
Roseburia intestinalis Modulates PYY Expression in a New a Multicellular Model including Enteroendocrine Cells
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Growth Culture
2.1.1. Endocrine Cells (NCI-H716) Differentiation
2.1.2. Cellular Model Composed of Four Cell Types (Quadricellular Model)
2.2. TEER Measurements
2.3. Transmission Electron Microscopy
2.4. Quantification of Selected Genes Expression Level by Quantitative Reverse Transcription PCR (RT-qPCR)
2.5. Cytotoxicity Evaluation by Measuring LDH (Lactate Deshydrogenase) Release
2.6. Bacteria Growth Conditions
2.7. Determination of Short Chain Fatty Acids (SCFAs) by Gas Chromatography Mass Spectrometry (GC/MS)
2.8. Co-Incubation of the Quadricellular Model with Bacteria or Their Cell-Free Supernatant
2.9. Statistical Analysis
3. Results
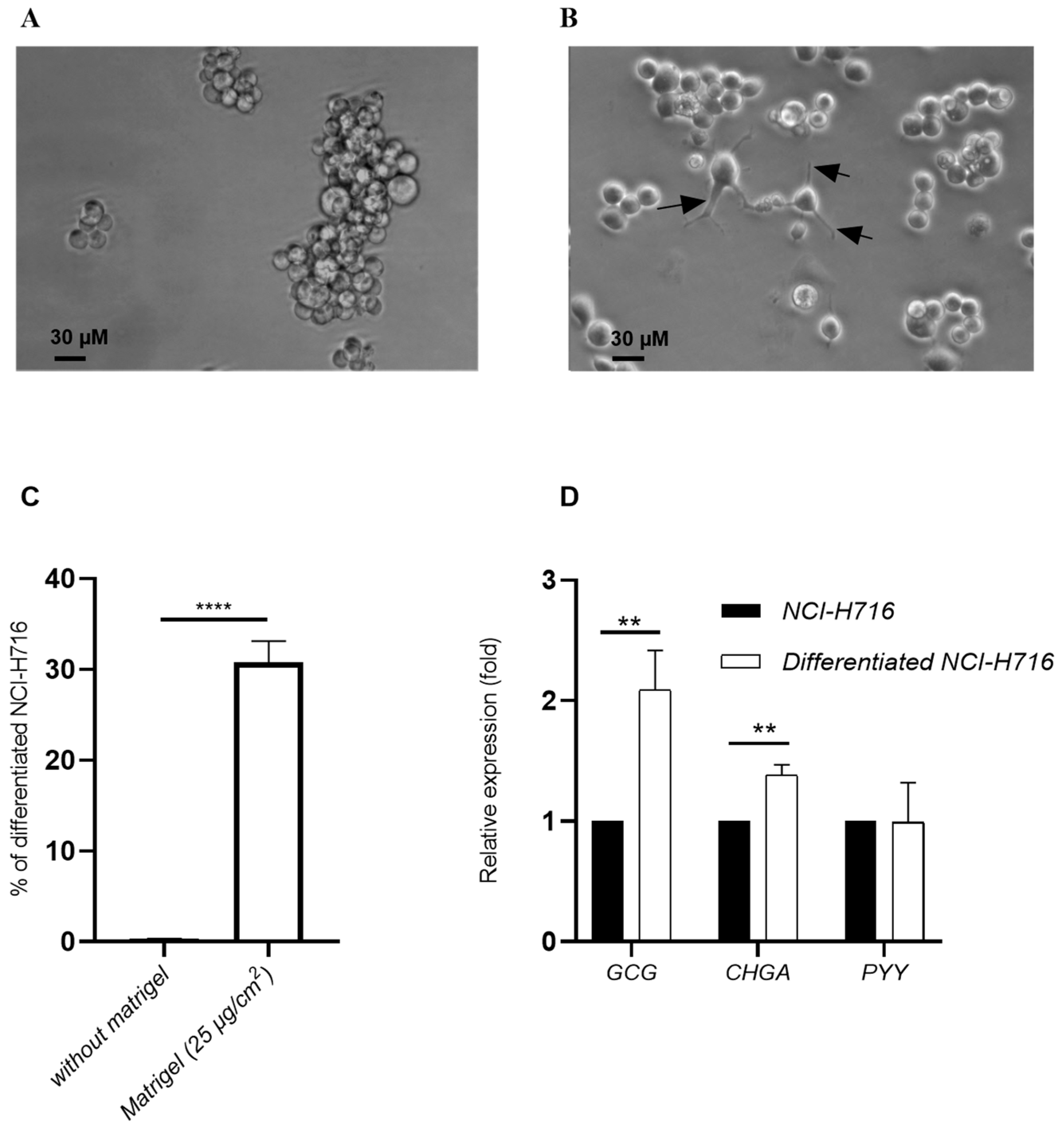
3.1. Differentiated NCI-H716 Cells Expressed Proglucagon (GCG) and Chromogranin A (CHGA) Genes
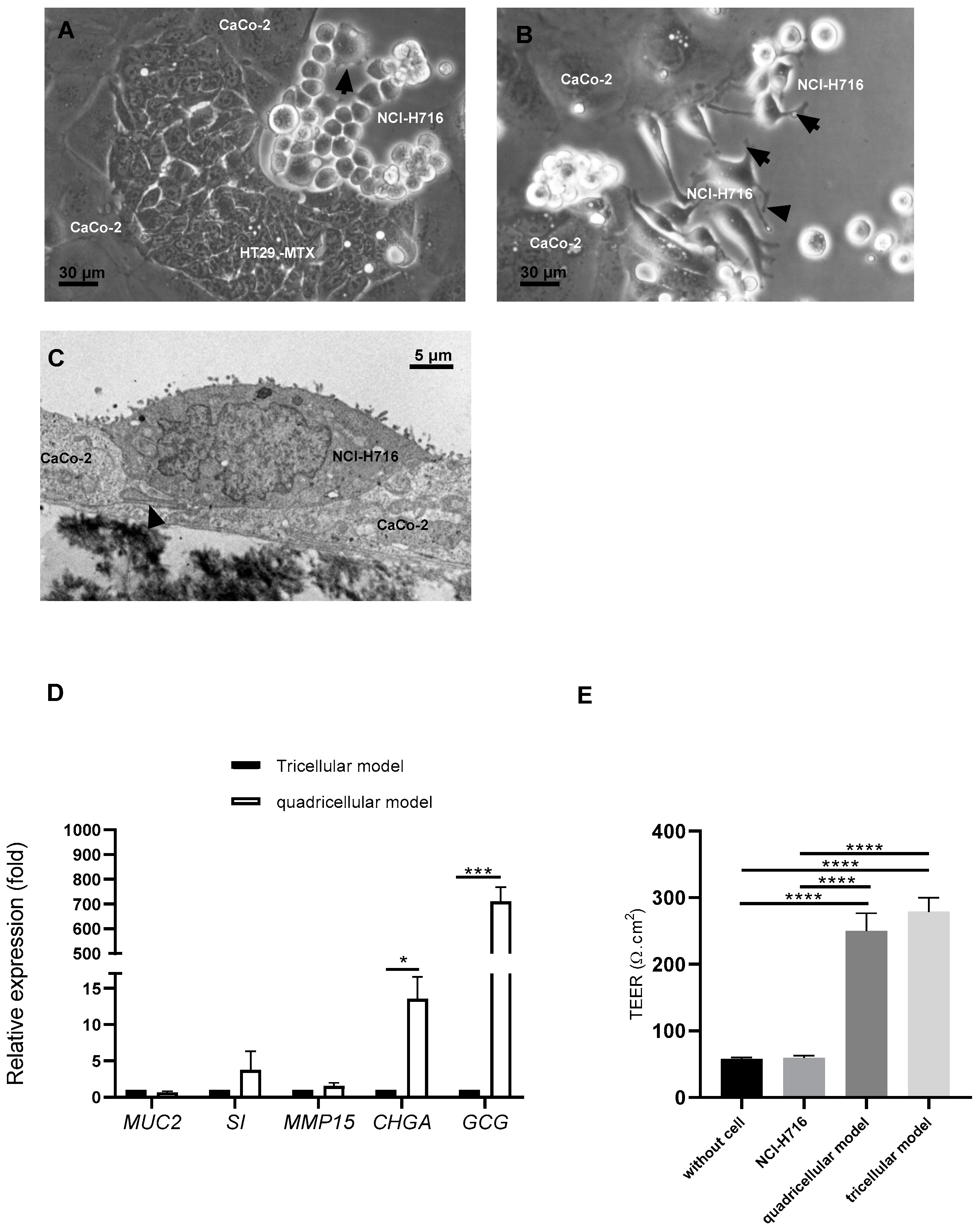
3.2. Endocrine-Differentiated NCI-H716 Are Functional in Co-Culture with Caco-2, HT-29 MTX and M Cells
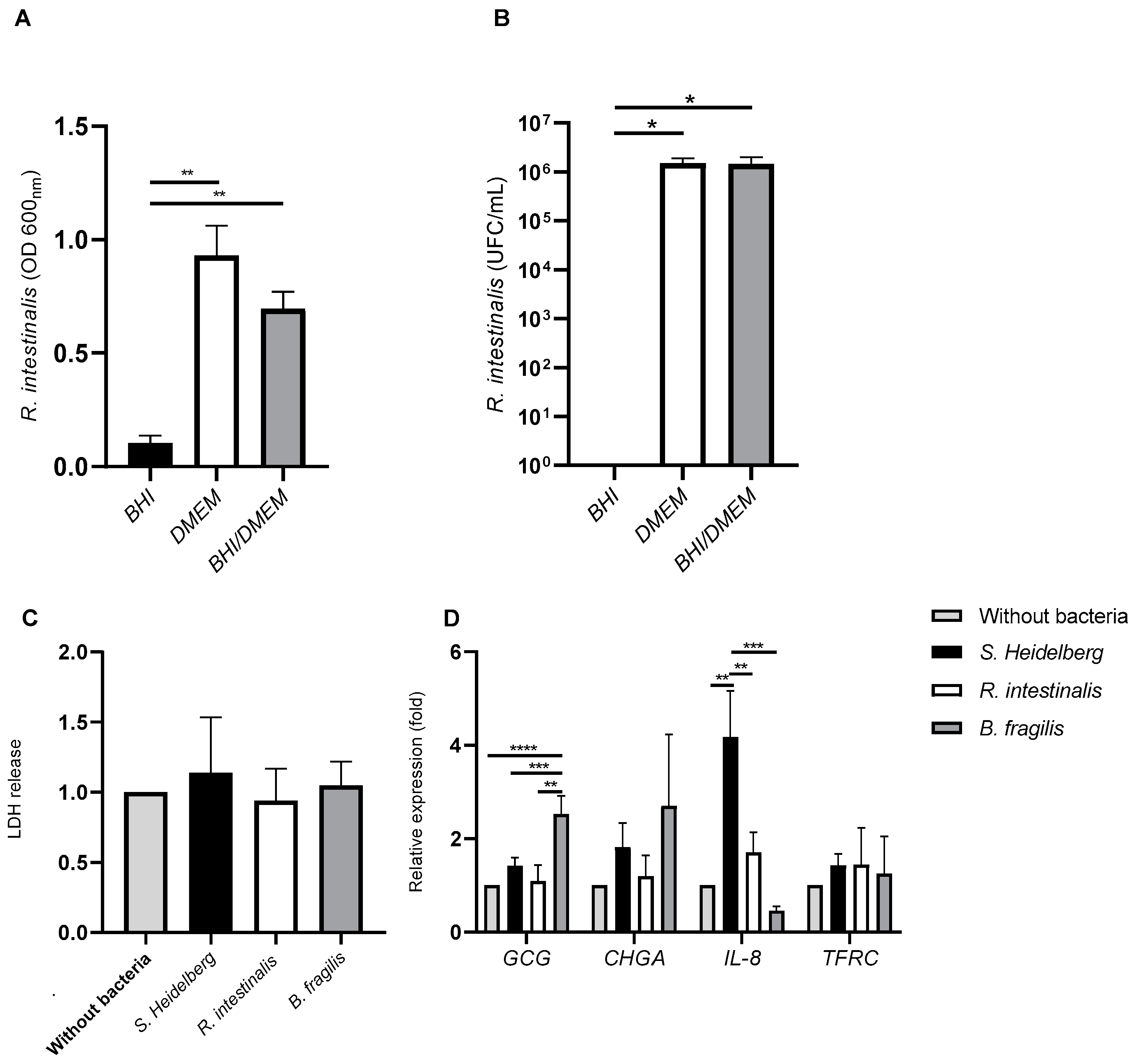
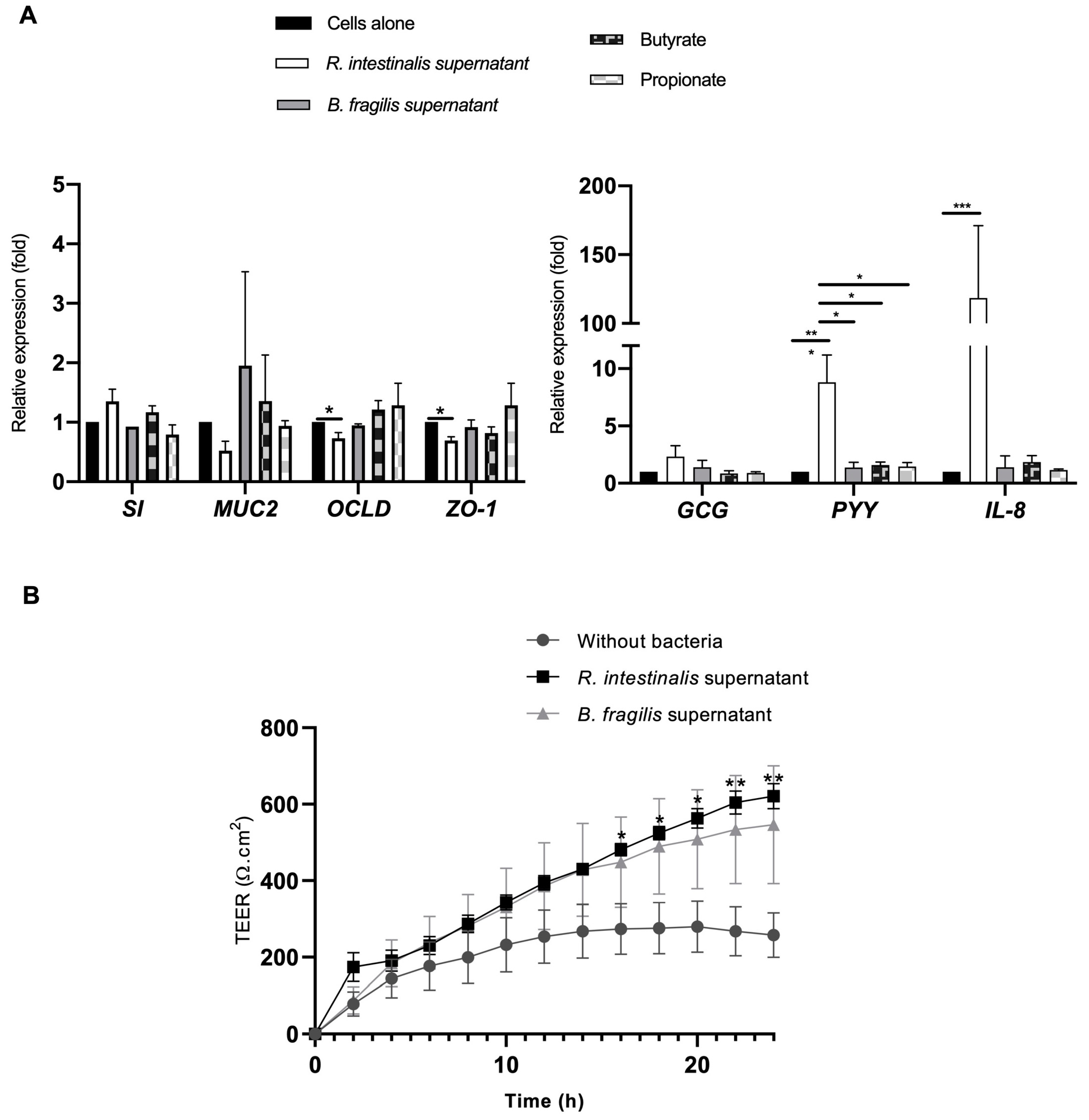
3.3. The Impact of Commensal and Pathogenic Bacteria on Quadricellular Model
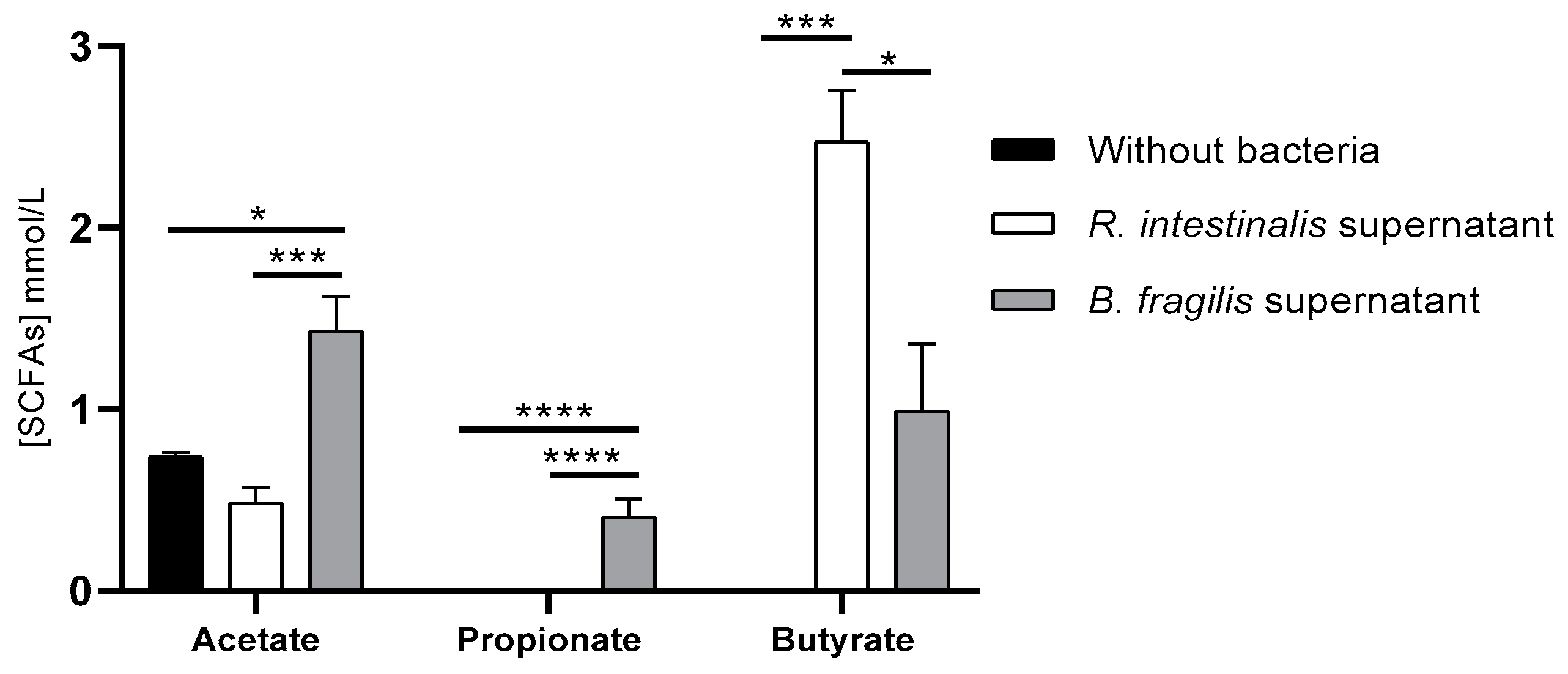
3.4. R. intestinalis and B. fragilis Produced SCFAs in DMEM
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- de Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut Microbiome and Health: Mechanistic Insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef] [PubMed]
- Sohail, M.U.; Althani, A.; Anwar, H.; Rizzi, R.; Marei, H.E. Role of the Gastrointestinal Tract Microbiome in the Pathophysiology of Diabetes Mellitus. J. Diabetes Res. 2017, 2017, 9631435. [Google Scholar] [CrossRef] [PubMed]
- Esser, N.; Legrand-Poels, S.; Piette, J.; Scheen, A.J.; Paquot, N. Inflammation as a Link between Obesity, Metabolic Syndrome and Type 2 Diabetes. Diabetes Res. Clin. Pract. 2014, 105, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashtanova, D.A.; Tkacheva, O.N.; Doudinskaya, E.N.; Strazhesko, I.D.; Kotovskaya, Y.V.; Popenko, A.S.; Tyakht, A.V.; Alexeev, D.G. Gut Microbiota in Patients with Different Metabolic Statuses: Moscow Study. Microorganisms 2018, 6, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takagi, T.; Naito, Y.; Kashiwagi, S.; Uchiyama, K.; Mizushima, K.; Kamada, K.; Ishikawa, T.; Inoue, R.; Okuda, K.; Tsujimoto, Y.; et al. Changes in the Gut Microbiota Are Associated with Hypertension, Hyperlipidemia, and Type 2 Diabetes Mellitus in Japanese Subjects. Nutrients 2020, 12, 2996. