
Phylogenomic Analyses and Molecular Signatures Elucidating the Evolutionary Relationships amongst the Chlorobia and Ignavibacteria Species: Robust Demarcation of Two Family-Level Clades within the Order Chlorobiales and Proposal for the Family Chloroherpetonaceae fam. nov


Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of Phylogenetic Trees
2.2. Identification of Conserved Signature Indels
3. Results
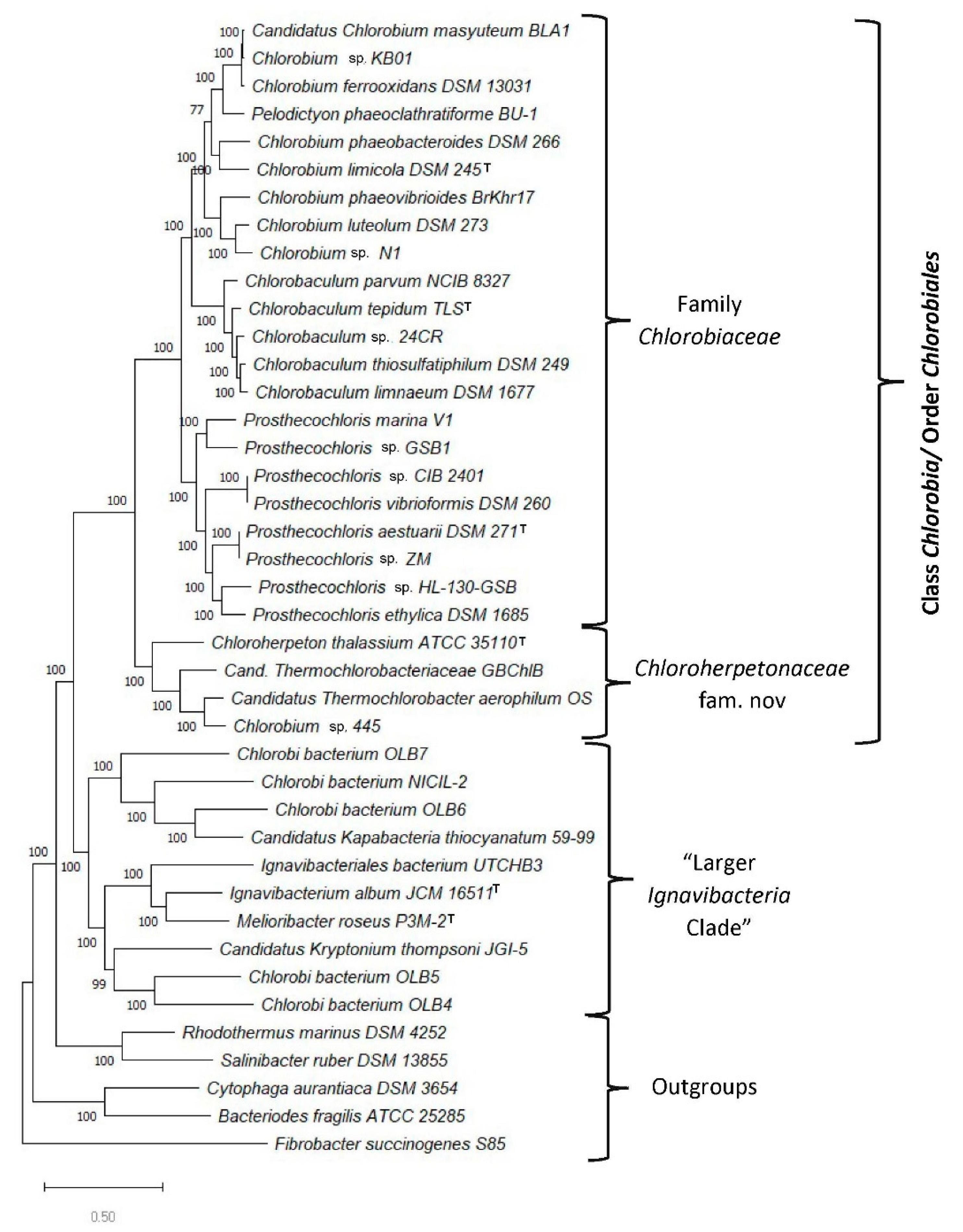
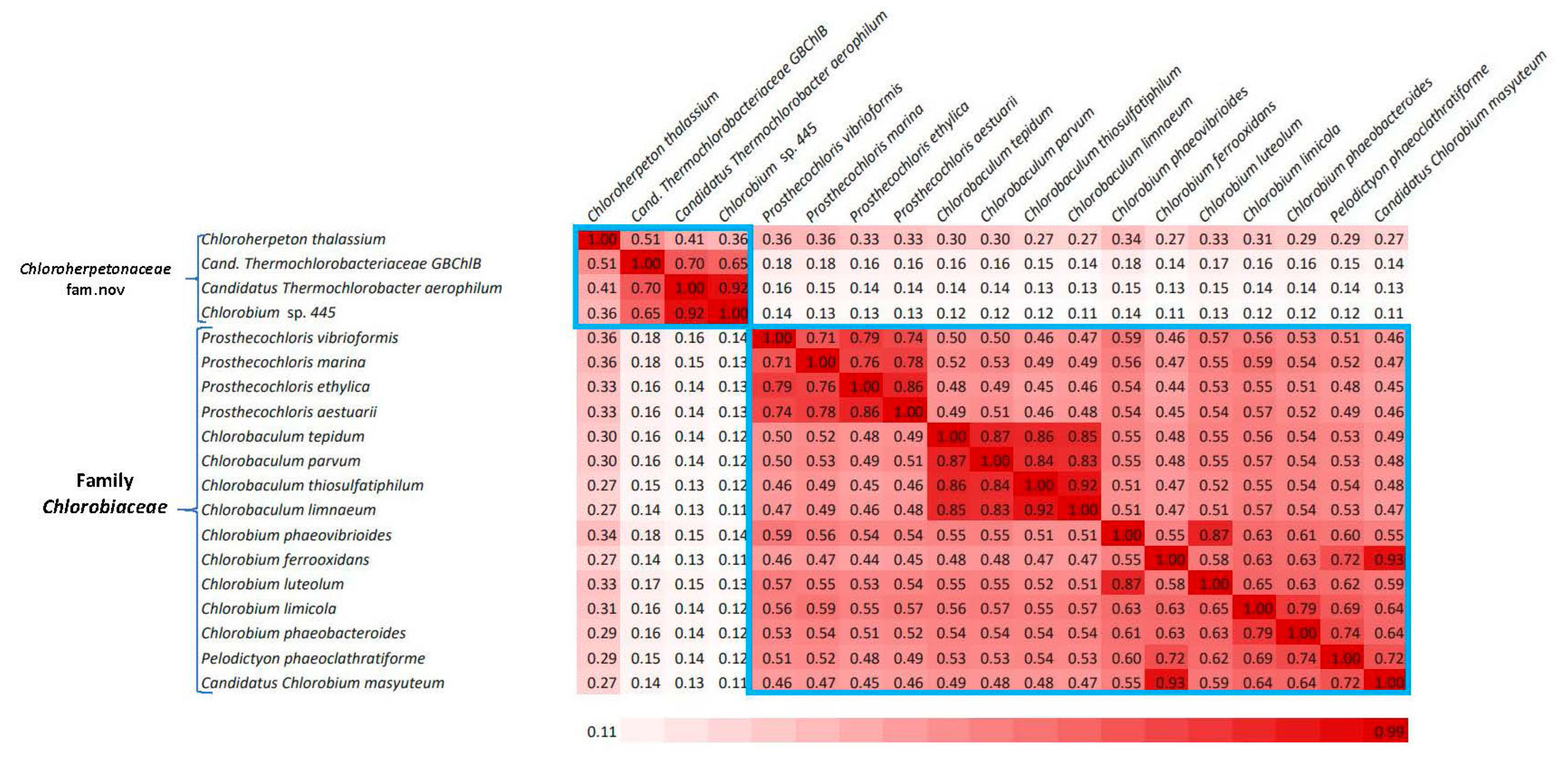
3.1. Phylogenetic Analysis of the Chlorobia/Ignavibacteria Species Based on Genome Sequences
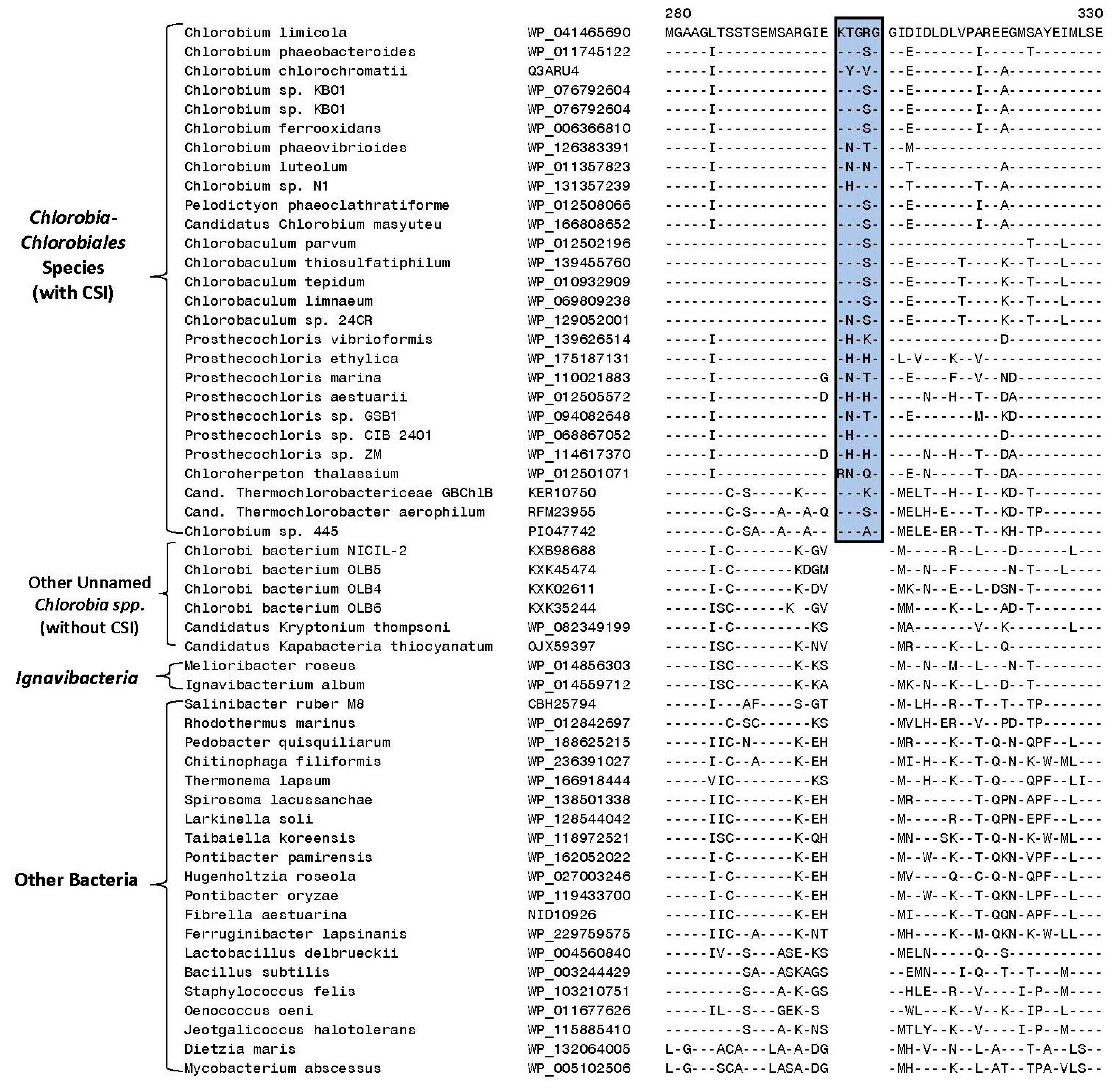
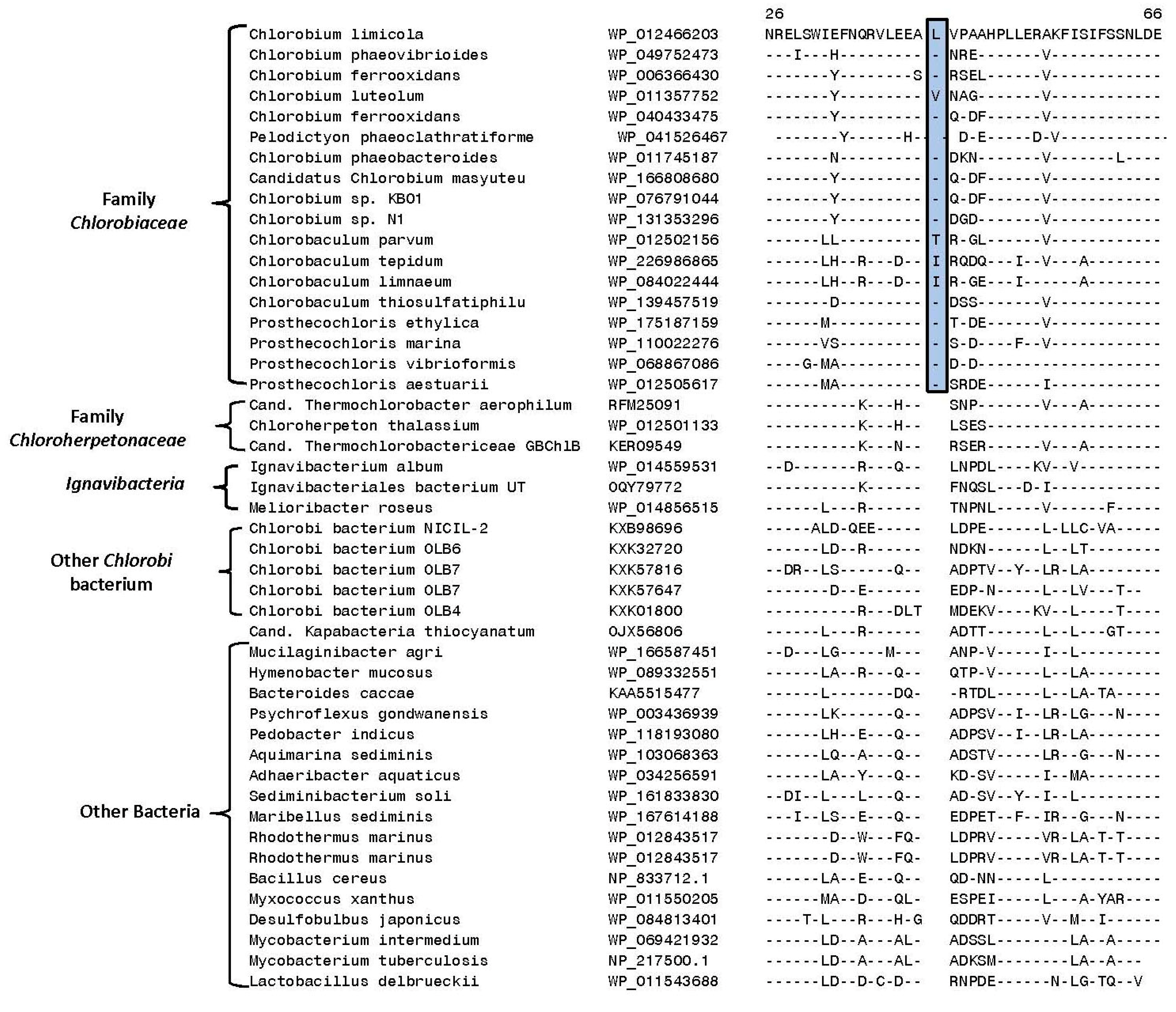
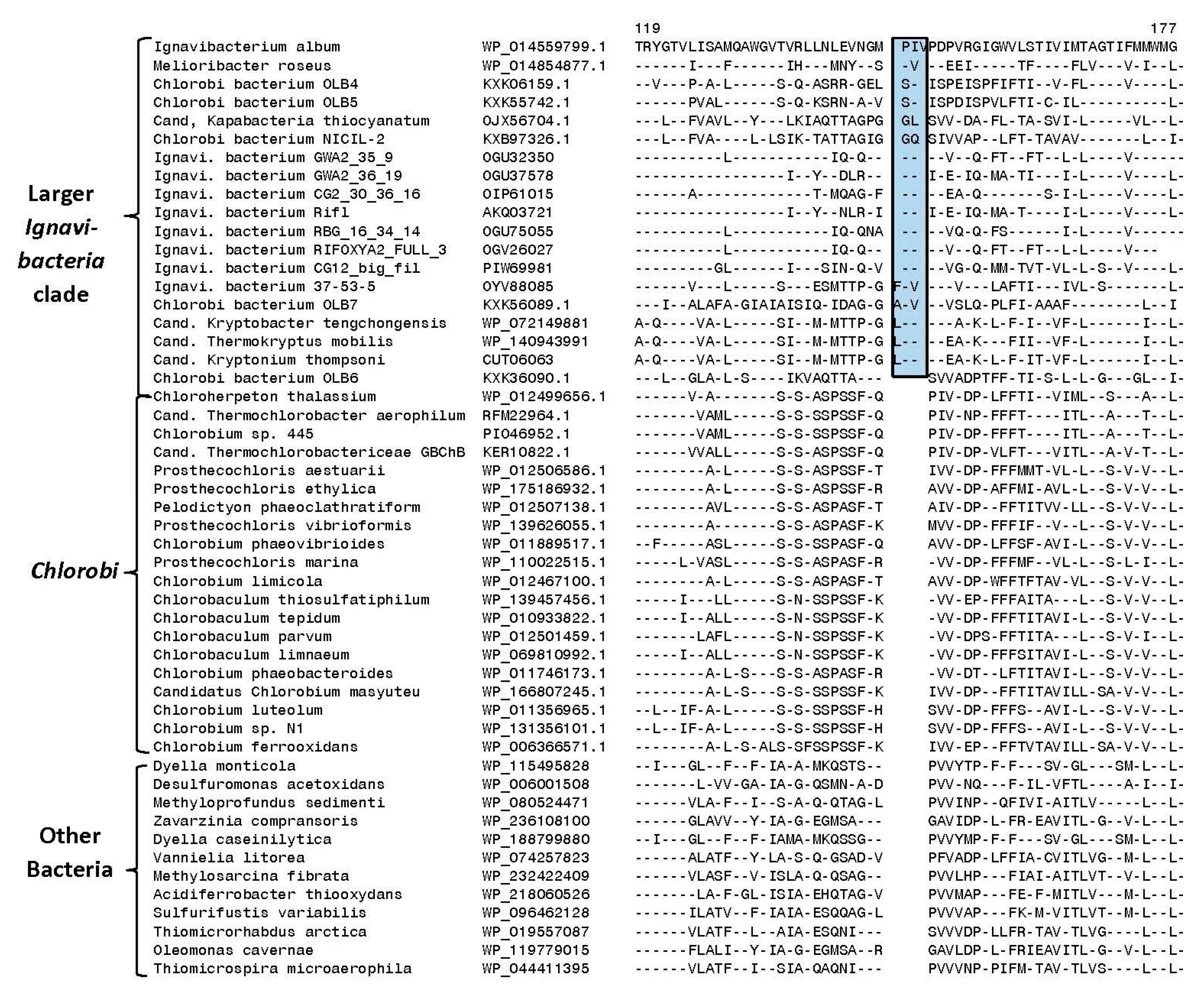
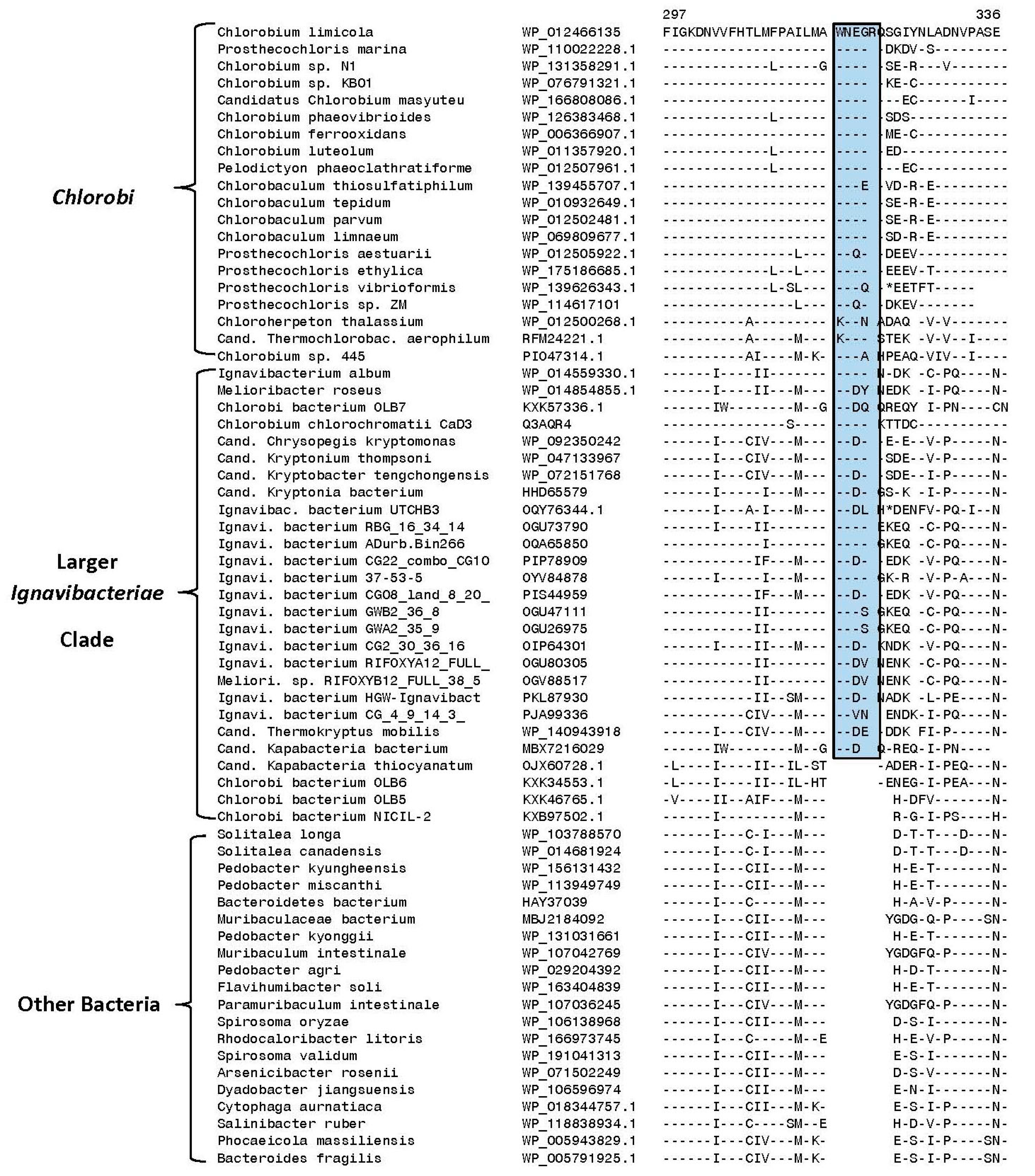
3.2. Identification of Molecular Markers Specific for the Main Clades of Chlorobia Species
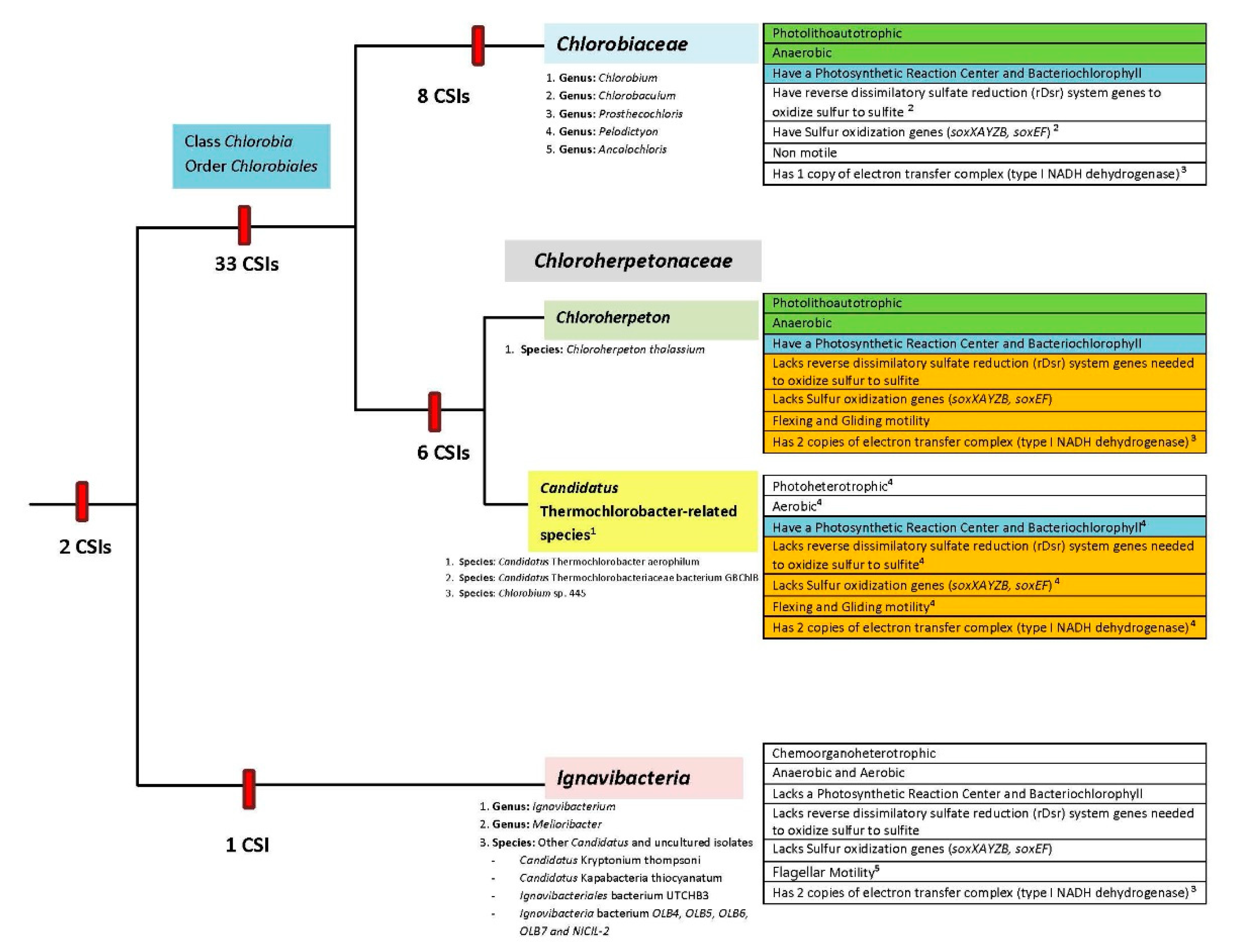
4. Discussion
- Description of the Class Chlorobia class nov. (Garrity and Holt 2001, 601EP)
- Emended Description of the order Chlorobiales Gibbons and Murray 1978 (Approved Lists 1980)
- Emended Description of the family Chlorobiaceae Copeland 1956 (Approved Lists 1980)
- Description of the family Chloroherpetonaceae fam. nov.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pfennig, N. Chlorobium. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Pfennig, N. Chlorobium phaeobacteroides nov. spec. and C. phaeovibrioides nov. spec., two new species of green sulfur bacteria. Arch. Mikrobiol. 1968, 63, 224–226. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Thiel, V. Phylogeny and taxonomy of Chlorobiaceae. Photosynth. Res. 2010, 104, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, J.F. The Family Chlorobiaceae. In The Prokaryotes—Other Major Lineages of Bacteria and the Archaea, 4th ed.; Rosenberg, E., De Long, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Overmann, J. The Family Chlorobiaceae. In The Prokaryotes, 3rd ed.; Springer: New York, NY, USA, 2003. [Google Scholar]
- Kondratieva, E.N.; Pfennig, N.; Truper, H.G. The Phototrophic Prokaryotes. In The Prokaryotes, 2nd ed.; Balows, A., Truper, H.G., Dworkin, M., Harder, W., Schleifer, K.H., Eds.; Springer-Verlag: New York, NY, USA, 1992; Volume 1, pp. 312–330. [Google Scholar]
- Iino, T.; Mori, K.; Uchino, Y.; Nakagawa, T.; Harayama, S.; Suzuki, K. Ignavibacterium album gen. nov., sp. nov., a moderately thermophilic anaerobic bacterium isolated from microbial mats at a terrestrial hot spring and proposal of Ignavibacteria classis nov., for a novel lineage at the periphery of green sulfur bacteria. Int. J. Syst. Evol. Microbiol. 2010, 60, 1376–1382. [Google Scholar] [CrossRef] [Green Version]
- Skerman, V.B.D.; McGowan, V.; Sneath, P.H.A. Approved lists of bacterial names. Int. J. Syst. Bacteriol. 1980, 30, 225–420. [Google Scholar] [CrossRef] [Green Version]
- Truper, H.G.; Pfennig, N. The Family Chlorobiaceae. In The Prokaryotes, 2nd ed.; Balows, A., Truper, H.G., Dworkin, M., Harder, W., Schleifer, K.H., Eds.; Springer: New York, NY, USA, 1992; pp. 3583–3592. [Google Scholar]
