
Genome-Based Analysis of Virulence Factors and Biofilm Formation in Novel P. aeruginosa Strains Isolated from Household Appliances

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Phenotypic Investigations
2.2.1. Cultivation
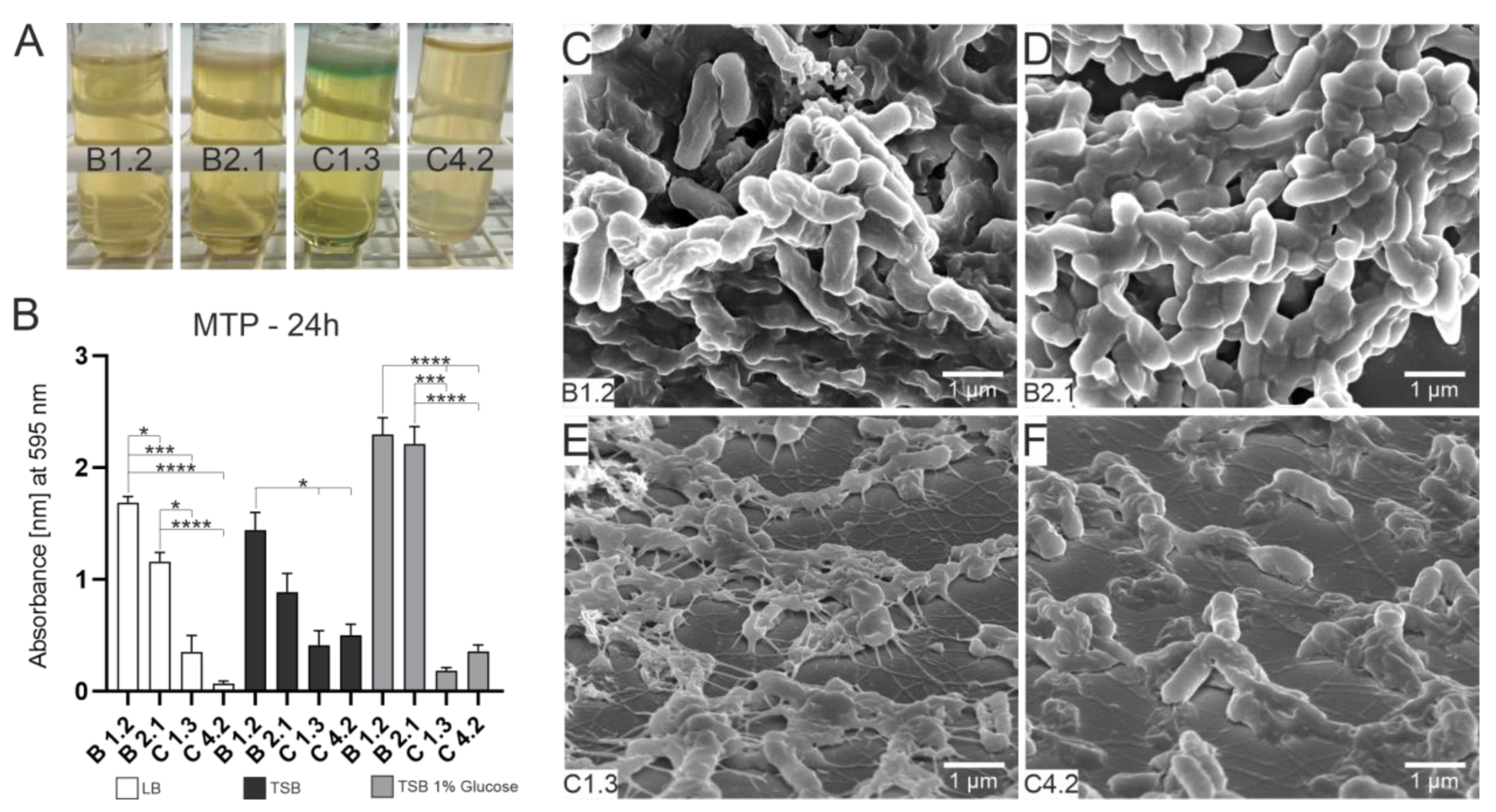
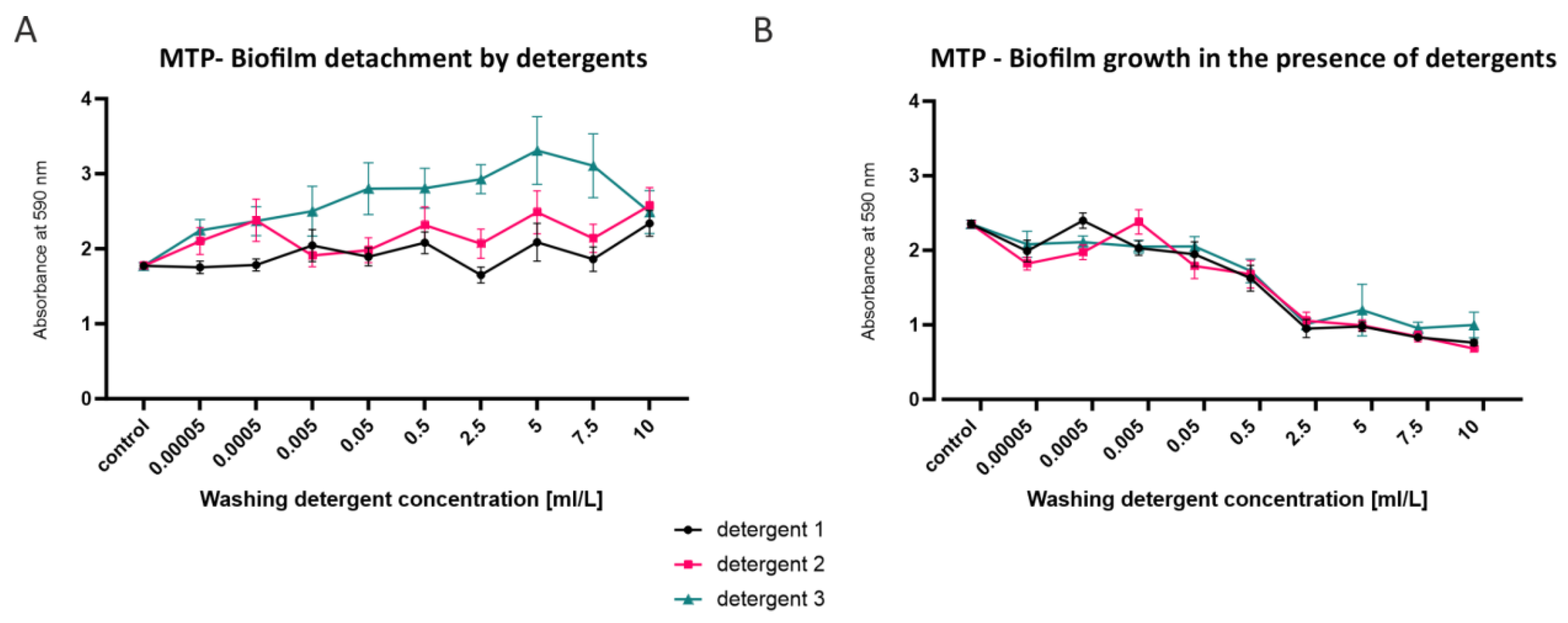
2.2.2. Microtiter Plate Assay (MTP)
2.2.3. SEM of Attached Cells and Biofilm Formation
2.3. Whole Genome Data Analysis
2.3.1. Whole Genome Sequencing and Genome Assembly
2.3.2. Nanopore Library Preparation and GridION Sequencing
2.3.3. Illumina Library Preparation and MiSeq Sequencing
2.3.4. Assembly and Annotation
2.3.5. Data Availability
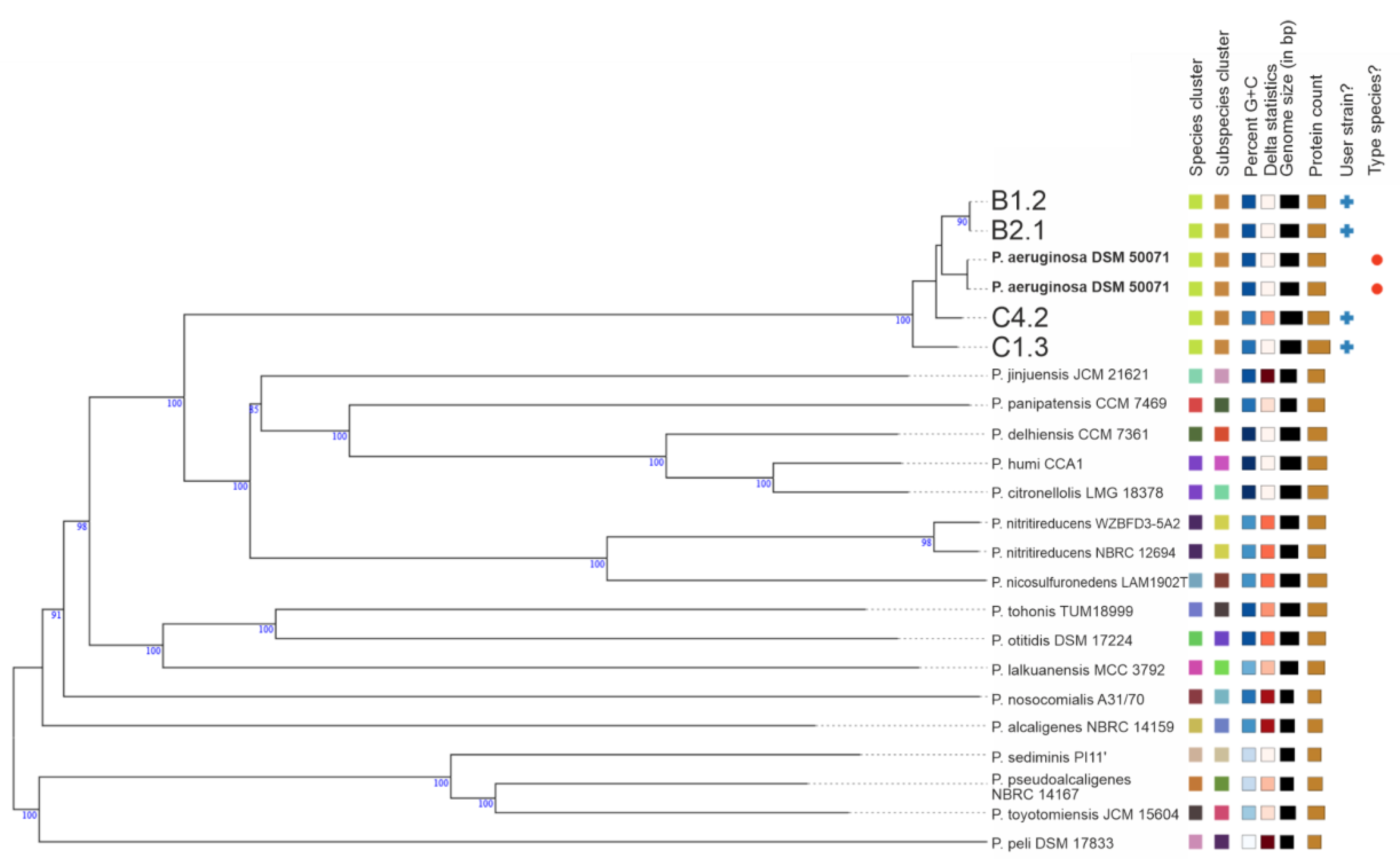
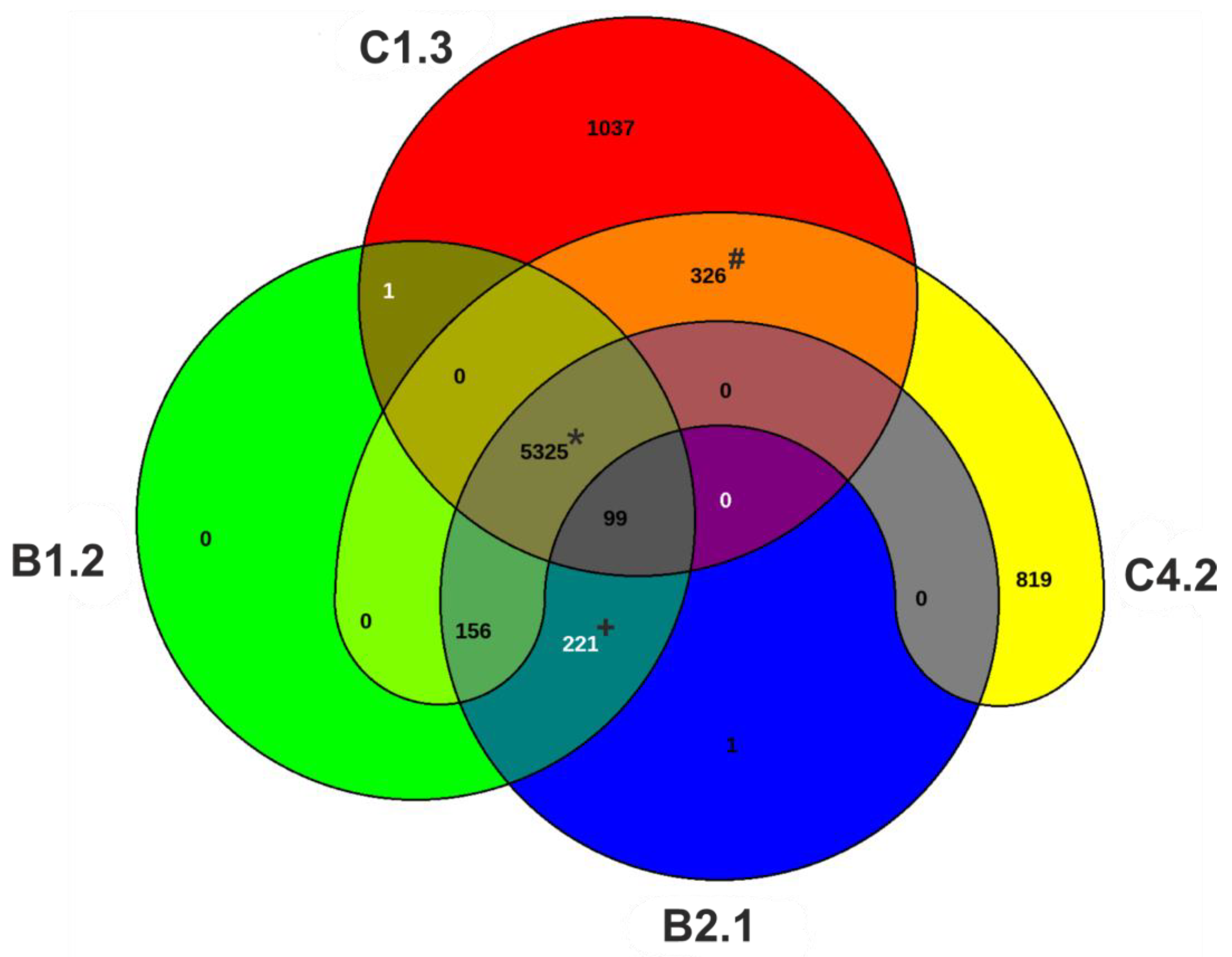
2.3.6. Phylogenetics, Pan-Genome, Typing and Prophage Investigation
2.3.7. Biofilm-Associated and Virulence Genes
2.3.8. Statistical Analysis
3. Results
3.1. Bacterial Growth, Attachment and Biofilm Formation
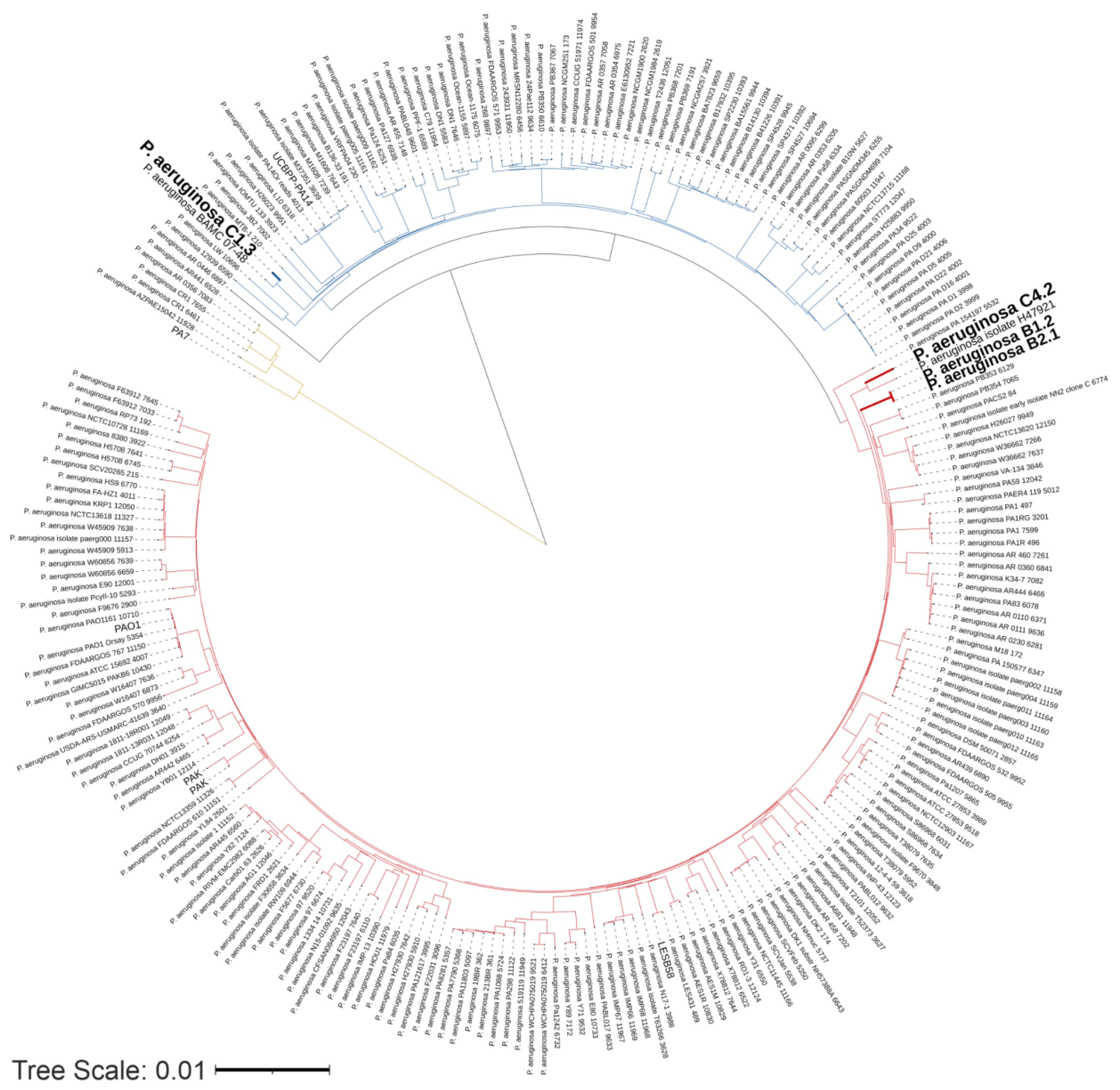
3.2. Genomic and Phylogenetic Features of P. aeruginosa WM Isolates
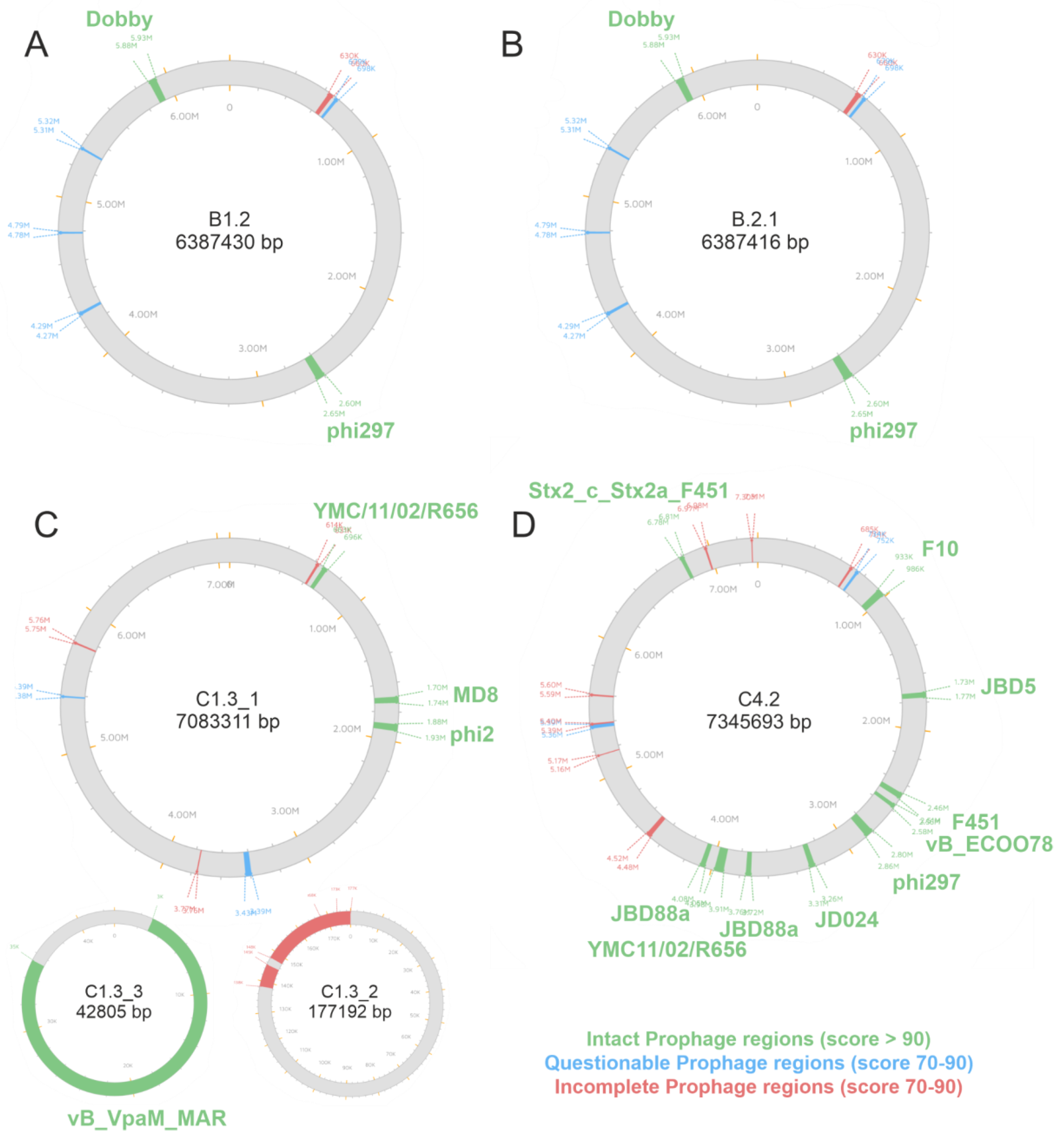
3.3. Identification of Prophages
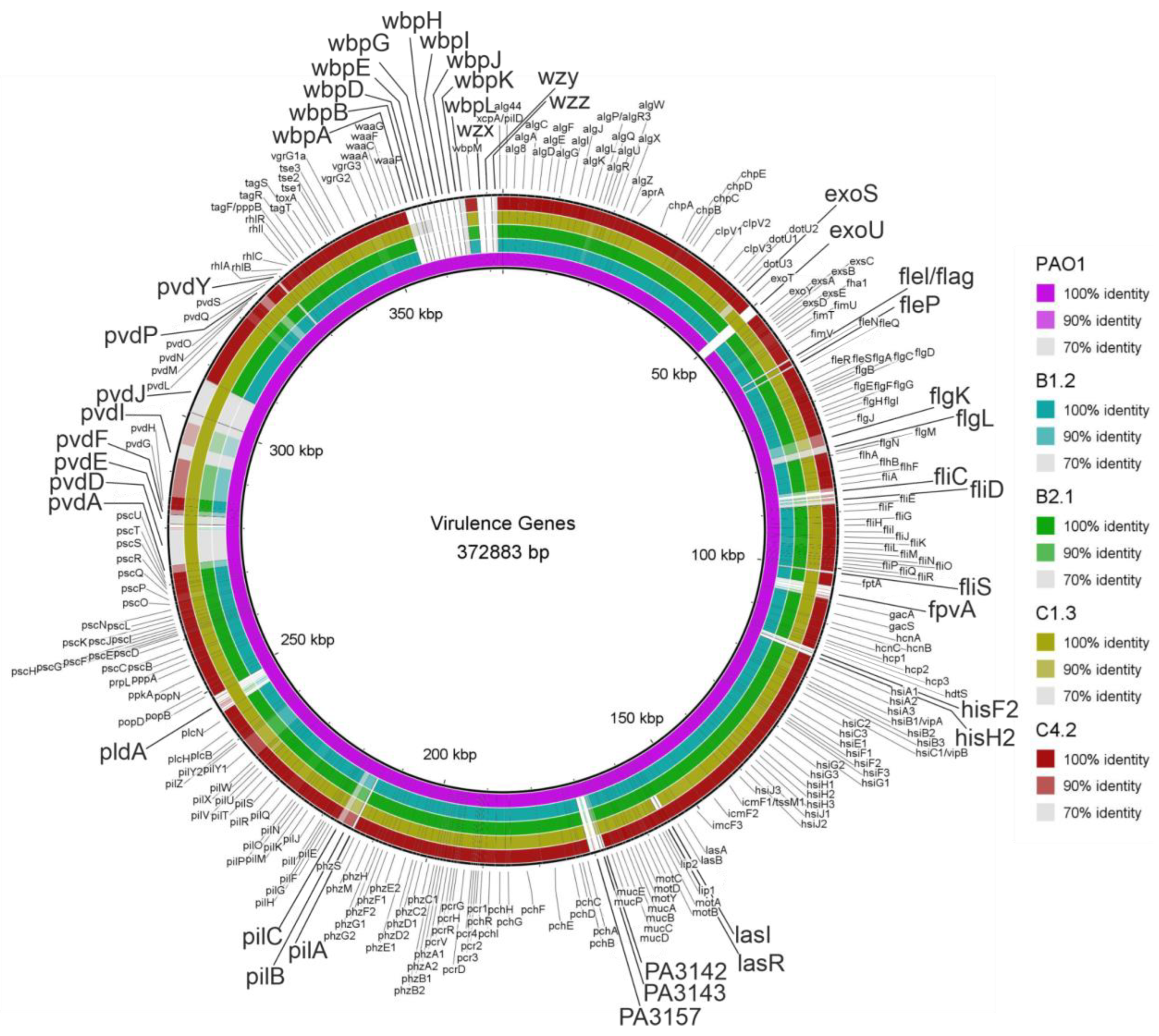
3.4. Virulence- and Biofilm-Associated Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.L.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete Genome Sequence of Pseudomonas Aeruginosa PAO1, an Opportunistic Pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callewaert, C.; Van Nevel, S.; Kerckhof, F.-M.; Granitsiotis, M.S.; Boon, N. Bacterial Exchange in Household Washing Machines. Front. Microbiol. 2015, 6, 1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gattlen, J.; Amberg, C.; Zinn, M.; Mauclaire, L. Biofilms Isolated from Washing Machines from Three Continents and Their Tolerance to a Standard Detergent. Biofouling 2010, 26, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Asghari, E.; Kiel, A.; Kaltschmidt, B.P.; Wortmann, M.; Schmidt, N.; Hüsgen, B.; Hütten, A.; Knabbe, C.; Kaltschmidt, C.; Kaltschmidt, B. Identification of Microorganisms from Several Surfaces by MALDI-TOF MS: P. Aeruginosa Is Leading in Biofilm Formation. Microorganisms 2021, 9, 992. [Google Scholar] [CrossRef]
- Jacksch, S.; Kaiser, D.; Weis, S.; Weide, M.; Ratering, S.; Schnell, S.; Egert, M. Influence of Sampling Site and Other Environmental Factors on the Bacterial Community Composition of Domestic Washing Machines. Microorganisms 2019, 8, 30. [Google Scholar] [CrossRef] [Green Version]