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Watanabe, K.; Kimura, I. Gut Microbiota Dysbiosis Drives and Implies Novel Therapeutic Strategies for Diabetes Mellitus and Related Metabolic Diseases. Front. Immunol. 2017, 8, 1882. [Google Scholar] [CrossRef]
- Schimpel, C.; Teubl, B.; Absenger, M.; Meindl, C.; Fröhlich, E.; Leitinger, G.; Zimmer, A.; Roblegg, E. Development of an Advanced Intestinal in Vitro Triple Culture Permeability Model to Study Transport of Nanoparticles. Mol. Pharm. 2014, 11, 808–818. [Google Scholar] [CrossRef]
- Vernay, T.; Cannie, I.; Gaboriau, F.; Gall, S.D.-L.; Tamanai-Shacoori, Z.; Burel, A.; Jolivet-Gougeon, A.; Loréal, O.; Bousarghin, L. Bacteroides Fragilis Prevents Salmonella Heidelberg Translocation in Co-Culture Model Mimicking Intestinal Epithelium. Benef. Microbes 2020, 11, 391–401. [Google Scholar] [CrossRef]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of Gut Microbiota in Type 2 Diabetes Pathophysiology. eBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut Bacteria Selectively Promoted by Dietary Fibers Alleviate Type 2 Diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef]
- Larraufie, P.; Roberts, G.P.; McGavigan, A.K.; Kay, R.G.; Li, J.; Leiter, A.; Melvin, A.; Biggs, E.K.; Ravn, P.; Davy, K.; et al. Important Role of the GLP-1 Axis for Glucose Homeostasis after Bariatric Surgery. Cell Rep. 2019, 26, 1399–1408.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, S.H.; Hold, G.L.; Barcenilla, A.; Stewart, C.S.; Flint, H.J. Roseburia Intestinalis Sp. Nov., a Novel Saccharolytic, Butyrate-Producing Bacterium from Human Faeces. Int. J. Syst. Evol. Microbiol. 2002, 52, 1615–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Cheng, Y.; Ruan, G.; Fan, L.; Tian, Y.; Xiao, Z.; Chen, D.; Wei, Y. New Pathway Ameliorating Ulcerative Colitis: Focus on Roseburia Intestinalis and the Gut–Brain Axis. Ther. Adv. Gastroenterol. 2021, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bickel, M. The Role of Interleukin-8 in Inflammation and Mechanisms of Regulation. J. Periodontol. 1993, 64, 456–460. [Google Scholar]