- Gibbons, N.E.; Murray, R.G.E. Proposals Concerning the Higher Taxa of Bacteri. Int. J. Syst. Bacteriol. 1978, 28, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Parte, A.C. LPSN—The List of Prokaryotic Names with Standing in Nomenclature. Int. J. Syst. Evol. Microbiol. 2018, 68, 1825–1829. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, J.F. Phylogenetic taxonomy of the family Chlorobiaceae on the basis of 16S rRNA and fmo (Fenna-Matthews-Olson protein) gene sequences. Int. J. Syst. Evol. Microbiol. 2003, 53, 941–951. [Google Scholar] [CrossRef] [Green Version]
- Gibson, J.; Pfennig, N.; Waterbury, J.B. Chloroherpeton thalassium gen. nov. et spec. nov., a non-filamentous, flexing and gliding green sulfur bacterium. Arch. Microbiol. 1984, 138, 96–101. [Google Scholar] [CrossRef]
- Gorlenko, V.M.; Lebedeva, E.V. New green sulfur bacteria with apophyses. Mikrobiologiia 1971, 40, 1035–1039. [Google Scholar]
- Alexander, B.; Andersen, J.H.; Cox, R.P.; Imhoff, J.F. Phylogeny of green sulfur bacteria on the basis of gene sequences of 16S rRNA and of the Fenna-Matthews-Olson protein. Arch. Microbiol. 2002, 178, 131–140. [Google Scholar] [CrossRef]
- Panwar, P.; Allen, M.A.; Williams, T.J.; Haque, S.; Brazendale, S.; Hancock, A.M.; Paez-Espino, D.; Cavicchioli, R. Remarkably coherent population structure for a dominant Antarctic Chlorobium species. Microbiome 2021, 9, 231. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, J.M.; Tran, N.; Schiff, S.L.; Venkiteswaran, J.J.; Molot, L.A.; Tank, M.; Hanada, S.; Neufeld, J.D. Anoxygenic photosynthesis and iron-sulfur metabolic potential of Chlorobia populations from seasonally anoxic Boreal Shield lakes. ISME J. 2020, 14, 2732–2747. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S.; Lorenzini, E. Phylogeny and molecular signatures (conserved proteins and indels) that are specific for the Bacteroidetes and Chlorobi species. BMC Evol. Biol. 2007, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.S. The Phylogeny and Signature Sequences characteristics of Fibrobacters, Chlorobi and Bacteroidetes. Crit. Rev. Microbiol. 2004, 30, 123–143. [Google Scholar] [CrossRef]
- Gupta, R.S. Impact of genomics on the understanding of microbial evolution and classification: The importance of Darwin’s views on classification. FEMS Microbiol. Rev. 2016, 40, 520–553. [Google Scholar] [CrossRef]
- Kadnikov, V.V.; Mardanov, A.V.; Podosokorskaya, O.A.; Gavrilov, S.N.; Kublanov, I.V.; Beletsky, A.V.; Bonch-Osmolovskaya, E.A.; Ravin, N.V. Genomic analysis of Melioribacter roseus, facultatively anaerobic organotrophic bacterium representing a novel deep lineage within Bacteriodetes/Chlorobi group. PLoS ONE 2013, 8, e53047. [Google Scholar] [CrossRef] [Green Version]
- Podosokorskaya, O.A.; Kadnikov, V.V.; Gavrilov, S.N.; Mardanov, A.V.; Merkel, A.Y.; Karnachuk, O.V.; Ravin, N.V.; Bonch-Osmolovskaya, E.A.; Kublanov, I.V. Characterization of Melioribacter roseus gen. nov., sp. nov., a novel facultatively anaerobic thermophilic cellulolytic bacterium from the class Ignavibacteria, and a proposal of a novel bacterial phylum Ignavibacteriae. Environ. Microbiol. 2013, 15, 1759–1771. [Google Scholar] [CrossRef]
