
Insights into the Biology of Leucocytozoon Species (Haemosporida, Leucocytozoidae): Why Is There Slow Research Progress on Agents of Leucocytozoonosis?

Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. Brief Outline of Leucocytozoon Life Cycle
3.2. Remarks on Some Unique Haemosporidian Life Cycle Features
3.3. Diversity of Leucocytozoon Species
3.3.1. The Classification Problems
3.3.2. Leucocytozoon Species Taxonomy
3.3.3. The Problem of Synonymous Species Names
3.3.4. Morphological Characters of Blood Stages
3.4. Host Cells of Leucocytozoon Parasites
3.5. The Obstacles in Research on Exo-Erythrocytic Development
3.6. Challenges of the Molecular Characterization of Leucocytozoon Parasites
3.7. The Obstacles in Leucocytozoon Parasite Vector Research
3.8. Puzzles of the Geographical Distribution
3.9. Pathogenicity and Related Health and the Host-Parasite Association Issues
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garnham, P.C.C. Malaria Parasites and Other Haemosporidia; Blackwell: Oxford, UK, 1966; p. 1114. [Google Scholar]
- Desser, S.S.; Bennett, G.F. The genera Leucocytozoon, Haemoproteus and Hepatocystis. In Parasitic Protozoa, 2nd ed.; Kreier, J.P., Baker, J.R., Eds.; Academic Press: San Diego, CA, USA, 1993; Volume 4, pp. 273–307. [Google Scholar]
- Forrester, D.J.; Greiner, E.C. Leucocytozoonosis. In Parasitic Diseases of Wild Birds; Atkinson, C.T., Thomas, N.J., Hunter, D.B., Eds.; Wiley-Blackwell: Oxford, UK, 2008; pp. 54–107. [Google Scholar]
- Valkiūnas, G. Avian Malaria Parasites and Other Haemosporidia; CRC: Boca Raton, FL, USA, 2005; pp. 1–932. [Google Scholar] [CrossRef]
- Santiago-Alarcon, D.; Marzal, A. Avian Malaria and Related Parasites in the Tropics: Ecology, Evolution and Systematics; Springer Nature Switzerland AG: Cham, Switzerland, 2020; p. 575. [Google Scholar] [CrossRef]
- Perkins, S.L. Malaria’s many mates: Past, present, and future of the systematics of the order Haemosporida. J. Parasitol. 2014, 10, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Galen, S.C.; Borner, J.; Martinsen, E.S.; Schaer, J.; Austin, C.C.; West, C.J.; Perkins, S.L. The polyphyly of Plasmodium: Comprehensive phylogenetic analyses of the malaria parasites (order Haemosporida) reveal widespread taxonomic conflict. R. Soc. Open Sci. 2018, 5, 171780. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, M.A.; Matta, N.E.; Valkiūnas, G.; Parker, P.G.; Mello, B.; Stanley, C.E., Jr.; Lentino, M.; Garcia-Amado, M.A.; Cranfield, M.; Pond, S.L.K.; et al. Mode and rate of evolution of Haemosporidian mitochondrial genomes: Timing the radiation of avian parasites. Mol. Biol. Evol. 2018, 35, 383–403. [Google Scholar] [CrossRef]
- Lotta, I.A.; Valkiūnas, G.; Pacheco, M.A.; Escalante, A.A.; Hernández, S.R.; Matta, N.E. Disentangling Leucocytozoon parasite diversity in the neotropics: Descriptions of two new species and shortcomings of molecular diagnostics for leucocytozoids. Int. J. Parasitol. Parasites Wildl. 2019, 9, 159–173. [Google Scholar] [CrossRef]
- Svobodová, M.; Čepička, I.; Zídková, L.; Kassahun, A.; Votýpka, J.; Peške, L.; Hrazdilová, K.; Brzoňová, J.; Voříšek, P.; Weidinger, K. Blood parasites (Trypanosoma, Leucocytozoon, Haemoproteus) in the Eurasian sparrowhawk (Accipiter nisus): Diversity, incidence and persistence of infection at the individual level. Parasites Vectors 2023, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, M.A.; Junge, R.E.; Menon, A.; McRoberts, J.; Valkiūnas, G.; Escalante, A.A. The evolution of primate malaria parasites: A study on the origin and diversification of Plasmodium in lemurs. Mol. Phylogenet. Evol. 2022, 174, 107551. [Google Scholar] [CrossRef]
- Venugopal, K.; Hentzschel, F.; Valkiūnas, G.; Marti, M. Plasmodium asexual growth and sexual development in the haematopoietic niche of the host. Nat. Rev. Microbiol. 2020, 18, 177–189. [Google Scholar] [CrossRef]
- Bensch, S.; Canbäck, B.; DeBarry, J.D.; Johansson, T.; Hellgren, O.; Kissinger, J.C.; Palinauskas, V.; Videvall, E.; Valkiūnas, G. The Genome of Haemoproteus tartakovskyi and Its Relationship to Human Malaria Parasites. Genome Biol. Evol. 2016, 8, 1361–1373. [Google Scholar] [CrossRef]
- Böhme, U.; Otto, T.D.; Cotton, J.A.; Steinbiss, S.; Sanders, M.; Oyola, S.O.; Nicot, A.; Gandon, S.; Patra, K.P.; Herd, C.; et al. Complete avian malaria parasite genomes reveal features associated with lineage-specific evolution in birds and mammals. Genome Res. 2018, 28, 547–560. [Google Scholar] [CrossRef]
- Videvall, E. Genomic Advances in Avian Malaria Research. Trends Parasitol. 2019, 35, 254–266. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A. Keys to the avian malaria parasites. Malar. J. 2018, 17, 212. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A. Keys to the avian Haemoproteus parasites (Haemosporida, Haemoproteidae). Malar. J. 2022, 21, 269. [Google Scholar] [CrossRef] [PubMed]
- Matta, N.E.; Lotta, I.A.; Valkiūnas, G.; González, A.D.; Pacheco, M.A.; Escalante, A.A.; Moncada, L.I.; Rodríguez-Fandiño, O.A. Description of Leucocytozoon quynzae sp. nov. (Haemosporida, Leucocytozoidae) from hummingbirds, with remarks on distribution and possible vectors of leucocytozoids in South America. Parasitol. Res. 2014, 113, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Bensch, S.; Hellgren, O.; Pérez-Tris, J. MalAvi: A public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol. Ecol. Resour. 2009, 9, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Walther, E.; Valkiūnas, G.; Wommack, E.A.; Bowie, R.C.; Iezhova, T.A.; Sehgal, R.N. Description and molecular characterization of a new Leucocytozoon parasite (Haemosporida: Leucocytozoidae), Leucocytozoon californicus sp. nov., found in American kestrels (Falco sparverius sparverius). Parasitol. Res. 2016, 115, 1853–1862. [Google Scholar] [CrossRef] [PubMed]
- Groff, T.C.; Lorenz, T.J.; Iezhova, T.A.; Valkiūnas, G.; Sehgal, R.N.M. Description and molecular characterization of novel Leucocytozoon parasite (Apicomplexa: Haemosporida: Leucocytozoidae), Leucocytozoon polynuclearis n. sp. found in North American woodpeckers. Syst. Parasitol. 2022, 99, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, R.S.; Sapp, S.G.H.; Potters, I.; Mathison, B.A.; Frean, J.; Mewara, A.; Sheorey, H.; Tamarozzi, F.; Couturier, M.R.; Chiodini, P.; et al. Where Have All the Diagnostic Morphological Parasitologists Gone? J. Clin. Microbiol. 2022, 60, e0098622. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, C.T.; Thomas, N.J.; Hunter, D.B. Parasitic Diseases of Wild Birds; Wiley-Blackwell: Oxford, UK, 2008; 595p. [Google Scholar]
- Greiner, E.C.; Bennett, G.F.; White, E.M.; Coombs, R.F. Distribution of the avian hematozoa of north America. Can. J. Zool. 1975, 53, 1762–1787. [Google Scholar] [CrossRef]
- McClure, H.E.; Poonswad, P.; Greiner, E.C.; Laird, M. Haematozoa in the Birds of Eastern and Southern Asia; Memorial University of Newfoundland: St. John’s, NL, Canada, 1978; 296p. [Google Scholar]
- Nandi, N.C. Index-catalogue of avian haematozoa from India. Rec. Zool. Surv. India Occ. Paper 1984, 48, 1–63. [Google Scholar]
- Huchzermeyer, F.W. A host-parasite list of the haematozoa of domestic poultry in sub-Saharan Africa and the isolation of Plasmodium durae Herman from turkeys and francolins in South Africa. Onderstepoort J. Vet. Res. 1993, 60, 15–21. [Google Scholar]
- Bennett, G.F.; Aguirre, A.A.; Cook, R.S. Blood parasites of some birds from northeastern Mexico. J. Parasitol. 1991, 77, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, R.N.; Valkiunas, G.; Iezhova, T.A.; Smith, T.B. Blood parasites of chickens in Uganda and Cameroon with molecular descriptions of Leucocytozoon schoutedeni and Trypanosoma gallinarum. J. Parasitol. 2006, 92, 1336–1343. [Google Scholar] [CrossRef] [PubMed]
- Chawengkirttikul, R.; Junsiri, W.; Watthanadirek, A.; Poolsawat, N.; Minsakorn, S.; Srionrod, N.; Anuracpreeda, P. Molecular detection and genetic diversity of Leucocytozoon sabrazesi in chickens in Thailand. Sci. Rep. 2021, 11, 16686. [Google Scholar] [CrossRef]
- Eide, A.; Fallis, A.M. Experimental studies of the life cycle of Leucocytozoon simondi in ducks in Norway. J. Protozool. 1972, 19, 414–416. [Google Scholar] [CrossRef] [PubMed]
- Valkiūnas, G.; Sruoga, A.; Paulauskas, A. On the geographycal distribution of Leucocytozoon simondi (Haemosporidia: Leucocytozoidae). Parazitologiya 1990, 24, 400–407. (In Russian) [Google Scholar]
- Ramey, A.M.; Buchheit, R.M.; Uher-Koch, B.D.; Reed, J.A.; Pacheco, M.A.; Escalante, A.A. Schmutz, J.A. Negligible evidence for detrimental effects of Leucocytozoon infections among Emperor Geese (Anser canagicus) breeding on the Yukon-Kuskokwim Delta, Alaska. Int. J. Parasitol. Parasites Wildl. 2021, 16, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.M.; Schmutz, J.; Apelgren, C.; Ramey, A.M. A real-time, quantitative PCR protocol for assessing the relative parasitemia of Leucocytozoon in waterfowl. J. Microbiol. Methods 2015, 111, 72–77. [Google Scholar] [CrossRef]
- Argilla, L.S.; Howe, L.; Gartrell, B.D.; Alley, M.R. High prevalence of Leucocytozoon spp. in the endangered yellow-eyed penguin (Megadyptes antipodes) in the sub-Antarctic regions of New Zealand. Parasitology 2013, 140, 672–682. [Google Scholar] [CrossRef]
- Gartrell, B.; Agnew, D.; Alley, M.; Carpenter, T.; Ha, H.J.; Howe, L.; Hunter, S.; McInnes, K.; Munday, R.; Roe, W.; et al. Investigation of a mortality cluster in wild adult yellow-eyed penguins (Megadyptes antipodes) at Otago Peninsula, New Zealand. Avian Pathol. 2017, 46, 278–288. [Google Scholar] [CrossRef]
- Buchheit, R.M.; Schmutz, J.A.; Reed, J.A.; Uher-Koch, B.; Ramey, A.M. Assessment of variation in the detection and prevalence of blood parasites among sympatrically breeding geese in western Alaska, USA. J. Wildl. Dis. 2021, 57, 799–807. [Google Scholar] [CrossRef]
- Van Hemert, C.; Meixell, B.W.; Smith, M.M.; Handel, C.M. Prevalence and diversity of avian blood parasites in a resident northern passerine. Parasites Vectors 2019, 12, 292. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.A.; Sexson, M.G.; Smith, M.M.; Schmutz, J.A.; Ramey, A.M. Evidence for Haemosporidian Parasite Infections in Spectacled Eiders (Somateria fischeri) Sampled in Alaska, USA during the Breeding Season. J. Wildl. Dis. 2018, 54, 877–880. [Google Scholar] [CrossRef] [PubMed]
- Oakgrove, K.S.; Harrigan, R.J.; Loiseau, C.; Guers, S.; Seppi, B.; Sehgal, R.N. Distribution, diversity and drivers of blood-borne parasite co-infections in Alaskan bird populations. Int. J. Parasitol. 2014, 44, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Reeves, A.B.; Smith, M.M.; Meixell, B.W.; Fleskes, J.P.; Ramey, A.M. Genetic diversity and host specificity varies across three genera of blood parasites in ducks of the Pacific Americas Flyway. PLoS ONE 2015, 10, e0116661. [Google Scholar] [CrossRef]
- Dodge, M.; Guers, S.L.; Sekercioğlu, Ç.H.; Sehgal, R.N. North American transmission of hemosporidian parasites in the Swainson’s thrush (Catharus ustulatus), a migratory songbird. J. Parasitol. 2013, 99, 548–553. [Google Scholar] [CrossRef]
- Smith, M.M.; Van Hemert, C.; Merizon, R. Haemosporidian parasite infections in grouse and ptarmigan: Prevalence and genetic diversity of blood parasites in resident Alaskan birds. Int. J. Parasitol. Parasites Wildl. 2016, 5, 229–239. [Google Scholar] [CrossRef]
- Deviche, P.; Greiner, E.C.; Manteca, X. Seasonal and age-related changes in blood parasite prevalence in Dark-eyed Juncos (Junco hyemalis, Aves, Passeriformes). J. Exp. Zool. 2001, 289, 456–466. [Google Scholar] [CrossRef]
- Deviche, P.; Greiner, E.C.; Manteca, X. Interspecific variability of prevalence in blood parasites of adult passerine birds during the breeding season in Alaska. J. Wildl. Dis. 2001, 37, 28–35. [Google Scholar] [CrossRef]
- Deviche, P.; Fokidis, H.B.; Lerbour, B.; Greiner, E. Blood parasitaemia in a high latitude flexible breeder, the white-winged crossbill, Loxia leucoptera: Contribution of seasonal relapse versus new inoculations. Parasitology 2010, 137, 261–273. [Google Scholar] [CrossRef]
- Kleinschmidt, B.; Dorsch, M.; Heinänen, S.; Morkūnas, J.; Schumm, Y.R.; Žydelis, R.; Quillfeldt, P. Prevalence of Haemosporidian Parasites in an Arctic Breeding Seabird Species-The Red-Throated Diver (Gavia stellata). Microorganisms 2022, 10, 2147. [Google Scholar] [CrossRef]
- Krylov, M.V.; Trjapicina, N.P. Blood protozoans of birds from Tadzhikistan. In Bird Diseases. Sbornik trudov Vses. NI inst. po bol. ptits; 1965; Volume 1, pp. 213–221. (In Russian) [Google Scholar]
