

Inhibition of Foodborne Pathogenic Bacteria by Excreted Metabolites of Serratia marcescens Strains Isolated from a Dairy-Producing Environment
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tested Bacterial Strains
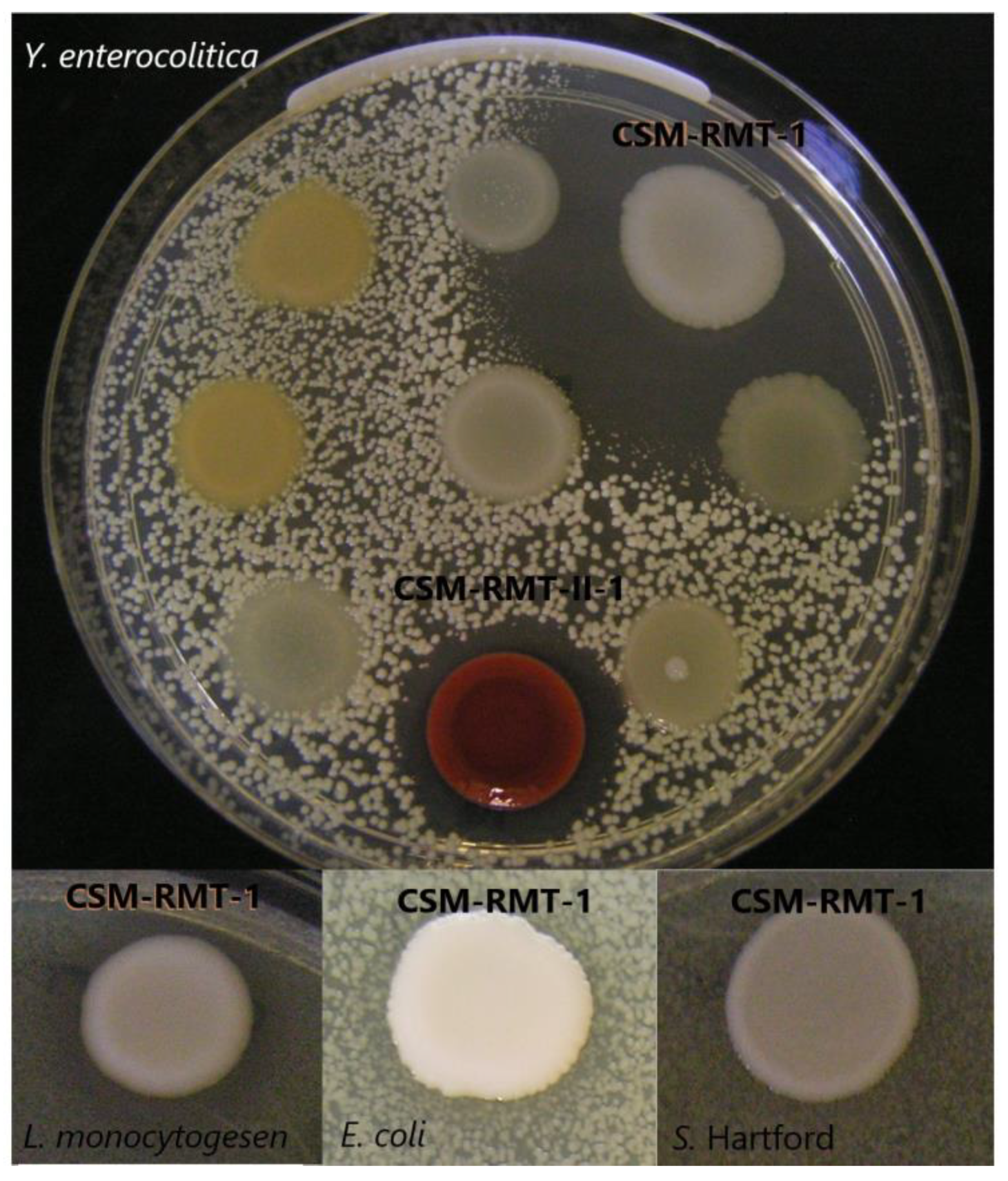
2.2. Screening for Antibacterial Activity of S. marcescens Strains and Testing the Inhibitory Effect of Their Cell-Free Supernatants
2.3. Cellophane Test
2.4. Detection of Proteolytic and Chitinase Activities of S. marcescens Strains
2.5. Detection of Prodigiosin-Producing Ability of S. marcescens Strains
2.6. PCR Analysis of Prodigiosin- and Chitinase-Encoding Genes
2.7. Co-Culturing of S. marcescens CSM-RMT-1 with a Food-Originated Salmonella enterica Subsp. enterica in Culture Broth
2.8. Statistical Analysis
3. Results
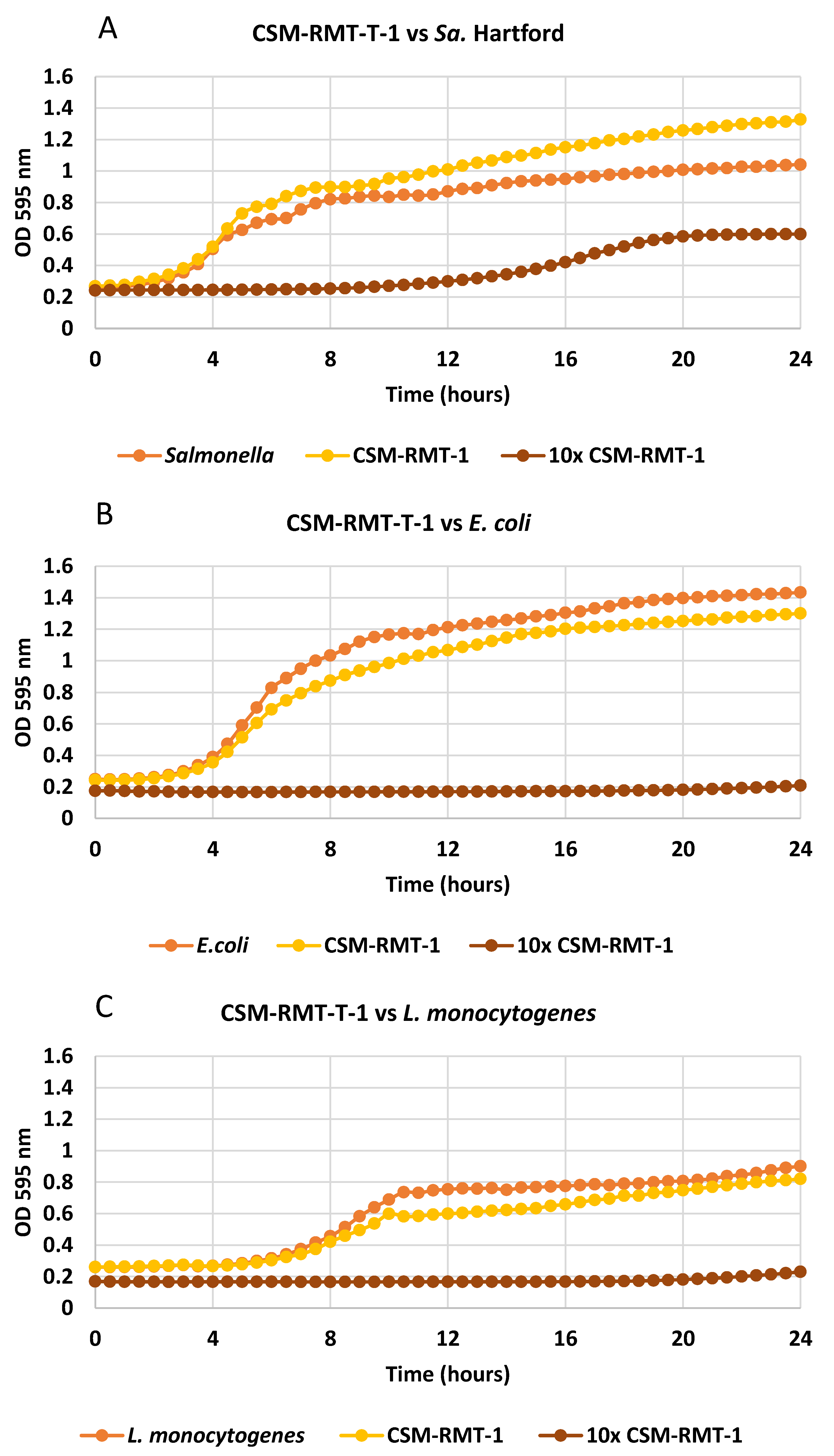
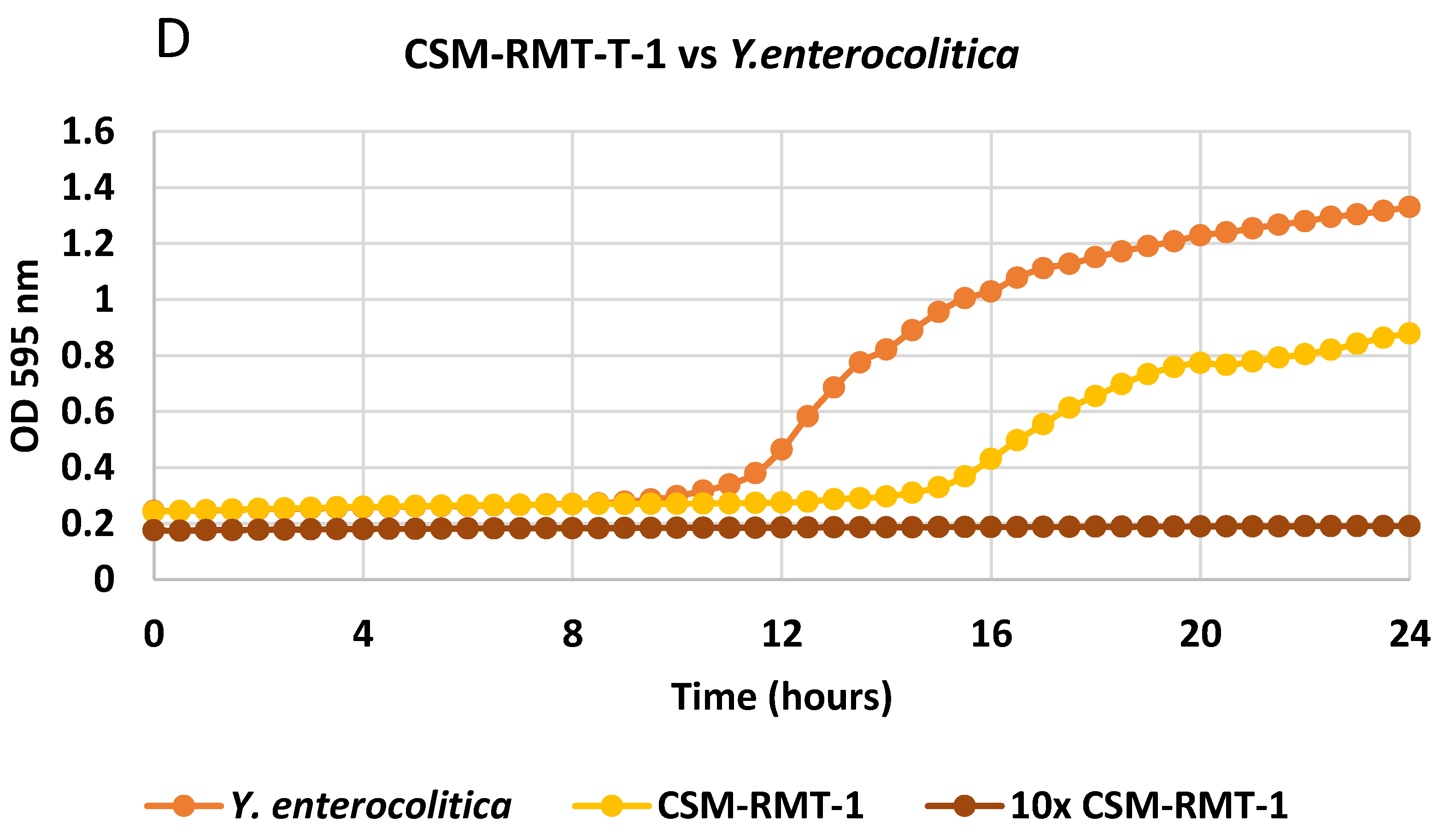
3.1. Inhibitory Activity of the Tested S. marcescens Strains on Different Foodborne Pathogenic Bacteria and the Effect of Cell-Free Supernatants
3.2. Protease and Chitinase Enzyme Activities of the S. marcescens Strains, and Their Prodigiosin Production
3.3. Presence of Prodigiosin- and Chitinase-Encoding Genes of S. marcescens Strains
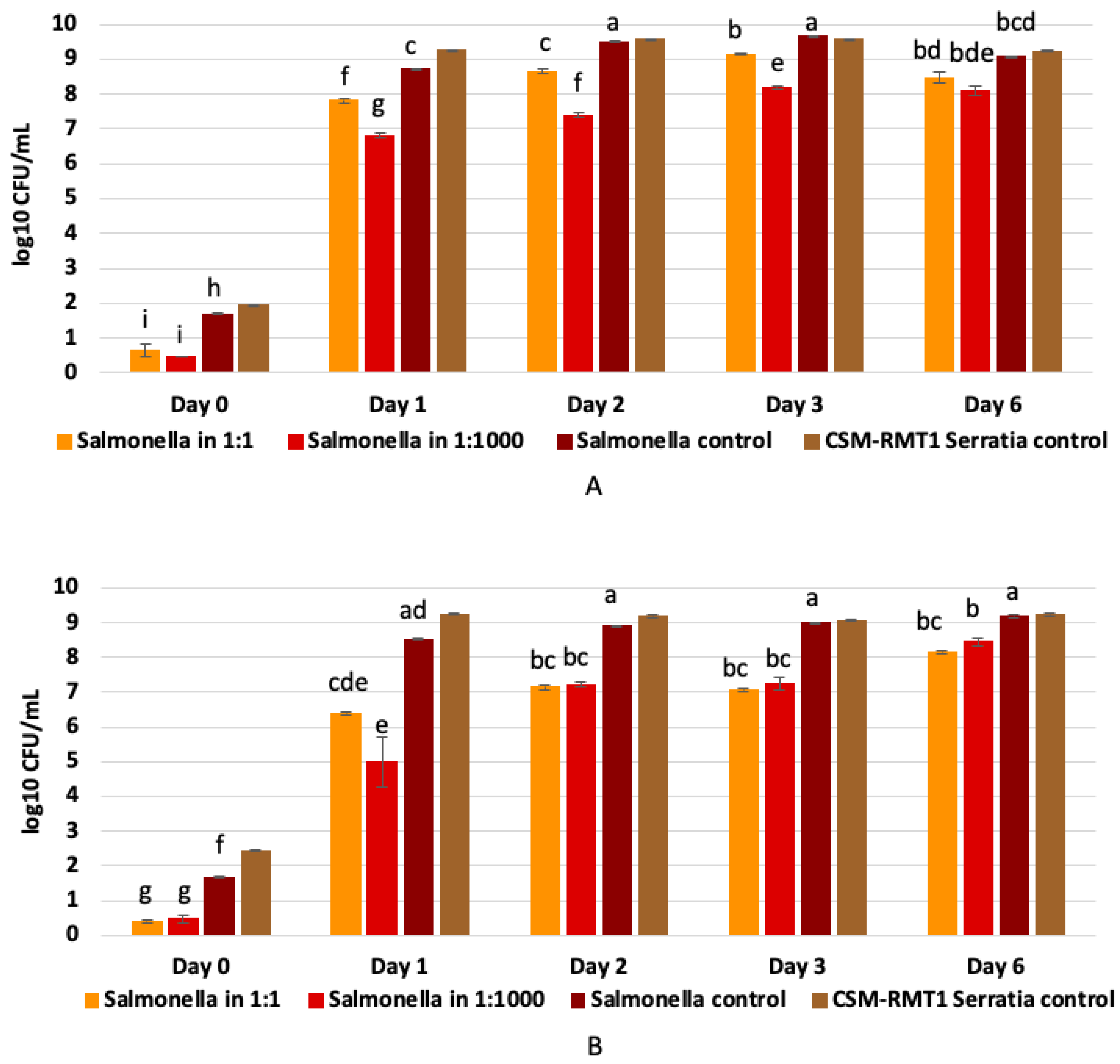
3.4. Results of Co-Culturing of Prodigiosin-Negative S. marcescens CSM-RMT-1 with A Food-Derived Salmonella enteritica Subsp. enteritica Strain
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EFSA. The European Union one health 2021 zoonoses report. EFSA J. 2022, 20, e07666. [Google Scholar] [CrossRef]
- Abdelhamid, A.G.; El-Dougdoug, N.K. Controlling foodborne pathogens with natural antimicrobials by biological control and antivirulence strategies. Heliyon 2020, 6, e05020. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, L.; Hudson, J.A.; Billington, C.; Withers, H. Biocontrol of foodborne bacteria: Past, present and future strategies. Food New Zealand 2007, 7, 25–36. [Google Scholar]
- Khanna, A.; Khanna, M.; Aggarwal, A. Serratia marcescens—A rare opportunistic nosocomial pathogen and measures to limit its spread in hospitalized patients. J. Clin. Diagn. Res. 2013, 7, 243–246. [Google Scholar]
- Yang, H.; Cheng, J.; Hu, L.; Zhu, Y.; Li, J. Mechanisms of antimicrobial resistance in Serratia marcescens. Afr. J. Microbiol. Res. 2012, 6, 4427–4437. [Google Scholar] [CrossRef]
- Ordentlich, A.; Elad, Y.; Chet, I. Rhizosphere colonization by Serratia marcescens for the control of Sclerotium rolfsii. Soil Biol. Biochem. 1987, 19, 747–751. [Google Scholar] [CrossRef]