- Arai, H. Regulation and Function of Versatile Aerobic and Anaerobic Respiratory Metabolism in Pseudomonas Aeruginosa. Front. Microbiol. 2011, 2, 103. [Google Scholar] [CrossRef] [Green Version]
- Kordes, A.; Grahl, N.; Koska, M.; Preusse, M.; Arce-Rodriguez, A.; Abraham, W.-R.; Kaever, V.; Häussler, S. Establishment of an Induced Memory Response in Pseudomonas Aeruginosa during Infection of a Eukaryotic Host. ISME J. 2019, 13, 2018–2030. [Google Scholar] [CrossRef]
- Mulcahy, L.R.; Isabella, V.M.; Lewis, K. Pseudomonas Aeruginosa Biofilms in Disease. Microb. Ecol. 2014, 68, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas Aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef]
- Høiby, N.; Ciofu, O.; Bjarnsholt, T. Pseudomonas Aeruginosa Biofilms in Cystic Fibrosis. Future Microbiol. 2010, 5, 1663–1674. [Google Scholar] [CrossRef]
- Winstanley, C.; O’Brien, S.; Brockhurst, M.A. Pseudomonas Aeruginosa Evolutionary Adaptation and Diversification in Cystic Fibrosis Chronic Lung Infections. Trends Microbiol. 2016, 24, 327–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thöming, J.G.; Tomasch, J.; Preusse, M.; Koska, M.; Grahl, N.; Pohl, S.; Willger, S.D.; Kaever, V.; Müsken, M.; Häussler, S. Parallel Evolutionary Paths to Produce More than One Pseudomonas Aeruginosa Biofilm Phenotype. Npj Biofilms Microbiomes 2020, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dötsch, A.; Schniederjans, M.; Khaledi, A.; Hornischer, K.; Schulz, S.; Bielecka, A.; Eckweiler, D.; Pohl, S.; Häussler, S. The Pseudomonas Aeruginosa Transcriptional Landscape Is Shaped by Environmental Heterogeneity and Genetic Variation. mBio 2015, 6, e00749-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, E.E.; Buckley, D.G.; Wu, Z.; Saenphimmachak, C.; Hoffman, L.R.; D’Argenio, D.A.; Miller, S.I.; Ramsey, B.W.; Speert, D.P.; Moskowitz, S.M.; et al. Genetic Adaptation by Pseudomonas Aeruginosa to the Airways of Cystic Fibrosis Patients. Proc. Natl. Acad. Sci. USA 2006, 103, 8487–8492. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.K.; Schaefer, A.L.; Parsek, M.R.; Moninger, T.O.; Welsh, M.J.; Greenberg, E.P. Quorum-Sensing Signals Indicate That Cystic Fibrosis Lungs Are Infected with Bacterial Biofilms. Nature 2000, 407, 762–764. [Google Scholar] [CrossRef] [PubMed]
- Parsek, M.R.; Singh, P.K. Bacterial Biofilms: An Emerging Link to Disease Pathogenesis. Annu. Rev. Microbiol. 2003, 57, 677–701. [Google Scholar] [CrossRef]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial Biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef]