- Mozaffer, F.; Menon, G.I.; Ishtiaq, F. Exploring the thermal limits of malaria transmission in the western Himalaya. Ecol. Evol. 2022, 12, e9278. [Google Scholar] [CrossRef] [PubMed]
- Barrow, L.N.; McNew, S.M.; Mitchell, N.; Galen, S.C.; Lutz, H.L.; Skeen, H.; Valqui, T.; Weckstein, J.D.; Witt, C.C. Deeply conserved susceptibility in a multi-host, multi-parasite system. Ecol. Lett. 2019, 22, 987–998. [Google Scholar] [CrossRef] [PubMed]
- Gil-Vargas, D.L.; Sedano-Cruz, R.E. Genetic variation of avian malaria in the tropical Andes: A relationship with the spatial distribution of hosts. Malar. J. 2019, 18, 129. [Google Scholar] [CrossRef] [PubMed]
- Mantilla, J.S.; Moncada, L.I.; Matta, N.E.; Adler, P.H. Distribution of black flies (Diptera: Simuliidae) along an elevational gradient in the Andes Mountains of Colombia during the El Niño Southern Oscillation. Acta Trop. 2018, 183, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Imura, T.; Suzuki, Y.; Ejiri, H.; Sato, Y.; Ishida, K.; Sumiyama, D.; Murata, K.; Yukawa, M. Prevalence of avian haematozoa in wild birds in a high-altitude forest in Japan. Vet. Parasitol. 2012, 183, 244–248. [Google Scholar] [CrossRef]
- Illera, J.C.; López, G.; García-Padilla, L.; Moreno, Á. Factors governing the prevalence and richness of avian haemosporidian communities within and between temperate mountains. PLoS ONE 2017, 12, e0184587. [Google Scholar] [CrossRef]
- Dunbar, M.R.; Tornquist, S.; Giordano, M.R. Blood parasites in sage-grouse from Nevada and Oregon. J. Wildl. Dis. 2003, 39, 203–208. [Google Scholar] [CrossRef]
- Murata, K.; Tamada, A.; Ichikawa, Y.; Hagihara, M.; Sato, Y.; Nakamura, H.; Nakamura, M.; Sakanakura, T.; Asakawa, M. Geographical distribution and seasonality of the prevalence of Leucocytozoon lovati in Japanese rock ptarmigans (Lagopus mutus japonicus) found in the alpine regions of Japan. J. Vet. Med. Sci. 2007, 69, 171–176. [Google Scholar] [CrossRef]
- Theodosopoulos, A.N.; Spellman, G.M.; Taylor, S.A. Survey of haemosporidian parasite infections in an endangered high alpine bird. Parasites Vectors 2023, 16, 67. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A. Distribution of lethal haemosporidoses pathogens in poultry. Parazitologiya 2001, 35, 27–34. (In Russian) [Google Scholar]
- Valkiūnas, G.; Atkinson, C.T. Introduction to life cycles, taxonomy, distribution and basic research techniques. In Avian Malaria and Related Parasites in the Tropics: Ecology, Evolution and Systematics; Springer Nature Switzerland AG: Cham, Switzerland, 2020; pp. 45–80. [Google Scholar] [CrossRef]
- Cook, R.S. Leucocytozoon Danilewsky 1890. In Infectious and Parasitic Diseases of Wild Birds; Davis, J.W., Anderson, R.C., Karstad, L., Trainer, D.O., Eds.; Iowa State University Press: Ames, IA, USA, 1971; pp. 291–299. [Google Scholar]
- Fallis, A.M.; Desser, S.S.; Khan, R.A. On species of Leucocytozoon. Adv. Parasitol. 1974, 12, 1–67. [Google Scholar] [PubMed]
- Atkinson, C.T.; van Riper, C. Pathogenicity and epizootiology of avian haematozoa: Plasmodium, Leucocytozoon, and Haemoproteus. In Bird–Parasite Interactions: Ecology, Evolution, and Behavior; Loye, J.E., Zuk, M., Eds.; Oxford University Press: Oxford, UK, 1991; pp. 19–48. [Google Scholar]
- Greiner, E.C. Leucocytozoonosis in waterfowl and wild galliform birds. Bull. Soc. Vector Ecol. 1991, 16, 84–93. [Google Scholar]
- Sambon, L.W. Remarks on the avian haemoprotozoa of the genus Leucocytozoon Danilewsky. J. Trop. Med. Hyg. 1908, 11, 245–248, 325–328. [Google Scholar]
- Coatney, G.R. A catalog and host-index of the genus Leucocytozoon. J. Parasitol. 1937, 23, 202–212. [Google Scholar] [CrossRef]
- Herman, C.M.; Greiner, E.C.; Bennett, G.F.; Laird, M. Bibliography of the Avian Blood-Inhabiting Protozoa; Memorial University of Newfoundland: St. John’s, NL, Canada, 1976; 123p. [Google Scholar]
- Bennett, G.F.; Whiteway, M.; Woodworth-Lynas, C. A host-parasite catalogue of the avian haematozoa. Mem. Univ. Nfld. Occ. Pap. Biol. 1982, 5, 1–243. [Google Scholar]
- Garnham, P.C.C.; Duggan, A.J. Catalogue of the Garnham Collection of Malaria Parasites and Other Haemosporidia; The Wellcome Trust: London, UK, 1986; 191p. [Google Scholar]
- Bennett, G.F.; Earlé, R.A.; Du Toit, H.; Huchzermeyer, F.W. A host–parasite catalogue of the haematozoa of the sub-Saharan birds. Onderstepoort J. Vet. Res. 1992, 59, 1–73. [Google Scholar]
- Bishop, M.A.; Bennett, G.F. Host-parasite catalogue of the avian haematozoa: Supplement 1, and Bibliography of the avian blood-inhabiting haematozoa. Mem. Univ. Nfld. Occ. Pap. Biol. 1992, 15, 1–244. [Google Scholar]
- Bennett, G.F.; Kučera, J.; Woodworth-Lynas, C.; Whiteway, M. Bibliography of the avian blood-inhabiting Protozoa. Supplement 1. Mem. Univ. Nfld. Occ. Pap. Biol. 1981, 4, 1–33. [Google Scholar]
- Akiba, K. Leucocytozoonosis of chickens. Nat. Inst. Anim. Hlth. Quart. 1970, 10, 131–147. [Google Scholar]
- Morii, T. A review of Leucocytozoon caulleryi infection in chickens. J. Protozool. Res. 1992, 2, 128–133. [Google Scholar]
- Morii, T.; Matsui, T.; Kobayashi, F.; Iijima, T.; Fukuda, M. Freeze fracture study of the schizonts and merozoites of Leucocytozoon caulleryi. Zbl. Bakt. Hyg. A 1987, 264, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Isobe, T.; Akiba, K. Early schizonts of Leucocytozoon caulleryi. J. Parasitol. 1990, 76, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Ilgūnas, M.; Himmel, T.; Harl, J.; Dagys, M.; Valkiūnas, G.; Weissenböck, H. Exo-Erythrocytic Development of Avian Haemosporidian Parasites in European Owls. Animals 2022, 12, 2212. [Google Scholar] [CrossRef]
- Desser, S.S. Schizogony and gametogony of Leucocytozoon simondi and associated reactions in the avian host. J. Protozool. 1967, 14, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.A.; Desser, S.S.; Fallis, A.M. Survival of sporozoites of Leucocytozoon in birds for 11 days. Can. J. Zool. 1969, 47, 347–350. [Google Scholar] [CrossRef]
- Khan, R.A.; Fallis, A.M. Life cycles of Leucocytozoon dubreuili Mathis and Léger, 1911 and L. fringillinarum Woodcock, 1910 (Haemosporidia: Leucocytozoidae). J. Protozool. 1970, 17, 642–658. [Google Scholar] [CrossRef]
- Khan, R.A.; Fallis, A.M. A note on the sporogony of Parahaemoproteus velans (=Haemoproteus velans Coatney and Roudabush) (Haemosporidia: Haemoproteidae) in species of Culicoides. Can. J. Zool. 1971, 49, 420–421. [Google Scholar] [CrossRef]
- Khan, R.A.; Fallis, A.M. Speciation, transmission, and schizogony of Leucocytozoon in corvid birds. Can. J. Zool. 1971, 49, 1363–1367. [Google Scholar] [CrossRef]
- Khan, R.A. Development of Leucocytozoon ziemanni (Laveran). J. Parasitol. 1975, 61, 449–457. [Google Scholar] [CrossRef]
- Allison, F.R.; Desser, S.S.; Whitten, L.K. Further observations on the life cycle and vectors of the haemosporidian Leucocytozoon tawaki and its transmission to the Fiordland crested penguin. N. Z. J. Zool. 1978, 5, 371–374. [Google Scholar] [CrossRef]
- Steele, E.J.; Noblet, G.P. Schizogonic development of Leucocytozoon smithi. J. Protozool. 1992, 39, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.W. Schizogony and gametocyte development of Leucocytozoon berestneffi in the yellow-billed magpie, Pica nuttalli. J. Protozool. 1965, 12, 584–589. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A. Exo-erythrocytic development of avian malaria and related haemosporidian parasites. Malar. J. 2017, 16, 101. [Google Scholar] [CrossRef] [PubMed]
- Desser, S.S. The fine structure of Leucocytozoon simondi. VI. Hepatic schizogony. Can. J. Zool. 1973, 51, 605–609. [Google Scholar] [CrossRef]
- Desser, S.S.; Fallis, A.M. The cytological development and encapsulation of megaloschizonts of Leucocytozoon simondi. Can. J. Zool. 1967, 45, 1061–1065. [Google Scholar] [CrossRef]
- Huff, C.G. Schizogony and gametocyte development in Leucocytozoon simondi, and comparison with Plasmodium and Haemoproteus. J. Infect. Dis. 1942, 71, 18–32. [Google Scholar] [CrossRef]
- Wingstrand, K.G. On some haematozoa of Swedish birds with remarks on the schizogony of Leucocytozoon sakharoffi. Kungl. Sven. Vetenskapsakad. Handl. 1947, 24, 1–31. [Google Scholar]
- Wingstrand, K.G. Further studies on Leucocytozoon sakharoffi. Kungl. Sven. Vetenskapsakad Handl. 1948, 24, 1–17. [Google Scholar]
- Hill, A.G.; Howe, L.; Gartrell, B.D.; Alley, M.R. Prevalence of Leucocytozoon spp., in the endangered yellow-eyed penguin Megadyptes antipodes. Parasitology 2010, 137, 1477–1485. [Google Scholar] [CrossRef]
- Harl, J.; Himmel, T.; Valkiūnas, G.; Ilgūnas, M.; Nedorost, N.; Matt, J.; Kübber-Heiss, A.; Alic, A.; Konicek, C.; Weissenböck, H. Avian haemosporidian parasites of accipitriform raptors. Malar. J. 2022, 21, 14. [Google Scholar] [CrossRef]
- Himmel, T.; Harl, J.; Kübber-Heiss, A.; Konicek, C.; Fernández, N.; Juan-Sallés, C.; Ilgūnas, M.; Valkiūnas, G.; Weissenböck, H. Molecular probes for the identification of avian Haemoproteus and Leucocytozoon parasites in tissue sections by chromogenic in situ hybridization. Parasites Vectors 2019, 12, 282. [Google Scholar] [CrossRef] [PubMed]
- Himmel, T.; Harl, J.; Pfanner, S.; Nedorost, N.; Nowotny, N.; Weissenböck, H. Haemosporidioses in wild Eurasian blackbirds (Turdus merula) and song thrushes (T. philomelos): An in situ hybridization study with emphasis on exo-erythrocytic parasite burden. Malar. J. 2020, 19, 69. [Google Scholar] [CrossRef] [PubMed]
- Dick, J.W.; Rice, J.T. The persistence of gametocytes of Leucocytozoon smithi in peripheral turkey blood. Poult. Sci. 1975, 54, 1753–1754. [Google Scholar] [CrossRef]
- Alverson, D.R.; Noblet, R. Spring relapse of Leucocytozoon smithi (Sporozoa: Leucocytozoidae) in turkeys. J. Med. Entomol. 1977, 14, 132–133. [Google Scholar] [CrossRef]
- Desser, S.S.; Fallis, A.M.; Garnham, P.C.C. Relapses in ducks chronically infected with Leucocytozoon simondi and Parahaemoproteus nettionis. Can. J. Zool. 1968, 46, 281–285. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Ilgūnas, M.; Bukauskaitė, D.; Žiegytė, R.; Bernotienė, R.; Jusys, V.; Eigirdas, V.; Fragner, K.; Weissenböck, H.; Iezhova, T.A. Plasmodium delichoni n. sp.: Description, molecular characterisation and remarks on the exoerythrocytic merogony, persistence, vectors and transmission. Parasitol. Res. 2016, 115, 2625–2636. [Google Scholar] [CrossRef]
- Khan, R.A.; Fallis, A.M. Relapses in birds infected with species of Leucocytozoon. Can. J. Zool. 1970, 48, 451–455. [Google Scholar] [CrossRef]
- Clarke, C.H.D. Organisms of a malarial type in ruffed grouse, with a description of the schizogony of Leucocytozoon bonasae. J. Wildl. Manag. 1938, 2, 146–150. [Google Scholar] [CrossRef]
- Chernin, E. The relapse phenomenon in the Leucocytozoon simondi infection of the domestic duck. Am. J. Hyg. 1952, 56, 101–118. [Google Scholar]
- Allan, R.A.; Mahrt, J.L. Influence of transmission period on primary and relapse patterns of infection of Leucocytozoon spp. and Haemoproteus mansoni. Am. Midl. Nat. 1989, 121, 341–349. [Google Scholar] [CrossRef]
- Noblet, G.P.; Noblet, R. Periodicity of Leucocytozoon smithi gametocytes in the peripheral blood of domestic turkeys. Poult. Sci. 1976, 55, 1088–1093. [Google Scholar] [CrossRef] [PubMed]
- Chagas, C.R.F.; Binkienė, R.; Valkiūnas, G. Description and Molecular Characterization of Two Species of Avian Blood Parasites, with Remarks on Circadian Rhythms of Avian Haematozoa Infections. Animals 2021, 11, 3490. [Google Scholar] [CrossRef]
- Carter, R.; Graves, P.M. Gametocytes. In Malaria. Principles and Practice of Malariology; Wernsdorfer, W.H., McGregor, I., Eds.; Churchill Livingston: Edinburgh, UK, 1988; pp. 253–305. [Google Scholar]
- Steele, E.J.; Noblet, G.P.; Noblet, R. Sporogonic development of Leucocytozoon smithi. J. Protozool. 1992, 39, 690–699. [Google Scholar] [CrossRef] [PubMed]
- Fallis, A.M.; Bennett, G.F. Observations on the sporogony of Leucocytozoon mirandae, L. bonasae, and L. fringillinarum (Sporozoa: Leucocytozoidae). Can. J. Zool. 1962, 40, 395–400. [Google Scholar] [CrossRef]
- Desser, S.S.; Fallis, A.M.; Allison, F.R. Nuclear changes preceding microgamete formation in Leucocytozoon simondi and Leucocytozoon tawaki. Can. J. Zool. 1976, 54, 799–801. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A. A comparison of the blood parasites in three subspecies of the yellow wagtail Motacilla flava. J. Parasitol. 2001, 87, 930–934. [Google Scholar] [CrossRef]
- Ashford, R.W.; Wyllie, I.; Newton, I. Leucocytozoon toddi in British sparrowhawks Accipiter nisus: Observations on the dynamics of infection. J. Nat. Hist. 1990, 24, 1101–1107. [Google Scholar] [CrossRef]
- Ashford, R.W.; Green, E.E.; Holmes, P.R.; Lucas, A.J. Leucocytozoon toddi in British sparrowhawks Accipiter nisus: Patterns of infection in nestlings. J. Nat. Hist. 1991, 25, 269–277. [Google Scholar] [CrossRef]