- Parani, K.; Shetty, G.P.; Saha, B.K. Isolation of Serratia marcescens SR1 as a source of chitinase having potentiality of using as a biocontrol agent. Indian J. Microbiol. 2011, 51, 247–250. [Google Scholar] [CrossRef]
- Purkayastha, G.D.; Mangar, P.; Saha, A.; Saha, D. Evaluation of the biocontrol efficacy of a Serratia marcescens strain indigenous to tea rhizosphere for the management of root rot disease in tea. PLoS ONE 2018, 13, e0191761. [Google Scholar] [CrossRef]
- Mohamed, Z.K.; El-Sayed, S.A.; Radwan, T.E.E.; Abd El-Wahab, S.; Ghada, G.S. Potency evaluation of Serratia marcescens and Pseudomonas fluorescens as biocontrol agents for root-knot nematodes in Egypt. J. Appl. Sci. Res. 2009, 5, 93–102. [Google Scholar]
- Tu, S.; Qiu, X.; Cao, L.; Han, R.; Zhang, Y.; Liu, X. Expression and characterization of the chitinases from Serratia marcescens GEI strain for the control of Varroa destructor, a honey bee parasite. J. Invertebr. Pathol. 2010, 104, 75–82. [Google Scholar] [CrossRef]
- Li, P.; Kwok, A.H.Y.; Jiang, J.; Ran, T.; Xu, D.; Wang, W.; Leung, F.C. Comparative genome analyses of Serratia marcescens FS14 reveals its high antagonistic potential. PLoS ONE 2015, 10, e0123061. [Google Scholar] [CrossRef] [PubMed]
- Chet, I.; Ordentlich, A.; Shapira, R.; Oppenheim, A. Mechanisms of biocontrol of soil-borne plant pathogens by Rhizobacteria. Plant Soil 1990, 129, 85–92. [Google Scholar] [CrossRef]
- Murdoch, S.L.; Trunk, K.; English, G.; Fritsch, M.J.; Pourkarimi, E.; Coulthurst, S.J. The opportunistic pathogen Serratia marcescens utilises Type VI secretion to target bacterial competitors. J. Bacteriol. 2011, 193, 6057–6069. [Google Scholar] [CrossRef] [Green Version]
- Gordee, R.S.; Matthews, T.R. Systemic antifungal activity of pyrrolnitrin. Appl. Microbiol. 1969, 17, 690–694. [Google Scholar] [CrossRef] [PubMed]
- El-Banna, N.; Winkelmann, G. Pyrrolnitrin from Burkholderia cepacia: Antibiotic activity against fungi and novel activities against streptomycetes. J. Appl. Microbiol. 1998, 85, 69–78. [Google Scholar] [CrossRef]
- Chernin, L.; Brandis, A.; Ismailov, Z.; Chet, I. Pyrrolnitrin production by an Enterobacter agglomerans strain with a broad spectrum of antagonistic activity towards fungal and bacterial phytopathogens. Curr. Microbiol. 1996, 32, 208–212. [Google Scholar] [CrossRef]