- Huang, N.; Katz, J.P.; Martin, D.R.; Wu, G.D. Inhibition of IL-8 Gene Expression in Caco-2 Cells by Compounds Which Induce Histone Hyperacetylation. Cytokine 1997, 9, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Fusunyan, R.D.; Quinn, J.J.; Fujimoto, M.; MacDermott, R.P.; Sanderson, I.R. Butyrate Switches the Pattern of Chemokine Secretion by Intestinal Epithelial Cells through Histone Acetylation. Mol. Med. 1999, 5, 631–640. [Google Scholar] [CrossRef] [Green Version]
- Gautier, T.; David-Le Gall, S.; Sweidan, A.; Tamanai-Shacoori, Z.; Jolivet-Gougeon, A.; Loréal, O.; Bousarghin, L. Next-Generation Probiotics and Their Metabolites in COVID-19. Microorganisms 2021, 9, 941. [Google Scholar] [CrossRef]
- Kasubuchi, M.; Hasegawa, S.; Hiramatsu, T.; Ichimura, A.; Kimura, I. Dietary Gut Microbial Metabolites, Short-Chain Fatty Acids, and Host Metabolic Regulation. Nutrients 2015, 7, 2839–2849. [Google Scholar] [CrossRef] [Green Version]
- Pingitore, A.; Chambers, E.S.; Hill, T.; Maldonado, I.R.; Liu, B.; Bewick, G.; Morrison, D.J.; Preston, T.; Wallis, G.A.; Tedford, C.; et al. The Diet-Derived Short Chain Fatty Acid Propionate Improves Beta-Cell Function in Humans and Stimulates Insulin Secretion from Human Islets in Vitro. Diabetes Obes. Metab. 2017, 19, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Zong, Y.; Chen, W.; Zhao, Y.; Suo, X.; Yang, X. Salmonella Infection Causes Hyperglycemia for Decreased GLP-1 Content by Enteroendocrine L Cells Pyroptosis in Pigs. Int. J. Mol. Sci. 2022, 23, 1272. [Google Scholar] [CrossRef]
- Tamanai-Shacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Roseburia Spp.: A Marker of Health? Future Microbiol. 2017, 12, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Song, K.; Shen, Z.; Quan, Y.; Tan, B.; Luo, W.; Wu, S.; Tang, K.; Yang, Z.; Wang, X. Roseburia Intestinalis Inhibits Interleukin-17 Excretion and Promotes Regulatory T Cells Differentiation in Colitis. Mol. Med. Rep. 2018, 17, 7567–7574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Duarte, M.E.; Kim, S.W. Dietary Inclusion of Multispecies Probiotics to Reduce the Severity of Post-Weaning Diarrhea Caused by Escherichia Coli F18+ in Pigs. Anim. Nutr. 2021, 7, 326–333. [Google Scholar] [CrossRef]
