

Polyhydroxybutyrate Production from Methane and Carbon Dioxide by a Syntrophic Consortium of Methanotrophs with Oxygenic Photogranules without an External Oxygen Supply
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Culture Conditions
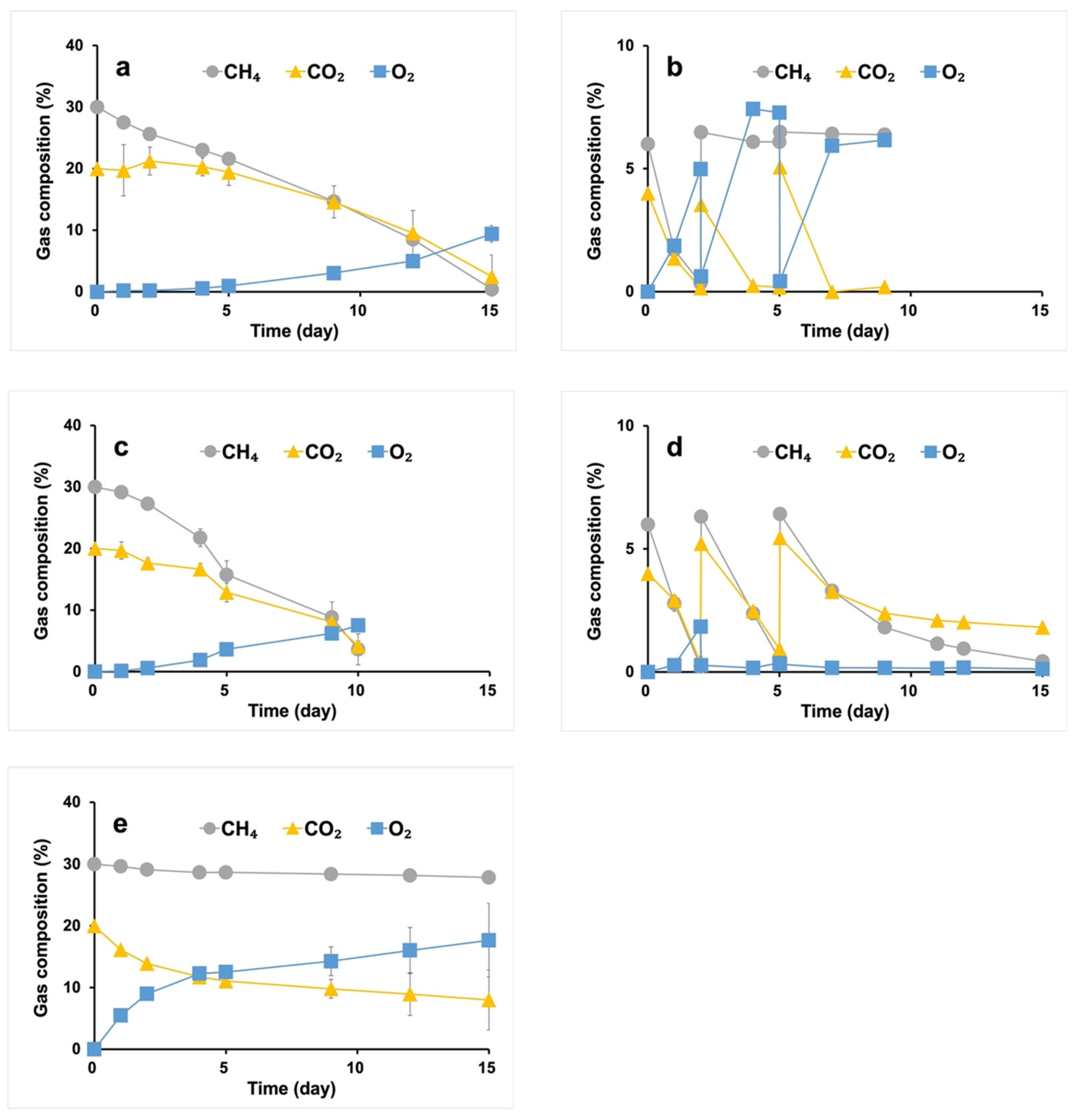
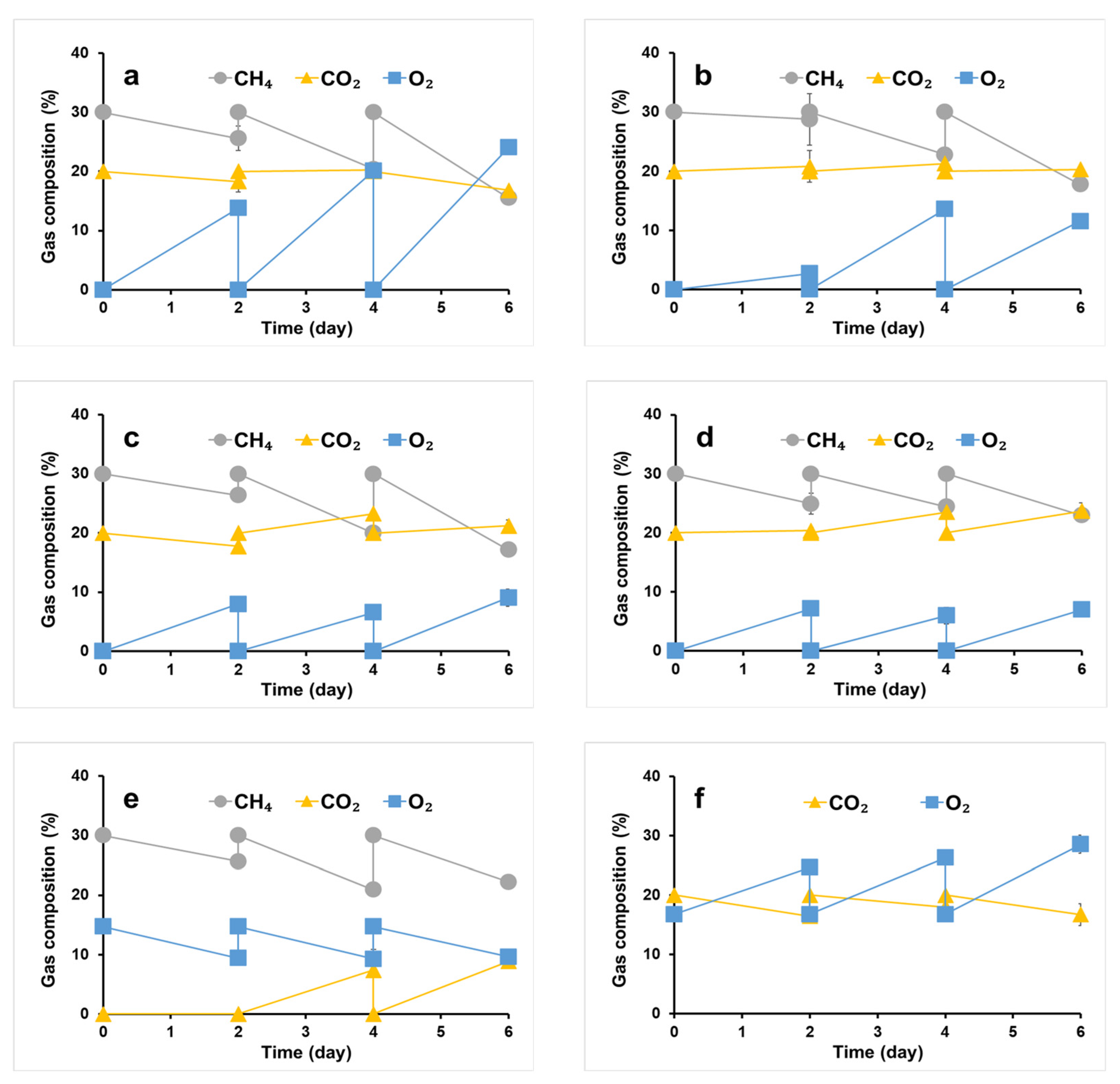
2.2. Gas Consumption of the Syntrophic Consortiums
2.3. 16S rRNA Amplicon Sequencing
2.4. Flask Cultivation of Syntrophic Consortium for PHB Production Using CH4 and CO2
2.5. Analytical Methods
3. Results and Discussion
3.1. Gas Consumption by the Syntrophic Consortiums under Different CH4 Contents
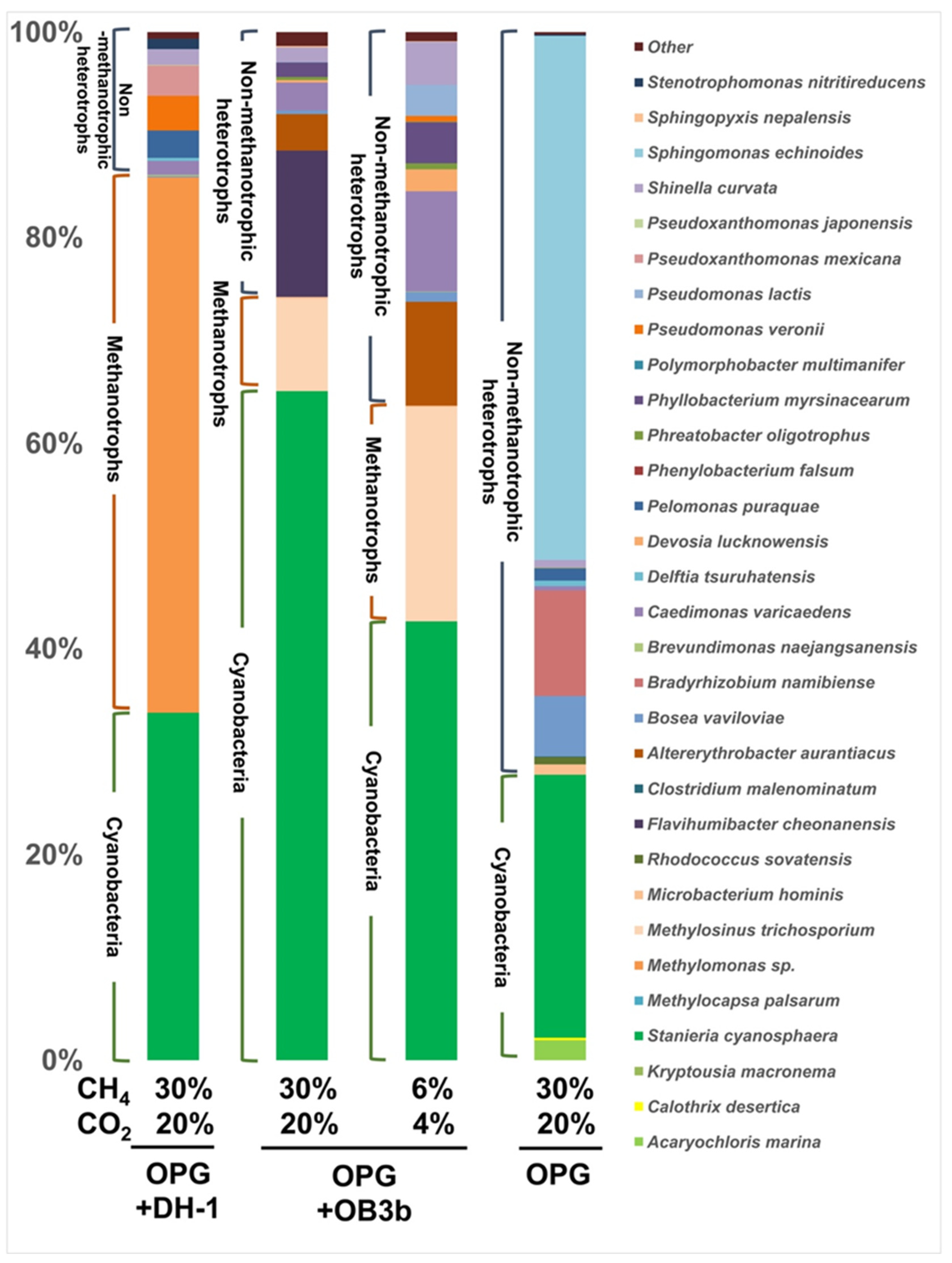
3.2. Metagenome Sequencing of the Syntrophic Consortiums after Cultivation with Different Gas Ratios
3.3. Improvement of PHB Accumulation by Adjusting Nitrogen Source Concentrations
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guglielmi, G. Methane leaks from US gas fields dwarf government estimates. Nature 2018, 558, 496–497. [Google Scholar] [CrossRef] [PubMed]
- Pathak, M.; Slade, R.; Pichs-Madruga, R.; Ürge-Vorsatz, D.; Shukla, P.R.; Skea, J. Climate Change 2022: Mitigation of Climate Change; Technical Summary; Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022. [Google Scholar]
- Strong, P.J.; Kalyuzhnaya, M.; Silverman, J.; Clarke, W.P. A methanotroph-based biorefinery: Potential scenarios for generating multiple products from a single fermentation. Bioresour. Technol. 2016, 215, 314–323. [Google Scholar] [CrossRef]
- Mackie, J.C. Partial oxidation of methane: The role of the gas phase reactions. Catal. Rev. 1991, 33, 169–240. [Google Scholar] [CrossRef]