- van Pée, K.-H. Chapter 2—Biosynthesis of halogenated alkaloids. In The Alkaloids: Chemistry and Biology, 1st ed.; Knölker, H.-J., Ed.; Academic Press: Oxford, UK, 2012; Volume 71, pp. 167–210. [Google Scholar] [CrossRef]
- Hu, D.X.; Withall, D.M.; Challis, G.L.; Thomson, R.J. Structure, chemical synthesis, and biosynthesis of prodiginine natural products. Chem. Rev. 2016, 116, 7818–7853. [Google Scholar] [CrossRef]
- Bennett, J.W.; Bentley, R. Seeing red: The story of prodigiosin. Adv. Appl. Microbiol. 2000, 47, 1–32. [Google Scholar] [CrossRef]
- Williamson, N.R.; Fineran, P.C.; Gristwood, T.; Leeper, F.J.; Salmond, G.P. The biosynthesis and regulation of bacterial prodiginines. Nat. Rev. Microbiol. 2006, 4, 887–899. [Google Scholar] [CrossRef]
- Roy, P.; Ahmed, N.H.; Grover, R.K. Non-pigmented strain of Serratia marcescens: An unusual pathogen causing pulmonary infection in a patient with malignancy. J. Clin. Diagn. Res. 2014, 8, DD05–DD06. [Google Scholar] [CrossRef]
- Ordentlich, A.; Elad, Y.; Chet, I. The role of chitinase of Serratia marcescens in biocontrol of Sclerotium rolfsii. Phytopathology 1988, 78, 84–87. [Google Scholar]
- Wang, K.; Yan, P.; Cao, L. Chitinase from a novel strain of Serratia marcescens JPP1 for biocontrol of aflatoxin: Molecular characterization and production optimization using response surface methodology. BioMed Res. Int. 2014, 2014, 482623. [Google Scholar] [CrossRef] [PubMed]
- Baráti-Deák, B.; Mohácsi-Farkas, C.; Belák, Á. Searching for antagonistic activity of bacterial isolates derived from food processing environments on some food-borne pathogenic bacteria. Acta Aliment. 2020, 49, 415–423. [Google Scholar] [CrossRef]
- Leisner, J.J.; Vogensen, F.K.; Kollmann, J.; Aideh, B.; Vandamme, P.; Vancanneyt, M.; Ingmer, H. α-Chitinase activity among lactic acid bacteria. Syst. Appl. Microbiol. 2008, 31, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-L.; Lan, C.-Y.; Fu, C.-C.; Juang, R.-S. Production of hexaoligochitin from colloidal chitin using a chitinase from Aeromonas schubertii. Int. J. Biol. Macromol. 2014, 69, 59–63. [Google Scholar] [CrossRef]
- Bharmal, M.H.; Jahagirdar, N.; Aruna, K. Study on optimization of prodigiosin production by Serratia marcescens MSK1 isolated from air. Int. J. Adv. Biol. Res. 2012, 2, 671–680. [Google Scholar]
- Harris, K.P.; Williamson, R.; Slater, H.; Cox, A.; Abbasi, S.; Foulds, I.; Simonsen, T.; Leeper, J.; Salmond, P.C. The Serratia gene cluster encoding biosynthesis of the red antibiotic, prodigiosin, shows species and strain dependent genome context variation. Microbiology 2004, 150, 3547–3560. [Google Scholar] [CrossRef]