- Tan, T.; Behary, P.; Tharakan, G.; Minnion, J.; Al-Najim, W.; Albrechtsen, N.J.W.; Holst, J.J.; Bloom, S.R. The Effect of a Subcutaneous Infusion of GLP-1, OXM, and PYY on Energy Intake and Expenditure in Obese Volunteers. J. Clin. Endocrinol. Metab. 2017, 102, 2364–2372. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.D.; Finan, B.; Bloom, S.R.; D’Alessio, D.; Drucker, D.J.; Flatt, P.R.; Fritsche, A.; Gribble, F.; Grill, H.J.; Habener, J.F.; et al. Glucagon-like Peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef] [PubMed]
- Svane, M.S.; Jørgensen, N.B.; Bojsen-Møller, K.N.; Dirksen, C.; Nielsen, S.; Kristiansen, V.B.; Toräng, S.; Wewer Albrechtsen, N.J.; Rehfeld, J.F.; Hartmann, B.; et al. Peptide YY and Glucagon-like Peptide-1 Contribute to Decreased Food Intake after Roux-En-Y Gastric Bypass Surgery. Int. J. Obes. 2016, 40, 1699–1706. [Google Scholar] [CrossRef]
- Tsai, C.-Y.; Lu, H.-C.; Chou, Y.-H.; Liu, P.-Y.; Chen, H.-Y.; Huang, M.-C.; Lin, C.-H.; Tsai, C.-N. Gut Microbial Signatures for Glycemic Responses of GLP-1 Receptor Agonists in Type 2 Diabetic Patients: A Pilot Study. Front. Endocrinol. 2022, 12, 814770. [Google Scholar] [CrossRef]
- Panwar, H.; Calderwood, D.; Gillespie, A.L.; Wylie, A.R.; Graham, S.F.; Grant, I.R.; Grover, S.; Green, B.D. Identification of Lactic Acid Bacteria Strains Modulating Incretin Hormone Secretion and Gene Expression in Enteroendocrine Cells. J. Funct. Foods 2016, 23, 348–358. [Google Scholar] [CrossRef] [Green Version]
- Park, J.G.; Oie, H.K.; Sugarbaker, P.H.; Henslee, J.G.; Chen, T.R.; Johnson, B.E.; Gazdar, A. Characteristics of Cell Lines Established from Human Colorectal Carcinoma. Cancer Res. 1987, 47, 6710–6718. [Google Scholar]
- Le Bars, H.; Le Gall-David, S.; Renoux, V.M.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A.; Bousarghin, L. Impact of a Mutator Phenotype on Motility and Cell Adherence in Salmonella Heidelberg. Vet. Microbiol. 2012, 159, 99–106. [Google Scholar] [CrossRef]
- Anderson, R.C.; MacGibbon, A.K.H.; Haggarty, N.; Armstrong, K.M.; Roy, N.C. Bovine Dairy Complex Lipids Improve in Vitro Measures of Small Intestinal Epithelial Barrier Integrity. PLoS ONE 2018, 13, e0190839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohórquez, D.V.; Chandra, R.; Samsa, L.A.; Vigna, S.R.; Liddle, R.A. Characterization of Basal Pseudopod-like Processes in Ileal and Colonic PYY Cells. J. Mol. Histol. 2011, 42, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Gall-David, S.L.; Zenbaa, N.; Bouchard, D.; Lavault, M.-T.