- Hanson, R.S.; Hanson, T.E. Methanotrophic bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.Y.; Rhie, M.N.; Jung, S.M.; Sohn, Y.J.; Yeon, Y.J.; Kim, M.S.; Park, C.; Lee, J.; Park, S.J.; Na, J.G. Hydrogen production from methane by Methylomonas sp. DH-1 under micro-aerobic conditions. Biotechnol. Bioprocess Eng. 2020, 25, 71–77. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Hwang, I.Y.; Na, J.G.; Lee, E.Y. Biological conversion of propane to 2-propanol using group Ⅰ and Ⅱ methanotrophs as biocatalysts. J. Ind. Microbiol. Biotechnol. 2019, 46, 675–685. [Google Scholar] [CrossRef]
- Patel, S.K.S.; Kalia, V.C.; Joo, J.B.; Kang, Y.C.; Lee, J.K. Biotransformation of methane into methanol by methanotrophs immobilized on coconut coir. Bioresour. Technol. 2020, 297, 122433. [Google Scholar] [CrossRef]
- Nguyen, D.T.N.; Lee, O.K.; Lim, C.; Lee, J.; Na, J.G.; Lee, E.Y. Metabolic engineering of type II methanotroph, Methylosinus trichosporium OB3b, for production of 3-hydroxypropionic acid from methane via a malonyl-CoA reductase-dependent pathway. Metab. Eng. 2020, 59, 142–150. [Google Scholar] [CrossRef]
- Carmona-Martínez, A.A.; Marcos-Rodrigo, E.; Bordel, S.; Marín, D.; Herrero-Lobo, R.; García-Encina, P.A.; Muñoz, R. Elucidating the key environmental parameters during the production of ectoines from biogas by mixed methanotrophic consortia. J. Environ. Manag. 2021, 298, 113462. [Google Scholar] [CrossRef]
- Cordova-Gonzalez, A.; Birgel, D.; Kappler, A.; Peckmann, J. Variation of salinity and nitrogen concentration affects the pentacyclic triterpenoid inventory of the haloalkaliphilic aerobic methanotrophic bacterium Methylotuvimicrobium alcaliphilum. Extremophiles 2021, 25, 285–299. [Google Scholar] [CrossRef]
- Tsapekos, P.; Zhu, X.; Pallis, E.; Angelidaki, I. Proteinaceous methanotrophs for feed additive using biowaste as carbon and nutrients source. Bioresour. Technol. 2020, 313, 123646. [Google Scholar] [CrossRef] [PubMed]
- Valverde-Pérez, B.; Xing, W.; Zachariae, A.A.; Skadborg, M.M.; Kjeldgaard, A.F.; Palomo, A.; Smets, B.F. Cultivation of methanotrophic bacteria in a novel bubble-free membrane bioreactor for microbial protein production. Bioresour. Technol. 2020, 310, 123388. [Google Scholar] [CrossRef]