- Ishtiaq, F.; Renner, S.C. Bird migration and vector-borne parasite transmission. In Avian Malaria and Related Parasites in the Tropics: Ecology, Evolution and Systematics; Santiago-Alarcon, D., Marzal, A., Eds.; Springer: Berlin, Germany, 2020; pp. 513–526. [Google Scholar]
- Valkiūnas, G. The role of seasonal migrations in the distribution of Haemosporidia of birds in North Palearctic. Ekologija 1993, 2, 57–67. [Google Scholar]
- Jenkins, T.; Owens, I.P. Biogeography of avian blood parasites (Leucocytozoon spp.) in two resident hosts across Europe: Phylogeographic structuring or the abundance-occupancy relationship? Mol. Ecol. 2011, 20, 3910–3920. [Google Scholar] [CrossRef]
- Vysotsky, V.G.; Valkiūnas, G. Investigation of the long-distance dispersal of migratory passerine birds using blood parasites as biological tags. Proc. Zool. Inst. Russ. Acad. Sci. 2001, 289, 173–180. [Google Scholar]
- Valkiūnas, G.; Iezhova, T.A.; Loiseau, C.; Sehgal, R.N. Nested cytochrome B polymerase chain reaction diagnostics detect sporozoites of hemosporidian parasites in peripheral blood of naturally infected birds. J. Parasitol. 2009, 95, 1512–1515. [Google Scholar] [CrossRef] [PubMed]
- De Amaral, F.; Wilson, R.E.; Sonsthagen, S.A.; Sehgal, R. Diversity, distribution, and methodological considerations of haemosporidian infections among Galliformes in Alaska. Int. J. Parasitol. Parasites Wildl. 2023, 20, 122–132. [Google Scholar] [CrossRef]
- Starkloff, N.C.; Galen, S.C. Coinfection rates of avian blood parasites increase with latitude in parapatric host species. Parasitology 2023, 150, 329–336. [Google Scholar] [CrossRef]
- Bennett, G.F.; Huchzermeyer, F.W.; Burger, W.P.; Earlé, R.A. The Leucocytozoidae of South African birds. Redescription of Leucocytozoon struthionis Walker, 1912. Ostrich 1992, 63, 83–85. [Google Scholar] [CrossRef]
- Ortiz-Catedral, L.; Brunton, D.; Stidworthy, M.F.; Elsheikha, H.M.; Pennycott, T.; Schulze, C.; Braun, M.; Wink, M.; Gerlach, H.; Pendl, H.; et al. Haemoproteus minutus is highly virulent for Australasian and South American parrots. Parasites Vectors 2019, 12, 40. [Google Scholar] [CrossRef] [PubMed]
- Peirce, M.A. Haematozoa of Zambian birds. IV. Description of Leucocytozoon balmorali sp. nov. from Malaconotidae. J. Nat. Hist. 1984, 18, 223–226. [Google Scholar] [CrossRef]
- Valkiūnas, G. Leucocytozoon bennetti sp. n. (Haemosporidia: Leucocytozoidae) from roller. Parazitologiya 1993, 27, 436–440. (In Russian) [Google Scholar]
- Coatney, G.R.; Roudabush, R.L. Some blood parasites from Nebraska birds. Am. Midl. Nat. 1937, 18, 1005–1030. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Sehgal, R.N.; Iezhova, T.A.; Hull, A.C. Identification of Leucocytozoon toddi group (Haemosporida: Leucocytozoidae), with remarks on the species taxonomy of leucocytozoids. J. Parasitol. 2010, 96, 170–177. [Google Scholar] [CrossRef]
- Kerandel, J. Trypanosomes et Leucocytozoon observés chez des oiseaux du Congo. Ann. Inst. Pasteur 1913, 27, 421–439. [Google Scholar]
- Omori, S.; Sato, Y.; Hirakawa, S.; Isobe, T.; Yukawa, M.; Murata, K. Two extra chromosomal genomes of Leucocytozoon caulleryi; complete nucleotide sequences of the mitochondrial genome and existence of the apicoplast genome. Parasitol. Res. 2008, 103, 953–957. [Google Scholar] [CrossRef] [PubMed]
- Fantham, H.B.; Cantab, M.A.; Lond, D.S. Some parasitic Protozoa found in South Africa: IV. S. Afr. J. Sci. 1921, 18, 164–170. [Google Scholar]
- Coatney, G.R.; West, E. Some blood parasites from Nebraska birds—II. Am. Midl. Nat. 1938, 19, 601–612. [Google Scholar] [CrossRef]
- Bennett, G.F.; Earlé, R.A.; Peirce, M.A.; Nandi, N.C. The Leucocytozoidae of South African birds. The Coliiformes and Coraciiformes. S.-Afr. Tydskr. Dierk. 1993, 28, 74–80. [Google Scholar] [CrossRef]
- Valkiūnas, G. Parasitic Protozoa of the blood of birds in the USSR. (4. Leucocytozoidae of Columbiformes, Caprimulgiformes, Coraciiformes, Upupiformes, Charadriiformes and Galliformes). Lietuvos TSR MA Darbai C Serija 1989, 2, 80–94. (In Russian) [Google Scholar]
- Ziemann, H. Ueber Malaria-und Andere Blutparasiten Nebst Anhang. Eine Wirksame Methode der Chromatin-und Blutfärbung; Fischer: Jena, Germany, 1898. [Google Scholar]
- Tendeiro, J. Acerca dos hematozoários de algumas aves da Guiné Portuguesa. Rev. Med. Vet. 1947, 42, 287–350. [Google Scholar]
- Mathis, C.; Léger, M. Recherches de Parasitologie et de Pathologie Humaines et Animales au Tonkin; Masson: Paris, France, 1911. [Google Scholar]
- Pacheco, M.A.; Cepeda, A.S.; Bernotienė, R.; Lotta, I.A.; Matta, N.E.; Valkiūnas, G.; Escalante, A.A. Primers targeting mitochondrial genes of avian haemosporidians: PCR detection and differential DNA amplification of parasites belonging to different genera. Int. J. Parasitol. 2018, 48, 657–670. [Google Scholar] [CrossRef]
- Woodcook, H.M. Studies on avian haematozoa. I. On certain parasites of the chaffinch (Fringilla coelebs) and the redpoll (Linota rufescens). Q. J. Microsc. Sci. 1910, 55, 641–740. [Google Scholar] [CrossRef]
- Bennett, G.F.; Khan, R.A.; Campbell, A.G. Leucocytozoon grusi sp. n. (Sporozoa: Leucocytozoidae) from a sandhill crane, Grus canadensis (L.). J. Parasitol. 1974, 60, 359–363. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A.; Mironov, S.V. Leucocytozoon hamiltoni n. sp. (Haemosporida, Leucocytozoidae) from the Bukharan great tit Parus bokharensis. J. Parasitol. 2002, 88, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Mathis, C.; Léger, M. Leucocytozoon d’un paon, d’un crabier et d’um bengali du Tonkin. C. R. Séances Soc. Biol. Fil. 1911, 70, 211–212. [Google Scholar]
- França, C. Leucocytozoon du geai, de l’épervier et la bécasse. Bull. Soc. Pathol. Exot. 1912, 5, 17–21. [Google Scholar]
- Seligmann, C.G.; Sambon, L.W. Preliminary note on a Leucocytozoon found in the blood of the reg grouse (Lagopus scoticus). Lancet 1907, 173, 829–830. [Google Scholar] [CrossRef]
- Sato, Y.; Hagihara, M.; Yamaguchi, T.; Yukawa, M.; Murata, K. Phylogenetic comparison of Leucocytozoon spp. from wild birds of Japan. J. Vet. Med. Sci. 2007, 69, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Greiner, E.C. Leucocytozoon maccluri sp. n. (Haemosporida: Leucocytozoidae) from a Thailand thrush, Zoothera marginata Blyth. J. Parasitol. 1976, 62, 545–547. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, J.; Xu, R.; Zhang, C.; Pang, Q.; Chen, X.; Liu, S.; Hong, L.; Yuan, J.; Li, X.; et al. The Gametocytes of Leucocytozoon sabrazesi Infect Chicken Thrombocytes, Not Other Blood Cells. PLoS ONE 2015, 10, e0133478. [Google Scholar] [CrossRef]
- Laveran, A. Sur une Haemamoeba d’une mésange (Parus major). C. R. Séances Soc. Biol. Fil. 1902, 54, 1121–1124. [Google Scholar]
- Mathis, C.; Léger, M. Leucocytozoon d’une Tourterelle (Turtur humilis) et d’une Sarcelle (Querquedula crecca) du Tonkin. C. R. Séances Soc. Biol. Fil. 1910, 68, 118–120. [Google Scholar]
- Covaleda Ortega, J.; Gállego Berenguer, J. Contribucion al estudio de algunos Leucocytozoon parasitos de aves españolas. Rev. Ibér. Parasitol. 1946, 6, 203–224. [Google Scholar]
- Balfour, A. Report of the travelling pathologist and protozoologist. Second report. Rep. Wellcome Res. Lab. Khartoum. 1906, 2, 183–204. [Google Scholar]
- Shamsuddin, M.; Mohammad, M.K. Haematozoa of some Iraqi birds with description of two new species, Haemoproteus pteroclis and Leucocytozoon nycticoraxi (Protozoa: Haemosporina). Bull. Nat. Hist. Res. Cent. 1980, 7, 111–155. [Google Scholar]
- Nandi, N.C. Leucocytozoon nyctyornis n. sp. from bluebeared bee-eater Nyctyornis athertoni (Jardine and Selby). Arch. Protistenkd. 1986, 132, 113–117. [Google Scholar] [CrossRef]
- Adlard, R.D.; Peirce, M.A.; Lederer, R. New species of Leucocytozoon from the avian families Otidae, Podargidae and Threskiornithidae. J. Nat. Hist. 2002, 36, 1261–1267. [Google Scholar] [CrossRef]
- Jiang, Y.; Brice, B.; Nguyen, M.; Loh, R.; Greay, T.; Adlard, R.; Ryan, U.; Yang, R. Further characterisation of Leucocytozoon podargii in wild tawny frogmouths (Podargus strigoides) in Western Australia. Parasitol. Res. 2019, 118, 1833–1840. [Google Scholar] [CrossRef] [PubMed]
- Lotta, I.A.; Gonzalez, A.D.; Pacheco, M.A.; Escalante, A.A.; Valkiūnas, G.; Moncada, L.I.; Matta, N.E. Leucocytozoon pterotenuis sp. nov. (Haemosporida, Leucocytozoidae): Description of the morphologically unique species from the Grallariidae birds, with remarks on the distribution of Leucocytozoon parasites in the Neotropics. Parasitol. Res. 2015, 114, 1031–1044. [Google Scholar] [CrossRef] [PubMed]
- Rodhain, J.; Pons, C.; Vandenbranden, F.; Bequaert, J. Rapport sur les Travaux de la Mission Scientifique du Katanga (Octobre 1910 à Septembre 1912); Hayez: Bruxelles, Belgium, 1913. [Google Scholar]
- Tartakovsky, M.G. Explanation of the Laboratory Exhibits at the All-Russian Hygienic Exhibition in St. Petersburg, May-October 1913; St. Petersburg, Russia, 1913. (In Russian) [Google Scholar]
- Knuth, P.; Magdeburg, F. Über ein durch Leukozytozoen verursachtes Sterben junger Gänse. Berl. Tierärztl. Wochenschr. 1922, 31, 359–361. [Google Scholar]
- Laveran, A.; Lucet, E. Deux hématozoaires de la perdrix ei du dindon. C. R. Acad. Sci. 1905, 141, 673–676. [Google Scholar]
- Nandi, N.C. Leucocytozoon squamatus sp. n. from scaly-bellied green woodpecker, Picus squamatus squamatus Vigors. Acta Protozool. 1986, 25, 223–226. [Google Scholar]
- Walker, J. Über ein Leucocytozoon beim Vögel. Z. Infekt. Haustier. 1912, 12, 372–375. [Google Scholar]
- Fallis, A.M.; Bisset, S.A.; Allison, F.R. Leucocytozoon tawaki n. sp. (Eucoccida: Leucocytozoidae) from the penguin Eudyptes pachyrhynchus, and preliminary observations on its development in Austrosimulium spp. (Diptera: Simuliidae). N. Z. J. Zool. 1976, 3, 11–16. [Google Scholar] [CrossRef]
- Greiner, E.C.; Kocan, A.A. Leucocytozoon (Haemosporida; Leucocytozoidae) of the Falconiformes. Can. J. Zool. 1977, 55, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Rodhain, J. Deux Leucocytozoon d’oiseaux du Congo belge. C. R. Hebd. Séances Mem. Soc. Biol. 1931, 107, 273–276. [Google Scholar]
- Peirce, M.A.; Adlard, R.D.; Lederer, R. A new species of Leucocytozoon Berestneff, 1904 (Apicomplexa: Leucocytozoidae) from the avian family Artamidae. Syst. Parasitol. 2005, 60, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Savage, A.F. Hematozoa of the avian family Philepittidae (the asities and sunbird asities). J. Parasitol. 2004, 90, 1473–1474. [Google Scholar] [CrossRef]
- Savage, A.F.; Ariey, F.; Greiner, E.C. Leucocytozoon atkinsoni n. sp. (Apicomplexa: Leucocytozoidae) from the avian family Timaliidae. Syst. Parasitol. 2006, 64, 105–109. [Google Scholar] [CrossRef]
- Barraclough, R.K.; Robert, V.; Peirce, M.A. New species of haematozoa from the avian families Campephagidae and Apodidae. Parasite 2008, 15, 105–110. [Google Scholar] [CrossRef]
- Jones, H.I.; Sehgal, R.N.; Smith, T.B. Leucocytozoon (Apicomplexa: Leucocytozoidae) from West African birds, with descriptions of two species. J. Parasitol. 2005, 91, 397–401. [Google Scholar] [CrossRef]
- International Commission on Zoological Nomenclature. Opinion 1971. Leucocytozoon (Protista, Haemosporida): Berestneff (1904) adopted as the author and date, and Leukocytozoen danilewskyi Ziemann, 1898 adopted as the type species. Bull. Zool. Nom. 2001, 58, 145–147. [Google Scholar]
- Ortego, J.; Cordero, P.J. PCR-based detection and genotyping of haematozoa (Protozoa) parasitizing eagle owls, Bubo bubo. Parasitol. Res. 2009, 104, 467–470. [Google Scholar] [CrossRef]
- Perkins, S.L. Molecular systematics of the three mitochondrial protein-coding genes of malaria parasites: Corroborative and new evidence for the origins of human malaria. Mitochondrial DNA 2008, 19, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Lotta, I.A.; Matta, N.E.; Torres, R.D.; Sandino, M.M.; Moncada, L.I. Leucocytozoon fringillinarum and Leucocytozoon dubreuili in Turdus fuscater from a Colombian Páramo ecosystem. J. Parasitol. 2013, 99, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.A.; González-Acuña, D.; Tkach, V.V. Haemosporidian Parasites of Chilean Ducks: The Importance of Biogeography and Nonpasserine Hosts. J. Parasitol. 2020, 106, 211–220. [Google Scholar] [CrossRef]
- Barraclough, R.K.; Duval, L.; Talman, A.M.; Ariey, F.; Robert, V. Attraction between sexes: Male-female gametocyte behaviour within a Leucocytozoon toddi (Haemosporida). Parasitol. Res. 2008, 102, 1321–1327. [Google Scholar] [CrossRef]
