
Candida albicans Strains Adapted to Caspofungin Due to Aneuploidy Become Highly Tolerant under Continued Drug Pressure

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media and Growth Conditions
2.2. Broth Microdilution Assay to Determine Minimum Inhibitory Concentration (MIC)
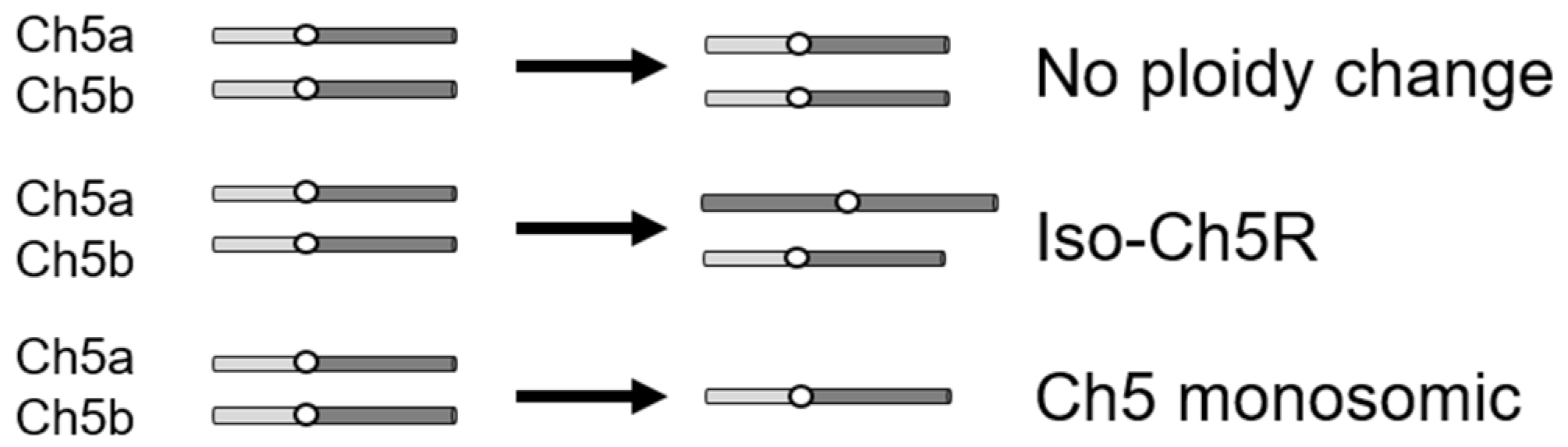
2.3. In Vitro Evolution of Strains
2.4. Sequencing of FKS1 Hot Spots HS1 and HS2 Regions
3. Results and Discussion
3.1. Caspofungin-Adapted Parental Mutants Have Decreased Susceptibility to Caspofungin Only
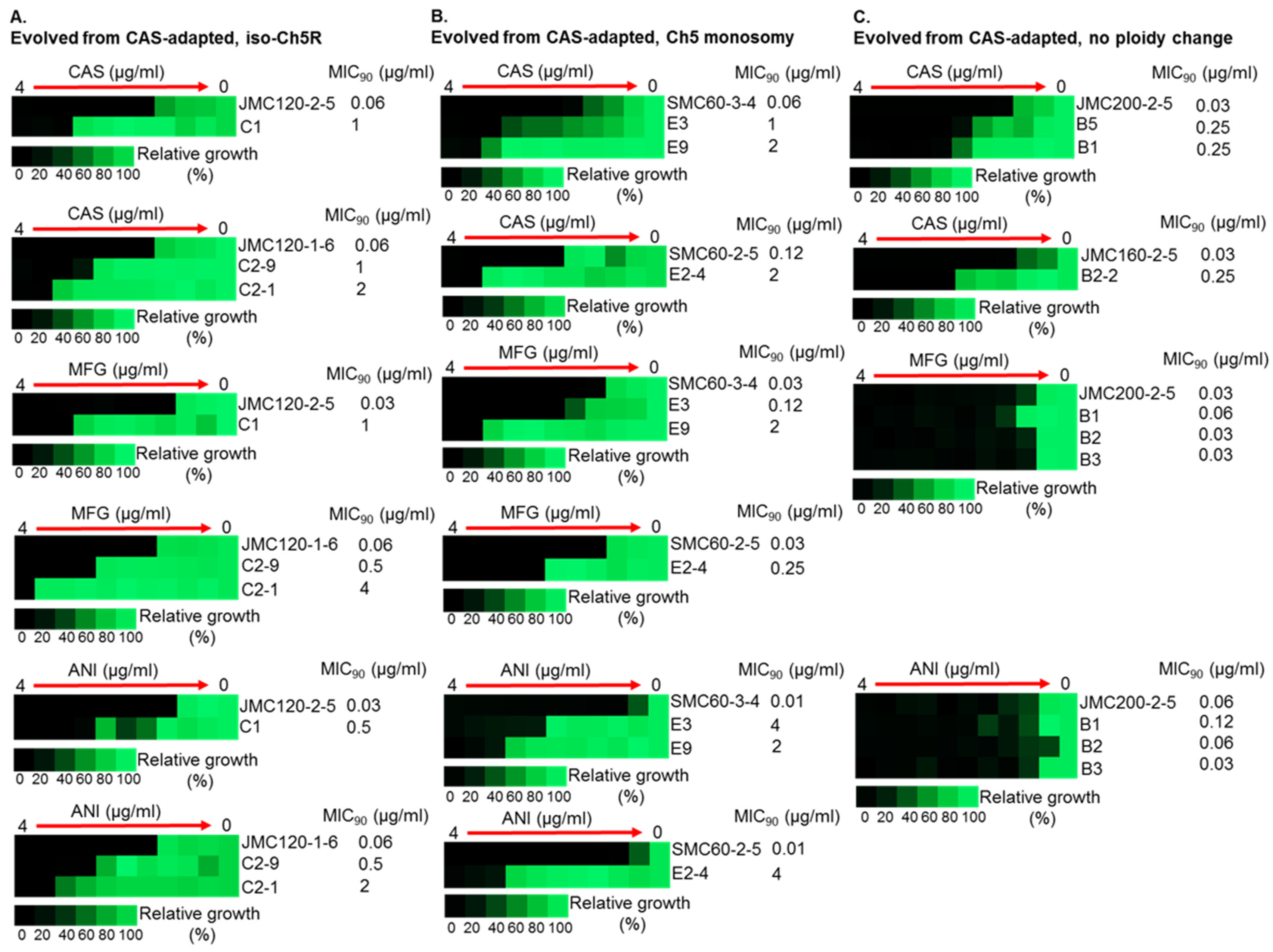
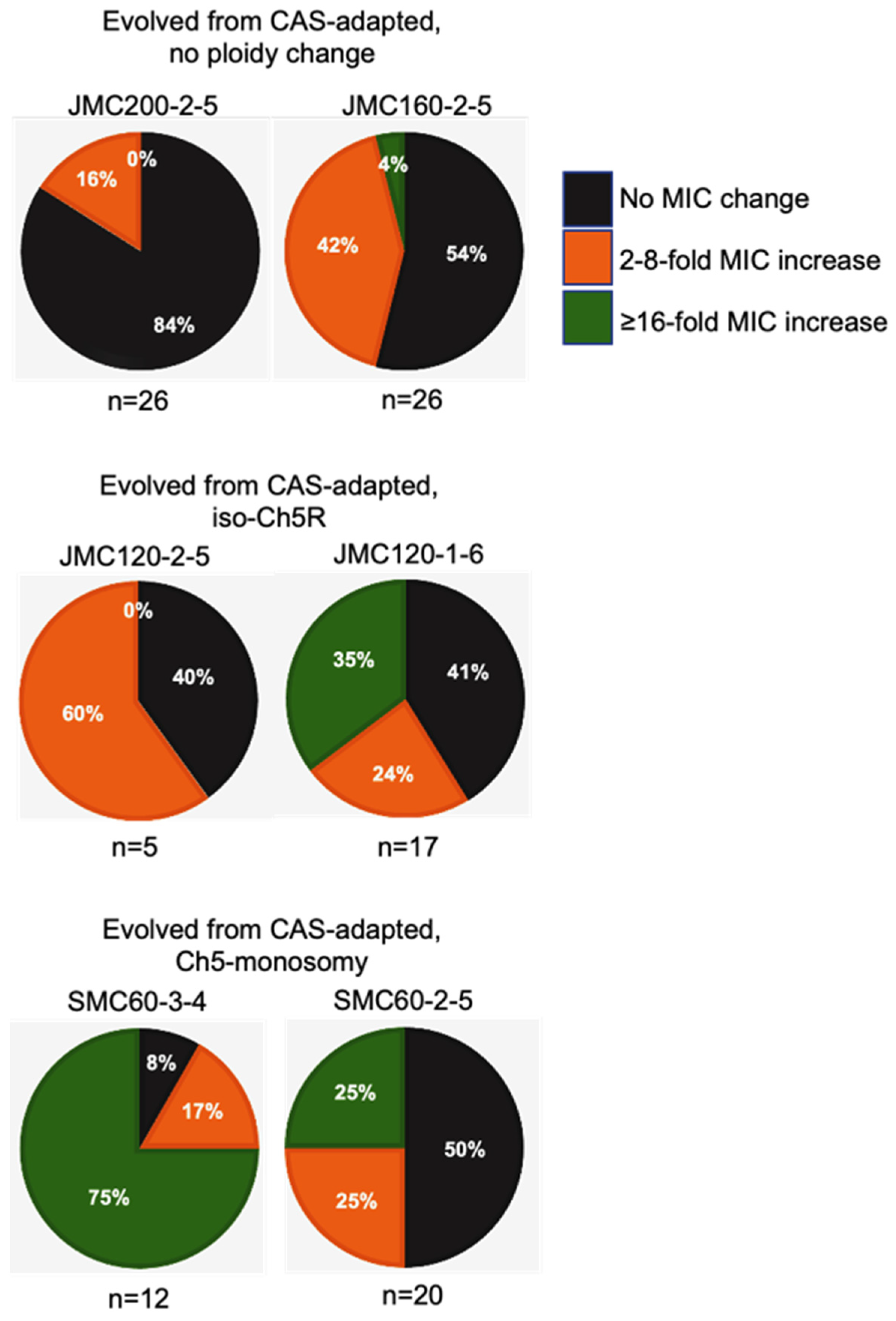
3.2. Generation and Properties of Strains That Evolved from Caspofungin-Adapted Mutants
3.3. Medical Importance
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heredia, M.Y.; Gunasekaran, D.; Ikeh, M.A.C.; Nobile, C.J.; Rauceo, J.M. Transcriptional regulation of the caspofungin-induced cell wall damage response in Candida albicans. Curr. Genet. 2020, 66, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Puumala, E.; Robbins, N.; Cowen, L.E. Antifungal drug resistance: Molecular mechanisms in Candida albicans and beyond. Chem. Rev. 2021, 121, 3390–3411. [Google Scholar] [CrossRef]
- Pristov, K.E.; Ghannoum, M.A. Resistance of Candida to azoles and echinocandins worldwide. Clin. Microbiol. Infect. 2019, 25, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Robbins, N.; Cowen, L.E. Antifungal drug resistance: Deciphering the mechanisms governing multidrug resistance in the fungal pathogen Candida glabrata. Curr. Biol. 2021, 31, R1520–R1523. [Google Scholar] [CrossRef] [PubMed]
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole antifungal resistance in Candida albicans and emerging non-albicans Candida species. Front. Microbiol. 2017, 7, 2173. [Google Scholar] [CrossRef] [Green Version]