- Iino, T. Ignavibacteriae. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2018; p. 1. [Google Scholar] [CrossRef]
- Roy, C.; Bakshi, U.; Rameez, M.J.; Mandal, S.; Haldar, P.K.; Pyne, P.; Ghosh, W. Phylogenomics of an uncultivated, aerobic and thermophilic, photoheterotrophic member of Chlorobia sheds light into the evolution of the phylum Chlorobi. Comput. Biol. Chem. 2019, 80, 206–216. [Google Scholar] [CrossRef]
- Hiras, J.; Wu, Y.W.; Eichorst, S.A.; Simmons, B.A.; Singer, S.W. Refining the phylum Chlorobi by resolving the phylogeny and metabolic potential of the representative of a deeply branching, uncultivated lineage. ISME J. 2016, 10, 833–845. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Klatt, C.G.; Ludwig, M.; Rusch, D.B.; Jensen, S.I.; Kuhl, M.; Ward, D.M.; Bryant, D.A. ‘Candidatus Thermochlorobacter aerophilum:’ an aerobic chlorophotoheterotrophic member of the phylum Chlorobi defined by metagenomics and metatranscriptomics. ISME J. 2012, 6, 1869–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Seshadri, R.; Varghese, N.J.; Eloe-Fadrosh, E.A.; Meier-Kolthoff, J.P.; Goker, M.; Coates, R.C.; Hadjithomas, M.; Pavlopoulos, G.A.; Paez-Espino, D.; et al. 1003 reference genomes of bacterial and archaeal isolates expand coverage of the tree of life. Nat. Biotechnol. 2017, 35, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Whitman, W.B. Genome sequences as the type material for taxonomic descriptions of prokaryotes 1. Syst. Appl. Microbiol. 2015, 38, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; McCluskey, K.; Desmeth, P.; Liu, S.; Hideaki, S.; Yin, Y.; Moriya, O.; Itoh, T.; Kim, C.Y.; Lee, J.S.; et al. The global catalogue of microorganisms 10K type strain sequencing project: Closing the genomic gaps for the validly published prokaryotic and fungi species. Gigascience 2018, 7, giy026. [Google Scholar] [CrossRef] [PubMed]
- Sayers, E.W.; Agarwala, R.; Bolton, E.E.; Brister, J.R.; Canese, K.; Clark, K.; Connor, R.; Fiorini, N.; Funk, K.; Hefferon, T.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2019, 47, D23–D28. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Chuvochina, M.; Waite, D.W.; Rinke, C.; Skarshewski, A.; Chaumeil, P.A.; Hugenholtz, P. A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat. Biotechnol. 2018, 36, 996–1004. [Google Scholar] [CrossRef]
- Rokas, A.; Williams, B.L.; King, N.; Carroll, S.B. Genome-scale approaches to resolving incongruence in molecular phylogenies. Nature 2003, 425, 798–804. [Google Scholar] [CrossRef]
- Wu, D.; Jospin, G.; Eisen, J.A. Systematic identification of gene families for use as “markers” for phylogenetic and phylogeny-driven ecological studies of bacteria and archaea and their major subgroups 1. PLoS ONE 2013, 8, e77033. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.S.; Patel, S.; Saini, N.; Chen, S. Robust demarcation of 17 distinct Bacillus species clades, proposed as novel Bacillaceae genera, by phylogenomics and comparative genomic analyses: Description of Robertmurraya kyonggiensis sp. nov. and proposal for an emended genus Bacillus limiting it only to the members of the Subtilis and Cereus clades of species. Int. J. Syst. Evol. Microbiol. 2020, 70, 5753–5798. [Google Scholar]
