

New Perspectives on BolA: A Still Mysterious Protein Connecting Morphogenesis, Biofilm Production, Virulence, Iron Metabolism, and Stress Survival

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
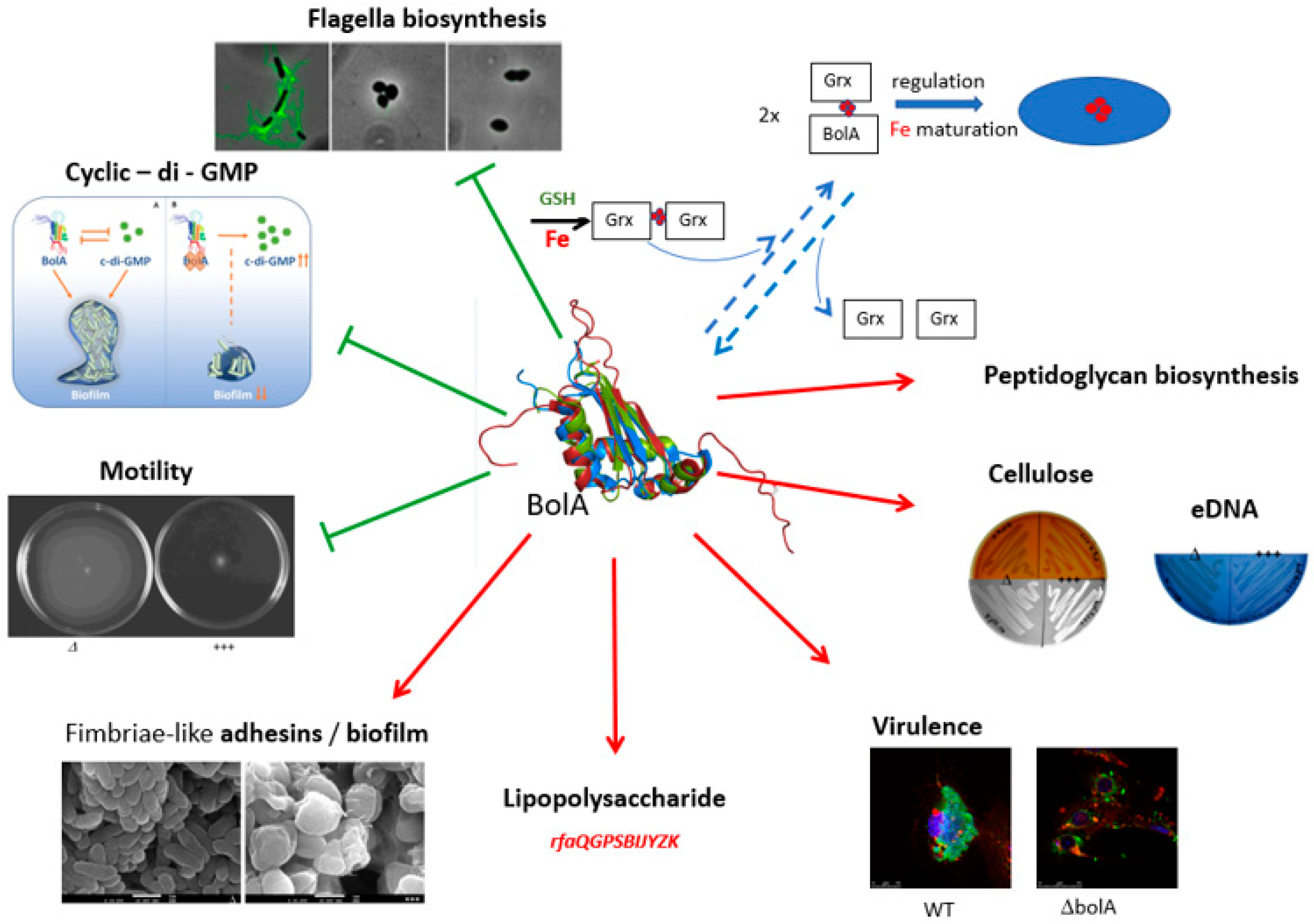
:1. BolA Function and Regulation
1.1. The Role of BolA in E. coli Survival
1.2. The Role of BolA in Virulence
2. BolA and Biofilms
3. BolA Phosphorylation and Interaction with Other Cellular Proteins
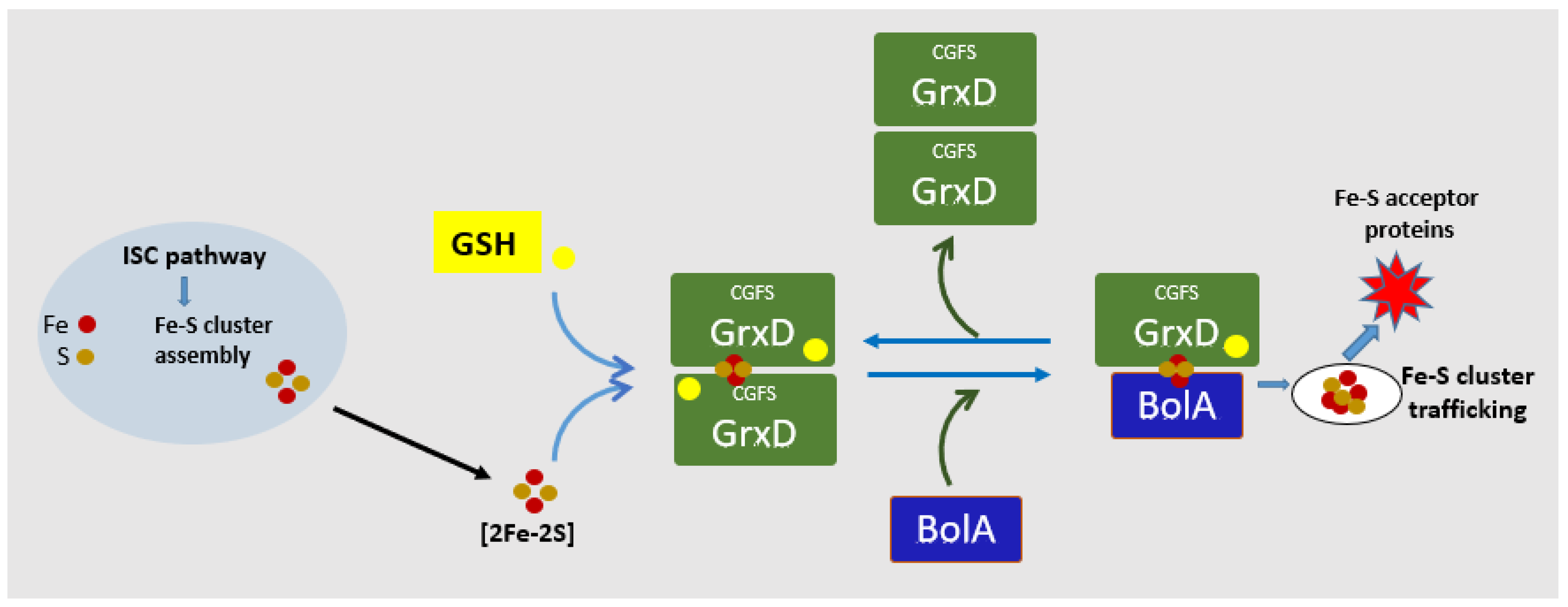
4. BolA Interacts with Grx Mediating Intracellular Iron Homeostasis
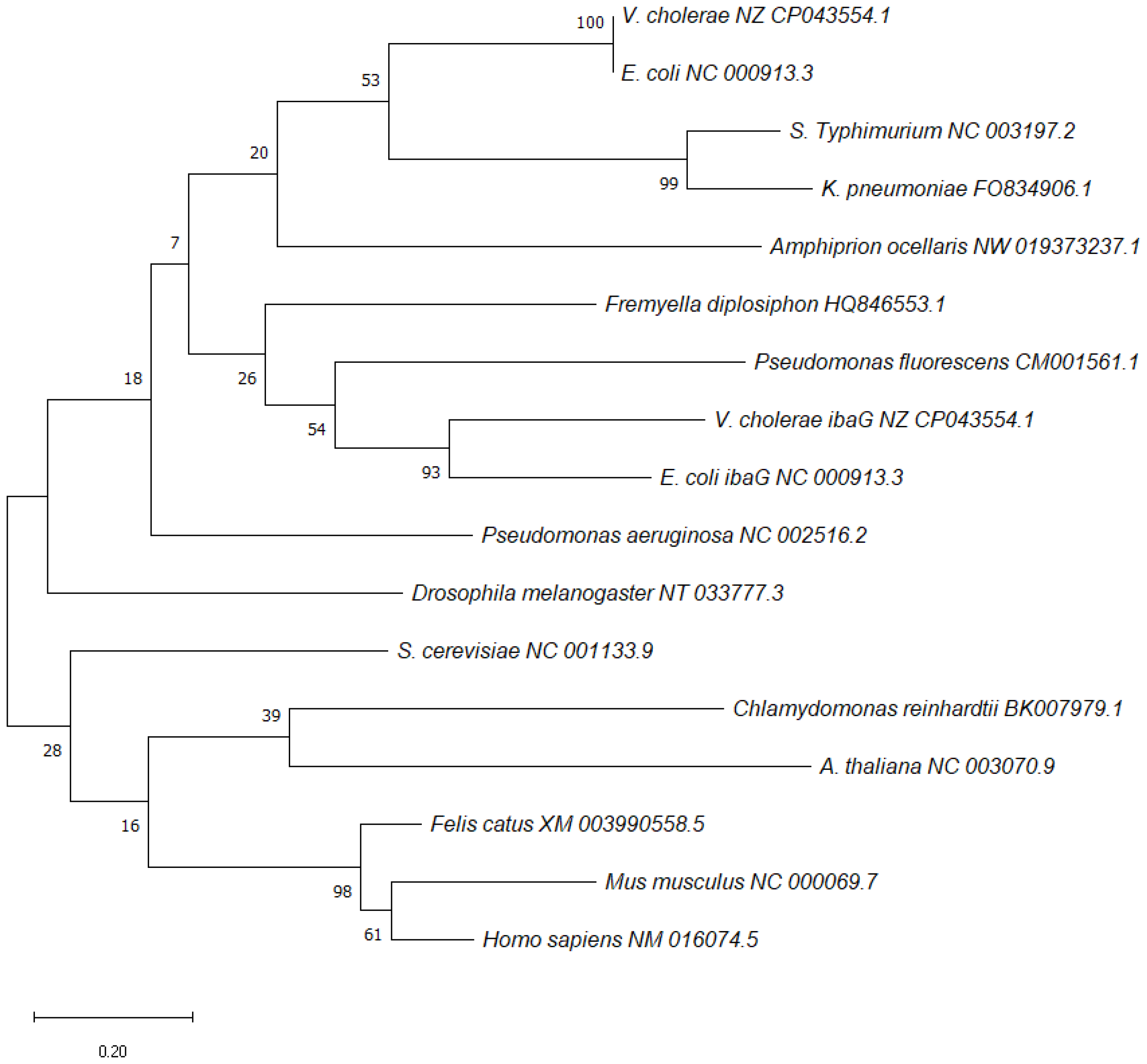
5. BolA Orthologues
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aldea, M.; Hernández-Chico, C.; de la Campa, A.G.; Kushner, S.R.; Vicente, M. Identification; cloning; and expression of bolA; an ftsZ-dependent morphogene of Escherichia coli. J. Bacteriol. 1988, 170, 5169–5176. [Google Scholar] [CrossRef] [Green Version]
- Lange, R.; Hengge-Aronis, R. Growth phase-regulated expression of bolA and morphology of stationary-phase Escherichia coli cells are controlled by the novel sigma factor sigma S. J. Bacteriol. 1991, 173, 4474–4481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, R.; Hengge-Aronis, R. Identification of a central regulator of stationary-phase gene expression in Escherichia coli. Mol. Microbiol. 1991, 5, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Aldea, M.; Garrido, T.; Hernández-Chico, C.; Vicente, M.; Kushner, S.R. Induction of a growth-phase-dependent promoter triggers transcription of bolA; an Escherichia coli morphogene. EMBO J. 1989, 8, 3923–3931. [Google Scholar] [CrossRef] [PubMed]
- Freire, P.; Vieira, H.A.; Furtado, A.R.; de Pedro, M.A.; Arraiano, C.M. Effect of the morphogene bolA on the permeability of the Escherichia coli outer membrane. FEMS Microbiol. Lett. 2006, 260, 106–111. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.M.; Freire, P.; Vicente, M.; Arraiano, C.M. The stationary-phase morphogene bolA from Escherichia coli is induced by stress during early stages of growth. Mol. Microbiol. 1999, 32, 789–798. [Google Scholar] [CrossRef] [Green Version]
- Vieira, H.A.; Freire, P.; Arraiano, C.M. Effect of Escherichia coli Morphogene bolA on Biofilms. Appl. Environ. Microbiol. 2004, 70, 5682–5684. [Google Scholar] [CrossRef] [Green Version]
- Guinote, I.B.; Moreira, R.N.; Barahona, S.; Freire, P.; Vicente, M.; Arraiano, C.M. Breaking through the stress barrier: The role of BolA in Gram-negative survival. World J. Microbiol. Biotechnol. 2014, 30, 2559–2566. [Google Scholar] [CrossRef]
- Qin, L.; Wang, M.; Zuo, J.; Feng, X.; Liang, X.; Wu, Z.; Ye, H. Cytosolic BolA Plays a Repressive Role in the Tolerance against Excess Iron and MV-Induced Oxidative Stress in Plants. PLoS ONE 2015, 10, e0124887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rey, P.; Taupin-Broggini, M.; Couturier, J.; Vignols, F.; Rouhier, N. Is There a Role for Glutaredoxins and BOLAs in the Perception of the Cellular Iron Status in Plants? Front. Plant Sci. 2019, 10, 712. [Google Scholar] [CrossRef] [Green Version]
