Ammonia effect on hydrogenotrophic methanogens and syntrophic acetate-oxidizing bacteria. FEMS Microbiol. Ecol. 2015, 91, fiv130. [Google Scholar] [CrossRef] [Green Version]
- Massé, D.I.; Rajagopal, R.; Singh, G. Technical and operational feasibility of psychrophilic anaerobic digestion biotechnology for processing ammonia-rich waste. Appl. Energy 2014, 120, 49–55. [Google Scholar] [CrossRef]
- Nzila, A. Mini review: Update on bioaugmentation in anaerobic processes for biogas production. Anaerobe 2017, 46, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-Y.; Inoue, D.; Ike, M. Mitigating ammonia-inhibition in anaerobic digestion by bioaugmentation: A review. J. Water Process Eng. 2023, 52, 103506. [Google Scholar] [CrossRef]
- Venkiteshwaran, K.; Bocher, B.; Maki, J.; Zitomer, D. Relating Anaerobic Digestion Microbial Community and Process Function: Supplementary Issue: Water Microbiology. Microbiol. Insights 2015, 8s2, MBI.S33593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Sun, H.; Zhao, Q.; Kurbonova, M.; Zhang, R.; Liu, G.; Wang, W. Long-term evaluation of bioaugmentation to alleviate ammonia inhibition during anaerobic digestion: Process monitoring, microbial community response, and methanogenic pathway modeling. Chem. Eng. J. 2020, 399, 125765. [Google Scholar] [CrossRef]
- Tian, H.; Mancini, E.; Treu, L.; Angelidaki, I.; Fotidis, I.A. Bioaugmentation strategy for overcoming ammonia inhibition during biomethanation of a protein-rich substrate. Chemosphere 2019, 231, 415–422. [Google Scholar] [CrossRef]
- Fotidis, I.A.; Treu, L.; Angelidaki, I. Enriched ammonia-tolerant methanogenic cultures as bioaugmentation inocula in continuous biomethanation processes. J. Clean. Prod. 2017, 166, 1305–1313. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Wang, W.; Liu, C.; Zhang, R.; Liu, G. Mitigation of ammonia inhibition through bioaugmentation with different microorganisms during anaerobic digestion: Selection of strains and reactor performance evaluation. Water Res. 2019, 155, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Christou, M.L.; Vasileiadis, S.; Karpouzas, D.G.; Angelidaki, I.; Kotsopoulos, T.A. Effects of organic loading rate and hydraulic retention time on bioaugmentation performance to tackle ammonia inhibition in anaerobic digestion. Bioresour. Technol. 2021, 334, 125246. [Google Scholar] [CrossRef] [PubMed]
- Kalamaras, S.D.; Vasileiadis, S.; Karas, P.; Angelidaki, I.; Kotsopoulos, T.A. Microbial adaptation to high ammonia concentrations during anaerobic digestion of manure-based feedstock: Biomethanation and 16S rRNA gene sequencing. J. Chem. Technol. Biotechnol. 2020, 95, 1970–1979. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Sun, Y.; Wu, S.; Kong, X.; Yuan, Z.; Dong, R. The performance efficiency of bioaugmentation to prevent anaerobic digestion failure from ammonia and propionate inhibition. Bioresour. Technol. 2017, 231, 94–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.-T.; Rao, L.; Wang, L.; Gou, M.; Sun, Z.-Y.; Xia, Z.-Y.; Song, W.-F.; Tang, Y.-Q. Bioaugmentation with syntrophic volatile fatty acids-oxidizing consortia to alleviate the ammonia inhibition in continuously anaerobic digestion of municipal sludge. Chemosphere 2022, 288, 132389. [Google Scholar] [CrossRef]