- Moradali, M.F.; Ghods, S.; Rehm, B.H.A. Pseudomonas Aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef] [Green Version]
- Amiel, E.; Lovewell, R.R.; O’Toole, G.A.; Hogan, D.A.; Berwin, B. Pseudomonas Aeruginosa Evasion of Phagocytosis Is Mediated by Loss of Swimming Motility and Is Independent of Flagellum Expression. Infect. Immun. 2010, 78, 2937–2945. [Google Scholar] [CrossRef] [Green Version]
- Häussler, S.; Parsek, M.R. Biofilms 2009: New Perspectives at the Heart of Surface-Associated Microbial Communities. J. Bacteriol. 2010, 192, 2941–2949. [Google Scholar] [CrossRef]
- de Beer, D.; Stoodley, P.; Lewandowski, Z. Liquid Flow in Heterogeneous Biofilms. Biotechnol. Bioeng. 1994, 44, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Parsek, M.R.; Greenberg, E.P. Sociomicrobiology: The Connections between Quorum Sensing and Biofilms. Trends Microbiol. 2005, 13, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial Biofilms: A Common Cause of Persistent Infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as Complex Differentiated Communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costerton, J.W.; Cheng, K.J.; Geesey, G.G.; Ladd, T.I.; Nickel, J.C.; Dasgupta, M.; Marrie, T.J. Bacterial Biofilms in Nature and Disease. Annu. Rev. Microbiol. 1987, 41, 435–464. [Google Scholar] [CrossRef] [PubMed]
- Simões, M.; Simões, L.C.; Machado, I.; Pereira, M.O.; Vieira, M.J. Control of Flow-Generated Biofilms with Surfactants: Evidence of Resistance and Recovery. Food Bioprod. Process. 2006, 84, 338–345. [Google Scholar] [CrossRef] [Green Version]
- Stewart, P.S. Theoretical Aspects of Antibiotic Diffusion into Microbial Biofilms. Antimicrob. Agents Chemother. 1996, 40, 2517–2522. [Google Scholar] [CrossRef] [Green Version]
- Liao, C.; Huang, X.; Wang, Q.; Yao, D.; Lu, W. Virulence Factors of Pseudomonas Aeruginosa and Antivirulence Strategies to Combat Its Drug Resistance. Front. Cell. Infect. Microbiol. 2022, 12, 926758. [Google Scholar] [CrossRef]
- Salacha, R.; Kovačić, F.; Brochier-Armanet, C.; Wilhelm, S.; Tommassen, J.; Filloux, A.; Voulhoux, R.; Bleves, S. The Pseudomonas Aeruginosa Patatin-like Protein PlpD Is the Archetype of a Novel Type V Secretion System. Environ. Microbiol. 2010, 12, 1498–1512. [Google Scholar] [CrossRef]
- Ma, Y.-X.; Wang, C.-Y.; Li, Y.-Y.; Li, J.; Wan, Q.-Q.; Chen, J.-H.; Tay, F.R.; Niu, L.-N. Considerations and Caveats in Combating ESKAPE Pathogens against Nosocomial Infections. Adv. Sci. 2020, 7, 1901872. [Google Scholar] [CrossRef]