- Hugenholtz, P.; Chuvochina, M.; Oren, A.; Parks, D.H.; Soo, R.M. Prokaryotic taxonomy and nomenclature in the age of big sequence data. ISME J. 2021, 15, 1879–1892. [Google Scholar] [CrossRef]
- Gupta, R.S. Microbial Taxonomy: How and Why Name Changes Occur and Their Significance for (Clinical) Microbiology. Clin. Chem. 2021, 68, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Gupta, R.S. A Phylogenomics and Comparative Genomic Framework for Resolving the Polyphyly of the Genus Bacillus: Proposal for six new genera of Bacillus species, Peribacillus gen. nov., Cytobacillus gen. nov., Mesobacillus gen. nov., Neobacillus gen. nov., Metabacillus gen. nov. and Alkalihalobacillus gen. nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 406–438. [Google Scholar]
- Gupta, R.S.; Lo, B.; Son, J. Phylogenomics and Comparative Genomic Studies Robustly Support Division of the Genus Mycobacterium into an Emended Genus Mycobacterium and Four Novel Genera. Front. Microbiol. 2018, 9, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar]
- Rokas, A.; Holland, P.W. Rare genomic changes as a tool for phylogenetics. Trends Ecol. Evol. 2000, 15, 454–459. [Google Scholar] [CrossRef]
- Gupta, R.S. Molecular signatures for the main phyla of photosynthetic bacteria and their subgroups. Photosynth. Res. 2010, 104, 357–372. [Google Scholar] [CrossRef]
- Gupta, R.S.; Chander, P.; George, S. Phylogenetic framework and molecular signatures for the class Chloroflexi and its different clades; proposal for division of the class Chloroflexia class. nov. [corrected] into the suborder Chloroflexineae subord. nov., consisting of the emended family Oscillochloridaceae and the family Chloroflexaceae fam. nov., and the suborder Roseiflexineae subord. nov., containing the family Roseiflexaceae fam. nov. Antonie Van Leeuwenhoek 2013, 103, 99–119. [Google Scholar]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Capella-Gutierrez, S.; Silla-Martinez, J.M.; Gabaldon, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2--approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Whelan, S.; Goldman, N. A general empirical model of protein evolution derived from multiple protein families using a maximum-likelihood approach. Mol. Biol. Evol. 2001, 18, 691–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Thompson, C.C.; Amaral, G.R.; Campeao, M.; Edwards, R.A.; Polz, M.F.; Dutilh, B.E.; Ussery, D.W.; Sawabe, T.; Swings, J.; Thompson, F.L. Microbial taxonomy in the post-genomic era: Rebuilding from scratch? 3. Arch. Microbiol. 2015, 197, 359–370. [Google Scholar] [CrossRef]
- Gupta, R.S. Distinction between Borrelia and Borreliella is more robustly supported by molecular and phenotypic characteristics than all other neighbouring prokaryotic genera: Response to Margos’ et al. “The genus Borrelia reloaded” (PLoS ONE 13(12): e0208432). PLoS ONE 2019, 14, e0221397. [Google Scholar] [CrossRef]