- Sehgal, R.N.; Hull, A.C.; Anderson, N.L.; Valkiūnas, G.; Markovets, M.J.; Kawamura, S.; Tell, L.A. Evidence for cryptic speciation of Leucocytozoon spp. (Haemosporida, Leucocytozoidae) in diurnal raptors. J. Parasitol. 2006, 92, 375–379. [Google Scholar] [CrossRef]
- Wong, T.C.; Desser, S.S. Ultrastructural observations on renal schizogony of Leucocytozoon dubreuili in the American robin. J. Protozool. 1978, 25, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Desser, S.S.; Baker, J.R.; Lake, P. The fine structure of Leucocytozoon simondi. I. Gametogenesis. Can. J. Zool. 1970, 48, 331–336. [Google Scholar] [CrossRef]
- Kocan, A.A.; Kocan, K.M. The fine structure of elongate gametocytes of Leucocytozoon ziemanni (Laveran). J. Parasitol. 1978, 64, 1057–1059. [Google Scholar] [CrossRef]
- Zachos, F.E. Species Concepts in Biology: Historical Development, Theoretical Foundations and Practical Relevance; Springer Nature: Cham, Switzerland, 2016; p. 220. [Google Scholar] [CrossRef]
- Santiago-Alarcon, D.; Palinauskas, V.; Schaefer, H.M. Diptera vectors of avian Haemosporidian parasites: Untangling parasite life cycles and their taxonomy. Biol. Rev. Camb. Philos. Soc. 2012, 87, 928–964. [Google Scholar] [CrossRef]
- Fallis, A.M.; Desser, S.S. On species of Leucocytozoon, Haemoproteus, and Hepatocystis. In Parasitic Protozoa; Kreier, J.P., Ed.; Academic Press: New York, NY, USA, 1977; Volume 3, pp. 239–266. [Google Scholar]
- Levine, N.D. The Protozoan Phylum Apicomplexa; Part 2; CRC Press: Boca Raton, FL, USA, 1988; 168p. [Google Scholar]
- Hsu, C.-K.; Campbell, G.R.; Levine, N.D. A check-list of the species of the genus Leucocytozoon (Apicomplexa, Plasmodiidae). J. Protozool. 1973, 20, 195–203. [Google Scholar] [CrossRef]
- Yang, G.; He, H.; Zhang, G.; Zhao, W.; Zhou, J.; Qian, Y.; Huang, X.; Dong, L. Neglected parasite reservoirs in wetlands: Prevalence and diversity of avian haemosporidians in waterbird communities in Northeast China. Int. J. Parasitol. Parasites Wildl. 2021, 29, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Harl, J.; Himmel, T.; Valkiūnas, G.; Ilgūnas, M.; Bakonyi, T.; Weissenböck, H. Geographic and host distribution of haemosporidian parasite lineages from birds of the family Turdidae. Malar. J. 2020, 19, 335. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, C.T.; Greiner, E.C.; Forrester, D.J. Pre-erythrocytic development and associated host responses to Haemoproteus meleagridis (Haemosporina: Haemoproteidae) in experimentally infected domestic turkeys. J. Protozool. 1986, 33, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Morii, T.; Fukuda, M. Observations on first-generation schizogony of Leucocytozoon caulleryi in chickens. J. Protozool. 1992, 39, 281–287. [Google Scholar] [CrossRef]
- Atkinson, C.T. Vectors, epizootiology, and pathogenicity of avian species of Haemoproteus (Haemosporina: Haemoproteidae). Bull. Soc. Vector Ecol. 1991, 16, 109–126. [Google Scholar]
- Lainson, R.; Schaw, J.J. A new haemosporidian of lizards, Saurocytozoon tupinambi gen. nov., sp. nov., in Tupinambus nigropunctatus (Teiidae). Parasitology 1969, 59, 159–162. [Google Scholar] [CrossRef]
- Lainson, G. A protozoologist in Amazonia: Neglected parasites, with particular reference to members of the Coccidia (Protozoa: Apicomplexa). Ciência Cult. 1992, 44, 81–93. [Google Scholar]
- Lainson, R.; Landau, I.; Shaw, J.J. Further parasites of the family Garniidae (Coccidiida: Haemosporidiidea) in Brazilian lizards. Fallisia effusa gen. nov., sp. nov. and Fallisia modesta gen. nov., sp. nov. Parasitology 1974, 68, 117–125. [Google Scholar] [CrossRef]
- Valkiūnas, G. Haemosporidians. In Protista: Handbook on Zoology; Nauka Press: St. Peterburg, Russia, 2007; Volume 2, pp. 256–330. (In Russian) [Google Scholar]
- Lainson, R. Atlas of Protozoan Parasites of the Amazonian Fauna of Brazil. Haemosporida of Reptiles; Instituto Evandro Chagas: Ananindeua, Brazil, 2012; Volume 1. [Google Scholar]
- Telford, S.R. The Hemoparasites of the Reptilian; CRC Press: Boca Raton, FL, USA, 2009; 376p. [Google Scholar]
- Berestneff, N. Über das Leucocytozoon Danilewskyi. Arch. Protistenkd. 1904, 3, 376–386. [Google Scholar]
- Valkiūnas, G. Bird Haemosporida. Acta Zool. Lit.; 3-5: A monograph; Institute of Ecology: Vilnius, Lithuania, 1997; 607p. (In Russian) [Google Scholar]
- Valkiūnas, G. Leucocytozoon (Protista, Haemosporida): Proposed adoption of Berestneff, 1904 as the author and of Leukocytozoen danilewskyi Ziemann, 1898 as the type species (Case 3089). Bull. Zool. Nom. 1999, 56, 168–170. [Google Scholar] [CrossRef]
- International Commission on Zoological Nomenclature. International Code of Zoological Nomenclature, 4th ed.; The International Trust for Zoological Nomenclature: London, UK, 1999. [Google Scholar]
- Sambon, L.W. Remarks on the avian haemoprotozoa of the genus Leucocytozoon. J. Trop. Med. Hyg. 1909, 12, 37–38. [Google Scholar]
- Léger, M.; Mathis, C. Leucocytozoon de la perorix du Tonkin. Ann. Inst. Pasteur Paris 1909, 23, 740–743. [Google Scholar]
- Prowazek, S.v. Beiträge zur Kenntnis der Protozoen und verwandter Organismen von Sumatra. Arch. Protistenkd. 1912, 26, 250–274. [Google Scholar]
- Léger, M. Hématozoaires d’oiseaux de la Corse. Bull. Soc. Pathol. Exot. 1913, 6, 515–523. [Google Scholar]
- Mathis, C.; Léger, M. Parasites sanguicoles d’un passerean du Tonkin (Ixus Hainanus, boulboul de l’lle d’Hainan). C. R. Séances Soc. Biol. Fil. 1910, 69, 30–32. [Google Scholar]
- de Mello, I.F. A contribution to the study of the blood parasites of some Indian birds. Proc. Indian Acad. Sci. (Sec. B) 1935, 1, 349–358. [Google Scholar] [CrossRef]
- Morii, T.; Kitaoka, S. Susceptibility of the gallinaceous birds to Akiba caulleryi. Nat. Inst. Anim. Hlth. Quart. 1971, 11, 170–171. [Google Scholar]
- Solis, J. Nonsusceptibility of some avian species to turkey Leucocytozoon infection. Poult. Sci. 1973, 52, 498–500. [Google Scholar] [CrossRef]
- Desser, S.S.; Ryckman, A.K. The development and pathogenesis of Leucocytozoon simondi in Canada and domestic geese in Algonquin Park, Ontario. Can. J. Zool. 1976, 54, 634–643. [Google Scholar] [CrossRef]
- Desser, S.S.; Stuht, J.; Fallis, A.M. Leucocytozoonosis in Canada geese in Upper Michigan. I. Strain differences among geese from different localities. J. Wildl. Dis. 1978, 14, 124–131. [Google Scholar] [CrossRef]
- Fallis, A.M.; Bennett, G.F. On the epizootiology of infections caused by Leucocytozoon simondi in Algonquin Park, Canada. Can. J. Zool. 1966, 44, 101–112. [Google Scholar] [CrossRef]
- Stephan, J. Ueber eine durch Leukozytozoen verursachte Ganseund Putenerkrankung. Dtsch. Tieraerztl. Wochenschr. 1922, 30, 589–592. [Google Scholar]
- Fallis, A.M.; Bennett, G.F.; Ritchie, R.C. Observations and experiments on Leucocytozoon with special reference to L. simondi Mathis and Léger, in ducks. In Proceedings of the XIV International Congress of Zoology, Copenhagen, Denmark, 5–12 August 1953. [Google Scholar]
- Fallis, A.M.; Pearson, J.C.; Bennett, G.F. On the specificity of Leucocytozoon. Can. J. Zool. 1954, 32, 120–124. [Google Scholar] [CrossRef]
- Anderson, J.R.; Trainer, D.O.; DeFoliart, G.R. Natural and experimental transmission of the waterfowl parasite, Leucocytozoon simondi M. et L., in Wisconsin. Zoon. Res. 1962, 1, 155–164. [Google Scholar]
- Skidmore, L.V. Leucocytozoon smithi infection in turkeys and its transmission by Simulium occidentale Townsend. J. Parasitol. 1931, 18, 130. [Google Scholar]
- Byrd, M.A. Observations on Leucocytozoon in pen-raised and free-ranging wild turkeys. J. Wildl. Manag. 1959, 23, 145–156. [Google Scholar] [CrossRef]
- Fallis, A.M.; Bennett, G.F. Transmission of Leucocytozoon bonasae Clarke to ruffed grouse (Bonasa umbellus L.) by the black flies Simulium latipes Mg. and Simulium aureum Fries. Can. J. Zool. 1958, 36, 533–539. [Google Scholar] [CrossRef]
- Walker, J. A short note on the occurrence of a Leucocytozoon infection. Host: The ostrich. Trans. R. Soc. S. Afr. 1913, 3, 35–38. [Google Scholar] [CrossRef]
- Forrester, D.J.; Spalding, M.G. Parasites and Diseases of Wild Birds in Florida; University Press of Florida: Gainesville, FL, USA, 2003. [Google Scholar]
- Valkiūnas, G.; Ashford, R.W. Natural host range is not a valid taxonomic character. Trends Parasitol. 2002, 18, 528–529. [Google Scholar] [CrossRef]
- Bennett, G.F.; Earlé, R.A.; Peirce, M.A.; Huchzermeyer, F.W.; Squires-Parsons, D. Avian Leucocytozoidae: The leucocytozoids of the Phasianidae sensu lato. J. Nat. Hist. 1991, 25, 1407–1428. [Google Scholar] [CrossRef]
- Bennett, G.F.; Earlé, R.A.; Peirce, M.A. The Leucocytozoidae of South African birds: Passeriformes. Onderstepoort J. Vet. Res. 1992, 59, 235–247. [Google Scholar] [PubMed]
- Bennett, G.F.; Peirce, M.A. Leucocytozoids of seven Old World passeriform families. J. Nat. Hist. 1992, 26, 693–707. [Google Scholar] [CrossRef]
- Bennett, G.F.; Squires-Parsons, D. The leucocytozoids of the avian families Anatidae and Emberizidae s. l., with descriptions of three new Leucocytozoon species. Can. J. Zool. 1992, 70, 2007–2014. [Google Scholar] [CrossRef]
- Bennett, G.F.; Earlé, R.A.; Peirce, M.A. The Leucocytozoidae of South African birds. The Muscicapidae sensu latu. S. Afr. J. Zool. 1993, 28, 81–87. [Google Scholar]
- Oliveros, C.; Field, D.J.; Ksepka, D.T.; Barker, F.K.; Aleixo, A.; Andersen, M.J.; Alström, P.; Benz, B.W.; Braun, E.L.; Braun, M.J.; et al. Earth history and the passerine superradiation. Proc. Natl. Acad. Sci. USA 2019, 116, 7916–7925. [Google Scholar] [CrossRef]
- Kuhl, H.; Frankl-Vilches, C.; Bakker, A.; Mayr, G.; Nikolaus, G.; Boerno, S.T.; Klages, S.; Timmermann, B.; Gahr, M. An unbiased molecular approach using 3′UTRs resolves the avian family-level tree of life. Mol. Biol. Evol. 2021, 38, 108–127. [Google Scholar] [CrossRef]
- Hanel, J.; Doležalová, J.; Stehlíková, Š.; Modrý, D.; Chudoba, J.; Synek, P.; Votýpka, J. Blood parasites in northern goshawk (Accipiter gentilis) with an emphasis to Leucocytozoon toddi. Parasitol. Res. 2016, 115, 263–270. [Google Scholar] [CrossRef]
- Ramey, A.M.; Schmutz, J.A.; Reed, J.A.; Fujita, G.; Scotton, B.D.; Casler, B.; Fleskes, J.P.; Konishi, K.; Uchida, K.; Yabsley, M.J. Evidence for intercontinental parasite exchange through molecular detection and characterization of haematozoa in northern pintails (Anas acuta) sampled throughout the North Pacific Basin. Int. J. Parasitol. Parasites Wildl. 2014, 4, 11–21. [Google Scholar] [CrossRef]
- Lotta, I.A.; Pacheco, M.A.; Escalante, A.A.; González, A.; Mantilla, J.S.; Moncada, L.I. Leucocytozoon Diversity and Possible Vectors in the Neotropical highlands of Colombia. Protist 2016, 167, 185–204. [Google Scholar] [CrossRef]
- Bennett, G.F.; Campbell, A.G. Avian Leucocytozoidae. I. Morphometric variation in three species of Leucocytozoon and some taxonomic implications. Can. J. Zool. 1975, 53, 800–812. [Google Scholar] [CrossRef]
- Steele, E.J.; Noblet, G.P. Gametocytogenesis of Leucocytozoon smithi. J. Euk. Microbiol. 1993, 40, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.P.; Underhill, G.W.; Cox, J.A.; Threlkeld, W.L. A blood protozoan of turkeys transmitted by Simulium nigroparvum (Twinn). Am. J. Hyg. 1938, 27, 649–665. [Google Scholar]
- Travassos Santos Dias, J.A. Hematozoários das Aves. Moçambique 1951, 67, 71–92. [Google Scholar]
- Gill, H.; Paperna, I. Leucocytozoonosis in the Israeli sparrow, Passer domesticus biblicus Hartert 1904. Parasitol. Res. 2005, 96, 373–377. [Google Scholar] [CrossRef]
- Himmel, T.; Harl, J.; Matt, J.; Weissenböck, H. A citizen science-based survey of avian mortality focusing on haemosporidian infections in wild passerine birds. Malar. J. 2021, 20, 417. [Google Scholar] [CrossRef]
- Wong, T.C.; Desser, S.S. An ultrastructural study of the schizogonic development of Leucocytozoon dubreuili in the liver of the american robin, Turdus migratorius. Protistologica 1981, 17, 11–27. [Google Scholar]
- Ramisz, A. Protozoa of the genus Leucocytozoon Danilewsky, 1890 in birds in the environs of Wroclaw. Acta Parasitol. Pol. 1962, 10, 39–52. [Google Scholar]
- Cook, A.R. The gametocyte development of Leucocytozoon simondi. Proc. Helminthol. Soc. Wash. 1954, 21, 1–9. [Google Scholar]
- Chagas, C.R.F.; Duc, M.; Gutierrez Liberato, G.A.; Valkiūnas, G. Host cells of Leucocytozoon (Haemosporida, Leucocytozoidae) gametocytes, with remarks on phylogenetic importance of this character. Pathogens 2023, in press. [Google Scholar]
- Aikawa, M.; Huff, C.G.; Strome, C.P.A. Morphological study of microgametogenesis of Leucocytozoon simondi. J. Ultrastr. Res. 1970, 32, 43–68. [Google Scholar] [CrossRef]