- Kasai, T.; Inoue, M.; Koshiba, S.; Yabuki, T.; Aoki, M.; Nunokawa, E.; Seki, E.; Matsuda, T.; Matsuda, N.; Tomo, Y.; et al. Solution structure of a BolA-like protein from Mus musculus. Protein Sci. 2004, 13, 545–548. [Google Scholar] [CrossRef] [PubMed]
- NCBI Resource Coordinators. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2016, 44, D7–D19. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Galego, L.; Barahona, S.; Romão, C.; Arraiano, C.M. Phosphorylation status of BolA affects its role in transcription and biofilm development. FEBS J. 2021, 288, 961–979. [Google Scholar] [CrossRef]
- Santos, J.M.; Lobo, M.; Matos, P.A.; de Pedro, M.A.; Arraiano, C.M. The gene bolA regulates dacA (PBP5); dacC (PBP6) and ampC (AmpC); promoting normal morphology in Escherichia coli. Mol. Microbiol. 2002, 45, 1729–1740. [Google Scholar] [CrossRef] [PubMed]
- Freire, P.; Moreira, R.N.; Arraiano, C.M. BolA Inhibits Cell Elongation and Regulates MreB Expression Levels. J. Mol. Biol. 2009, 385, 1345–1351. [Google Scholar] [CrossRef]
- Guinote, I.B.; Matos, R.G.; Freire, P.; Arraiano, C.M. BolA Affects Cell Growth; and Binds to the Promoters of Penicillin-Binding Proteins 5 and 6 and Regulates Their Expression. J. Microbiol. Biotechnol. 2011, 21, 243–251. [Google Scholar] [CrossRef]
- Moreira, R.N.; Dressaire, C.; Domingues, S.; Arraiano, C.M. A new target for an old regulator: H-NS represses transcription of bolA morphogene by direct binding to both promoters. Biochem. Biophys. Res. Commun. 2011, 411, 50–55. [Google Scholar] [CrossRef]
- Yamamoto, K. Negative regulation of the bolA1p of Escherichia coli K-12 by the transcription factor OmpR for osmolarity response genes. FEMS Microbiol. Lett. 2000, 186, 257–262. [Google Scholar] [CrossRef]
- Freire, P.; Amaral, J.; Santos, J.; Arraiano, C. Adaptation to carbon starvation: RNase III ensures normal expression levels of bolA1p mRNA and σS. Biochimie 2006, 88, 341–346. [Google Scholar] [CrossRef]
- Cao, G.; Sarkar, N. Stationary Phase-Specific mRNAs in Escherichia coli Are Polyadenylated. Biochem. Biophys. Res. Commun. 1997, 239, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Dressaire, C.; Moreira, R.N.; Barahona, S.; Alves de Matos, A.P.; Arraiano, C.M. BolA Is a Transcriptional Switch That Turns off Motility and Turns on Biofilm Development. mBio 2015, 6, e02352-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lhocine, N.; Arena, E.T.; Bomme, P.; Ubelmann, F.; Prévost, M.-C.; Robine, S.; Sansonetti, P.J. Apical Invasion of Intestinal Epithelial Cells by Salmonella Typhimurium Requires Villin to Remodel the Brush Border Actin Cytoskeleton. Cell Host Microbe 2015, 17, 164–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fàbrega, A.; Vila, J. Salmonella enterica Serovar Typhimurium Skills To Succeed in the Host: Virulence and Regulation. Clin. Microbiol. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mil-Homens, D.; Barahona, S.; Moreira, R.N.; Silva, I.J.; Pinto, S.N.; Fialho, A.M.; Arraiano, C.M. Stress Response Protein BolA Influences Fitness and Promotes Salmonella enterica Serovar Typhimurium Virulence. Appl. Environ. Microbiol. 2018, 84, e02850-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graça-Lopes, G.; Graça, G.; Barahona, S.; Moreira, R.N.; Arraiano, C.M.; Gonçalves, L.G. NMR-Metabolomics Shows That BolA Is an Important Modulator of Salmonella Typhimurium Metabolic Processes under Virulence Conditions. Metabolites 2019, 9, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Yan, X.; Bai, J.; Xiang, L.; Ding, M.; Li, Q.; Zhang, B.; Liang, Q.; Zhou, Y. Identification of the BolA Protein Reveals a Novel Virulence Factor in K. pneumoniae that Contributes to Survival in Host. Microbiol. Spectr. 2022, 10, e00378-22. [Google Scholar] [CrossRef]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival Mechanisms of Clinically Relevant Microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Sousa, A.M.; Machado, I.; Pereira, M.O.; Khan, S.; Morton, G.; Hadi, S. Role of bolA and rpoS genes in biofilm formation and adherence pattern by Escherichia coli K-12 MG1655 on polypropylene; stainless steel; and silicone surfaces. Acta Microbiol. Immunol. Hung. 2017, 64, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Guttenplan, S.B.; Kearns, D.B. Regulation of flagellar motility during biofilm formation. FEMS Microbiol. Rev. 2013, 37, 849–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azam, M.W.; Zuberi, A.; Khan, A.U. bolA gene involved in curli amyloids and fimbriae production in E. coli: Exploring pathways to inhibit biofilm and amyloid formation. J. Biol. Res.-Thessalon. 2020, 27, 10. [Google Scholar] [CrossRef]
- Povolotsky, T.L.; Hengge, R. ‘Life-style’ control networks in Escherichia coli: Signaling by the second messenger c-di-GMP. J. Biotechnol. 2012, 160, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Paul, R.; Weiser, S.; Amiot, N.C.; Chan, C.; Schirmer, T.; Giese, B.; Jenal, U. Cell cycle-dependent dynamic localization of a bacterial response regulator with a novel di-guanylate cyclase output domain. Genes Dev. 2004, 18, 715–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simm, R.; Morr, M.; Kader, A.; Nimtz, M.; Römling, U. GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessility to motility. Mol. Microbiol. 2004, 53, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Boyd, C.D.; O’Toole, G.A. Second Messenger Regulation of Biofilm Formation: Breakthroughs in Understanding c-di-GMP Effector Systems. Annu. Rev. Cell Dev. Biol. 2012, 28, 439–462. [Google Scholar] [CrossRef] [Green Version]
- Christen, M.; Christen, B.; Folcher, M.; Schauerte, A.; Jenal, U. Identification and Characterization of a Cyclic di-GMP-specific Phosphodiesterase and Its Allosteric Control by GTP. J. Biol. Chem. 2005, 280, 30829–30837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, R.N.; Dressaire, C.; Barahona, S.; Galego, L.; Kaever, V.; Jenal, U.; Arraiano, C.M. BolA Is Required for the Accurate Regulation of c-di-GMP, a Central Player in Biofilm Formation. mBio 2017, 8, e00443-17. [Google Scholar] [CrossRef]