- Perlin, D.S. Mechanisms of echinocandin antifungal drug resistance: Mechanism of echinocandin drug resistance. Ann. N.Y. Acad. Sci. 2015, 1354, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Kelly, R.; Kahn, J.N.; Robles, J.; Hsu, M.-J.; Register, E.; Li, W.; Vyas, V.; Fan, H.; Abruzzo, G.; et al. Specific substitutions in the echinocandin target Fks1p account for reduced susceptibility of rare laboratory and clinical Candida sp. isolates. Antimicrob. Agents. Chemother. 2005, 49, 3264–3273. [Google Scholar] [CrossRef] [Green Version]
- Espinel-Ingroff, A.; Arendrup, M.; Cantón, E.; Cordoba, S.; Dannaoui, E.; García-Rodríguez, J.; Gonzalez, G.M.; Govender, N.P.; Martin-Mazuelos, E.; Lackner, M.; et al. Multicenter study of method-dependent epidemiological cutoff values for detection of resistance in Candida spp. and Aspergillus spp. to amphotericin B and echinocandins for the Etest agar diffusion method. Antimicrob. Agents. Chemother. 2017, 61, e01792-16. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, E.; Boyken, L.; Tendolkar, S.; McDanel, J.; Castanheira, M.; Pfaller, M.; Diekema, D. Candidemia surveillance in Iowa: Emergence of echinocandin resistance. Diagn. Microbiol. Infect Dis. 2014, 79, 205–208. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Messer, S.A.; Diekema, D.J.; Jones, R.N.; Castanheira, M. Use of micafungin as a surrogate marker to predict susceptibility and resistance to caspofungin among 3764 clinical isolates of Candida by use of CLSI methods and interpretive criteria. J. Clin. Microbiol. 2014, 52, 108–114. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Messer, S.A.; Woosley, L.N.; Jones, R.N.; Castanheira, M. Echinocandin and triazole antifungal susceptibility profiles for clinical opportunistic yeast and mold isolates collected from 2010 to 2011: Application of new CLSI clinical breakpoints and epidemiological cutoff values for characterization of geographic and temporal trends of antifungal resistance. J. Clin. Microbiol. 2013, 51, 2571–2581. [Google Scholar] [PubMed] [Green Version]