- Ramaiah, N.; Hill, R.T.; Chun, J.; Ravel, J.; Matte, M.H.; Straube, W.L.; Colwell, R.R. Use of a chiA probe for detection of chitinase genes in bacteria from the Chesapeake Bay. FEMS Microbiol. Ecol. 2000, 34, 63–71. [Google Scholar] [CrossRef]
- Rastogi, V.; Purohit, P.; Peters, B.P.; Nirwan, P.S. Pulmonary infection with Serratia marcescens. Indian J. Med. Microbiol. 2002, 20, 167–168. [Google Scholar] [CrossRef]
- Mřretrř, T.; Langsrud, S. Residential bacteria on surfaces in the food industry and their implications for food safety and quality. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1022–1041. [Google Scholar] [CrossRef]
- Amorim, A.M.B.; Melo, D.H.; Souza, B.V.; Medeiros, L.M.; Mattoso, J.M.V.; Nascimento, J.S. A reddish problem: Antibiotic-resistant Serratia marcescens in dairy food commercialized in Rio de Janeiro. Int. Food Res. J. 2018, 25, 880–883. [Google Scholar]
- Cherif-Antar, A.; Moussa–Boudjemâa, B.; Didouh, N.; Medjahdi, K.; Mayo, B.; Flóre, A.B. Diversity and biofilm-forming capability of bacteria recovered from stainless steel pipes of a milk-processing dairy plant. Dairy Sci. Technol. 2016, 96, 27–38. [Google Scholar] [CrossRef]
- Darshan, N.; Manonmani, H.K. Prodigiosin and its potential applications. J. Food Sci. Technol. 2015, 52, 5393–5407. [Google Scholar] [CrossRef] [PubMed]
- Lapenda, J.C.; Silva, P.A.; Vicalvi, M.; Sena, K.X.F.R.; Nascimento, S.C. Antimicrobial activity of prodigiosin isolated from Serratia marcescens UFPEDA 398. World J. Microbiol. Biotechnol. 2014, 31, 399–406. [Google Scholar] [CrossRef]
- Venil, C.K.; Velmurugan, P.; Lakshmanaperumalsamy, P. Genomic environment of cueR and copA genes for prodigiosin biosynthesis by Serratia marcescens SB08. Rom. Biotechnol. Lett. 2009, 14, 4812–4819. [Google Scholar]
- Vaaje-Kolstad, G.; Horn, S.J.; Sørlie, M.; Eijsink, V.G.H. The chitinolytic machinery of Serratia marcescens—A model system for enzymatic degradation of recalcitrant polysaccharides. FEBS J. 2013, 280, 3028–3049. [Google Scholar] [CrossRef]
- Karthik, N.; Binod, P.; Pandey, A. Chitinases. In Current Developments in Biotechnology and Bioengineering: Production, Isolation and Purification of Industrial Products; Pandey, A., Negi, A., Soccol, C.R., Eds.; Elsevier: Oxford, UK, 2017; Chapter 15; pp. 335–368. [Google Scholar] [CrossRef]