- Qin, Q.L.; Xie, B.B.; Zhang, X.Y.; Chen, X.L.; Zhou, B.C.; Zhou, J.; Oren, A.; Zhang, Y.Z. A proposed genus boundary for the prokaryotes based on genomic insights. J. Bacteriol. 2014, 196, 2210–2215. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glockner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Gupta, R.S. Identification of Conserved Indels that are Useful for Classification and Evolutionary Studies. In Bacterial Taxonomy, Methods in Microbiology Volume 41; Goodfellow, M., Sutcliffe, I.C., Chun, J., Eds.; Elsevier: London, UK, 2014; pp. 153–182. [Google Scholar]
- Naushad, H.S.; Lee, B.; Gupta, R.S. Conserved signature indels and signature proteins as novel tools for understanding microbial phylogeny and systematics: Identification of molecular signatures that are specific for the phytopathogenic genera Dickeya, Pectobacterium and Brenneria. Int. J. Syst. Evol. Microbiol. 2014, 64, 366–383. [Google Scholar] [CrossRef] [Green Version]
- Gibson, J. Chloroherpeton. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Kantor, R.S.; van Zyl, A.W.; van Hille, R.P.; Thomas, B.C.; Harrison, S.T.; Banfield, J.F. Bioreactor microbial ecosystems for thiocyanate and cyanide degradation unravelled with genome-resolved metagenomics. Environ. Microbiol. 2015, 17, 4929–4941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eloe-Fadrosh, E.A.; Paez-Espino, D.; Jarett, J.; Dunfield, P.F.; Hedlund, B.P.; Dekas, A.E.; Grasby, S.E.; Brady, A.L.; Dong, H.; Briggs, B.R.; et al. Global metagenomic survey reveals a new bacterial candidate phylum in geothermal springs. Nat. Commun. 2016, 7, 10476. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, V.; Gupta, R.S. Phylum Thermotogae. In The Prokaryotes—Other Major Lineages of Bacteria and the Archaea, 4th ed.; Rosenberg, E., DeLong, E., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: New York, NY, USA, 2014; Volume 11, pp. 989–1015. [Google Scholar]
- Jiang, L.; Wang, D.; Kim, J.S.; Lee, J.H.; Kim, D.H.; Kim, S.W.; Lee, J. Reclassification of genus Izhakiella into the family Erwiniaceae based on phylogenetic and genomic analyses. Int. J. Syst. Evol. Microbiol. 2020, 70, 3541–3546. [Google Scholar] [CrossRef] [PubMed]
- Dobritsa, A.P.; Samadpour, M. Reclassification of Burkholderia insecticola as Caballeronia insecticola comb. nov. and reliability of conserved signature indels as molecular synapomorphies. Int. J. Syst. Evol. Microbiol. 2019, 69, 2057–2063. [Google Scholar] [CrossRef]
- Hohmann-Marriott, M.F.; Blankenship, R.E. Evolution of photosynthesis. Annu. Rev. Plant. Biol. 2011, 62, 515–548. [Google Scholar] [CrossRef] [Green Version]
- Bryant, D.A.; Costas, A.M.; Maresca, J.A.; Chew, A.G.; Klatt, C.G.; Bateson, M.M.; Tallon, L.J.; Hostetler, J.; Nelson, W.C.; Heidelberg, J.F.; et al. Candidatus Chloracidobacterium thermophilum: An aerobic phototrophic Acidobacterium. Science 2007, 317, 523–526. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Frigaard, N.U.; Vogl, K.; Iino, T.; Ohkuma, M.; Overmann, J.; Bryant, D.A. Complete Genome of Ignavibacterium album, a Metabolically Versatile, Flagellated, Facultative Anaerobe from the Phylum Chlorobi. Front. Microbiol. 2012, 3, 185. [Google Scholar] [CrossRef] [Green Version]