- Cantera, S.; Muñoz, R.; Lebrero, R.; López, J.C.; Rodríguez, Y.; García-Encina, P.A. Technologies for the bioconversion of methane into more valuable products. Curr. Opin. Biotechnol. 2018, 50, 128–135. [Google Scholar] [CrossRef] [PubMed]
- García-Pérez, T.; López, J.C.; Passos, F.; Lebrero, R.; Revah, S.; Muñoz, R. Simultaneous methane abatement and PHB production by Methylocystis hirsuta in a novel gas-recycling bubble column bioreactor. Chem. Eng. J. 2018, 334, 691–697. [Google Scholar] [CrossRef]
- Rodríguez, Y.; Firmino, P.I.M.; Pérez, V.; Lebrero, R.; Muñoz, R. Biogas valorization via continuous polyhydroxybutyrate production by Methylocystis hirsuta in a bubble column bioreactor. Waste Manag. 2020, 113, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, O.P.; Chidambarampadmavathy, K.; Cirés, S.; Heimann, K. Review of sustainable methane mitigation and bio-polymer production. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1579–1610. [Google Scholar] [CrossRef]
- Singh, R.; Ryu, J.; Kim, S.W. An overview on methanotrophs and the role of Methylosinus trichosporium OB3b for biotechnological applications. Biotechnol. Bioprocess Eng. 2022, 27, 468–481. [Google Scholar] [CrossRef]
- Pieja, A.J.; Sundstrom, E.R.; Criddle, C.S. Cyclic, alternating methane and nitrogen limitation increases PHB production in a methanotrophic community. Bioresour. Technol. 2012, 107, 385–392. [Google Scholar] [CrossRef]
- Hecke, W.V.; Bockrath, R.; Wever, H.D. Effects of moderately elevated pressure on gas fermentation processes. Bioresour. Technol. 2019, 293, 122129. [Google Scholar] [CrossRef]
- Velasco, P.; Jegatheesan, V.; Othman, M. Recovery of dissolved methane from anaerobic membrane bioreactor using degassing membrane contactors. Front. Environ. Sci. 2018, 6, 151. [Google Scholar] [CrossRef]