- Yang, Y.J.; Desser, S.S.; Fallis, A.M. Elongate and round gametocytes of Leucocytozoon simondi (Mathis et Léger) in ducks inoculated with megaloschizonts. J. Protozool. 1971, 18, 553–556. [Google Scholar] [CrossRef] [PubMed]
- Akiba, K.; Inui, S.; Ishitani, R. Morphology and distribution of intracellular schizonts in chickens experimentally infected with Akiba caulleryi. Nat. Inst. Anim. Hlth Quart. 1971, 11, 109–121. [Google Scholar]
- Morii, T.; Nakamura, K.; Lee, Y.C.; Iijima, T.; Hoji, K. Observations on the Taiwanese strain of Leucocytozoon caulleryi (Haemosporina) in chickens. J. Protozool. 1986, 33, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Richey, D.J.; Ware, R.E. Schizonts of Leucocytozoon smithi in artificially infected turkeys. Cornell Vet. 1955, 45, 642–643. [Google Scholar]
- Newberne, J.W. The pathology of Leucocytozoon infection in turkeys with a note on its tissue stages. Am. J. Vet. Res. 1955, 16, 593–597. [Google Scholar]
- Wehr, E.E. Studies on leucocytozoonosis of turkeys, with notes on schizogony, transmission, and control of Leucocytozoon smithi. Avian Dis. 1962, 6, 195–210. [Google Scholar] [CrossRef]
- Chew, M. Megaloschizonts of Leucocytozoon in the eyes and sciatic nerves of the domestic fowl. Vet. Rec. 1968, 83, 518–519. [Google Scholar] [CrossRef]
- Omar, A.R. Haemoprotozoan infections of poultry in Malaysia. Kajian Vet. 1968, 1, 109–124. [Google Scholar]
- Kitaoka, S.; Morii, T.; Fujisaki, K. Quantitative observation on schizogony of Akiba caulleryi in chickens. Nat. Inst. Anim. Hlth. Quart. 1972, 12, 151–160. [Google Scholar]
- Peirce, M.A.; Cheke, A.S.; Cheke, R.A. A survey of blood parasites of birds in the Mascarene Islands, Indian Ocean with descriptions of two new species and taxonomic discussion. Ibis 1977, 119, 451–461. [Google Scholar] [CrossRef]
- Dinhopl, N.; Mostegl, M.M.; Richter, B.; Nedorost, N.; Maderner, A.; Fragner, K.; Weissenböck, H. Application of in-situ hybridization for the detection and identifcation of avian malaria parasites in parafn wax-embedded tissues from captive penguins. Avian Pathol. 2011, 40, 315–320. [Google Scholar] [CrossRef]
- Dinhopl, N.; Nedorost, N.; Mostegl, M.M.; Weissenbacher-Lang, C.; Weissenböck, H. In situ hybridization and sequence analysis reveal an association of Plasmodium spp. with mortalities in wild passerine birds in Austria. Parasitol. Res. 2015, 114, 1455–1462. [Google Scholar] [CrossRef] [PubMed]
- Harl, J.; Himmel, T.; Valkiūnas, G.; Weissenböck, H. The nuclear 18S ribosomal DNAs of avian haemosporidian parasites. Malar. J. 2019, 18, 305. [Google Scholar] [CrossRef] [PubMed]
- Valkiūnas, G.; Iezhova, T.A.; Shapoval, A.P. High prevalence of blood parasites in hawfnch Coccothraustes coccothraustes. J. Nat. Hist. 2003, 37, 2647–2652. [Google Scholar] [CrossRef]
- Gutiérrez, R.J. Hematozoa from the spotted owl. J. Wildl. Dis. 1989, 25, 614–618. [Google Scholar] [CrossRef]
- Galen, S.C.; Speer, K.A.; Perkins, S.L. Evolutionary lability of host associations promotes phylogenetic overdispersion of co-infecting blood parasites. J. Anim. Ecol. 2019, 88, 1936–1949. [Google Scholar] [CrossRef]
- Nooroong, P.; Watthanadirek, A.; Minsakorn, S.; Poolsawat, N.; Junsiri, W.; Srionrod, N.; Sangchuai, S.; Chawengkirttikul, R.; Anuracpreeda, P. Molecular genetic diversity and bioinformatic analysis of Leucocytozoon sabrazesi based on the mitochondrial genes cytb, coxI and coxIII and co-infection of Plasmodium spp. Parasite 2022, 29, 22. [Google Scholar] [CrossRef]
- Reis, S.; Melo, M.; Covas, R.; Doutrelant, C.; Pereira, H.; Lima, R.; Loiseau, C. Influence of land use and host species on parasite richness, prevalence and co-infection patterns. Int. J. Parasitol. 2021, 51, 83–94. [Google Scholar] [CrossRef]
- Xuan, M.N.T.; Kaewlamun, W.; Saiwichai, T.; Thanee, S.; Poofery, J.; Tiawsirisup, S.; Channumsin, M.; Kaewthamasorn, M. Development and application of a novel multiplex PCR assay for the differentiation of four haemosporidian parasites in the chicken Gallus gallus domesticus. Vet. Parasitol. 2021, 293, 109431. [Google Scholar] [CrossRef]
- Pigeault, R.; Chevalier, M.; Cozzarolo, C.S.; Baur, M.; Arlettaz, M.; Cibois, A.; Keiser, A.; Guisan, A.; Christe, P.; Glaizot, O. Determinants of haemosporidian single- and co-infection risks in western palearctic birds. Int. J. Parasitol. 2022, 52, 617–627. [Google Scholar] [CrossRef]
- Rooyen, J.V.; Lalubin, F.; Glaizot, O.; Christe, P. Altitudinal variation in haemosporidian parasite distribution in great tit populations. Parasites Vectors 2013, 6, 139. [Google Scholar] [CrossRef] [PubMed]
- Meixell, B.W.; Arnold, T.W.; Lindberg, M.S.; Smith, M.M.; Runstadler, J.A.; Ramey, A.M. Detection, prevalence, and transmission of avian hematozoa in waterfowl at the Arctic/sub-Arctic interface: Co-infections, viral interactions, and sources of variation. Parasites Vectors 2016, 9, 390. [Google Scholar] [CrossRef] [PubMed]
- Musa, S.; Mackenstedt, U.; Woog, F.; Dinkel, A. Untangling the actual infection status: Detection of avian haemosporidian parasites of three Malagasy bird species using microscopy, multiplex PCR, and nested PCR methods. Parasitol. Res. 2022, 121, 2817–2829. [Google Scholar] [CrossRef]
- Jia, T.; Huang, X.; Valkiūnas, G.; Yang, M.; Zheng, C.; Pu, T.; Zhang, Y.; Dong, L.; Suo, X.; Zhang, C. Malaria parasites and related haemosporidians cause mortality in cranes: A study on the parasites diversity, prevalence and distribution in Beijing Zoo. Malar. J. 2018, 17, 234. [Google Scholar] [CrossRef] [PubMed]
- Inumaru, M.; Nishiumi, I.; Kawakami, K.; Sato, Y. A widespread survey of avian haemosporidia in deceased wild birds of Japan: The hidden value of personally collected samples. J. Vet. Med. Sci. 2022, 84, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Ciloglu, A.; Yildirim, A.; Pekmezci, D.; Yetismis, G.; Sursal Simsek, N.; Simsek, E.; Duzlu, O.; Onder, Z.; Kokcu, N.D.; Pekmezci, G.Z.; et al. A novel one-step multiplex PCR protocol to detect avian haemosporidian parasites in the subgenus Haemoproteus (Kruse, 1890) used to quantify parasite prevalence in domestic pigeons (Columba livia) in Turkey. Vet. Res. Commun. 2022. [Google Scholar] [CrossRef]
- Ciloglu, A.; Ergen, A.G.; Inci, A.; Dik, B.; Duzlu, O.; Onder, Z.; Yetismis, G.; Bensch, S.; Valkiūnas, G.; Yildirim, A. Prevalence and genetic diversity of avian haemosporidian parasites at an intersection point of bird migration routes: Sultan Marshes National Park, Turkey. Acta Trop. 2020, 210, 105465. [Google Scholar] [CrossRef]
- Ciloglu, A.; Yildirim, A.; Duzlu, O.; Onder, Z.; Dogan, Z.; Inci, A. Investigation of avian haemosporidian parasites from raptor birds in Turkey, with molecular characterisation and microscopic confirmation. Folia Parasitol. 2016, 63, 2016.023. [Google Scholar] [CrossRef]
- Taioe, M.O.; Mnisi, M.C.; Chaisi, M.; Mwale, M.; Mabunda, N.; Phetla, V.; Thekisoe, O.M. Mosquito identification and haemosporidian parasites detection in the enclosure of the African penguins (Spheniscus demersus) at the SANBI zoological garden. Int. J. Parasitol. Parasites Wildl. 2020, 13, 98–105. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Duc, M.; Iezhova, T.A. Increase of avian Plasmodium circumflexum prevalence, but not of other malaria parasites and related haemosporidians in northern Europe during the past 40 years. Malar. J. 2022, 21, 105. [Google Scholar] [CrossRef]
- Huang, X.; Dong, L.; Zhang, C.; Zhang, Y. Genetic diversity, temporal dynamics, and host specificity in blood parasites of passerines in north China. Parasitol. Res. 2015, 114, 4513–4520. [Google Scholar] [CrossRef] [PubMed]
- Honjo, Y.; Fukumoto, S.; Sakamoto, H.; Hikosaka, K. New PCR primers targeting the cytochrome b gene reveal diversity of Leucocytozoon lineages in an individual host. Parasitol. Res. 2022, 121, 3313–3320. [Google Scholar] [CrossRef] [PubMed]
- Silva-Iturriza, A.; Ketmaier, V.; Tiedemann, R. Prevalence of avian haemosporidian parasites and their host fidelity in the central Philippine islands. Parasitol. Int. 2012, 61, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Schmid, S.; Fachet, K.; Dinkel, A.; Mackenstedt, U.; Woog, F. Carrion crows (Corvus corone) of southwest Germany: Important hosts for haemosporidian parasites. Malar. J. 2017, 16, 369. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rodríguez, A.; de la Hera, I.; Fernández-González, S.; Pérez-Tris, J. Global warming will reshuffle the areas of high prevalence and richness of three genera of avian blood parasites. Glob. Chang. Biol. 2014, 20, 2406–2416. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.F.; Peirce, M.A.; Ashford, R.W. Avian haematozoa: Mortality and pathogenicity. J. Nat. Hist. 1993, 27, 993–1001. [Google Scholar] [CrossRef]
- Duc, M.; Ilgūnas, M.; Valkiūnas, G. Patterns of Haemoproteus majoris (Haemosporida, Haemoproteidae) megalomeront development. Acta Trop. 2020, 212, 105706. [Google Scholar] [CrossRef]
- Duc, M.; Ilgūnas, M.; Kubiliūnaitė, M.; Valkiūnas, G. First Report of Haemoproteus (Haemosporida, Haemoproteidae) Megalomeronts in the Brain of an Avian Host, with Description of Megalomerogony of Haemoproteus pastoris, the Blood Parasite of the Common Starling. Animals 2021, 11, 2824. [Google Scholar] [CrossRef]
- Hellgren, O.; Waldenström, J.; Peréz-Tris, J.; Szöll, E.; Si, O.; Hasselquist, D.; Križanauskienė, A.; Ottosson, U.; Bensch, S. Detecting shifts of transmission areas in avian blood parasites: A phylogenetic approach. Mol. Ecol. 2007, 16, 1281–1290. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Kazlauskienė, R.; Bernotienė, R.; Bukauskaitė, D.; Palinauskas, V.; Iezhova, T.A. Haemoproteus infections (Haemosporida, Haemoproteidae) kill bird-biting mosquitoes. Parasitol. Res. 2014, 113, 1011–1018. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A.; Palinauskas, V.; Ilgūnas, M.; Bernotienė, R. The evidence for rapid gametocyte viability changes in the course of parasitemia in Haemoproteus parasites. Parasitol. Res. 2015, 114, 2903–2909. [Google Scholar] [CrossRef] [PubMed]
- Valkiūnas, G.; Žiegytė, R.; Palinauskas, V.; Bernotienė, R.; Bukauskaitė, D.; Ilgūnas, M.; Dimitrov, D.; Iezhova, T.A. Complete sporogony of Plasmodium relictum (lineage pGRW4) in mosquitoes Culex pipiens pipiens, with implications on avian malaria epidemiology. Parasitol. Res. 2015, 114, 3075–3085. [Google Scholar] [CrossRef] [PubMed]
- Bensch, S.; Hellgren, O. The use of molecular methods in studies of avian haemosporidians. In Avian Malaria and Related Parasites in the Tropics: Ecology, Evolution and Systematics; Santiago-Alarcon, D., Marzal, A., Eds.; Springer: Berlin, Germany, 2020; pp. 113–136. [Google Scholar]
- Nilsson, E.; Taubert, H.; Hellgren, O.; Huang, X.; Palinauskas, V.; Markovets, M.Y.; Valkiūnas, G.; Bensch, S. Multiple cryptic species of sympatric generalists within the avian blood parasite Haemoproteus majoris. J. Evol. Biol. 2016, 29, 1812–1826. [Google Scholar] [CrossRef] [PubMed]
- Freund, D.; Wheeler, S.S.; Townsend, A.K.; Boyce, W.M.; Ernest, H.B.; Cicero, C.; Sehgal, R.N.M. Genetic sequence data reveals widespread sharing of Leucocytozoon lineages in corvids. Parasitol. Res. 2016, 115, 3557–3565. [Google Scholar] [CrossRef]
- Mantilla, J.S.; González, A.D.; Lotta, I.A.; Moens, M.; Pacheco, M.A.; Escalante, A.A.; Valkiūnas, G.; Moncada, L.I.; Pérez-Tris, J.; Matta, N.E. Haemoproteus erythrogravidus n. sp. (Haemosporida, Haemoproteidae): Description and molecular characterization of a widespread blood parasite of birds in South America. Acta Trop. 2016, 159, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K. (Eds.) International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017; Koeltz Botanical Books: Glashütten, Germany, 2018; 254p. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Ashford, R.W.; Bensch, S.; Killick-Kendrick, R.; Perkins, S. A cautionary note concerning Plasmodium in apes. Trends Parasitol. 2011, 6, 231–232. [Google Scholar] [CrossRef]
- Outlaw, D.C.; Ricklefs, R.E. Species limits in avian malaria parasites (Haemosporida): How to move forward in the molecular era. Parasitology 2014, 141, 1223–1232. [Google Scholar] [CrossRef]
- O’Roke, E.C. The incidence, pathogenicity and transmission of Leucocytozoon anatis of ducks. J. Parasitol. 1930, 17, 112. [Google Scholar]
- Desser, S.S.; Yang, Y.J. Sporogony of Leucocytozoon spp. in mammalophilic simuliids. Can. J. Zool. 1973, 51, 793. [Google Scholar] [CrossRef]
- Fallis, A.M.; Bennett, G.F. Sporogony of Leucocytozoon and Haemoproteus in simuliids and ceratopogonids and a revised classification of the Haemosporidiida. Can. J. Zool. 1961, 39, 215–228. [Google Scholar] [CrossRef]
- Baker, J.R. Transmission of Leucocytozoon sakharoffi in England by Simulium angustitarse. Parasitology 1970, 60, 417–423. [Google Scholar] [CrossRef]