- Gregersen, L.H.; Bryant, D.A.; Frigaard, N.U. Mechanisms and evolution of oxidative sulfur metabolism in green sulfur bacteria. Front. Microbiol. 2011, 2, 116. [Google Scholar] [CrossRef] [Green Version]
- Holkenbrink, C.; Barbas, S.O.; Mellerup, A.; Otaki, H.; Frigaard, N.U. Sulfur globule oxidation in green sulfur bacteria is dependent on the dissimilatory sulfite reductase system. Microbiology 2011, 157, 1229–1239. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Inferring Phylogenies; Sinauer Associates, Inc.: Sunderland, MA, USA, 2004. [Google Scholar]
- Baldauf, S.L. Phylogeny for the faint of heart: A tutorial. Trends Genet. 2003, 19, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.S. Protein phylogenies and signature sequences: A reappraisal of evolutionary relationships among archaebacteria, eubacteria, and eukaryotes. Microbiol. Mol. Biol. Rev. 1998, 62, 1435–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbour, A.G.; Adeolu, M.; Gupta, R.S. Division of the genus Borrelia into two genera (corresponding to Lyme disease and relapsing fever groups) reflects their genetic and phenotypic distinctiveness and will lead to a better understanding of these two groups of microbes (Margos et al. (2016) There is inadequate evidence to support the division of the genus Borrelia. Int. J. Syst. Evol. Microbiol. doi: 10.1099/ijsem.0.001717). Int. J. Syst. Evol. Microbiol. 2017, 67, 2058–2067. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S. Protein signatures (molecular synapomorphies) that are distinctive characteristics of the major cyanobacterial clades. Int. J. Syst. Evol. Microbiol. 2009, 59, 2510–2526. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Gupta, R.S. Conserved indels in protein sequences that are characteristic of the phylum Actinobacteria. Int. J. Syst. Evol. Microbiol. 2005, 55, 2401–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmod, N.Z.; Gupta, R.S.; Shah, H.N. Identification of a Bacillus anthracis specific indel in the yeaC gene and development of a rapid pyrosequencing assay for distinguishing B. anthracis from the B. cereus group. J. Microbiol. Methods 2011, 87, 278–285. [Google Scholar] [CrossRef]
- Wong, S.Y.; Paschos, A.; Gupta, R.S.; Schellhorn, H.E. Insertion/deletion-based approach for the detection of Escherichia coli O157:H7 in freshwater environments. Environ. Sci. Technol. 2014, 48, 11462–11470. [Google Scholar] [CrossRef]
- Singh, B.; Gupta, R.S. Conserved inserts in the Hsp60 (GroEL) and Hsp70 (DnaK) proteins are essential for cellular growth. Mol. Genet. Genom. 2009, 281, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Khadka, B.; Gupta, R.S. Identification of a Conserved 8 aa Insert in the PIP5K Protein in the Saccharomycetaceae family of Fungi and the Molecular Dynamics Simulations and Structural Analysis to investigate its Potential Functional Role. Proteins 2017, 85, 1454–1467. [Google Scholar] [CrossRef]
- Khadka, B.; Adeolu, M.; Blankenship, R.E.; Gupta, R.S. Novel insights into the origin and diversification of photosynthesis based on analyses of conserved indels in the core reaction center proteins. Photosynth. Res. 2016, 131, 159–171. [Google Scholar] [CrossRef]