- Deutscher, J.; Saier, M.H., Jr. Ser/Thr/Tyr Protein Phosphorylation in Bacteria—For Long Time Neglected, Now Well Established. Microb. Physiol. 2005, 9, 125–131. [Google Scholar] [CrossRef]
- Garcia, T.G.; Poncet, S.; Derouiche, A.; Shi, L.; Mijakovic, I.; Noirot-Gros, M.-F. Role of Protein Phosphorylation in the Regulation of Cell Cycle and DNA-Related Processes in Bacteria. Front. Microbiol. 2016, 7, 184. [Google Scholar] [CrossRef]
- Kruse, T. Dysfunctional MreB inhibits chromosome segregation in Escherichia coli. EMBO J. 2003, 22, 5283–5292. [Google Scholar] [CrossRef] [Green Version]
- Van den Ent, F.; Izoré, T.; Bharat, T.A.; Johnson, C.M.; Löwe, J. Bacterial actin MreB forms antiparallel double filaments. eLife 2014, 3, e02634. [Google Scholar] [CrossRef]
- Kruse, T.; Bork-Jensen, J.; Gerdes, K. The morphogenetic MreBCD proteins of Escherichia coli form an essential membrane-bound complex. Mol. Microbiol. 2004, 55, 78–89. [Google Scholar] [CrossRef]
- Singh, S.P.; Montgomery, B.L. Morphogenes bolA and mreB mediate the photoregulation of cellular morphology during complementary chromatic acclimation in Fremyella diplosiphon. Mol. Microbiol. 2014, 93, 167–182. [Google Scholar] [CrossRef]
- Li, H.; Outten, C.E. Monothiol CGFS Glutaredoxins and BolA-like Proteins: [2Fe-2S] Binding Partners in Iron Homeostasis. Biochemistry 2012, 51, 4377–4389. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Mapolelo, D.T.; Dingra, N.N.; Keller, G.; Riggs-Gelasco, P.J.; Winge, D.R.; Johnson, M.K.; Outten, C.E. Histidine 103 in Fra2 Is an Iron-Sulfur Cluster Ligand in the [2Fe-2S] Fra2-Grx3 Complex and Is Required for in Vivo Iron Signaling in Yeast. J. Biol. Chem. 2011, 286, 867–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, N.; Gold, B.; Liu, N.L.; Prathapam, R.; Sterling, H.J.; Willams, E.R.; Butland, G. The E. coli Monothiol Glutaredoxin GrxD Forms Homodimeric and Heterodimeric FeS Cluster Containing Complexes. Biochemistry 2011, 50, 8957–8969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dlouhy, A.C.; Li, H.; Albetel, A.-N.; Zhang, B.; Mapolelo, D.T.; Randeniya, S.; Holland, A.A.; Johnson, M.K.; Outten, C.E. The Escherichia coli BolA Protein IbaG Forms a Histidine-Ligated [2Fe-2S]-Bridged Complex with Grx4. Biochemistry 2016, 55, 6869–6879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynen, M.A.; Spronk, E.M.; Gabaldón, T.; Snel, B. Combining data from genomes; Y2H and 3D structure indicates that BolA is a reductase interacting with a glutaredoxin. FEBS Lett. 2005, 579, 591–596. [Google Scholar] [CrossRef] [Green Version]
- Couturier, J.; Jacquot, J.P.; Rouhier, N. Evolution and diversity of glutaredoxins in photosynthetic organisms. Cell. Mol. Life Sci. 2009, 66, 2539–2557. [Google Scholar] [CrossRef]