- Sah, S.K.; Hayes, J.J.; Rustchenko, E. The role of aneuploidy in the emergence of echinocandin resistance in human fungal pathogen Candida albicans. PLoS Pathog. 2021, 17, e1009564. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, L.; Wakabayashi, H.; Myers, J.; Jiang, Y.; Cao, Y.; Jimenez-Ortigosa, C.; Perlin, D.S.; Rustchenko, E. Tolerance to caspofungin in Candida albicans is associated with at least three distinctive mechanisms that govern expression of FKS genes and cell wall remodeling. Antimicrob. Agents. Chemother. 2017, 61, e00071-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Healey, K.R.; Katiyar, S.K.; Raj, S.; Edlind, T.D. CRS-MIS in Candida glabrata: Sphingolipids modulate echinocandin-Fks interaction: Sphingolipids modulate echinocandin-Fks interaction. Mol. Microbiol. 2012, 86, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Kabir, M.A.; Kravets, A.; Andaluz, E.; Larriba, G.; Rustchenko, E. Chromosome instability and unusual features of some widely used strains of Candida albicans. Yeast 2008, 25, 433–448. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. M27-A3 Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts: Approved Standard, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]
- Locke, J.B.; Almaguer, A.L.; Zuill, D.E.; Bartizal, K. Characterization of in vitro resistance development to the novel echinocandin CD101 in Candida species. Antimicrob. Agents. Chemother. 2016, 60, 6100–6107. [Google Scholar] [CrossRef] [Green Version]