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A.; Bousarghin, L. Hypermutator Salmonella Heidelberg Induces an Early Cell Death in Epithelial Cells. Vet. Microbiol. 2015, 180, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Harel, E.; Rubinstein, A.; Nissan, A.; Khazanov, E.; Milbauer, M.N.; Barenholz, Y.; Tirosh, B. Enhanced Transferrin Receptor Expression by Proinflammatory Cytokines in Enterocytes as a Means for Local Delivery of Drugs to Inflamed Gut Mucosa. PLoS ONE 2011, 6, e24202. [Google Scholar] [CrossRef] [Green Version]
- Reimer, R.A.; Darimont, C.; Gremlich, S.; Nicolas-Métral, V.; Rüegg, U.T.; Macé, K. A Human Cellular Model for Studying the Regulation of Glucagon-like Peptide-1 Secretion. Endocrinology 2001, 142, 4522–4528. [Google Scholar] [CrossRef] [PubMed]
- Anini, Y.; Brubaker, P.L. Role of Leptin in the Regulation of Glucagon-like Peptide-1 Secretion. Diabetes 2003, 52, 252–259. [Google Scholar] [CrossRef] [Green Version]
- During, M.J.; Cao, L.; Zuzga, D.S.; Francis, J.S.; Fitzsimons, H.L.; Jiao, X.; Bland, R.J.; Klugmann, M.; Banks, W.A.; Drucker, D.J.; et al. Glucagon-like Peptide-1 Receptor Is Involved in Learning and Neuroprotection. Nat. Med. 2003, 9, 1173–1179. [Google Scholar] [CrossRef]
- Kuhre, R.E.; Albrechtsen, N.J.W.; Deacon, C.F.; Balk-Møller, E.; Rehfeld, J.F.; Reimann, F.; Gribble, F.M.; Holst, J.J. Peptide Production and Secretion in GLUTag, NCI-H716 and STC-1 Cells: A Comparison to Native L-Cells. J. Mol. Endocrinol. 2016, 56, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Sjölund, K.; Sandén, G.; Håkanson, R.; Sundler, F. Endocrine Cells in Human Intestine: An Immunocytochemical Study. Gastroenterology 1983, 85, 1120–1130. [Google Scholar] [CrossRef]
- Xie, N.; Huang, X.; Yang, C.; Dai, M.; Cai, L.; Deng, S.; Hardiman, P.J.; Zhou, J. Artificial Sweeteners Affect the Glucose Transport Rate in the Caco-2/NCI-H716 Co-Culture Model. J. Sci. Food Agric. 2020, 100, 4887–4892. [Google Scholar] [CrossRef]
- Tolhurst, G.; Reimann, F.; Gribble, F.M. Nutritional Regulation of Glucagon-like Peptide-1 Secretion. J. Physiol. 2009, 587, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial Signature of Dysbiosis in Gut Microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Ménard, S.; Lacroix-Lamandé, S.; Ehrhardt, K.; Yan, J.; Grassl, G.A.; Wiedemann, A. Cross-Talk Between the Intestinal Epithelium and Salmonella Typhimurium. Front. Microbiol. 