- Waterman, K.C.; Adami, R.C. Accelerated aging: Prediction of chemical stability of pharmaceuticals. Int. J. Pharm. 2005, 293, 101–125. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Ru, D.; Wang, Y.; Zhang, J.; Jiang, L.; Xu, X.; Nie, L. Optimization of a nitrite-dependent anaerobic methane oxidation (n-damo) process by enhancing methane availability. Bioresour. Technol. 2019, 275, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Rosero-Chasoy, G.; Rodríguez-Jasso, R.M.; Aguilar, C.N.; Buitrón, G.; Chairez, I.; Ruiz, H.A. Microbial co-culturing strategies for the production high value compounds, a reliable framework towards sustainable biorefinery implementation—An overview. Bioresour. Technol. 2021, 321, 124458. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.; de Roy, K.; Thas, O.; De Neve, J.; Hoefman, S.; Vandamme, P.; Heylen, K.; Boon, N. The more, the merrier: Heterotroph richness stimulates methanotrophic activity. ISME J. 2014, 8, 1945–1948. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Li, W.; Guo, Y.; Zhang, Z.; Shi, W.; Cui, F.; Lens, P.N.L.; Tay, J.H. Microalgal-bacterial consortia: From interspecies interactions to biotechnological applications. Renew. Sustain. Energy Rev. 2020, 118, 109563. [Google Scholar] [CrossRef]
- Sahoo, K.K.; Goswami, G.; Das, D. Biotransformation of methane and carbon dioxide into high-value products by methanotrophs: Current state of art and future prospects. Front. Microbiol. 2021, 12, 636486. [Google Scholar] [CrossRef]
- Safitri, A.S.; Hamelin, J.; Kommedal, R.; Milferstedt, K. Engineered methanotrophic syntrophy in photogranule communities removes dissolved methane. Water Res. X 2021, 12, 100106. [Google Scholar] [CrossRef]
- Whittenbury, R.; Phillips, K.C.; Wilkinson, J.F. Enrichment, isolation and some properties of methane-utilizing bacteria. J. Gen. Microbiol. 1970, 61, 205–218. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Braunegg, G.; Sonnleitner, B.; Lafferty, R.M. A rapid gas chromatographic method for the determination of poly-β-hydroxybutyric acid in microbial biomass. Eur. J. Appl. Microbiol. Biotechnol. 1978, 6, 29–37. [Google Scholar] [CrossRef]