- Chen, L.; Zou, Y.; She, P.; Wu, Y. Composition, Function, and Regulation of T6SS in Pseudomonas Aeruginosa. Microbiol. Res. 2015, 172, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Gallique, M.; Bouteiller, M.; Merieau, A. The Type VI Secretion System: A Dynamic System for Bacterial Communication? Front. Microbiol. 2017, 8, 1454. [Google Scholar] [CrossRef] [PubMed]
- Basler, M.; Ho, B.T.; Mekalanos, J.J. Tit-for-Tat: Type VI Secretion System Counterattack during Bacterial Cell-Cell Interactions. Cell 2013, 152, 884–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, A.K.; Mende-Mueller, L.; Selzer, J.; Barbieri, J.T. Pseudomonas Aeruginosa Exoenzyme S, a Double ADP-Ribosyltransferase, Resembles Vertebrate Mono-ADP-Ribosyltransferases. J. Biol. Chem. 1999, 274, 9503–9508. [Google Scholar] [CrossRef] [Green Version]
- Krall, R.; Schmidt, G.; Aktories, K.; Barbieri, J.T. Pseudomonas Aeruginosa ExoT Is a Rho GTPase-Activating Protein. Infect. Immun. 2000, 68, 6066–6068. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Frank, D.W.; Hillard, C.J.; Feix, J.B.; Pankhaniya, R.R.; Moriyama, K.; Finck-Barbançon, V.; Buchaklian, A.; Lei, M.; Long, R.M.; et al. The Mechanism of Action of the Pseudomonas Aeruginosa-Encoded Type III Cytotoxin, ExoU. EMBO J. 2003, 22, 2959–2969. [Google Scholar] [CrossRef] [Green Version]
- Yahr, T.L.; Vallis, A.J.; Hancock, M.K.; Barbieri, J.T.; Frank, D.W. ExoY, an Adenylate Cyclase Secreted by the Pseudomonas Aeruginosa Type III System. Proc. Natl. Acad. Sci. USA 1998, 95, 13899–13904. [Google Scholar] [CrossRef] [Green Version]
- Rutherford, S.T.; Bassler, B.L. Bacterial Quorum Sensing: Its Role in Virulence and Possibilities for Its Control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef]
- Nanda, A.M.; Thormann, K.; Frunzke, J. Impact of Spontaneous Prophage Induction on the Fitness of Bacterial Populations and Host-Microbe Interactions. J. Bacteriol. 2015, 197, 410–419. [Google Scholar] [CrossRef] [Green Version]
- Canchaya, C.; Fournous, G.; Brüssow, H. The Impact of Prophages on Bacterial Chromosomes. Mol. Microbiol. 2004, 53, 9–18. [Google Scholar] [CrossRef]
- Dobrindt, U.; Hochhut, B.; Hentschel, U.; Hacker, J. Genomic Islands in Pathogenic and Environmental Microorganisms. Nat. Rev. Microbiol. 2004, 2, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Ambroa, A.; Blasco, L.; López-Causapé, C.; Trastoy, R.; Fernandez-García, L.; Bleriot, I.; Ponce-Alonso, M.; Pacios, O.; López, M.; Cantón, R.; et al. Temperate Bacteriophages (Prophages) in Pseudomonas Aeruginosa Isolates Belonging to the International Cystic Fibrosis Clone (CC274). Front. Microbiol. 2020, 11, 556706. [Google Scholar] [CrossRef] [PubMed]