2022, 13, 906238. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.-C. Regulation of Salmonella Flagellin-Induced Interleukin-8 in Intestinal Epithelial Cells by Muramyl Dipeptide. Cell. Immunol. 2012, 278, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Quan, Y.; Song, K.; Zhang, Y.; Zhu, C.; Shen, Z.; Wu, S.; Luo, W.; Tan, B.; Yang, Z.; Wang, X. Roseburia Intestinalis-Derived Flagellin Is a Negative Regulator of Intestinal Inflammation. Biochem. Biophys. Res. Commun. 2018, 501, 791–799. [Google Scholar] [CrossRef]
- Chen, L.; Yang, M.; Zhu, W.; Su, Y.; Li, D.; Wang, T. Multi-Omics Analysis After Vaginal Administration of Bacteroides Fragilis in Chickens. Front. Microbiol. 2022, 13, 846011. [Google Scholar] [CrossRef]
- Chan, J.L.; Wu, S.; Geis, A.L.; Chan, G.V.; Gomes, T.A.M.; Beck, S.E.; Wu, X.; Fan, H.; Tam, A.J.; Chung, L.; et al. Non-Toxigenic Bacteroides Fragilis (NTBF) Administration Reduces Bacteria-Driven Chronic Colitis and Tumor Development Independent of Polysaccharide A. Mucosal Immunol. 2019, 12, 164–177. [Google Scholar] [CrossRef] [Green Version]
- Qu, D.; Sun, F.; Feng, S.; Yu, L.; Tian, F.; Zhang, H.; Chen, W.; Zhai, Q. Protective Effects of Bacteroides Fragilis against Lipopolysaccharide-Induced Systemic Inflammation and Their Potential Functional Genes. Food Funct. 2022, 13, 1015–1025. [Google Scholar] [CrossRef]
- Pathak, P.; Xie, C.; Nichols, R.G.; Ferrell, J.M.; Boehme, S.; Krausz, K.W.; Patterson, A.D.; Gonzalez, F.J.; Chiang, J.Y.L. Intestine Farnesoid X Receptor Agonist and the Gut Microbiota Activate G-Protein Bile Acid Receptor-1 Signaling to Improve Metabolism. Hepatology 2018, 68, 1574–1588. [Google Scholar] [CrossRef] [Green Version]
- Princen, H.M.; Meijer, P. Maintenance of Bile Acid Synthesis and Cholesterol 7 Alpha-Hydroxylase Activity in Cultured Rat Hepatocytes. Biochem. J. 1990, 272, 273–275. [Google Scholar] [CrossRef] [Green Version]
- Gautier, T.; Olivieiro, N.; Ferron, S.; Le Pogam, P.; David-Le Gall, S.; Sauvager, A.; Leroyer, P.; Cannie, I.; Dion, S.; Sweidan, A.; et al. Bacteroides fragilis derived metabolites, identified by molecular networking, decrease Salmonella virulence in mice model. Front. Microbiol. 2022, 13, 1023315. [Google Scholar] [CrossRef]