- Yang, Z.; Sun, H.; Zhou, L.; Arhin, S.G.; Papadakis, V.G.; Goula, M.A.; Liu, G.; Zhang, Y.; Wang, W. Bioaugmentation with well-constructed consortia can effectively alleviate ammonia inhibition of practical manure anaerobic digestion. Water Res. 2022, 215, 118244. [Google Scholar] [CrossRef]
- Aziz, C.E.; Wymore, R.A.; Steffan, R.J. Bioaugmentation Considerations. In Bioaugmentation for Groundwater Remediation; Stroo, H.F., Leeson, A., Ward, C.H., Eds.; Springer: New York, NY, USA, 2013; pp. 141–169. [Google Scholar]
- Fotidis, I.A.; Wang, H.; Fiedel, N.R.; Luo, G.; Karakashev, D.B.; Angelidaki, I. Bioaugmentation as a Solution To Increase Methane Production from an Ammonia-Rich Substrate. Environ. Sci. Technol. 2014, 48, 7669–7676. [Google Scholar] [CrossRef]
- Fotidis, I.A.; Karakashev, D.; Angelidaki, I. Bioaugmentation with an acetate-oxidising consortium as a tool to tackle ammonia inhibition of anaerobic digestion. Bioresour. Technol. 2013, 146, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.; Yan, M.; Treu, L.; Angelidaki, I.; Fotidis, I.A. Hydrogenotrophic methanogens are the key for a successful bioaugmentation to alleviate ammonia inhibition in thermophilic anaerobic digesters. Bioresour. Technol. 2019, 293, 122070. [Google Scholar] [CrossRef]
- Yan, M.; Treu, L.; Campanaro, S.; Tian, H.; Zhu, X.; Khoshnevisan, B.; Tsapekos, P.; Angelidaki, I.; Fotidis, I.A. Effect of ammonia on anaerobic digestion of municipal solid waste: Inhibitory performance, bioaugmentation and microbiome functional reconstruction. Chem. Eng. J. 2020, 401, 126159. [Google Scholar] [CrossRef]
- Bridgewater, L.; American Public Health Association; American Water Works Association; Water Environment Federation. Standard Methods for the Examination of Water and Wastewater, 21st ed.; APHA-AWWA-WEF: Washington, DC, USA, 2005. [Google Scholar]
- Kalamaras, S.D.; Vitoulis, G.; Christou, M.L.; Sfetsas, T.; Tziakas, S.; Fragos, V.; Samaras, P.; Kotsopoulos, T.A. The Effect of Ammonia Toxicity on Methane Production of a Full-Scale Biogas Plant—An Estimation Method. Energies 2021, 14, 5031. [Google Scholar] [CrossRef]
- Katsoula, A.; Vasileiadis, S.; Sapountzi, M.; Karpouzas, D.G. The response of soil and phyllosphere microbial communities to repeated application of the fungicide iprodione: Accelerated biodegradation or toxicity? FEMS Microbiol. Ecol. 2020, 96, fiaa056. [Google Scholar] [CrossRef]
- Berry, D.; Mahfoudh, K.B.; Wagner, M.; Loy, A. Barcoded Primers Used in Multiplex Amplicon Pyrosequencing Bias Amplification. Appl. Environ. Microbiol. 2011, 77, 7846–7849. [Google Scholar] [CrossRef] [Green Version]
- Apprill, A.; McNally, S.; Parsons, R.; Weber, L. Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Alma, E.; Needham, D.; Fuhrman, J. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef]
- Christou, M.L.; Vasileiadis, S.; Kalamaras, S.D.; Karpouzas, D.G.; Angelidaki, I.; Kotsopoulos, T.A. Ammonia-induced inhibition of manure-based continuous biomethanation process under different organic loading rates and associated microbial community dynamics. Bioresour. Technol. 2021, 320, 124323. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing, Reference Index Version 3.3.3; R Foundation for Statistical Computing: Indianapolis, IN, USA, 2017. [Google Scholar]