- Dlouhy, A.C.; Outten, C.E. The Iron Metallome in Eukaryotic Organisms. In Metallomics and the Cell; Springer: Dordrecht, The Netherlands, 2013; pp. 241–278. [Google Scholar] [CrossRef] [Green Version]
- Banci, L.; Brancaccio, D.; Ciofi-Baffoni, S.; Del Conte, R.; Gadepalli, R.; Mikolajczyk, M.; Neri, S.; Piccioli, M.; Winkelmann, J. [2Fe-2S] cluster transfer in iron–sulfur protein biogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, 6203–6208. [Google Scholar] [CrossRef] [Green Version]
- Mühlenhoff, U.; Molik, S.; Godoy, J.R.; Uzarska, M.A.; Richter, N.; Seubert, A.; Zhang, Y.; Stubbe, J.; Pierrel, F.; Herrero, E.; et al. Cytosolic Monothiol Glutaredoxins Function in Intracellular Iron Sensing and Trafficking via Their Bound Iron-Sulfur Cluster. Cell Metab. 2010, 12, 373–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, A.; Fladvad, M.; Berndt, C.; Andrésen, C.; Lillig, C.H.; Neubauer, P.; Sunnerhagen, M.; Holmgren, A.; Vlamis, A. A Novel Monothiol Glutaredoxin (Grx4) from Escherichia coli Can Serve as a Substrate for Thioredoxin Reductase. J. Biol. Chem. 2005, 280, 24544–24552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Mapolelo, D.T.; Dingra, N.N.; Naik, S.G.; Lees, N.S.; Hoffman, B.M.; Riggs-Gelasco, P.J.; Huynh, B.H.; Johnson, M.K.; Outten, C.E. The Yeast Iron Regulatory Proteins Grx3/4 and Fra2 Form Heterodimeric Complexes Containing a [2Fe-2S] Cluster with Cysteinyl and Histidyl Ligation. Biochemistry 2009, 48, 9569–9581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, E.; Belli, G.; Casas, C. Structural and Functional Diversity of Glutaredoxins in Yeast. Curr. Protein Pept. Sci. 2010, 11, 659–668. [Google Scholar] [CrossRef] [Green Version]
- Poor, C.B.; Wegner, S.V.; Li, H.; Dlouhy, A.C.; Schuermann, J.P.; Sanishvili, R.; Hinshaw, J.R.; Riggs-Gelasco, P.J.; Outten, C.E.; He, C. Molecular mechanism and structure of the Saccharomyces cerevisiae iron regulator Aft2. Proc. Natl. Acad. Sci. USA 2014, 111, 4043–4048. [Google Scholar] [CrossRef] [Green Version]
- Dlouhy, A.C.; Beaudoin, J.; Labbé, S.; Outten, C.E. Schizosaccharomyces pombe Grx4 regulates the transcriptional repressor Php4 via [2Fe–2S] cluster binding. Metallomics 2017, 9, 1096–1105. [Google Scholar] [CrossRef]
- Ye, H.; Rouault, T.A. Human Iron−Sulfur Cluster Assembly, Cellular Iron Homeostasis, and Disease. Biochemistry 2010, 49, 4945–4956. [Google Scholar] [CrossRef]
- Rouhier, N.; Couturier, J.; Johnson, M.K.; Jacquot, J.P. Glutaredoxins: Roles in iron homeostasis. Trends Biochem. Sci. 2010, 35, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Iwema, T.; Picciocchi, A.; Traore, D.A.K.; Ferrer, J.-L.; Chauvat, F.; Jacquamet, L. Structural Basis for Delivery of the Intact [Fe2S2] Cluster by Monothiol Glutaredoxin. Biochemistry 2009, 48, 6041–6043. [Google Scholar] [CrossRef]
- Haunhorst, P.; Berndt, C.; Eitner, S.; Godoy, J.R.; Lillig, C.H. Characterization of the human monothiol glutaredoxin 3 (PICOT) as iron–sulfur protein. Biochem. Biophys. Res. Commun. 2010, 394, 372–376. [Google Scholar] [CrossRef]