- de Medeiros, S.C.; Monteiro-Júnior, J.E.; Sales, G.W.P.; Grangeiro, T.B.; Pinto Nogueira, N.A. Chitinases as antibacterial proteins: A systematic review. J. Young Pharm. 2018, 10, 144–148. [Google Scholar] [CrossRef]
- Tariq, A.L.; Reyaz, A.L. Characterization of an indigenous Serratia marcescens strain TRL isolated from fish market soil and cloning of its chiA gene. World J. Fish Mar. Sci. 2015, 7, 428–434. [Google Scholar] [CrossRef]
- Watanabe, T.; Kimura, K.; Sumiya, T.; Nikaidou, N.; Suzuki, K.; Suzuki, M.; Taiyoji, M.; Ferrer, S.; Regue, M. Genetic analysis of the chitinase system of Serratia marcescens 2170. J. Bacteriol. 1997, 179, 7111–7117. [Google Scholar] [CrossRef]
- Brurberg, M.B.; Eijsink, V.G.H.; Nes, I.F. Characterization of a chitinase gene (ch/A) from Serratia marcescens BJL200 and one-step purification of the gene product. FEMS Microbiol. Lett. 1994, 124, 399–404. [Google Scholar] [CrossRef]
- Ekstrand, B. Antimicrobial factors in milk—A review. Food Biotechnol. 1989, 3, 105–126. [Google Scholar] [CrossRef]
- Someya, N.; Nakajima, M.; Watanabe, K.; Hibi, T.; Akutsu, K. Potential of Serratia marcescens strain B2 for biological control of rice sheath blight. Biocontrol Sci. Technol. 2005, 15, 105–109. [Google Scholar] [CrossRef]
- Ferreira, L.C.; Maul, J.E.; Viana, M.V.C.; de Sousa, T.J.; de Carvalho Azevedo, V.A.; Roberts, D.P.; de Souza, J.T. Complete genome sequence of the biocontrol agent Serratia marcescens strain N4-5 uncovers an assembly artefact. Braz. J. Microbiol. 2021, 52, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Trejo-López, J.A.; Rangel-Vargas, E.; Gómez-Aldapa, C.A.; Villagómez-Ibarra, J.R.; Falfán-Cortes, R.N.; Acevedo- Sandoval, O.A.; Castro-Rosas, J. Isolation and molecular identification of Serratia strains producing chitinases, glucanases, cellulases, and prodigiosin and determination of their antifungal effect against Colletotrichum siamense and Alternaria alternata in vitro and on mango fruit. Int. J. Plant Biol. 2022, 13, 281–297. [Google Scholar] [CrossRef]
- Tao, A.; Wang, T.; Pang, F.; Zheng, X.; Ayra-Pardo, C.; Huang, S.; Xu, R.; Liu, F.; Li, J.; Wei, Y.; et al. Characterization of a novel chitinolytic Serratia marcescens strain TC-1 with broad insecticidal spectrum. AMB Express 2022, 12, 100. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Region or Gene | Primer Name | Sequence of the Primer (5′–3′) | Size of the PCR Product (bp) | References |
|---|---|---|---|---|