- Rice, S.A.; Tan, C.H.; Mikkelsen, P.J.; Kung, V.; Woo, J.; Tay, M.; Hauser, A.; McDougald, D.; Webb, J.S.; Kjelleberg, S. The Biofilm Life Cycle and Virulence of Pseudomonas Aeruginosa Are Dependent on a Filamentous Prophage. ISME J. 2009, 3, 271–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Secor, P.R.; Burgener, E.B.; Kinnersley, M.; Jennings, L.K.; Roman-Cruz, V.; Popescu, M.; Van Belleghem, J.D.; Haddock, N.; Copeland, C.; Michaels, L.A.; et al. Pf Bacteriophage and Their Impact on Pseudomonas Virulence, Mammalian Immunity, and Chronic Infections. Front. Immunol. 2020, 11, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisht, K.; Moore, J.L.; Caprioli, R.M.; Skaar, E.P.; Wakeman, C.A. Impact of Temperature-Dependent Phage Expression on Pseudomonas Aeruginosa Biofilm Formation. Npj Biofilms Microbiomes 2021, 7, 1–9. [Google Scholar] [CrossRef]
- Livny, J.; Friedman, D.I. Characterizing Spontaneous Induction of Stx Encoding Phages Using a Selectable Reporter System. Mol. Microbiol. 2004, 51, 1691–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allesen-Holm, M.; Barken, K.B.; Yang, L.; Klausen, M.; Webb, J.S.; Kjelleberg, S.; Molin, S.; Givskov, M.; Tolker-Nielsen, T. A Characterization of DNA Release in Pseudomonas Aeruginosa Cultures and Biofilms. Mol. Microbiol. 2006, 59, 1114–1128. [Google Scholar] [CrossRef]
- Kirov, S.M.; Webb, J.S.; O’May, C.Y.; Reid, D.W.; Woo, J.K.K.; Rice, S.A.; Kjelleberg, S. Biofilm Differentiation and Dispersal in Mucoid Pseudomonas Aeruginosa Isolates from Patients with Cystic Fibrosis. Microbiology 2007, 153, 3264–3274. [Google Scholar] [CrossRef] [Green Version]
- Webb, J.S.; Thompson, L.S.; James, S.; Charlton, T.; Tolker-Nielsen, T.; Koch, B.; Givskov, M.; Kjelleberg, S. Cell Death in Pseudomonas Aeruginosa Biofilm Development. J. Bacteriol. 2003, 185, 4585–4592. [Google Scholar] [CrossRef] [Green Version]
- Christensen, G.D.; Simpson, W.A.; Younger, J.J.; Baddour, L.M.; Barrett, F.F.; Melton, D.M.; Beachey, E.H. Adherence of Coagulase-Negative Staphylococci to Plastic Tissue Culture Plates: A Quantitative Model for the Adherence of Staphylococci to Medical Devices. J. Clin. Microbiol. 1985, 22, 996–1006. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 1 March 2022).