- Lillig, C.H.; Berndt, C. Cellular functions of glutathione. Biochim. Biophys. Acta BBA Gen. Subj. 2013, 1830, 3137–3138. [Google Scholar] [CrossRef]
- Ito, T.; Tashiro, K.; Muta, S.; Ozawa, R.; Chiba, T.; Nishizawa, M.; Yamamoto, K.; Kuhara, S.; Sakaki, Y. Toward a protein–protein interaction map of the budding yeast: A comprehensive system to examine two-hybrid interactions in all possible combinations between the yeast proteins. Proc. Natl. Acad. Sci. USA 2000, 97, 1143–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumánovics, A.; Chen, O.S.; Li, L.; Bagley, D.; Adkins, E.M.; Lin, H.; Dingra, N.N.; Outten, C.E.; Keller, G.; Winge, D.; et al. Identification of FRA1 and FRA2 as Genes Involved in Regulating the Yeast Iron Regulon in Response to Decreased Mitochondrial Iron-Sulfur Cluster Synthesis. J. Biol. Chem. 2008, 283, 10276–10286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey, A.G.; Palenchar, D.J.; Wildemann, J.D.; Philpott, C.C. A Glutaredoxin·BolA Complex Serves as an Iron-Sulfur Cluster Chaperone for the Cytosolic Cluster Assembly Machinery. J. Biol. Chem. 2016, 291, 22344–22356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uzarska, M.A.; Nasta, V.; Weiler, B.D.; Spantgar, F.; Ciofi-Baffoni, S.; Saviello, M.R.; Gonnelli, L.; Mühlenhoff, U.; Banci, L.; Lill, R. Mitochondrial Bol1 and Bol3 function as assembly factors for specific iron-sulfur proteins. eLife 2016, 5, e16673. [Google Scholar] [CrossRef] [PubMed]
- Frazzon, J. Formation of iron–sulfur clusters in bacteria: An emerging field in bioinorganic chemistry. Curr. Opin. Chem. Biol. 2003, 7, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, J.C.; Ojeda, L.; Balk, J.; Mühlenhoff, U.; Lill, R.; Winge, D.R. Activation of the Iron Regulon by the Yeast Aft1/Aft2 Transcription Factors Depends on Mitochondrial but Not Cytosolic Iron-Sulfur Protein Biogenesis. J. Biol. Chem. 2005, 280, 10135–10140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojeda, L.; Keller, G.; Muhlenhoff, U.; Rutherford, J.C.; Lill, R.; Winge, D.R. Role of Glutaredoxin-3 and Glutaredoxin-4 in the Iron Regulation of the Aft1 Transcriptional Activator in Saccharomyces cerevisiae. J. Biol. Chem. 2006, 281, 17661–17669. [Google Scholar] [CrossRef] [Green Version]
- Lesuisse, E.; Knight, A.B.; Courel, M.; Santos, R.; Camadro, J.M.; Dancis, A. Genome-Wide Screen for Genes with Effects on Distinct Iron Uptake Activities in Saccharomyces cerevisiae. Genetics 2005, 169, 107–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Manzaneque, M.T.; Tamarit, J.; Bellí, G.; Ros, J.; Herrero, E. Grx5 Is a Mitochondrial Glutaredoxin Required for the Activity of Iron/Sulfur Enzymes. Mol. Biol. Cell 2002, 13, 1109–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayala-Castro, C.; Saini, A.; Outten, F.W. Fe-S Cluster Assembly Pathways in Bacteria. Microbiol. Mol. Biol. Rev. 2008, 72, 