- Khang, T.U.; Kim, M.J.; Yoo, J.I.; Sohn, Y.J.; Jeon, S.G.; Park, S.J.; Na, J.-G. Rapid analysis of polyhydroxyalkanoate contents and its monomer compositions by pyrolysis-gas chromatography combined with mass spectrometry (Py-GC/MS). Int. J. Biol. Macromol. 2021, 174, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Lebranchu, A.; Blanchard, F.; Fick, M.; Pacaud, S.; Olmos, E.; Delaunay, S. Pilot-scale biomethanation of cattle manure using dense membranes. Bioresour. Technol. 2019, 284, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Legoix, L.N.; Ruffine, L.; Deusner, C.; Haeckel, M. Experimental study of mixed gas hydrates from gas feed containing CH4, CO2 and N2: Phase equilibrium in the presence of excess water and gas exchange. Energies 2018, 11, 1984. [Google Scholar] [CrossRef]
- Amaral, J.A.; Knowles, R. Growth of methanotrophs in methane and oxygen counter gradients. FEMS Microbiol. Lett. 1995, 126, 215–220. [Google Scholar] [CrossRef]
- Komarek, J.; Hindak, F. Taxonomy of the new isolated strains of Chroococcidiopsis (Cyanophyceae). Algol. Stud. 1975, 13, 311–329. [Google Scholar]
- Milferstedt, K.; Kuo-Dahab, W.C.; Butler, C.S.; Hamelin, J.; Abouhend, A.S.; Stauch-White, K.; McNair, A.; Watt, C.; Carbajal-González, B.I.; Dolan, S.; et al. The importance of filamentous cyanobacteria in the development of oxygenic photogranules. Sci. Rep. 2017, 7, 17944. [Google Scholar] [CrossRef]
- Jo, S.Y.; Son, J.; Sohn, Y.J.; Lim, S.H.; Lee, J.Y.; Yoo, J.I.; Park, S.Y.; Na, J.G.; Park, S.J. A shortcut to carbon-neutral bioplastic production: Recent advances in microbial production of polyhydroxyalkanoates from C1 resources. Int. J. Biol. Macromol. 2021, 192, 978–998. [Google Scholar] [CrossRef]