| cueR-pigA | cueR | TCGTAAAAACGAATCGTC | 505 | [28] |
| PE1 | GCAAAACTCTGAGCGGATTCGC | |||
| pigN-copA | ab77 | GAAACACTTAACCTGACG | 244 | |
| PE2 | CGCAGTTCATGCAGGACAGC | |||
| chiA | chiFEMSF | GATATCGACTGGGAGTTCCC | 225 | [29] |
| chiFEMSR | CATAGAAGTCGTAGGTCATC |
| Tested Foodborne Bacterial Pathogens | Potential Inhibitory S. marcescens Strains | |
|---|---|---|
| CSM-RMT-1 | CSM-RMT-II-1 | |
| L. monocytogenes CCM 4699 | + | − |
| Sa. Hartford NCAIM B.01310 | (+) | − |
| Y. enterocolitica HNCMB 98002 | + | + |
| E. coli NCAIM B.01909 | + | − |
| Days of Incubation | Tested Temperatures | |||||||
|---|---|---|---|---|---|---|---|---|
| 5 °C | 10 °C | 15 °C | 20 °C | 25 °C | 30 °C | 37 °C | 42 °C | |
| Inhibition of Sa. Hartford by S. marcescens CSM-RMT-1 | ||||||||
| 1 | − | − | − | − | − | − | − | − |
| 2 | − | − | − | − | − | − | − | − |
| 3 | − | (+) | − | − | − | − | − | − |
| 6 | − | − | − | − | − | − | − | − |
| Inhibition of E. coli by S. marcescens CSM−RMT−1 | ||||||||
| 1 | − | − | − | − | − | + | − | − |
| 2 | − | − | − | − | − | (+) | − | − |
| 3 | − | − | − | − | − | (+) | − | − |
| 6 | − | − | − | − | − | (+) | − | − |
| Inhibition of L. monocytogenes by S. marcescens CSM-RMT-1 | ||||||||
| 1 | − | − | − | (+) | (+) | − | − | − |
| 2 | − | − | + | (+) | (+) | (+) | − | − |
| 3 | − | (+) | + | (+) | (+) | (+) | − | − |
| 6 | − | (+) | + | (+) | − | (+) | − | − |
| Inhibition of Y. enterocolitica by S. marcescens CSM−RMT−1 | ||||||||
| 1 | − | − | − | − | − | + | − | − |
| 2 | − | − | − | + | + | + | − | − |
| 3 | − | − | − | + | + | + | − | − |
| 6 | − | − | + | + | + | + | − | − |
| Time | Temp. | Mean | Std. Deviation | 24 h | 48 h | 120 h | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 20 °C | 25 °C | 30 °C | 20 °C | 25 °C | 30 °C | 20 °C | 25 °C | 30 °C | ||||
| S. marcescens CSM-RMT-1 | ||||||||||||
| 24 h | 20 °C | 4.333 | 0.5774 | 0.951 | 0.319 | 0.173 | 0.086 | 0.004 | 0.000 | 0.004 | 0.001 | |
| 25 °C | 4.000 | 0.0000 | 0.168 | 0.038 | 0.024 | 0.018 | 0.005 | 0.014 | 0.001 | |||
| 30 °C | 5.667 | 0.5774 | 1.000 | 0.699 | 0.013 | 0.000 | 0.006 | 0.001 | ||||
| 48 h | 20 °C | 5.833 | 0.2887 | 0.553 | 0.019 | 0.001 | 0.013 | 0.000 | ||||
| 25 °C | 6.333 | 0.2887 | 0.029 | 0.002 | 0.015 | 0.000 | ||||||
| 30 °C | 9.333 | 0.5774 | 0.003 | 0.019 | 0.003 | |||||||
| 120 h | 20 °C | 14.667 | 0.5774 | 0.533 | 0.135 | |||||||
| 25°C | 16.333 | 1.1547 | 1.000 | |||||||||
| 30 °C | 16.333 | 0.2887 | ||||||||||