- Yang, J.-Y.; Lee, Y.-S.; Kim, Y.; Lee, S.-H.; Ryu, S.; Fukuda, S.; Hase, K.; Yang, C.-S.; Lim, H.S.; Kim, M.-S.; et al. Gut Commensal Bacteroides Acidifaciens Prevents Obesity and Improves Insulin Sensitivity in Mice. Mucosal Immunol. 2017, 10, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Huang, X.; Ma, Y.; Gao, M.; Wang, O.; Gao, T.; Shen, Y.; Liu, X. Interleukin-8 Regulates Endothelial Permeability by down-Regulation of Tight Junction but Not Dependent on Integrins Induced Focal Adhesions. Int. J. Biol. Sci. 2013, 9, 966–979. [Google Scholar] [CrossRef]
- Kasahara, K.; Krautkramer, K.A.; Org, E.; Romano, K.A.; Kerby, R.L.; Vivas, E.I.; Mehrabian, M.; Denu, J.M.; Bäckhed, F.; Lusis, A.J.; et al. Interactions between Roseburia Intestinalis and Diet Modulate Atherogenesis in a Murine Model. Nat. Microbiol. 2018, 3, 1461–1471. [Google Scholar] [CrossRef] [PubMed]
- Navab-Moghadam, F.; Sedighi, M.; Khamseh, M.E.; Alaei-Shahmiri, F.; Talebi, M.; Razavi, S.; Amirmozafari, N. The Association of Type II Diabetes with Gut Microbiota Composition. Microb. Pathog. 2017, 110, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; van der Beek, C.M.; Jocken, J.W.E.; Goossens, G.H.; Holst, J.J.; Olde Damink, S.W.M.; Lenaerts, K.; Dejong, C.H.C.; Blaak, E.E. Colonic Infusions of Short-Chain Fatty Acid Mixtures Promote Energy Metabolism in Overweight/Obese Men: A Randomized Crossover Trial. Sci. Rep. 2017, 7, 2360. [Google Scholar] [CrossRef]
- Christiansen, C.B.; Gabe, M.B.N.; Svendsen, B.; Dragsted, L.O.; Rosenkilde, M.M.; Holst, J.J. The Impact of Short-Chain Fatty Acids on GLP-1 and PYY Secretion from the Isolated Perfused Rat Colon. Am. J. Physiol.-Gastrointest. Liver Physiol. 2018, 315, G53–G65. [Google Scholar] [CrossRef] [Green Version]
- Covasa, M.; Stephens, R.W.; Toderean, R.; Cobuz, C. Intestinal Sensing by Gut Microbiota: Targeting Gut Peptides. Front. Endocrinol. 2019, 10, 82. [Google Scholar] [CrossRef] [Green Version]
- Pizarroso, N.A.; Fuciños, P.; Gonçalves, C.; Pastrana, L.; Amado, I.R. A Review on the Role of Food-Derived Bioactive Molecules and the Microbiota-Gut-Brain Axis in Satiety Regulation. Nutrients 2021, 13, 632. [Google Scholar] [CrossRef]
- Xia, F.; Wen, L.-P.; Ge, B.-C.; Li, Y.-X.; Li, F.-P.; Zhou, B.-J. Gut Microbiota as a Target for Prevention and Treatment of Type 2 Diabetes: Mechanisms and Dietary Natural Products. World J. Diabetes 2021, 12, 1146–1163. [Google Scholar] [CrossRef]