- Gupta, R.S. Moelcular Markers for Photosynthetic Bacteria and Insights into the Origin and Spread of Photosynthesis. Adv. Bot. Res. 2013, 66, 37–66. [Google Scholar]
- Nadson, G.A. The morphology of inferior Algae. III. Chlorobium limicola Nads., the green chlorophyll bearing microbe. Bull. Du Jard. Bot. St. Petersbourg 1906, 6, 190. [Google Scholar]
- Gibson, J.; Pfenning, N.; Waterbury, J.B. In Validation list no. 17. Validation of publication of new names and new combinations previously effectively published outside the IJSB. Int. J. Syst. Bacteriol. 1985, 35, 223–225. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Accession/GI Number | Figure No. | Indel Size | Indel Position |
|---|---|---|---|---|
| Phosphoribosylformylglycinamidine synthase II | WP_041465690 | Figure 4 | 5 aa ins | 270–320 |
| Glycyl-tRNA synthetase | 78185909 | Figure S4 | 1 aa ins | 248–314 |
| RNA polymerase sigma-32 subunit | 194337394 | Figure S5 | 12 aa ins | 24–79 |
| Uroporphyrinogen decarboxylase | 189347814 | Figure S6 | 3 aa ins | 284–333 |
| Glutamyl-tRNA reductase | 194336061 | Figure S7 | 1 aa ins | 211–250 |
| Glutamate-1-semialdehyde aminotransferase | 21674908 | Figure S8 | 1 aa ins | 60–109 |
| fructose-1,6-bisphosphate aldolase, class II | 189500266 | Figure S9 | 11 aa del | 233–282 |
| A major facilitator superfamily protein | 193212482 | Figure S10 | 1 aa ins | 325–365 |
| DNA polymerase III subunit alpha | 78187124 | Figure S11 | 2–4 aa ins | 294–340 |
| Acetyl-CoA carboxylase carboxyltransferase subunit alpha | 119356086 | Figure S12 | 6 aa ins | 103–137 |
| N-acetyl-alpha-D-glucosaminyl L-malate synthase BshA | 193214252 | Figure S13 | 3 aa ins | 88–132 |
| Serine hydroxymethyltransferase | 193215659 | Figure S14 | 2 aa ins | 228–280 |
| Deoxyribonucleotide triphosphate pyrophosphatase | 193215546 | Figure S15 | 3 aa ins | 94–144 |
| Polynucleotide phosphorylase/polyadenylase | 189346136 | Figure S16 | 1 aa ins | 37–69 |
| Polynucleotide phosphorylase/polyadenylase | 189346136 | Figure S17 | 17 aa ins | 264–335 |
| 7-carboxy-7-deazaguanine synthase QueE | 500067752 | Figure S18 | 3–7 aa ins | 119–164 |
| DNA polymerase III subunits gamma and tau | 193213861 | Figure S19 | 2 aa ins | 47–93 |
| DNA polymerase III subunits gamma and tau | 193213861 | Figure S20 | 4 aa ins | 220–268 |
| Cytidylate kinase | 21673125 | Figure S21 | 4 aa ins | 146–199 |
| 4-hydroxy-3-methylbut-2-enyl diphosphate reductase | 194333263 | Figure S22 | 2–5 aa ins | 264–311 |
| molecular chaperone DnaK | 193214979 | Figure S23 | 1 aa del | 70–119 |
| Tryptophanyl-tRNA synthetase | 78188056 | Figure S24 | 8 aa ins | 16–59 |
| Tryptophanyl-tRNA synthase | 493409794 | Figure S25 | 5 aa ins | 153–204 |
| Succinyl-CoA synthetase subunit alpha | 193216377 | Figure S26 | 7 aa ins | 31–87 |
| A Gfo/Idh/MocA family oxidoreductase | 21673819 | Figure S27 | 4 aa ins | 2–48 |
| A Gfo/Idh/MocA family oxidoreductase | 21673819 | Figure S28 | 2 aa ins | 209–249 |
| Ribonuclease R | 193213033 | Figure S29 | 7–8 aa ins | 176–223 |