- Baker, J.R. Sporogony and schizogony of Leucocytozoon sakharoffi in England. J. Protozool. 1971, 18, 40. [Google Scholar]
- Fallis, A.M.; Jacobson, R.L.; Raybould, J.N. Haematozoa in domestic chickens and guinea fowl in Tanzania and transmission of Leucocytozoon neavei and L. schoutedeni. J. Protozool. 1973, 20, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Desser, S.S.; Allison, F.B. Aspects of the sporogonic development of Leucocytozoon tawaki of the fiordland crested penguin in its primary vector, Austrosimulium ungulatum: An ultrastructural study. J. Parasitol. 1979, 65, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.F. On some ornithophilic blood-sucking Diptera in Algonquin Park, Ontario, Canada. Can. J. Zool. 1960, 38, 377–389. [Google Scholar] [CrossRef]
- Barrow, J.H.; Kelker, N.; Miller, H. The transmission of Leucocytozoon simondi to birds by Simulium rugglesi in Northern Michigan. Am. Midl. Nat. 1968, 79, 197–204. [Google Scholar] [CrossRef]
- Chakarov, N.; Kampen, H.; Wiegmann, A.; Werner, D.; Bensch, S. Blood parasites in vectors reveal a united blackfly community in the upper canopy. Parasites Vectors 2020, 13, 309. [Google Scholar] [CrossRef]
- Herman, C.M.; Bennett, G.F. Use of sentinel ducks in epizootiological studies of anatid blood Protozoa. Can. J. Zool. 1976, 54, 1038–1043. [Google Scholar] [CrossRef]
- Glukhova, V.M.; Valkiūnas, G. On the fauna and ecology of biting midges (Ceratopogonidae: Culicoides) on the Curonian Spit, the methods of their collection from birds and experimental infection with haemoproteids (Haemosporidia: Haemoproteidae). Ekologija 1993, 2, 68–73. (In Russian) [Google Scholar]
- Bukauskaitė, D.; Chagas, C.R.F.; Bernotienė, R.; Žiegytė, R.; Ilgūnas, M.; Iezhova, T.; Valkiūnas, G. A new methodology for sporogony research of avian haemoproteids in laboratory-reared Culicoides spp., with a description of the complete sporogonic development of Haemoproteus pastoris. Parasites Vectors 2019, 12, 582. [Google Scholar] [CrossRef]
- Waite, J.L.; Henry, A.R.; Owen, J.P.; Clayton, D.H. An experimental test of the effects of behavioral and immunological defenses against vectors: Do they interact to protect birds from blood parasites? Parasites Vectors 2014, 7, 104. [Google Scholar] [CrossRef] [PubMed]
- Coral, A.A.; Valkiūnas, G.; González, A.D.; Matta, N.E. In vitro development of Haemoproteus columbae (Haemosporida: Haemoproteidae), with perspectives for genomic studies of avian haemosporidian parasites. Exp. Parasitol. 2015, 157, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Ziegytė, R.; Palinauskas, V.; Bernotienė, R.; Iezhova, T.A.; Valkiūnas, G. Haemoproteus minutus and Haemoproteus belopolskyi (Haemoproteidae): Complete sporogony in the biting midge Culicoides impunctatus (Ceratopogonidae), with implications on epidemiology of haemoproteosis. Exp. Parasitol. 2014, 145, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Rubtsov, I.A.; Yankovsky, A.V. Key to the Genera of Black Flies of the Palearctic; Nauka Press: Leningrad, Russia, 1984. (In Russian) [Google Scholar]
- Adler, P.H.; Currie, D.C.; Wood, D.M. The Black Flies (Simuliidae) of North America; Cornell University Press: New York, NY, USA, 2004; 941p. [Google Scholar]
- Palma, L.; Ceballos, S.J.; Johnson, P.C.; Niemeier, D.; Pitesky, M.; VanderGheynst, J.S. Cultivation of black soldier fly larvae on almond byproducts: Impacts of aeration and moisture on larvae growth and composition. J. Sci. Food Agric. 2018, 98, 5893–5900. [Google Scholar] [CrossRef]
- Edman, J.D.; Simmons, K.R. Rearing and colonization of black flies (Diptera: Simuliidae). J. Med. Entomol. 1985, 22, 1–17. [Google Scholar] [CrossRef]
- Ham, P.J.; Bianco, A.E. Maintenance of Simulium wilhelmia lineatum Meigen and Simulium erythrocephalum de Geer through successive generations in the laboratory. Can. J. Zool. 1984, 62, 870–877. [Google Scholar] [CrossRef]
- Lacoursière, J.O.; Boisvert, J.L. Short-term maintenance system for black fly larvae (Diptera: Simuliidae). J. Med. Entomol. 1987, 24, 463–466. [Google Scholar] [CrossRef]
- Bernotienė, R.; Žiegytė, R.; Vaitkutė, G.; Valkiūnas, G. Identification of a new vector species of avian haemoproteids, with a description of methodology for the determination of natural vectors of haemosporidian parasites. Parasites Vectors 2019, 12, 307. [Google Scholar] [CrossRef]
- King, R.S.; Adler, P.H. Development and evaluation of methods to assess populations of black flies (Diptera: Simuliidae) at nests of the endangered whooping crane (Grus americana). J. Vector. Ecol. 2012, 37, 298–306. [Google Scholar] [CrossRef]
- Otabil, K.B.; Gyasi, S.F.; Awuah, E.; Obeng-Ofori, D.; Rodríguez-Pérez, M.A.; Katholi, C.R.; Unnasch, T.R. The Search for an Efficient Black Fly Trap for Xenomonitoring of Onchocerciasis. J. Parasitol. Res. 2018, 2018, 5902367. [Google Scholar] [CrossRef]
- Žiegytė, R.; Bernotiene, R. Contribution to the knowledge on black flies (Diptera: Simuliidae) as vectors of Leucocytozoon (Haemosporida) parasites in Lithuania. Parasitol. Int. 2022, 87, 102515. [Google Scholar] [CrossRef] [PubMed]
- Bernotienė, R.; Valkiūnas, G. PCR detection of malaria parasites and related haemosporidians: The sensitive methodology in determining bird-biting insects. Malar. J. 2016, 15, 283. [Google Scholar] [CrossRef] [PubMed]
- Chagas, C.R.F.; Hernández-Lara, C.; Duc, M.; Valavičiūtė-Pocienė, K.; Bernotienė, R. What Can Haemosporidian Lineages Found in Culicoides Biting Midges Tell Us about Their Feeding Preferences? Diversity 2022, 14, 957. [Google Scholar] [CrossRef]
- Žiegyte, R.; Bernotienė, R.; Palinauskas, V. Culicoides segnis and Culicoides pictipennis Biting Midges (Diptera, Ceratopogonidae), New Reported Vectors of Haemoproteus Parasites. Microorganisms 2022, 10, 898. [Google Scholar] [CrossRef] [PubMed]
- Holmstad, P.R.; Anwar, A.; Iezhova, T.; Skorping, A. Standard sampling techniques underestimate prevalence of avian hematozoa in willow ptarmigan (Lagopus lagopus). J. Wildl. Dis. 2003, 39, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Mahrt, J.L.; Zwickel, F.C.; Tessier, T.G. Blood parasites of blue grouse (Dendragapus obscurus) in western North America. J. Wildl. Dis. 1991, 27, 482–485. [Google Scholar] [CrossRef]
- Shurulinkov, P.; Spasov, L.; Stoyanov, G.; Chakarov, N. Blood parasite infections in a wild population of ravens (Corvus corax) in Bulgaria. Malar. J. 2018, 17, 33. [Google Scholar] [CrossRef]
- King, J.C.; Dubay, S.A.; Huspeni, T.C.; VanLanen, A.R.; Gerhold, R.W. Parasite infections in nestling red-shouldered hawks (Buteo lineatus) in northeast Wisconsin. J. Parasitol. 2010, 96, 535–540. [Google Scholar] [CrossRef]
- Starkloff, N.C.; Kirchman, J.J.; Jones, A.W.; Winger, B.M.; Huang, Y.H.; Pulgarín-R, P.C.; Turner, W.C. Drivers of community turnover differ between avian hemoparasite genera along a North American latitudinal gradient. Ecol. Evol. 2020, 10, 5402–5415. [Google Scholar] [CrossRef]
- Morii, T.; Kitaoka, S. Influence of temperature on the sporogony of Akiba caulleryi in three Culicoides species. Nat. Inst. Anim. Hlth. Quart. 1968, 8, 210–216. [Google Scholar]
- Huchzermeyer, F.W.; Sutherland, B. Leucocytozoon smithi in South African turkeys. Avian Pathol. 1978, 7, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Glushchenko, V.V. New data on the distribution of turkey leucocytozoonosis in the territory of Ukraine. In Problems of Parasitology. Trudy Ukrainsk. Resp. Obsh. Parazitologov; 1961; Volume 1, pp. 353–355. (In Russian) [Google Scholar]
- Kairullaev, K.K.; Osipov, P.P. The finding of Leucocytozoon (Akiba) caulleryi (Haemosporidia) in hens of Kazakhstan. Parazitologiya 1981, 15, 369–370. (In Russian) [Google Scholar]
- Elbestawy, A.R.; Ellakany, H.F.; Abd El-Hamid, H.S.; Gado, A.R.; Geneedy, A.M.; Noreldin, A.E.; Menshawy, S.; El-Neweshy, M.; El-Shall, N.A.; Salaheldin, A.H. Leucocytozoon caulleryi in Broiler Chicken Flocks: Clinical, Hematologic, Histopathologic, and Molecular Detection. Avian Dis. 2021, 65, 407–413. [Google Scholar] [CrossRef] [PubMed]
- El-Azm, K.I.A.; Hamed, M.F.; Matter, A.; Rozmyslowicz, T.; Rahman, S.A.E.; Gaulton, G.N.; Bau, H.H.; El-Tholoth, M. Molecular and pathological characterization of natural co-infection of poultry farms with the recently emerged Leucocytozoon caulleryi and chicken anemia virus in Egypt. Trop. Anim. Health Prod. 2022, 54, 91. [Google Scholar] [CrossRef] [PubMed]
- Morii, T.; Kitaoka, S.; Akiba, K. Some investigations on the sporogony of Leucocytozoon caulleryi in laboratory-reared biting midges of four Culicoides species. Nat. Inst. Anim. Hlth. Quart. 1965, 5, 109–110. [Google Scholar]
- Pohuang, T.; Jittimanee, S.; Junnu, S. Pathology and molecular characterization of Leucocytozoon caulleryi from backyard chickens in Khon Kaen Province, Thailand. Vet. World 2021, 14, 2634–2639. [Google Scholar] [CrossRef]
- Lee, H.R.; Koo, B.S.; Jeon, E.O.; Han, M.S.; Min, K.C.; Lee, S.B. Pathology and molecular characterization of recent Leucocytozoon caulleryi cases in layer flocks. J. Biomed. Res. 2016, 30, 517–524. [Google Scholar] [CrossRef]
- Imura, T.; Sato, S.; Sato, Y.; Sakamoto, D.; Isobe, T.; Murata, K.; Holder, A.A.; Yukawa, M. The apicoplast genome of Leucocytozoon caulleryi, a pathogenic apicomplexan parasite of the chicken. Parasitol. Res. 2014, 113, 823–828. [Google Scholar] [CrossRef]
- Umali, D.V.; Ito, A.; Del Valle, F.P.; Shirota, K.; Katoh, H. Antibody response and protective immunity of chickens vaccinated with booster dose of recombinant oil-adjuvanted Leucocytozoon caulleryi subunit vaccine. Avian Dis. 2014, 58, 609–615. [Google Scholar] [CrossRef]
- Ito, A.; Gotanda, T. Field efficacy of recombinant R7 vaccine against chicken leucocytozoonosis. J. Vet. Med. Sci. 2004, 66, 483–487. [Google Scholar] [CrossRef]
- Morii, T.; Fujita, J.; Akiba, K.; Isobe, T.; Nakamoto, K.; Ishihara, H. Protective immunity to Leucocytozoon caulleryi in chickens by a killed vaccine. Parasitol. Res. 1990, 76, 630–632. [Google Scholar] [CrossRef]
- Gratz, N.G.; Steffen, R.; Cocksedge, W. Why aircraft disinsection? Bull. World Health Organ. 2000, 78, 995–1004. [Google Scholar] [PubMed]
- Alenou, L.D.; Etang, J. Airport Malaria in Non-Endemic Areas: New Insights into Mosquito Vectors, Case Management and Major Challenges. Microorganisms 2021, 9, 2160. [Google Scholar] [CrossRef]
- Pondorfer, S.G.; Jaeger, V.K.; Scholz-Kreisel, P.; Horn, J.; Krumkamp, R.; Kreuels, B.; Mikolajczyk, R.T.; Karch, A. Risk estimation for air travel-induced malaria transmission in central Europe—A mathematical modelling study. Travel. Med. Infect. Dis. 2020, 36, 101564. [Google Scholar] [CrossRef]
- Ricklefs, R.E.; Medeiros, M.; Ellis, V.A.; Svensson-Coelho, M.; Blake, J.G.; Loiselle, B.A.; Soares, L.; Fecchio, A.; Outlaw, D.; Marra, P.P.; et al. Avian migration and the distribution of malaria parasites in New World passerine birds. J. Biogeogr. 2017, 44, 1113–1123. [Google Scholar] [CrossRef]
- White, E.M.; Greiner, E.C.; Bennett, G.F.; Herman, C.M. Distribution of the hematozoa of Neotropical birds. Rev. Biol. Trop. 1978, 26, 43–102. [Google Scholar] [PubMed]
- Fecchio, A.; Silveira, P.; Weckstein, J.D.; Dispoto, J.H.; Anciães, M.; Boshln, M.; Tkach, V.V.; Bell, J.A. First Record of Leucocytozoon (Haemosporida: Leucocytozoidae) in Amazonia: Evidence for Rarity in Neotropical Lowlands or Lack of Sampling for This Parasite Genus? J. Parasitol. 2018, 104, 168–172. [Google Scholar] [CrossRef] [PubMed]
- González-Olvera, M.; Hernandez-Colina, A.; Pérez, J.G.; Ulloa, G.M.; Montero, S.; Maguiña, J.L.; Lescano, A.G.; Santolalla, M.L.; Baylis, M.; Mayor, P. Haemosporidians from a Neglected Group of Terrestrial Wild Birds in the Peruvian Amazonia. Ecohealth 2022, 19, 402–416. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.M.; Ferreira, F.C.; Araújo, A.C.; Hirano, L.Q.L.; Paludo, G.R.; Braga, É.M. Molecular detection of Leucocytozoon in red-legged seriemas (Cariama cristata), a non-migratory bird species in the Brazilian Cerrado. Vet. Parasitol. Reg. Stud. Rep. 2022, 31, 100652. [Google Scholar] [CrossRef] [PubMed]
- Chagas, C.R.; Guimarães, L.O.; Monteiro, E.F.; Valkiūnas, G.; Katayama, M.V.; Santos, S.V.; Guida, F.J.V.; Simões, R.F.; Kirchgatter, K. Hemosporidian parasites of free-living birds in the São Paulo Zoo, Brazil. Parasitol. Res. 2016, 115, 1443–1452. [Google Scholar] [CrossRef] [PubMed]
- Adler, P.H. World Blackflies (Diptera: Simuliidae): A Comprehensive Revision of the Taxonomic and Geographical Inventory. 2022, pp. 1–145. Available online: https://biomia.sites.clemson.edu/pdfs/blackflyinventory.pdf (accessed on 20 April 2023).