- Gray, N.D.; Sherry, A.; Larter, S.R.; Erdmann, M.; Leyris, J.; Liengen, T.; Beeder, J.; Head, I.M. Biogenic methane production in formation waters from a large gas field in the North Sea. Extremophiles 2009, 13, 511–519. [Google Scholar] [CrossRef]
- Edward, M.; Edwards, S.; Egwu, U.; Sallis, P. Bio-methane potential test (BMP) using inert gas sampling bags with macroalgae feedstock. Biomass Bioenergy 2015, 83, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Hansen, T.L.; Schmidt, J.E.; Angelidaki, I.; Marca, E.; Jansen, J.l.C.; Mosbæk, H.; Christensen, T.H. Method for determination of methane potentials of solid organic waste. Waste Manag. 2004, 24, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Fotidis, I.A.; Mancini, E.; Angelidaki, I. Different cultivation methods to acclimatise ammonia-tolerant methanogenic consortia. Bioresour. Technol. 2017, 232, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franke-Whittle, I.H.; Walter, A.; Ebner, C.; Insam, H. Investigation into the effect of high concentrations of volatile fatty acids in anaerobic digestion on methanogenic communities. Waste Manag. 2014, 34, 2080–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, T.J.; Allen, M.A.; Berengut, J.F.; Cavicchioli, R. Shedding Light on Microbial “Dark Matter”: Insights Into Novel Cloacimonadota and Omnitrophota From an Antarctic Lake. Front. Microbiol. 2021, 12, 741077. [Google Scholar] [CrossRef]
- Johnson, L.A.; Hug, L.A. Cloacimonadota metabolisms include adaptations in engineered environments that are reflected in the evolutionary history of the phylum. Environ. Microbiol. Rep. 2022, 14, 520–529. [Google Scholar] [CrossRef]
- Kara Murdoch, F.; Murdoch, R.W.; Gürakan, G.C.; Sanin, F.D. Change of microbial community composition in anaerobic digesters during the degradation of nonylphenol diethoxylate. Int. Biodeterior. Biodegrad. 2018, 135, 1–8. [Google Scholar] [CrossRef]
- Li, C.; Hao, L.; Lü, F.; Duan, H.; Zhang, H.; He, P. Syntrophic Acetate-Oxidizing Microbial Consortia Enriched from Full-Scale Mesophilic Food Waste Anaerobic Digesters Showing High Biodiversity and Functional Redundancy. mSystems 2022, 7, e00339-22. [Google Scholar] [CrossRef]
- Zhang, C.; Yuan, Q.; Lu, Y. Inhibitory effects of ammonia on syntrophic propionate oxidation in anaerobic digester sludge. Water Res. 2018, 146, 275–287. [Google Scholar] [CrossRef]
- Leite, A.F.; Janke, L.; Lv, Z.; Harms, H.; Richnow, H.-H.; Nikolausz, M. Improved Monitoring of Semi-Continuous Anaerobic Digestion of Sugarcane Waste: Effects of Increasing Organic Loading Rate on Methanogenic Community Dynamics. Int. J. Mol. Sci. 2015, 16, 23210–23226. [Google Scholar] [CrossRef] [Green Version]
- Buan, N.; Kulkarni, G.; Metcalf, W. Chapter two—Genetic Methods for Methanosarcina Species. In Methods in Enzymology, Rosenzweig, A.C., Ragsdale, S.W., Eds.; Academic Press: Cambridge, MA, USA, 2011; Volume 494, pp. 23–42. [Google Scholar]
- Demirel, B.; Scherer, P. The roles of acetotrophic and hydrogenotrophic methanogens during anaerobic conversion of biomass to methane: A review. Rev. Environ. Sci. Bio/Technol. 2008, 7, 173–190. [Google Scholar] [CrossRef]