- Balashov, S.V.; Park, S.; Perlin, D.S. Assessing resistance to the echinocandin antifungal drug caspofungin in Candida albicans by profiling mutations in FKS1. Antimicrob. Agents. Chemother. 2006, 50, 2058–2063. [Google Scholar] [CrossRef] [Green Version]
- Heintz, N.; Gong, S. Small-scale preparations of yeast DNA. Cold. Spring. Harb. Protoc. 2020, 2020, pdb.prot098152. [Google Scholar] [CrossRef]
- Tóth, Z.; Forgács, L.; Kardos, T.; Kovács, R.; Locke, J.B.; Kardos, G.; Nagy, F.; Borman, A.M.; Adnan, A.; Majoros, L. Relative frequency of paradoxical growth and trailing effect with caspofungin, micafungin, anidulafungin, and the novel echinocandin rezafungin against Candida species. JoF 2020, 6, 136. [Google Scholar] [CrossRef]
- Li, H.; Cao, Y.; Yan, T.; Jiang, Y.; Yang, F. Aneuploidy underlies paradoxical growth of rezafungin and enables cross-tolerance to echinocandins in Candida albicans. J. Infect. 2022, 85, 702–769. [Google Scholar] [CrossRef]
- Healey, K.R.; Katiyar, S.K.; Castanheira, M.; Pfaller, M.A.; Edlind, T.D. Candida glabrata mutants demonstrating paradoxical reduced caspofungin susceptibility but increased micafungin susceptibility. Antimicrob. Agents. Chemother. 2011, 55, 3947–3949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, M.A.; Diekema, D.J.; Andes, D.; Arendrup, M.C.; Brown, S.D.; Lockhart, S.R.; Motyl, M.; Perlin, D.S. CLSI Subcommittee for antifungal Testing. Clinical breakpoints for the echinocandins and Candida revisited: Integration of molecular, clinical, and microbiological data to arrive at species-specific interpretive criteria. Drug Resist. Updat. 2011, 14, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Sah, S.K.; Bhattacharya, S.; Yadav, A.; Husain, F.; Ndiaye, A.B.K.T.; Kruppa, M.D.; Hayes, J.J.; Rustchenko, E. Multiple genes of Candida albicans influencing echinocandin susceptibility in caspofungin-adapted mutants. Antimicrob. Agents. Chemother. 2022, e00977-22. [Google Scholar] [CrossRef] [PubMed]