| S. marcescens CSM-RMT-II-1 | ||||||||||||
| 24 h | 20 °C | 4.000 | 0.0000 | 0.951 | 1.000 | 0.091 | 0.038 | 0.004 | 0.001 | 0.008 | 0.008 | |
| 25 °C | 3.500 | 0.8660 | 1.000 | 0.042 | 0.013 | 0.019 | 0.004 | 0.001 | 0.000 | |||
| 30 °C | 3.833 | 1.4434 | 0.183 | 0.084 | 0.083 | 0.021 | 0.005 | 0.005 | ||||
| 48 h | 20 °C | 7.500 | 0.8660 | 0.604 | 0.198 | 0.011 | 0.003 | 0.002 | ||||
| 25 °C | 8.833 | 0.7638 | 0.710 | 0.012 | 0.004 | 0.003 | ||||||
| 30 °C | 9.667 | 0.2887 | 0.000 | 0.018 | 0.015 | |||||||
| 120 h | 20 °C | 15.167 | 0.2887 | 0.780 | 0.463 | |||||||
| 25 °C | 16.000 | 0.8660 | 0.995 | |||||||||
| 30 °C | 16.500 | 0.8660 | ||||||||||
| Sample | Sampling Time | Mean | Std. Deviation | Salmonella Control | Salmonella 1:1 | Salmonella 1:1000 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Day 0 | Day 1 | Day 2 | Day 3 | Day 6 | Day 0 | Day 1 | Day 2 | Day 3 | Day 6 | Day 0 | Day 1 | Day 2 | Day 3 | Day 6 | ||||
| Co-culturing in TSB | ||||||||||||||||||
| Salmonella control | Day 0 | 1.666 | 0.0205 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| Day 1 | 8.528 | 0.0254 | 0.000 | 0.000 | 0.214 | 0.000 | 0.000 | 1.000 | 0.041 | 0.039 | 0.000 | 0.000 | 0.000 | 0.000 | 0.048 | |||
| Day 2 | 8.897 | 0.0311 | 0.968 | 0.460 | 0.000 | 0.000 | 0.000 | 0.020 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | ||||
| Day 3 | 8.991 | 0.0188 | 0.006 | 0.000 | 0.000 | 0.000 | 0.017 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |||||
| Day 6 | 9.185 | 0.0738 | 0.000 | 0.000 | 0.080 | 1.000 | 0.057 | 0.000 | 0.000 | 0.000 | 0.000 | 0.051 | ||||||
| Salmonella 1:1 | Day 0 | 0.391 | 0.0086 | 0.000 | 0.000 | 0.000 | 0.000 | 0.947 | 0.000 | 0.000 | 0.000 | 0.000 | ||||||
| Day 1 | 6.389 | 0.1245 | 0.000 | 0.000 | 0.005 | 0.000 | 0.000 | 0.101 | 0.018 | 0.000 | ||||||||
| Day 2 | 7.135 | 0.1336 | 0.028 | 0.037 | 0.000 | 0.000 | 0.000 | 0.042 | 0.020 | |||||||||
| Day 3 | 7.056 | 0.0805 | 0.110 | 0.000 | 0.000 | 0.000 | 0.000 | 0.120 | ||||||||||
| Day 6 | 8.147 | 0.1022 | 0.000 | 0.000 | 0.000 | 0.000 | 0.929 | |||||||||||
| Salmonella 1:1000 | Day 0 | 0.473 | 0.0016 | 0.000 | 0.000 | 0.000 | 0.000 | |||||||||||
| Day 1 | 5.000 | 1.4142 | 0.007 | 0.000 | 0.000 | |||||||||||||
| Day 2 | 7.223 | 0.1095 | 0.000 | 0.000 | ||||||||||||||
| Day 3 | 7.238 | 0.3373 | 0.091 | |||||||||||||||
| Day 6 | 8.451 | 0.2128 | ||||||||||||||||
| Co-culturing in Milk | ||||||||||||||||||