- Karra, E.; Chandarana, K.; Batterham, R.L. The Role of Peptide YY in Appetite Regulation and Obesity. J. Physiol. 2009, 587, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Dominique, M.; Lucas, N.; Legrand, R.; Bouleté, I.-M.; Bôle-Feysot, C.; Deroissart, C.; Léon, F.; Nobis, S.; do Rego, J.-C.; Lambert, G.; et al. Effects of Bacterial CLPB Protein Fragments on Food Intake and PYY Secretion. Nutrients 2021, 13, 2223. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Encoded Protein | Left Primer | Right Primer |
|---|---|---|---|
| tbp | TATA Box protein | CCGGAATCCCTATCTTTAGTCC | GGGTCAGTCCAGTGCCATAAG |
| MUC2 | Mucin- 2 | CAGCACCGATTGCTGACTTG | GCTGGTCATCTCAATGGCAG |
| SI | Sucrase isomaltase | CATCCTACCATGTCAAGAGCCA | GCTTGTTAAGGTGGTCTGGTTT |
| MMP15 | Metalloprotease 15 | ACAACTATCCCATGCCCATC | ACCTGTCCTCTTGGAAGAAG |
| PYY | Peptide Tyrosine-Tyrosine | GCCTTGACCACAGTGCTTC | CTCTTTTCCCATACCGCTGC |
| GCG | Proglucagon | ACATTGCCAAACGTCACGA | GCGACCTCTTCTGGGAAATC |
| OCLN | Occludin | CCAATGTCGAGGAGTGGG | CGCTGCTGTAACGAGGCT |
| ZO-1 | Zonula occludens 1 | ATCCCTCAAGGAGCCATTC | CACTTGTTTTGCCAGGTTTTA |
| IL-8 | Interleukin-8 | AGACTTCCAAGCTGGCCGTGGCT | TCTCAGCCCTCTTCAAAAACTTCTC |
| CHGA | Chromogranin-A | CTACGCGCCTTGTCTCCTAC | AGTTGTGCCCAGTGGATAGG |
| TFRC | Transferrine receptor | GCTTTCCCTTTCCTTGCATATTCT | GGTGGTACCCAAATAAGGATAATCTGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gautier, T.; Fahet, N.; Tamanai-Shacoori, Z.; Oliviero, N.; Blot, M.; Sauvager, A.; Burel, A.; Gall, S.D.-L.; Tomasi, S.; Blat, S.; et al. Roseburia intestinalis Modulates PYY Expression in a New a Multicellular Model including Enteroendocrine Cells. Microorganisms 2022, 10, 2263. https://doi.org/10.3390/microorganisms10112263
Gautier T, Fahet N, Tamanai-Shacoori Z, Oliviero N, Blot M, Sauvager A, Burel A, Gall SD-L, Tomasi S, Blat S, et al. Roseburia intestinalis Modulates PYY Expression in a New a Multicellular Model including Enteroendocrine Cells. Microorganisms. 2022; 10(11):2263. https://doi.org/10.3390/microorganisms10112263
Chicago/Turabian StyleGautier, Thomas, Nelly Fahet, Zohreh Tamanai-Shacoori, Nolwenn Oliviero, Marielle Blot, Aurélie Sauvager, Agnes Burel, Sandrine David-Le Gall, Sophie Tomasi, Sophie Blat, and et al. 2022. "Roseburia intestinalis Modulates PYY Expression in a New a Multicellular Model including Enteroendocrine Cells" Microorganisms 10, no. 11: 2263. https://doi.org/10.3390/microorganisms10112263