- Kravets, A.; Qin, H.; Ahmad, A.; Bethlendy, G.; Gao, Q.; Rustchenko, E. Widespread occurrence of dosage compensation in Candida albicans. PLoS ONE 2010, 5, e10856. [Google Scholar] [CrossRef] [Green Version]
- Wakabayashi, H.; Tucker, C.; Bethlendy, G.; Kravets, A.; Welle, S.L.; Bulger, M.; Hayes, J.J.; Rustchenko, E. NuA4 histone acetyltransferase activity is required for H4 acetylation on a dosage-compensated monosomic chromosome that confers resistance to fungal toxins. Epigenetics Chromatine 2017, 10, 49. [Google Scholar] [CrossRef] [Green Version]
- Gow, N.A.R.; Johnson, C.; Berman, J.; Coste, A.T.; Cuomo, C.A.; Perlin, D.S.; Bicanic, T.; Harrison, T.S.; Wiederhold, N.; Bromley, M.; et al. The importance of antimicrobial resistance in medical mycology. Nat. Commun. 2022, 13, 5352. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | CAS | MFG ** | ANI ** |
|---|---|---|---|
| No ploidy change | |||
| Parent JRCT1 JMC200-2-5 JMC160-2-5 | − + + | − + + | − + + |
| Ch5 monosomic | |||
| Parent SC5314 SMC60-3-4 SMC60-2-5 | − + + | − − − − − | − − − − − |
| Iso-Ch5R | |||
| Parent JRCT1 JMC120-1-6 JMC120-2-5 | − + + | − − − − − | − INC INC |
| D | CAS | MFG | ANI | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CAS-Adapted Parental Mutant * | 2X MIC Conc. (µg/mL) | No. of Recove-red Cols. | No. of Evolved Colonies—Increase of MICs | No. of Colonies W. 16- or 32-Fold Increase of MICs | % of Colonies w. 16- or 32-Fold Increase of MICs | No. of Evolved Colonies—Increase of MICs | No. of Colonies W. 16- or 32-Fold Increase of MICs | % of Colonies W. 16- or 32-Fold Increase of MICs | No. of Evolved Colonies—Increase of MICs | No. of Colonies W. 16- or 32-Fold Increase of MICs | % of Colonies W. 16- or 32-Fold Increase of MICs ** |