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of Long, Error-Prone Reads Using Repeat Graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive Visualization of de Novo Genome Assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; O’Neill, K.R.; Haft, D.H.; DiCuccio, M.; Chetvernin, V.; Badretdin, A.; Coulouris, G.; Chitsaz, F.; Derbyshire, M.K.; Durkin, A.S.; et al. RefSeq: Expanding the Prokaryotic Genome Annotation Pipeline Reach with Protein Family Model Curation. Nucleic Acids Res. 2021, 49, D1020–D1028. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI Prokaryotic Genome Annotation Pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A Database Tandem for Fast and Reliable Genome-Based Classification and Nomenclature of Prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M. TYGS Is an Automated High-Throughput Platform for State-of-the-Art Genome-Based Taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [Green Version]
- Winsor, G.L.; Van Rossum, T.; Lo, R.; Khaira, B.; Whiteside, M.D.; Hancock, R.E.W.; Brinkman, F.S.L. Pseudomonas Genome Database: Facilitating User-Friendly, Comprehensive Comparisons of Microbial Genomes. Nucleic Acids Res. 2009, 37, D483–D488. [Google Scholar] [CrossRef] [Green Version]
- Winsor, G.L.; Griffiths, E.J.; Lo, R.; Dhillon, B.K.; Shay, J.A.; Brinkman, F.S.L. Enhanced Annotations and Features for Comparing Thousands of Pseudomonas Genomes in the Pseudomonas Genome Database. Nucleic Acids Res. 2016, 44, D646–D653. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.-H.; Ha, S.; Lim, J.; Kwon, S.; Chun, J. A Large-Scale Evaluation of Algorithms to Calculate Average Nucleotide Identity. Antonie Van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Blom, J.; Albaum, S.P.; Doppmeier, D.; Pühler, A.; Vorhölter, F.-J.; Zakrzewski, M.; Goesmann, A. EDGAR: A Software Framework for the Comparative Analysis of Prokaryotic Genomes. BMC Bioinformatics 2009, 10, 154. [Google Scholar] [CrossRef] [Green Version]
- Dieckmann, M.A.; Beyvers, S.; Nkouamedjo-Fankep, R.C.; Hanel, P.H.G.; Jelonek, L.; Blom, J.; Goesmann, A. EDGAR3.0: Comparative Genomics and Phylogenomics on a Scalable Infrastructure. Nucleic Acids Res. 2021, 49, W185–W192. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Thrane, S.W.; Taylor, V.L.; Lund, O.; Lam, J.S.; Jelsbak, L. Application of Whole-Genome Sequencing Data for O-Specific Antigen Analysis and In Silico Serotyping of Pseudomonas Aeruginosa Isolates. J. Clin. Microbiol. 2016, 54, 1782–1788. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A Better, Faster Version of the PHAST Phage Search Tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A Reference Database for Bacterial Virulence Factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef]
- Colvin, K.M.; Irie, Y.; Tart, C.S.; Urbano, R.; Whitney, J.C.; Ryder, C.; Howell, P.L.; Wozniak, D.J.; Parsek, M.R. The Pel and Psl Polysaccharides Provide Pseudomonas Aeruginosa Structural Redundancy within the Biofilm Matrix. Environ. Microbiol. 2012, 14. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.K.; Almpani, M.; Maura, D.; Kitao, T.; Ferrari, L.; Fontana, S.; Bergamini, G.; Calcaterra, E.; Pignaffo, C.; Negri, M.; et al. Tackling Recalcitrant Pseudomonas Aeruginosa Infections in Critical Illness via Anti-Virulence Monotherapy. Nat. Commun. 2022, 13, 5103. [Google Scholar] [CrossRef] [PubMed]
- Alikhan, N.-F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple Prokaryote Genome Comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, P.H.; Tetu, S.G.; Larouche, A.; Elbourne, L.; Tremblay, S.; Ren, Q.; Dodson, R.; Harkins, D.; Shay, R.; Watkins, K.; et al. Complete Genome Sequence of the Multiresistant Taxonomic Outlier Pseudomonas Aeruginosa PA7. PLoS ONE 2010, 5, e8842. [Google Scholar] [CrossRef] [PubMed]
- Stewart, L.; Ford, A.; Sangal, V.; Jeukens, J.; Boyle, B.; Kukavica-Ibrulj, I.; Caim, S.; Crossman, L.; Hoskisson, P.A.; Levesque, R.; et al. Draft Genomes of 12 Host-Adapted and Environmental Isolates of Pseudomonas Aeruginosa and Their Positions in the Core Genome Phylogeny. Pathog. Dis. 2014, 71, 20–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kos, V.N.; Déraspe, M.; McLaughlin, R.E.; Whiteaker, J.D.; Roy, P.H.; Alm, R.A.; Corbeil, J.; Gardner, H. The Resistome of Pseudomonas Aeruginosa in Relationship to Phenotypic Susceptibility. Antimicrob. Agents Chemother. 2014, 59, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Freschi, L.; Jeukens, J.; Kukavica-Ibrulj, I.; Boyle, B.; Dupont, M.-J.; Laroche, J.; Larose, S.; Maaroufi, H.; Fothergill, J.L.; Moore, M.; et al. Clinical Utilization of Genomics Data Produced by the International Pseudomonas Aeruginosa Consortium. Front. Microbiol. 2015, 6, 1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, K.; Smyth, R.L.; Govan, J.R.; Doherty, C.; Winstanley, C.; Denning, N.; Heaf, D.P.; van Saene, H.; Hart, C.A. Spread of β-Lactam-Resistant Pseudomonas Aeruginosa in a Cystic Fibrosis Clinic. Lancet 1996, 348, 639–642. [Google Scholar] [CrossRef]
- Yan, J.; Deforet, M.; Boyle, K.E.; Rahman, R.; Liang, R.; Okegbe, C.; Dietrich, L.E.P.; Qiu, W.; Xavier, J.B. Bow-Tie Signaling in c-Di-GMP: Machine Learning in a Simple Biochemical Network. PLoS Comput. Biol. 2017, 13, e1005677. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.G.; Urbach, J.M.; Wu, G.; Liberati, N.T.; Feinbaum, R.L.; Miyata, S.; Diggins, L.T.; He, J.; Saucier, M.; Déziel, E.; et al. Genomic Analysis Reveals That Pseudomonas Aeruginosa Virulence Is Combinatorial. Genome Biol. 2006, 7, R90. [Google Scholar] [CrossRef] [PubMed]
- Sanjar, F.; Karna, S.L.R.; Chen, T.; Chen, P.; Abercrombie, J.J.; Leung, K.P. Whole-Genome Sequence of Multidrug-Resistant Pseudomonas Aeruginosa Strain BAMCPA07-48, Isolated from a Combat Injury Wound. Genome Announc. 2016, 4, e00547-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghupathi, P.K.; Zupančič, J.; Brejnrod, A.D.; Jacquiod, S.; Houf, K.; Burmølle, M.; Gunde-Cimerman, N.; Sørensen, S.J. Microbial Diversity and Putative Opportunistic Pathogens in Dishwasher Biofilm Communities. Appl. Environ. Microbiol. 2018, 84, e02755-17. [Google Scholar] [CrossRef] [Green Version]