- Forrester, D.J.; Foster, G.W.; Morrison, J.L. Leucocytozoon toddi and Haemoproteus tinnunculi (Protozoa: Haemosporina) in the Chimango caracara (Milvago chimango) in southern Chile. Mem. Inst. Oswaldo Cruz. 2001, 97, 1023–1024. [Google Scholar] [CrossRef] [PubMed]
- Peirce, M.A.; Greenwood, A.G.; Swinnerton, K. Pathogenicity of Leucocytozoon marchouxi in the pink pigeon (Columba mayeri) in Mauritius. Vet. Rec. 1997, 140, 155–156. [Google Scholar] [CrossRef] [PubMed]
- Galosi, L.; Scaglione, F.E.; Magi, G.E.; Cork, S.C.; Peirce, M.A.; Ferraro, S.; Cucuzza, L.S.; Cannizzo, F.T.; Rossi, G. Fatal Leucocytozoon Infection in a Captive Grey-headed Parrot (Poicephalus robustus suahelicus). J. Avian Med. Surg. 2019, 33, 179–183. [Google Scholar] [CrossRef]
- Davies, D.M. Longevity of black flies in captivity. Can. J. Zool. 1953, 31, 304–312. [Google Scholar] [CrossRef]
- Bukauskaitė, D.; Bernotienė, R.; Iezhova, T.A.; Valkiūnas, G. Mechanisms of mortality in Culicoides biting midges due to Haemoproteus infection. Parasitology 2016, 143, 1748–1754. [Google Scholar] [CrossRef] [PubMed]
- Chong, D.L.A.; McHale, B.; Garrett, K.B.; Yabsley, M.J. Fatal Systemic Haemosporidiosis in a Free-ranging Greater Sage-grouse (Centrocercus urophasianus). J. Wildl. Dis. 2023, 59, 207–211. [Google Scholar] [CrossRef]
- Martín-Maldonado, B.; Mencía-Gutiérrez, A.; Andreu-Vázquez, C.; Fernández, R.; Pastor-Tiburón, N.; Alvarado, A.; Carrero, A.; Fernández-Novo, A.; Esperón, F.; González, F. A Four-Year Survey of Hemoparasites from Nocturnal Raptors (Strigiformes) Confirms a Relation between Leucocytozoon and Low Hematocrit and Body Condition Scores of Parasitized Birds. Vet. Sci. 2023, 10, 54. [Google Scholar] [CrossRef]
- Wobeser, G.A. Diseases of Wild Waterfowl, 2nd ed.; Plenum Press: New York, NY, USA, 1997; 324p. [Google Scholar]
- Wickware, A.B. Is Leucocytozoon anatis the cause of a new disease in ducks? Parasitology 1915, 8, 17–21. [Google Scholar] [CrossRef]
- O’Roke, E.C. A malaria-like disease of ducks caused by Leucocytozoon anatis Wickware. Univ. Mich. Sch. Cons. Bull. 1934, 4, 1–44. [Google Scholar]
- Ivanić, M. Zur Kenntnis der gewöhnlichen Zweiteilung und der in den Leberzellen der Hausgans (Anser domesticus L.) vorkommenden multiplen Teilung (Schizogonie) bei Leucocytozoon anseris Knuth u. Magdeburg. Arch. Protistenkd. 1937, 89, 16–44. [Google Scholar]
- Fallis, A.M.; Davies, D.M.; Vickers, M.A. Life history of Leucocytozoon simondi Mathis and Léger in natural and experimental infections and blood changes produced in the avian host. Can. J. Zool. 1951, 29, 305–328. [Google Scholar] [CrossRef]
- Cowan, A.B. Reactions against the megaloschizonts of Leucocytozoon simondi Mathis and Léger in ducks. J. Infect. Dis. 1957, 100, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Kocan, R.M.; Clark, D.T. Anemia in ducks infected with Leucocytozoon simondi. J. Protozool. 1966, 13, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Novilla, M.N.; Masangkay, J.S.; Santiago, B.A. Pathology of chicken naturally infected with Leucocytozoon caulleryi. Philipp. J. Vet. Med. 1971, 10, 130–138. [Google Scholar]
- Jones, J.E.; Barnett, B.D.; Solis, J. The effect of Leucocytozoon smithi infection on production, fertility, and hatchability of broad breasted white turkey hens. Poult. Sci. 1972, 51, 1543–1545. [Google Scholar] [CrossRef]
- Commichau, C.; Jonas, D. Eine durch Leukozytozoon simondi verursachte Erkrankung bei Entenküken unter besonderer Berücksichtigung des histologischen Nachweises. Zbl. Vet. Med. B 1977, 24, 662–667. [Google Scholar] [CrossRef]
- Oosthuizen, J.H.; Markus, M.B. The hematozoa of South African birds. IV. Leucocytozoonosis in nestlings. J. Parasitol. 1968, 54, 1244–1246. [Google Scholar] [CrossRef]
- Peirce, M.A. Haematozoa of Zambian birds. I. General survey. J. Nat. Hist. 1984, 18, 105–122. [Google Scholar] [CrossRef]
- Raida, S.R.; Jaensch, S.M. Central nervous disease and blindness in Nankeen kestrels (Falco cenchroides) due to a novel Leucocytozoon-like infection. Avian Pathol. 2000, 29, 51–56. [Google Scholar] [CrossRef]
- Tarello, W. Leucocytozoon toddi in falcons from Kuwait: Epidemiology, clinical signs and response to melarsomine. Parasite 2006, 13, 179. [Google Scholar]
- Jansson, D.S.; Otman, F.; Bagge, E.; Lindgren, Y.; Etterlin, P.E.; Eriksson, H. Retrospective analysis of post-mortem findings in domestic ducks and geese from non-commercial flocks in Sweden, 2011–2020. Acta Vet. Scand. 2021, 63, 47. [Google Scholar] [CrossRef]
- Siccardi, F.J.; Derieux, W.T.; Rutherford, H.O. Pathology and prevention of Leucocytozoon smithi infection in turkeys. J. Am. Vet. Med. Assoc. 1971, 158, 1901–1902. [Google Scholar] [CrossRef]
- Lee, Y.C.; Chang, C.H.; Fu, T.H. Studies on the leucocytozoonosis of chickens. V. Symptoms and damages caused by the infection of Leucocytozoon sp. Mem. Coll. Agric. 1969, 10, 16–23. [Google Scholar]
- Newberne, J.W. Studies on the histopathology of Leucocytozoon simondi infection. Am. J. Vet. Res. 1957, 18, 191–199. [Google Scholar] [PubMed]
- Miller, R.E.; Trampel, D.W.; Desser, S.S.; Boever, W.J. Leucocytozoon simondi infection in European and American eiders. J. Am. Vet. Med. Assoc. 1983, 183, 1241–1244. [Google Scholar]
- Peirce, M.A.; Lederer, R.; Adlard, R.D.; O’Donoghue, P.J. Pathology associated with endogenous development of haematozoa in birds from southeast Queensland. Avian Pathol. 2004, 33, 445–450. [Google Scholar] [CrossRef]
- Kocan, R.M. Anemia and mechanism of erythrocyte destruction in ducks with acute Leucocytozoon infections. J. Protozool. 1968, 15, 455–462. [Google Scholar] [CrossRef]
- Maley, G.J.M.; Desser, S.S. Anemia in Leucocytozoon simondi infections. I. Quantification of anemia, gametocytemia, and osmotic fragility of erythrocytes in naturally infected Pekin ducklings. Can. J. Zool. 1977, 55, 355–358. [Google Scholar] [CrossRef]
- Siccardi, F.J.; Rutherford, H.O.; Derieux, W.T. Pathology and prevention of Leucocytozoon smithi infection of turkeys. Avian Dis. 1974, 18, 21–32. [Google Scholar] [CrossRef]
- Peirce, M.A.; Greenwood, A.G.; Stidworthy, M.F. Leucocytozoon in captive penguins. Vet. Rec. 2005, 157, 819–820. [Google Scholar] [CrossRef]
- Hunter, D.B.; Rohner, C.; Currie, D.C. Mortality in fledgling great horned owls from black fly hematophaga and leucocytozoonosis. J. Wildl. Dis. 1997, 33, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Niedringhaus, K.D.; Fenton, H.M.A.; Cleveland, C.A.; Anderson, A.N.; Schwartz, D.; Alex, C.E.; Rogers, K.H.; Mete, A.; Yabsley, M.J. Case Series: Virulent hemosporidiosis infections in juvenile great horned owls (Bubo virginianus) from Louisiana and California, USA. Vet. Parasitol. Reg. Stud. Rep. 2018, 12, 49–54. [Google Scholar] [CrossRef]
- Khan, R.A.; Fallis, A.M. Comparison of infections with Leucocytozoon simondi in black ducks (Anas rubripes), mallards (Anas platyrhynchos), and white Pekins (Anas boschas). Can. J. Zool. 1968, 46, 773–780. [Google Scholar] [CrossRef]