| Salmonella control | Day 0 | 1.693 | 0.0296 | 0.000 | 0.000 | 0.000 | 0.000 | 0.009 | 0.000 | 0.000 | 0.000 | 0.000 | 0.008 | 0.000 | 0.000 | 0.000 | 0.000 | |
| Day 1 | 8.771 | 0.0059 | 0.999 | 0.993 | 0.902 | 0.000 | 0.003 | 0.003 | 0.024 | 0.049 | 0.000 | 0.000 | 0.039 | 0.041 | 0.049 | |||
| Day 2 | 9.501 | 0.0386 | 1.000 | 1.000 | 0.000 | 0.001 | 0.020 | 0.014 | 0.480 | 0.000 | 0.000 | 0.030 | 0.032 | 0.048 | ||||
| Day 3 | 9.667 | 0.0198 | 1.000 | 0.000 | 0.000 | 0.013 | 0.009 | 0.049 | 0.000 | 0.000 | 0.020 | 0.021 | 0.049 | |||||
| Day 6 | 9.069 | 0.0574 | 0.000 | 0.000 | 0.005 | 0.004 | 0.041 | 0.000 | 0.000 | 0.008 | 0.009 | 0.039 | ||||||
| Salmonella 1:1 | Day 0 | 0.652 | 0.0287 | 0.000 | 0.000 | 0.000 | 0.000 | 0.904 | 0.000 | 0.000 | 0.000 | 0.000 | ||||||
| Day 1 | 7.801 | 0.1443 | 0.817 | 0.901 | 0.045 | 0.000 | 0.341 | 0.699 | 0.677 | 0.015 | ||||||||
| Day 2 | 8.642 | 0.1388 | 1.000 | 0.441 | 0.000 | 0.004 | 0.695 | 1.000 | 0.145 | |||||||||
| Day 3 | 9.138 | 0.0490 | 0.341 | 0.000 | 0.005 | 1.000 | 1.000 | 0.104 | ||||||||||
| Day 6 | 8.404 | 0.3234 | 0.000 | 0.000 | 0.566 | 0.588 | 0.906 | |||||||||||
| Salmonella 1:1000 | Day 0 | 0.474 | 0.0248 | 0.000 | 0.000 | 0.000 | 0.000 | |||||||||||
| Day 1 | 6.801 | 0.1443 | 0.002 | 0.002 | 0.000 | |||||||||||||
| Day 2 | 7.389 | 0.1245 | 1.000 | 0.206 | ||||||||||||||
| Day 3 | 8.172 | 0.0823 | 0.218 | |||||||||||||||
| Day 6 | 8.590 | 0.0157 | ||||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baráti-Deák, B.; Da Costa Arruda, G.C.; Perjéssy, J.; Klupács, A.; Zalán, Z.; Mohácsi-Farkas, C.; Belák, Á. Inhibition of Foodborne Pathogenic Bacteria by Excreted Metabolites of Serratia marcescens Strains Isolated from a Dairy-Producing Environment. Microorganisms 2023, 11, 403. https://doi.org/10.3390/microorganisms11020403
Baráti-Deák B, Da Costa Arruda GC, Perjéssy J, Klupács A, Zalán Z, Mohácsi-Farkas C, Belák Á. Inhibition of Foodborne Pathogenic Bacteria by Excreted Metabolites of Serratia marcescens Strains Isolated from a Dairy-Producing Environment. Microorganisms. 2023; 11(2):403. https://doi.org/10.3390/microorganisms11020403
Chicago/Turabian StyleBaráti-Deák, Bernadett, Giseli Cristina Da Costa Arruda, Judit Perjéssy, Adél Klupács, Zsolt Zalán, Csilla Mohácsi-Farkas, and Ágnes Belák. 2023. "Inhibition of Foodborne Pathogenic Bacteria by Excreted Metabolites of Serratia marcescens Strains Isolated from a Dairy-Producing Environment" Microorganisms 11, no. 2: 403. https://doi.org/10.3390/microorganisms11020403