- Rasouli, Z.; Valverde-Pérez, B.; D’Este, M.; De Francisci, D.; Angelidaki, I. Nutrient recovery from industrial wastewater as single cell protein by a co-culture of green microalgae and methanotrophs. Biochem. Eng. J. 2018, 134, 129–135. [Google Scholar] [CrossRef]
- Murrell, J.C.; Jetten, M.S.M. The microbial methane cycle. Environ. Microbiol. Rep. 2009, 1, 279–284. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Nitrogen Source Concentration (mM) | Biomass (g/L) | PHB Content (mg/100 mg Biomass) | PHB Titer (mg/L) |
|---|---|---|---|
| 11.5 | 1.30 ± 0.03 | 2.6 ± 0.2 | 33.3 ± 2.1 |
| 5.7 | 1.29 ± 0.00 | 2.8 ± 0.4 | 36.6 ± 4.6 |
| 2.9 | 1.13 ± 0.04 | 7.4 ± 0.0 | 83.0 ± 2.6 |
| 1.4 | 0.50 ± 0.06 | 13.7 ± 1.5 | 67.3 ± 1.4 |
| 9.9 (OB3b control) | 0.44 ± 0.01 | 38.1 ± 0.0 | 167.6 ± 5.4 |
| 1.6 (OPGs control) | 0.53 ± 0.05 | 0.2 ± 0.0 | 1.1 ± 0.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashoor, S.; Jun, S.-H.; Ko, H.D.; Lee, J.; Hamelin, J.; Milferstedt, K.; Na, J.-G. Polyhydroxybutyrate Production from Methane and Carbon Dioxide by a Syntrophic Consortium of Methanotrophs with Oxygenic Photogranules without an External Oxygen Supply. Microorganisms 2023, 11, 1110. https://doi.org/10.3390/microorganisms11051110
Ashoor S, Jun S-H, Ko HD, Lee J, Hamelin J, Milferstedt K, Na J-G. Polyhydroxybutyrate Production from Methane and Carbon Dioxide by a Syntrophic Consortium of Methanotrophs with Oxygenic Photogranules without an External Oxygen Supply. Microorganisms. 2023; 11(5):1110. https://doi.org/10.3390/microorganisms11051110
Chicago/Turabian StyleAshoor, Selim, Seong-Hoon Jun, Han Do Ko, Jinwon Lee, Jérôme Hamelin, Kim Milferstedt, and Jeong-Geol Na. 2023. "Polyhydroxybutyrate Production from Methane and Carbon Dioxide by a Syntrophic Consortium of Methanotrophs with Oxygenic Photogranules without an External Oxygen Supply" Microorganisms 11, no. 5: 1110. https://doi.org/10.3390/microorganisms11051110