- Goberna, M.; Gadermaier, M.; García, C.; Wett, B.; Insam, H. Adaptation of Methanogenic Communities to the Cofermentation of Cattle Excreta and Olive Mill Wastes at 37 °C and 55 °C. Appl. Environ. Microbiol. 2010, 76, 6564–6571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fazi, S.; Amalfitano, S.; Venturi, S.; Pacini, N.; Vazquez, E.; Olaka, L.A.; Tassi, F.; Crognale, S.; Herzsprung, P.; Lechtenfeld, O.J.; et al. High concentrations of dissolved biogenic methane associated with cyanobacterial blooms in East African lake surface water. Commun. Biol. 2021, 4, 845. [Google Scholar] [CrossRef] [PubMed]
- Quispe-Cardenas, E.; Rogers, S. Microbial adaptation and response to high ammonia concentrations and precipitates during anaerobic digestion under psychrophilic and mesophilic conditions. Water Res. 2021, 204, 117596. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Fotidis, I.A.; Mancini, E.; Treu, L.; Mahdy, A.; Ballesteros, M.; González-Fernández, C.; Angelidaki, I. Acclimation to extremely high ammonia levels in continuous biomethanation process and the associated microbial community dynamics. Bioresour. Technol. 2018, 247, 616–623. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter (Unit) | Inoculum for AD | Inoculum for Bioaugmentation Culture | Bioaugmented Culture | Cattle Manure |
|---|---|---|---|---|
| Total Solids–TS (g L−1) | 67.10 ± 0.35 | 65.2 ± 0.34 | 37.28 ± 0.84 | 66.6 ± 0.42 |
| Volatile Solids–VS (g L−1) | 52.52 ± 0.42 | 50.48 ± 0.45 | 30.85 ± 0.93 | 56.05 ± 0.54 |
| Total Kjeldahl Nitrogen–TKN (g L−1) | 4.06 ± 0.34 | 4.00 ± 0.28 | 5.45 ± 0.69 | 3.60 ± 0.58 |
| Total Ammonia Nitrogen–TAN (g L−1) | 2.03 ± 0.18 | 2.53 ± 0.25 | 5.20 ± 0.19 | 1.57 ± 0.12 |
| pH | 7.8 ± 0.05 | 7.7 ± 0.05 | 7.8 ± 0.03 | 7.60 ± 0.04 |
| Total Volatile Fatty Acids–VFAs (g L−1) | 0.10 ± 0.003 | 0.10 ± 0.001 | 0.12 ± 0.001 | 11.61 ± 0.15 |
| AD Reactors | Inoculum for AD (g) | Cattle Manure (g) | Ammonia (NH4Cl 50 g L−1) (g) | Bioaugmented Culture (g) | Water (g) | Total (g) |
|---|---|---|---|---|---|---|
| PSA | 112 | 18 | 8.90 | - | 21.10 | 160 |
| PS | 112 | 18 | - | - | 30.00 | 160 |
| IN | 112 | - | - | - | 48.00 | 160 |
| PBIOA | - | - | 13.50 | 10.09 | 136.41 | 160 |
| BIOmin | 112 | 18 | 8.50 | 6.96 | 14.54 | 160 |
| BIOmed | 112 | 18 | 8.10 | 13.90 | 8.00 | 160 |
| BIOmax | 112 | 18 | 7.70 | 20.85 | 1.45 | 160 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalamaras, S.D.; Christou, M.L.; Tzenos, C.A.; Vasileiadis, S.; Karpouzas, D.G.; Kotsopoulos, T.A. Investigation of the Critical Biomass of Acclimated Microbial Communities to High Ammonia Concentrations for a Successful Bioaugmentation of Biogas Anaerobic Reactors with Ammonia Inhibition. Microorganisms 2023, 11, 1710. https://doi.org/10.3390/microorganisms11071710
Kalamaras SD, Christou ML, Tzenos CA, Vasileiadis S, Karpouzas DG, Kotsopoulos TA. Investigation of the Critical Biomass of Acclimated Microbial Communities to High Ammonia Concentrations for a Successful Bioaugmentation of Biogas Anaerobic Reactors with Ammonia Inhibition. Microorganisms. 2023; 11(7):1710. https://doi.org/10.3390/microorganisms11071710
Chicago/Turabian StyleKalamaras, Sotirios D., Maria Lida Christou, Christos A. Tzenos, Sotirios Vasileiadis, Dimitrios G. Karpouzas, and Thomas A. Kotsopoulos. 2023. "Investigation of the Critical Biomass of Acclimated Microbial Communities to High Ammonia Concentrations for a Successful Bioaugmentation of Biogas Anaerobic Reactors with Ammonia Inhibition" Microorganisms 11, no. 7: 1710. https://doi.org/10.3390/microorganisms11071710