110–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fladvad, M.; Bellanda, M.; Fernandes, A.P.; Mammi, S.; Vlamis-Gardikas, A.; Holmgren, A.; Sunnerhagen, M. Molecular Mapping of Functionalities in the Solution Structure of Reduced Grx4, a Monothiol Glutaredoxin from Escherichia coli. J. Biol. Chem. 2005, 280, 24553–24561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, E.; de la Torre-Ruiz, M.A. Monothiol glutaredoxins: A common domain for multiple functions. Cell. Mol. Life Sci. 2007, 64, 1518–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winger, J.A.; Marletta, M.A. Expression and Characterization of the Catalytic Domains of Soluble Guanylate Cyclase: Interaction with the Heme Domain. Biochemistry 2005, 44, 4083–4090. [Google Scholar] [CrossRef]
- Ye, H.; Abdel-Ghany, S.E.; Anderson, T.D.; Pilon-Smits, A.H.; Pilon, M. CpSufE Activates the Cysteine Desulfurase CpNifS for Chloroplastic Fe-S Cluster Formation. J. Biol. Chem. 2006, 281, 8958–8969. [Google Scholar] [CrossRef] [Green Version]
- Roret, T.; Tsan, P.; Couturier, J.; Zhang, B.; Johnsonet, M.K.; Rouhier, N.; Didierjean, C. Structural and Spectroscopic Insights into BolA-Glutaredoxin Complexes. J. Biol. Chem. 2014, 289, 24588–24598. [Google Scholar] [CrossRef] [Green Version]
- Couturier, J.; Wu, H.C.; Dhalleine, T.; Pégeot, H.; Sudre, S.; Gualberto, J.M.; Jacquot, J.-P.; Gaymard, F.; Vignols, F.; Rouhier, N. Monothiol Glutaredoxin–BolA Interactions: Redox Control of Arabidopsis thaliana BolA2 and SufE1. Mol. Plant. 2014, 7, 187–205. [Google Scholar] [CrossRef] [Green Version]
- Banci, L.; Ciofi-Baffoni, S.; Gajda, K.; Muzzioli, R.; Peruzzini, R.; Winkelmann, J. N-terminal domains mediate [2Fe-2S] cluster transfer from glutaredoxin-3 to anamorsin. Nat. Chem. Biol. 2015, 11, 772–778. [Google Scholar] [CrossRef]
- Uzarska, M.A.; Dutkiewicz, R.; Freibert, S.A.; Lill, R.; Mühlenhoff, U. The mitochondrial Hsp70 chaperone Ssq1 facilitates Fe/S cluster transfer from Isu1 to Grx5 by complex formation. Mol. Biol. Cell 2013, 24, 1830–1841. [Google Scholar] [CrossRef]
- Cameron, J.M.; Janer, A.; Levandovskiy, V.; MacKay, N.; Rouault, T.A.; Tong, W.-H.; Ogilvie, I.; Shoubridge, E.A.; Robinson, B.H. Mutations in Iron-Sulfur Cluster Scaffold Genes NFU1 and BOLA3 Cause a Fatal Deficiency of Multiple Respiratory Chain and 2-Oxoacid Dehydrogenase Enzymes. Am. J. Hum. Genet. 2011, 89, 486–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lill, R.; Hoffmann, B.; Molik, S.; Pierik, A.J.; Rietzschel, N.; Stehling, O.; Uzarska, M.A.; Webert, H.; Wilbrecht, C.; Mühlenhoff, U. The role of mitochondria in cellular iron–sulfur protein biogenesis and iron metabolism. Biochim. Biophys. Acta BBA Mol. Cell Res. 2012, 1823, 1491–1508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhlenhoff, U. Components involved in assembly and dislocation of iron-sulfur clusters on the scaffold protein Isu1p. EMBO J. 2003, 22, 4815–4825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willems, P.; Wanschers, F.J.; Esseling, J.; Szklarczyk, R.; Kudlaet, U.; Duarte, I.; Forkink, M.; Nooteboom, M.; Swarts, H.; Gloerich, J.; et al. BOLA1 Is an Aerobic Protein That Prevents Mitochondrial Morphology Changes Induced by Glutathione Depletion. Antioxid. Redox Signal. 