| JMC200-2-5 No ploidy change | 0.500 | 26 | 3–4-fold 1–8-fold | 0 | NA | 1–2-fold | 0 | NA | 1–2-fold | 0 | NA |
| JMC160-2-5 No ploidy change | 0.500 | 26 | 4–2-fold 5–4-fold 2–8-fold 1–128-fold | 1 | 3.8% | 0 | 0 | NA | 0 | 0 | NA |
| JMC120-2-5 iso-Ch5 | 0.125 | 5 | 3–8-fold | 0 | NA | 2–32-fold 1–128-fold | 3 | 60% | 2–16-fold 1–64-fold | 3 | 60% |
| JMC120-1-6 iso-Ch5R | 0.125 | 17 | 2–4-fold 2–8-fold 1–16-fold 3–32-fold 2–64-fold | 6 | 35% | 2–4-fold, 1–8-fold 1–16-fold 5–64-fold | 6 | 35% | 1–4-fold, 2–8-fold 3–16-fold 2–32-fold 1-64-fold | 6 | 35% |
| SMC60-3-4 Ch5 mono | 0.250 | 12 | 2–2-fold 3–16-fold 4–32-fold 2–64-fold | 9 | 75% | 1–4-fold 1–8-fold 4–16-fold 1–32-fold 1–64-fold 1–128-fold | 7 | 58.3% | 1–8-fold 3–16-fold 1–64-fold 3–128-fold 1–256-fold | 8 | 66.7% |
| SMC60-2-5 Ch5 mono | 0.250 | 20 | 2–2-fold 2–4-fold 1–8-fold 2–16-fold 3–32-fold | 5 | 25% | 1–8-fold 3–16-fold 1–32-fold 2–64-fold 2–128-fold | 8 | 40% | 2–4-fold 1–8-fold 1–32-fold 2–128-fold 3–256-fold | 6 | 30% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Husain, F.; Yadav, A.; Sah, S.K.; Hayes, J.J.; Rustchenko, E. Candida albicans Strains Adapted to Caspofungin Due to Aneuploidy Become Highly Tolerant under Continued Drug Pressure. Microorganisms 2023, 11, 23. https://doi.org/10.3390/microorganisms11010023
Husain F, Yadav A, Sah SK, Hayes JJ, Rustchenko E. Candida albicans Strains Adapted to Caspofungin Due to Aneuploidy Become Highly Tolerant under Continued Drug Pressure. Microorganisms. 2023; 11(1):23. https://doi.org/10.3390/microorganisms11010023
Chicago/Turabian StyleHusain, Farha, Anshuman Yadav, Sudisht K. Sah, Jeffrey J. Hayes, and Elena Rustchenko. 2023. "Candida albicans Strains Adapted to Caspofungin Due to Aneuploidy Become Highly Tolerant under Continued Drug Pressure" Microorganisms 11, no. 1: 23. https://doi.org/10.3390/microorganisms11010023