- Mörner, T.; Wahlström, K. Infektion med bloodparasiten Leucocytozoon simondi—En vanlig dödsorsak hos knölsvanungar Cygnus olor. Vår. Fågelvarld. 1983, 42, 389–394. [Google Scholar]
- Karstad, L. A case of leucocytozoonosis in a wild mallard. Bull. Wildl. Dis. Ass. 1965, 1, 33–34. [Google Scholar] [CrossRef]
- Herman, C.M.; Barrow, J.H.; Tarshis, I.B. Leucocytozoonosis in Canada geese at the Seney National Wildlife Refuge. J. Wildl. Dis. 1975, 11, 404–411. [Google Scholar] [CrossRef]
- Korpimäki, E.; Hakkarainen, H.; Bennett, G.F. Blood parasites and reproductive success of Tengmalm’s owls: Detrimental effects on females but not on males? Funct. Ecol. 1993, 7, 420–426. [Google Scholar] [CrossRef]
- Norris, K.; Anwar, M.; Read, A. Reproductive effort influences the prevalence of haematozoan parasites in great tits. J. Anim. Ecol. 1994, 63, 601–610. [Google Scholar] [CrossRef]
- Shutler, D.; Lowe, A.G.; Robinson, S.R. Relationships between circulating leucocytes and Leucocytozoon simondi in mallard, Anas platyrhynchos, ducklings. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 156, 46–49. [Google Scholar] [CrossRef]
- Møller, A.P.; Nielsen, J.T. Malaria and risk of predation: A comparative study of birds. Ecology 2007, 88, 871–881. [Google Scholar] [CrossRef]
- Trefiak, W.D.; Desser, S.S. Crystalloid inclusions in species of Leucocytozoon, Parahaemoproteus, and Plasmodium. J. Protozool. 1973, 20, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Terzakis, J.A.; Vanderberg, J.P.; Weiss, M.M. Viruslike particles in malaria parasites. J. Parasitol. 1976, 62, 366–371. [Google Scholar] [CrossRef]
- Charon, J.; Grigg, M.J.; Eden, J.-S.; Piera, K.A.; Rana, H.; William, T.; Rose, K.; Davenport, M.P.; Anstey, N.M.; Holmes, E.C. Novel RNA viruses associated with Plasmodium vivax in human malaria and Leucocytozoon parasites in avian disease. PLoS Pathog. 2019, 15, e1008216. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.R.; Roy, S.W.; Sehgal, R.N.M. Novel RNA viruses associated with avian haemosporidian parasites. PLoS ONE 2022, 17, e0269881. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Arreaza, A.; Haenni, A.-L.; Dunia, I.; Avilán, L. Viruses of parasites as actors in the parasite-host relationship: A “ménage à trois”. Acta Trop. 2017, 166, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Fujisaki, K.; Takamatsu, H.; Kitaoka, S.; Suzuki, K.; Kamio, T. Direct immunoflourescent staining of Leucocytozoon caulleryi of different developmental stages. Nat. Inst. Anim. Hlth. Quart. 1981, 21, 73–79. [Google Scholar]
- Isobe, T.; Shimizu, S.Y.; Yoshihara, S.; Suzuki, K. Immunoblot analysis of humoral responses to Leucocytozoon caulleryi in chickens. J. Parasitol. 1998, 84, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Ito, A.; Gotanda, T. A rapid assay for detecting antibody against leucocytozoonosis in chickens with a latex agglutination test using recombinant R7 antigen. Avian Pathol. 2005, 34, 15–19. [Google Scholar] [CrossRef]
- Horosko, S.; Noblet, R. Black fly control and suppression of leucocytozoonosis in turkeys. J. Agric. Entomol. 1968, 3, 10–24. [Google Scholar]





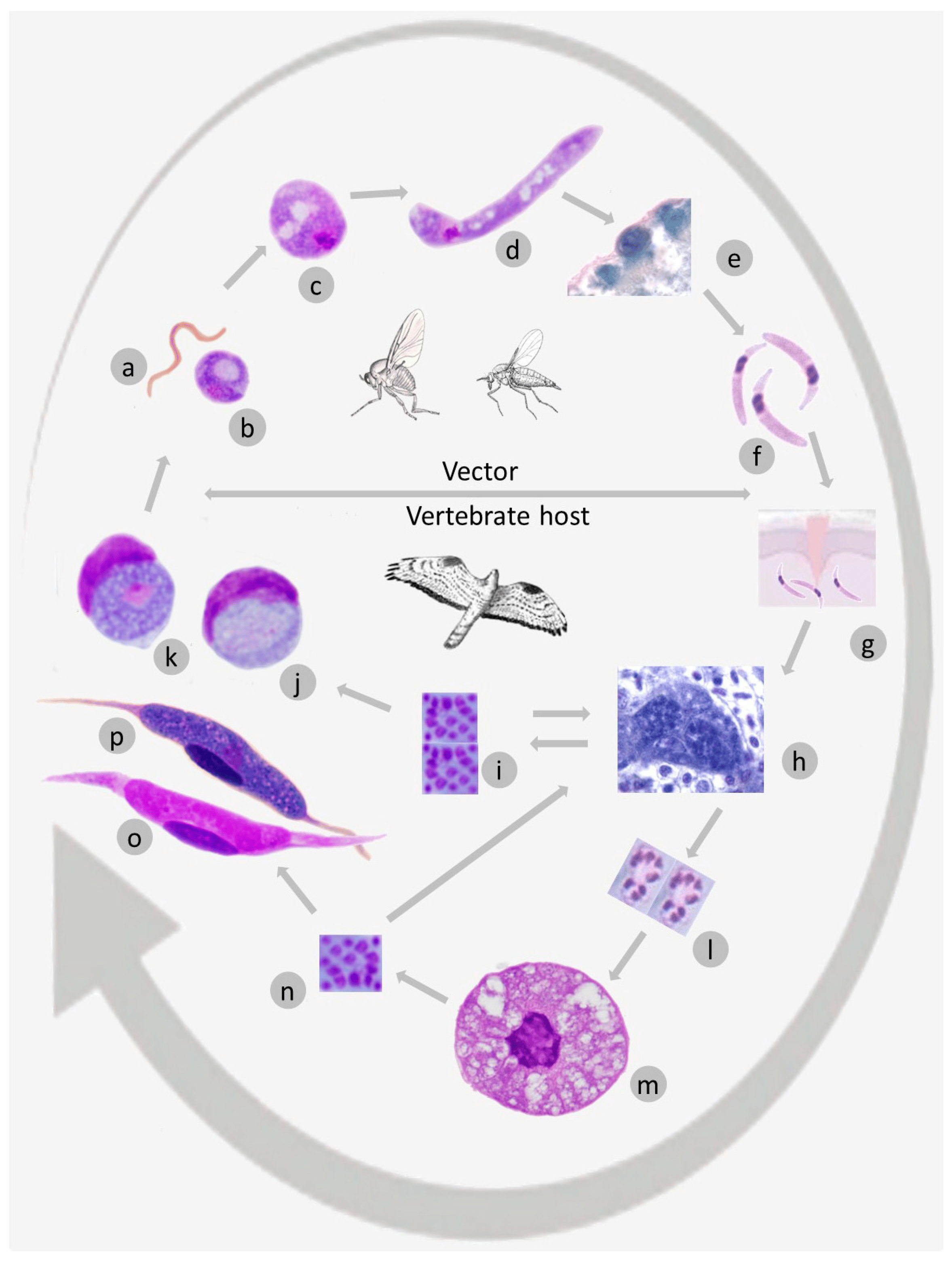{kind=link}
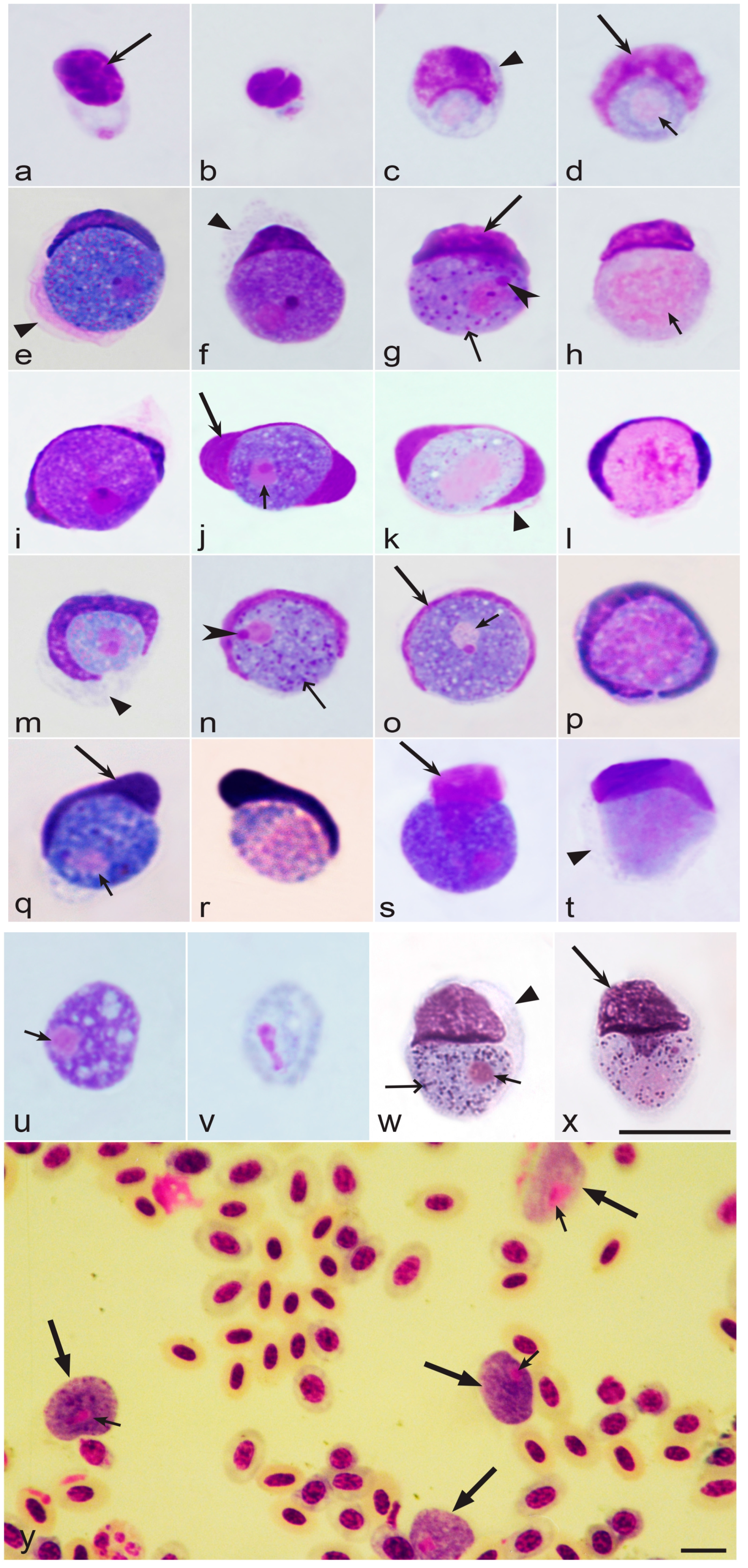{kind=link}
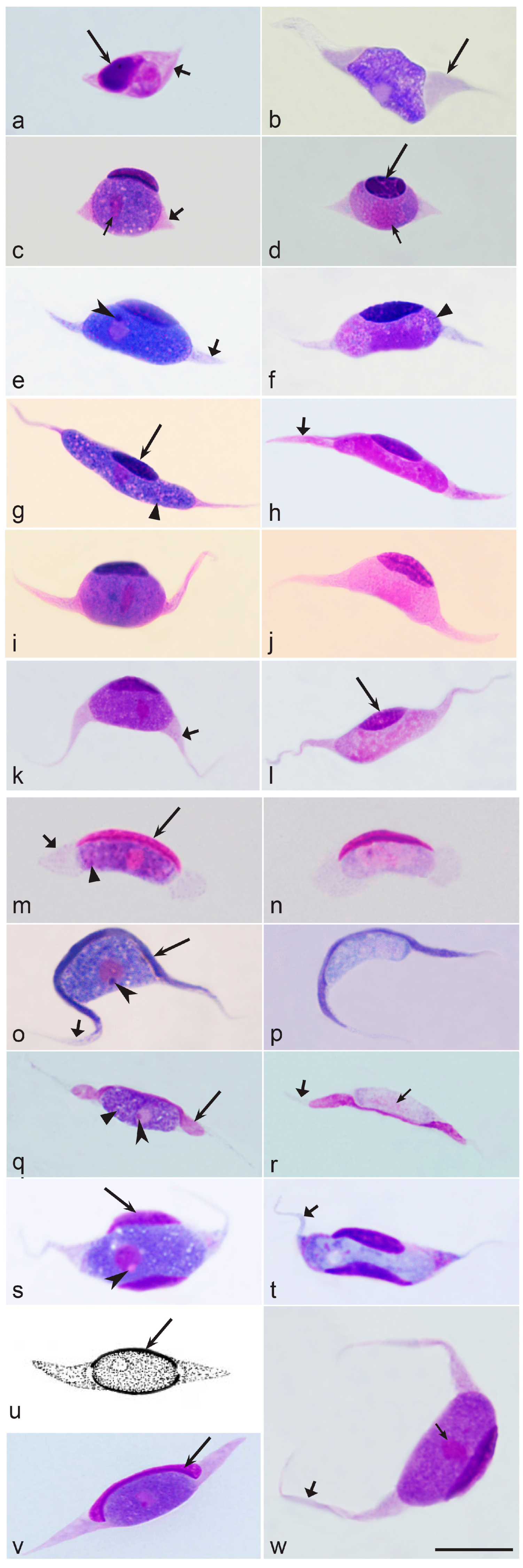{kind=link}
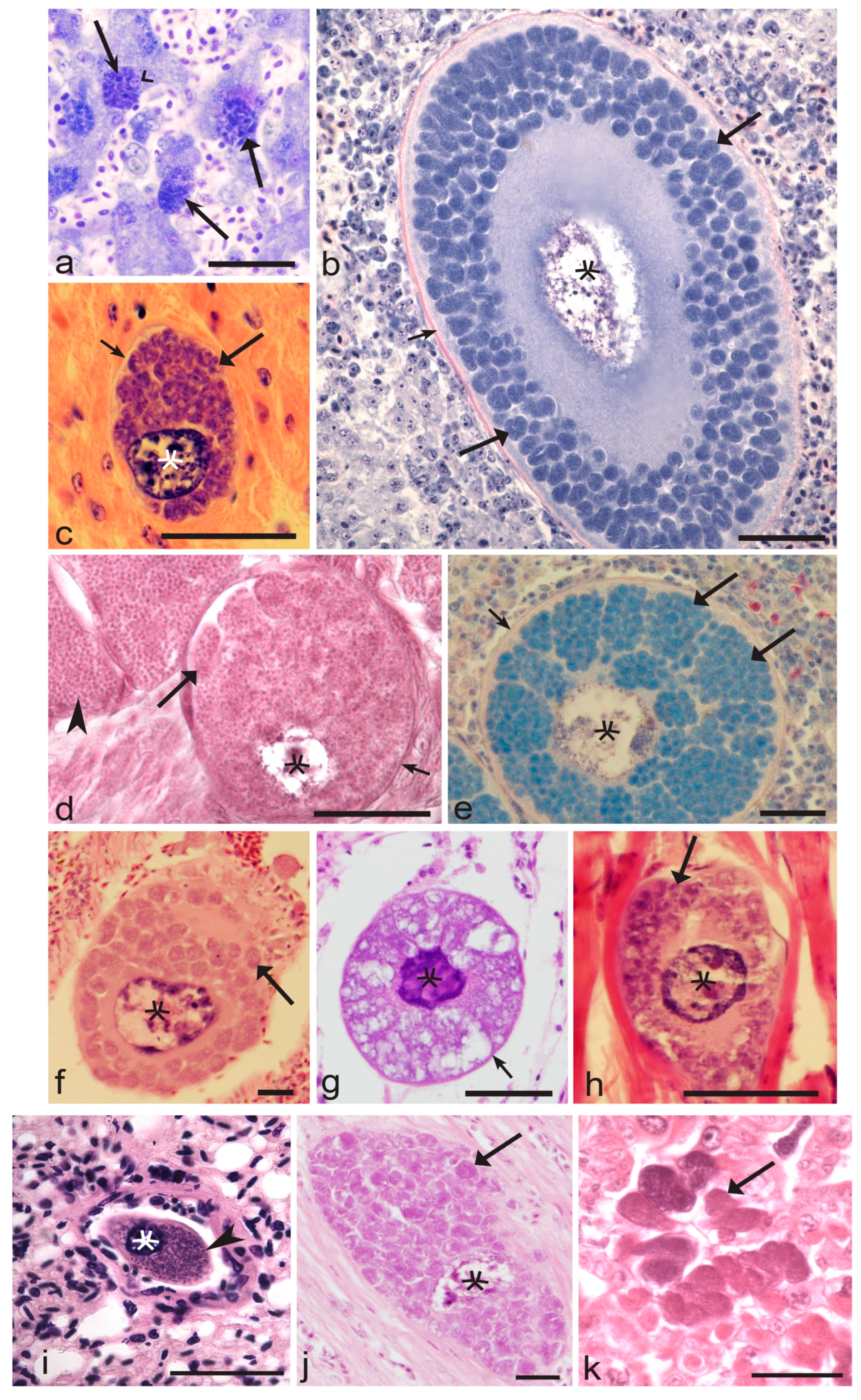{kind=link}
| Parasite Species 1 | GenBank Accession and Lineage Code (in Parentheses) 2 | Reference 3 |
|---|---|---|
| L. balmorali (R, F) | Not available | [4,122] |
| L. bennetti (R) | Not available | [4,123] |
| L. berestneffi (R) | Not available | [4,64,85,90] |
| L. buteonis (F) 4 | DQ177253 (BUBT2), DQ177273 (BUTJAM10), DQ177264 (BUTREG01) | [4,124,125] |
| L. californicus (R) | FASPA02 (KR422359) | [20] |
| L. caprimulgi (R, F) | Not available | [4,126] |
| L. caulleryi (R) | GALLUS05 (AB302215), complete mitochondrial DNA genome | [4,127] |
| L. centropi (R) | Not available | [4,128,129] |
| L. cheissini (F) | Not available | [4,48] |
| L. colius (R) | Not available | [4,130] |
| L. communis (R) | Not available | [4,131] |
| L. danilewskyi (R, F) 5 | Not available | [4,124,132] |
| L. dizini (R) | Not available | [4,133] |
| L. dubreuili (R) 6 | TUMER09 (KY653795) | [4,90,134,135] |
| L. eurystomi (R, R) | Not available | [4,126] |
| L. fringillinarum (R) 7 | Not available | [4,136] |
| L. grallariae (F) | GRSQU01 (MK103895), partial mitochondrial DNA genome | [9] |
| L. grusi (R, F) | Not available | [4,137] |
| L. hamiltoni (F) | Not available | [4,138] |
| L. leboeufi (R) | Not available | [4,139] |
| L. legeri (R) | Not available | [4,140] |
| L. lovati (R, F) 8 | LAMUT01 (AB183550) | [4,56,141,142] |
| L. maccluri (R, F) | Not available | [4,143] |
| L. macleani (R, F) 9 | Not available | [4,30,64,144] |
| L. majoris (R) 10 | Not available | [4,145] |
| L. marchouxi (R) | Not available | [4,146,147] |
| L. mathisi (F) 4 | ACNI04 (DQ177252), ACCOP01 (DQ177250) | [125,140] |
| L. neavei (F) | Not available | [4,126,148] |
| L. neotropicalis (F) | PIRIE01 (MK103894), partial mitochondrial DNA genome | [9] |
| L. nycticoraxi (R) | Not available | [4,149] |
| L. nyctyornis (R) | Not available | [4,150] |
| L. podargii (R) | Isolate WA (MK358451) | [151,152] |
| L. polynuclearis (R) | COLAUR01 (MW626894), DRYALB01 (MW626892) | [21] |
| L. pterotenuis (F) 11 | (KM610046) partial mitochondrial DNA genome, GRARUF01 (KM272250) | [9,153] |
| L. quynzae (R) | HEAME01 (KF479480) partial mitochondrial DNA genome, HELIAM01 (KF309188) | [18] |
| L. sakharoffi (R) | Not available | [4,64,90] |
| L. schoutedeni (R) | GALLUS06 (DQ676823), GALLUS07 (DQ676824) | [4,29,154] |
| L. simondi (R, F) 12 | Not available | [4,146,155,156] |
| L. smithi (R, F) | Not available | [4,157] |
| L. sousadiasi (F) | Not available | [4,133] |
| L. squamatus (R) | Not available | [4,158] |
| L. struthionis (R) | Not available | [4,159] |
| L. tawaki (R) | Not available | [4,160] |
| L. toddi (F, occationally R) 13 | Not available | [4,64,161] |
| L. vandenbrandeni (R) | Not available | [4,162] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valkiūnas, G.; Iezhova, T.A. Insights into the Biology of Leucocytozoon Species (Haemosporida, Leucocytozoidae): Why Is There Slow Research Progress on Agents of Leucocytozoonosis? Microorganisms 2023, 11, 1251. https://doi.org/10.3390/microorganisms11051251
Valkiūnas G, Iezhova TA. Insights into the Biology of Leucocytozoon Species (Haemosporida, Leucocytozoidae): Why Is There Slow Research Progress on Agents of Leucocytozoonosis? Microorganisms. 2023; 11(5):1251. https://doi.org/10.3390/microorganisms11051251
Chicago/Turabian StyleValkiūnas, Gediminas, and Tatjana A. Iezhova. 2023. "Insights into the Biology of Leucocytozoon Species (Haemosporida, Leucocytozoidae): Why Is There Slow Research Progress on Agents of Leucocytozoonosis?" Microorganisms 11, no. 5: 1251. https://doi.org/10.3390/microorganisms11051251