- Zupančič, J.; Turk, M.; Črnigoj, M.; Ambrožič Avguštin, J.; Gunde-Cimerman, N. The Dishwasher Rubber Seal Acts as a Reservoir of Bacteria in the Home Environment. BMC Microbiol. 2019, 19, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikkelsen, H.; McMullan, R.; Filloux, A. The Pseudomonas Aeruginosa Reference Strain PA14 Displays Increased Virulence Due to a Mutation in LadS. PLoS ONE 2011, 6, e29113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgener, E.B.; Sweere, J.M.; Bach, M.S.; Secor, P.R.; Haddock, N.; Jennings, L.K.; Marvig, R.L.; Johansen, H.K.; Rossi, E.; Cao, X.; et al. Filamentous Bacteriophages Are Associated with Chronic Pseudomonas Lung Infections and Antibiotic Resistance in Cystic Fibrosis. Sci. Transl. Med. 2019, 11, eaau9748. [Google Scholar] [CrossRef] [PubMed]
- Secor, P.R.; Sweere, J.M.; Michaels, L.A.; Malkovskiy, A.V.; Lazzareschi, D.; Katznelson, E.; Rajadas, J.; Birnbaum, M.E.; Arrigoni, A.; Braun, K.R.; et al. Filamentous Bacteriophage Promote Biofilm Assembly and Function. Cell Host Microbe 2015, 18, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Secor, P.R.; Jennings, L.K.; Michaels, L.A.; Sweere, J.M.; Singh, P.K.; Parks, W.C.; Bollyky, P.L. Biofilm Assembly Becomes Crystal Clear-Filamentous Bacteriophage Organize the Pseudomonas Aeruginosa Biofilm Matrix into a Liquid Crystal. Microb. Cell Graz Austria 2015, 3, 49–52. [Google Scholar] [CrossRef]
- Petrova, O.E.; Schurr, J.R.; Schurr, M.J.; Sauer, K. The Novel Pseudomonas Aeruginosa Two-Component Regulator BfmR Controls Bacteriophage-Mediated Lysis and DNA Release during Biofilm Development through PhdA. Mol. Microbiol. 2011, 81, 767–783. [Google Scholar] [CrossRef] [Green Version]
- Johnson, G.; Wolfe, A.J.; Putonti, C. Characterization of the ΦCTX-like Pseudomonas Aeruginosa Phage Dobby Isolated from the Kidney Stone Microbiota. Access Microbiol. 2019, 1, e000002. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An Emergent Form of Bacterial Life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Friedman, L.; Kolter, R. Genes Involved in Matrix Formation in Pseudomonas Aeruginosa PA14 Biofilms. Mol. Microbiol. 2004, 51, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Lam, J.; Taylor, V.; Islam, S.; Hao, Y.; Kocíncová, D. Genetic and Functional Diversity of Pseudomonas Aeruginosa Lipopolysaccharide. Front. Microbiol. 2011, 2, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, K.; Takashima, K.; Ishihara, H.; Shinomiya, T.; Kageyama, M.; Kanaya, S.; Ohnishi, M.; Murata, T.; Mori, H.; Hayashi, T. The R-Type Pyocin of Pseudomonas Aeruginosa Is Related to P2 Phage, and the F-Type Is Related to Lambda Phage. Mol. Microbiol. 2000, 38, 213–231. [Google Scholar] [CrossRef] [PubMed]
- Köhler, T.; Donner, V.; van Delden, C. Lipopolysaccharide as Shield and Receptor for R-Pyocin-Mediated Killing in Pseudomonas Aeruginosa. J. Bacteriol. 2010, 192, 1921–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beveridge, T.J.; Makin, S.A.; Kadurugamuwa, J.L.; Li, Z. Interactions between Biofilms and the Environment. FEMS Microbiol. Rev. 1997, 20, 291–303. [Google Scholar] [CrossRef]
- Liu, P.V. Comparison of the Chinese Schema and the International Antigenic Typing System for Serotyping Pseudomonas Aeruginosa. J. Clin. Microbiol. 1987, 25, 824–826. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.V.; Wang, S. Three New Major Somatic Antigens of Pseudomonas Aeruginosa. J. Clin. Microbiol. 1990, 28, 922–925. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.V.; Matsumoto, H.; Kusama, H.; Bergan, T. Survey of Heat-Stable, Major Somatic Antigens of Pseudomonas Aeruginosa. Int. J. Syst. Evol. Microbiol. 1983, 33, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Raymond, C.K.; Sims, E.H.; Kas, A.; Spencer, D.H.; Kutyavin, T.V.; Ivey, R.G.; Zhou, Y.; Kaul, R.; Clendenning, J.B.; Olson, M.V. Genetic Variation at the O-Antigen Biosynthetic Locus in Pseudomonas Aeruginosa. J. Bacteriol. 2002, 184, 3614–3622. [Google Scholar] [CrossRef] [Green Version]
- Nasrin, S.; Hegerle, N.; Sen, S.; Nkeze, J.; Sen, S.; Permala-Booth, J.; Choi, M.; Sinclair, J.; Tapia, M.D.; Johnson, J.K.; et al. Distribution of Serotypes and Antibiotic Resistance of Invasive Pseudomonas Aeruginosa in a Multi-Country Collection. BMC Microbiol. 2022, 22, 13. [Google Scholar] [CrossRef]
- Lu, Q.; Eggimann, P.; Luyt, C.-E.; Wolff, M.; Tamm, M.; François, B.; Mercier, E.; Garbino, J.; Laterre, P.-F.; Koch, H.; et al. Pseudomonas Aeruginosa Serotypes in Nosocomial Pneumonia: Prevalence and Clinical Outcomes. Crit. Care 2014, 18, R17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witney, A.A.; Gould, K.A.; Pope, C.F.; Bolt, F.; Stoker, N.G.; Cubbon, M.D.; Bradley, C.R.; Fraise, A.; Breathnach, A.S.; Butcher, P.D.; et al. Genome Sequencing and Characterization of an Extensively Drug-Resistant Sequence Type 111 Serotype O12 Hospital Outbreak Strain of Pseudomonas Aeruginosa. Clin. Microbiol. Infect. 2014, 20, O609–O618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bert, F.; Lambert-Zechovsky, N.; Seymour, A.C. Comparitive Distribution of Resistance Patterns and Serotypes in Pseudomonas Aeruginosa Isolates from Intensive Care Units and Other Wards. J. Antimicrob. Chemother. 1996, 37, 809–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirnay, J.-P.; Bilocq, F.; Pot, B.; Cornelis, P.; Zizi, M.; Eldere, J.V.; Deschaght, P.; Vaneechoutte, M.; Jennes, S.; Pitt, T.; et al. Pseudomonas Aeruginosa Population Structure Revisited. PLoS ONE 2009, 4, e7740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, D.S.; Rizo, A.G.; Monroy, S.P.; Reyes, T. Antimicrobial susceptibility and serotyping of Pseudomonas aeruginosa isolated from HIV/AIDS patients. Rev. Cubana Med. Trop. 2002, 54, 142–146. [Google Scholar]