2013, 18, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Guinote, I.B.; Moreira, R.N.; Freire, P.; Arraiano, C.M. Characterization of the BolA Homolog IbaG: A New Gene Involved in Acid Resistance. J. Microbiol. Biotechnol. 2012, 22, 484–493. [Google Scholar] [CrossRef] [Green Version]
- Fleurie, A.; Zoued, A.; Alvarez, L.; Hines, K.M.; Cava, F.; Xu, L.; Davis, B.M.; Waldor, M.K. A Vibrio cholerae BolA-Like Protein Is Required for Proper Cell Shape and Cell Envelope Integrity. mBio 2019, 10, e00790-19. [Google Scholar] [CrossRef] [Green Version]
- Koch, B.; Nybroe, O. Initial characterization of a bolA homologue from Pseudomonas fluorescens indicates different roles for BolA-like proteins in P. fluorescens and Escherichia coli. FEMS Microbiol. Lett. 2006, 262, 48–56. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, M.; Lee, J.K.; Kim, M.J.; Jin, Y.H.; Seong, R.H.; Hong, S.H.; Joe, C.O.; Park, S.D. Identification and expression of uvi31+; a UV-inducible gene from Schizosaccharomyces pombe. Environ. Mol. Mutagen. 1997, 30, 72–81. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, H.S.; Lee, J.K.; Lee, C.B.; Park, S.D. Regulation of septation and cytokinesis during resumption of cell division requires uvi31+, a UV-inducible gene of fission yeast. Mol Cells 2002, 14, 425–430. [Google Scholar]
- Khona, D.K.; Dongre, S.S.; Arraiano, C.M.; D’Souza, J.S. A BolA-like morphogene from the alga Chlamydomonas reinhardtii changes morphology and induces biofilm formation in Escherichia coli. FEMS Microbiol. Lett. 2013, 339, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Touati, D. Iron and Oxidative Stress in Bacteria. Arch. Biochem. Biophys. 2000, 373, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, P.; Wei, Q.; Andrews, S.C.; Vinckx, T. Iron homeostasis and management of oxidative stress response in bacteria. Metallomics 2011, 3, 540. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, A.A.; Galego, L.; Arraiano, C.M. New Perspectives on BolA: A Still Mysterious Protein Connecting Morphogenesis, Biofilm Production, Virulence, Iron Metabolism, and Stress Survival. Microorganisms 2023, 11, 632. https://doi.org/10.3390/microorganisms11030632
da Silva AA, Galego L, Arraiano CM. New Perspectives on BolA: A Still Mysterious Protein Connecting Morphogenesis, Biofilm Production, Virulence, Iron Metabolism, and Stress Survival. Microorganisms. 2023; 11(3):632. https://doi.org/10.3390/microorganisms11030632
Chicago/Turabian Styleda Silva, Ana Alves, Lisete Galego, and Cecília Maria Arraiano. 2023. "New Perspectives on BolA: A Still Mysterious Protein Connecting Morphogenesis, Biofilm Production, Virulence, Iron Metabolism, and Stress Survival" Microorganisms 11, no. 3: 632. https://doi.org/10.3390/microorganisms11030632