| Ribonuclease R | 193213033 | Figure S30 | 4–5 aa ins | 558–608 |
| Ribonuclease R | 193213033 | Figure S31 | 4 aa ins | 640–720 |
| Bifunctional 5,10-methylene-THF dehydrogenase | 78186789 | Figure S32 | 2 aa ins | 128–161 |
| Lactoylglutathione lyase | 78187149 | Figure S33 | 2 aa ins | 5–46 |
| Thiazole synthase | 78187419 | Figure S34 | 1 aa ins | 55–106 |
| DNA mismatch repair protein * | 193213813 | Figure S35 | 1 aa del | 599–644 |
| Protein Name | Accession/GI Number | Figure No. | Indel Size | Indel Position | Specificity |
|---|---|---|---|---|---|
| Polyphosphate kinase | WP_012466203 | Figure 5 | 1 aa ins | 25–65 | Family Chlorobiaceae |
| Peptide chain release factor 3 | 193212508 | Figure S36 | 1 aa del | 400–447 | |
| Peptidase U32 | 493410285 | Figure S37 | 6 aa ins | 260–316 | |
| MiaB-like tRNA modifying protein | 145220314 | Figure S38 | 5 aa ins | 329–382 | |
| Molecular chaperone HtpG | 145219831 | Figure S39 | 3 aa ins | 259–314 | |
| DegT/DnrJ/EryC1/StrS aminotransferase | 193212751 | Figure S40 | 1 aa del | 111–156 | |
| biogenesis of lysosome-related organelles complex 1 subunit 2 | 194336576 | Figure S41 | 1 aa del | 238–279 | |
| DNA gyrase subunit A | 194335275 | Figure S42 | 1 aa ins | 234–268 | |
| UDP-glucose/GDP-mannose dehydrogenase family protein | WP_012498759 | Figure 6 | 2 aa ins | 287–330 | Family Chloroherpetonaceae |
| hypoxanthine phosphoribosyltransferase | PIO48526 | Figure S43 | 1 aa ins | 57–94 | |
| dihydrolipoyl dehydrogenase | PIO48610 | Figure S44 | 1 aa ins | 30–64 | |
| SDR family oxidoreductase | WP_012499003 | Figure S45 | 4 aa ins | 236–280 | |
| RecQ family ATP-dependent DNA helicase | WP_012499407 | Figure S46 | 2 aa ins | 164- 203 | |
| alkaline phosphatase family protein | RFM24133 | Figure S47 | 5 aa ins | 33–78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bello, S.; Howard-Azzeh, M.; Schellhorn, H.E.; Gupta, R.S. Phylogenomic Analyses and Molecular Signatures Elucidating the Evolutionary Relationships amongst the Chlorobia and Ignavibacteria Species: Robust Demarcation of Two Family-Level Clades within the Order Chlorobiales and Proposal for the Family Chloroherpetonaceae fam. nov. Microorganisms 2022, 10, 1312. https://doi.org/10.3390/microorganisms10071312
Bello S, Howard-Azzeh M, Schellhorn HE, Gupta RS. Phylogenomic Analyses and Molecular Signatures Elucidating the Evolutionary Relationships amongst the Chlorobia and Ignavibacteria Species: Robust Demarcation of Two Family-Level Clades within the Order Chlorobiales and Proposal for the Family Chloroherpetonaceae fam. nov. Microorganisms. 2022; 10(7):1312. https://doi.org/10.3390/microorganisms10071312
Chicago/Turabian StyleBello, Sarah, Mohammad Howard-Azzeh, Herb E. Schellhorn, and Radhey S. Gupta. 2022. "Phylogenomic Analyses and Molecular Signatures Elucidating the Evolutionary Relationships amongst the Chlorobia and Ignavibacteria Species: Robust Demarcation of Two Family-Level Clades within the Order Chlorobiales and Proposal for the Family Chloroherpetonaceae fam. nov" Microorganisms 10, no. 7: 1312. https://doi.org/10.3390/microorganisms10071312