- Estahbanati, H.K.; Kashani, P.P.; Ghanaatpisheh, F. Frequency of Pseudomonas Aeruginosa Serotypes in Burn Wound Infections and Their Resistance to Antibiotics. Burns 2002, 28, 340–348. [Google Scholar] [CrossRef]
- Meyer, J.-M.; Stintzi, A. Iron Metabolism and Siderophores in Pseudomonas and Related Species. In Pseudomonas; Biotechnology Handbooks; Montie, T.C., Ed.; Springer: Boston, MA, USA, 1998; pp. 201–243. ISBN 978-1-4899-0120-0. [Google Scholar]
- Meyer, J.M.; Neely, A.; Stintzi, A.; Georges, C.; Holder, I.A. Pyoverdin Is Essential for Virulence of Pseudomonas Aeruginosa. Infect. Immun. 1996, 64, 518–523. [Google Scholar] [CrossRef] [Green Version]
- McMorran, B.J.; Merriman, M.E.; Rombel, I.T.; Lamont, I.L. Characterisation of the PvdE Gene Which Is Required for Pyoverdine Synthesis in Pseudomonas Aeruginosa. Gene 1996, 176, 55–59. [Google Scholar] [CrossRef]
- Subedi, D.; Vijay, A.K.; Kohli, G.S.; Rice, S.A.; Willcox, M. Comparative Genomics of Clinical Strains of Pseudomonas Aeruginosa Strains Isolated from Different Geographic Sites. Sci. Rep. 2018, 8, 15668. [Google Scholar] [CrossRef] [Green Version]
- Stewart, R.M.K.; Wiehlmann, L.; Ashelford, K.E.; Preston, S.J.; Frimmersdorf, E.; Campbell, B.J.; Neal, T.J.; Hall, N.; Tuft, S.; Kaye, S.B.; et al. Genetic Characterization Indicates That a Specific Subpopulation of Pseudomonas Aeruginosa Is Associated with Keratitis Infections. J. Clin. Microbiol. 2011, 49, 993–1003. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, S.; Suzuki, T.; Kobayashi, T.; Oka, N.; Ishikawa, E.; Shinomiya, H.; Ohashi, Y. Genotypic Analysis of Pseudomonas Aeruginosa Isolated from Ocular Infection. J. Infect. Chemother. 2014, 20, 407–411. [Google Scholar] [CrossRef]
- Tingpej, P.; Smith, L.; Rose, B.; Zhu, H.; Conibear, T.; Al Nassafi, K.; Manos, J.; Elkins, M.; Bye, P.; Willcox, M.; et al. Phenotypic Characterization of Clonal and Nonclonal Pseudomonas Aeruginosa Strains Isolated from Lungs of Adults with Cystic Fibrosis. J. Clin. Microbiol. 2007, 45, 1697–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilderman, P.J.; Vasil, A.I.; Johnson, Z.; Vasil, M.L. Genetic and Biochemical Analyses of a Eukaryotic-like Phospholipase D of Pseudomonas Aeruginosa Suggest Horizontal Acquisition and a Role for Persistence in a Chronic Pulmonary Infection Model. Mol. Microbiol. 2001, 39, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Spencer, C.; Brown, H.A. Biochemical Characterization of a Pseudomonas Aeruginosa Phospholipase D. Biochemistry 2015, 54, 1208–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rumbaugh, K.P.; Griswold, J.A.; Hamood, A.N. The Role of Quorum Sensing in the in Vivo Virulence of Pseudomonas Aeruginosa. Microbes Infect. 2000, 2, 1721–1731. [Google Scholar] [CrossRef]
- Davies, D.G.; Parsek, M.R.; Pearson, J.P.; Iglewski, B.H.; Costerton, J.W.; Greenberg, E.P. The Involvement of Cell-to-Cell Signals in the Development of a Bacterial Biofilm. Science 1998, 280, 295–298. [Google Scholar] [CrossRef] [Green Version]
- Ozer, E.; Yaniv, K.; Chetrit, E.; Boyarski, A.; Meijler, M.M.; Berkovich, R.; Kushmaro, A.; Alfonta, L. An inside Look at a Biofilm: Pseudomonas Aeruginosa Flagella Biotracking. Sci. Adv. 2021, 7, eabg8581. [Google Scholar] [CrossRef]
- Dasgupta, N.; Arora, S.K.; Ramphal, R. The Flagellar System of Pseudomonas Aeruginosa. In Pseudomonas: Volume 1 Genomics, Life Style and Molecular Architecture; Ramos, J.-L., Ed.; Springer: Boston, MA, USA, 2004; pp. 675–698. ISBN 978-1-4419-9086-0. [Google Scholar]
- Gellatly, S.L.; Hancock, R.E.W. Pseudomonas Aeruginosa: New Insights into Pathogenesis and Host Defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [Green Version]
- Miao, E.A.; Andersen-Nissen, E.; Warren, S.E.; Aderem, A. TLR5 and Ipaf: Dual Sensors of Bacterial Flagellin in the Innate Immune System. Semin. Immunopathol. 2007, 29, 275–288. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WM Compartment | Name of WM Isolates |
|---|---|
| detergent compartment | B1.2 |
| detergent enema | B2.1 |
| detergent compartment | C1.3 |
| sealing rubber | C4.2 |
| Sampling Site | Isolate | Length (bp) | GC (%) | Total Genes (CDS) | Sequence Type | Serotype Group |
|---|---|---|---|---|---|---|
| WM isolates | ||||||
| detergent compartment | B1.2 | 6,387,430 | 66 | 5930 (5851) | 369 | O6 |
| detergent enema | B2.1 | 6,387,416 | 66 | 5930 (5851) | 369 | O6 |
| detergent compartment | C1.3 | 7,303,308 | 65 | 7034 (6952) | 313 | O1 |
| sealing rubber | C4.2 | 7,345,693 | 66 | 6987 (6900) | 2844 | O13 |
| Reference strains | ||||||
| human wound | PAO1 | 6,264,404 * | 67 * | 5708 (5587) * | 549 | O5 |
| human burn wound | UCBPP-PA14 | 6,537,648 * | 66 * | 5983 (5894) * | 253 | O10 |
| human non-respiratory clinical isolate | PA7 | 6,588,339 * | 66 * | 6369 (6286) * | 1195 | O12 |
| human sputum | LESB58 | 6,601,757 * | 66 * | 6028 (5927) * | 146 | O6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiel, A.; Creutz, I.; Rückert, C.; Kaltschmidt, B.P.; Hütten, A.; Niehaus, K.; Busche, T.; Kaltschmidt, B.; Kaltschmidt, C. Genome-Based Analysis of Virulence Factors and Biofilm Formation in Novel P. aeruginosa Strains Isolated from Household Appliances. Microorganisms 2022, 10, 2508. https://doi.org/10.3390/microorganisms10122508
Kiel A, Creutz I, Rückert C, Kaltschmidt BP, Hütten A, Niehaus K, Busche T, Kaltschmidt B, Kaltschmidt C. Genome-Based Analysis of Virulence Factors and Biofilm Formation in Novel P. aeruginosa Strains Isolated from Household Appliances. Microorganisms. 2022; 10(12):2508. https://doi.org/10.3390/microorganisms10122508
Chicago/Turabian StyleKiel, Annika, Ines Creutz, Christian Rückert, Bernhard Peter Kaltschmidt, Andreas Hütten, Karsten Niehaus, Tobias Busche, Barbara Kaltschmidt, and Christian Kaltschmidt. 2022. "Genome-Based Analysis of Virulence Factors and Biofilm Formation in Novel P. aeruginosa Strains Isolated from Household Appliances" Microorganisms 10, no. 12: 2508. https://doi.org/10.3390/microorganisms10122508