
Impact of Elevated Levels of Dissolved CO2 on Performance and Proteome Response of an Industrial 2′-Fucosyllactose Producing Escherichia coli Strain


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain
2.2. Precultures
2.3. Fed-Batch Bioreactor Cultivations
2.4. CO2 Enrichment
2.5. Calibration of CO2 Probe
2.6. Sampling and Analytical Procedures
2.7. Proteomics Analysis
2.8. Data Analysis
3. Results
3.1. pCO2 Levels in Industrial and Laboratory Scale
3.2. Dual Limitation in the Fermentation Process
3.3. Fermentation Performance with and without pCO2 Enrichment
3.4. Proteome Analysis
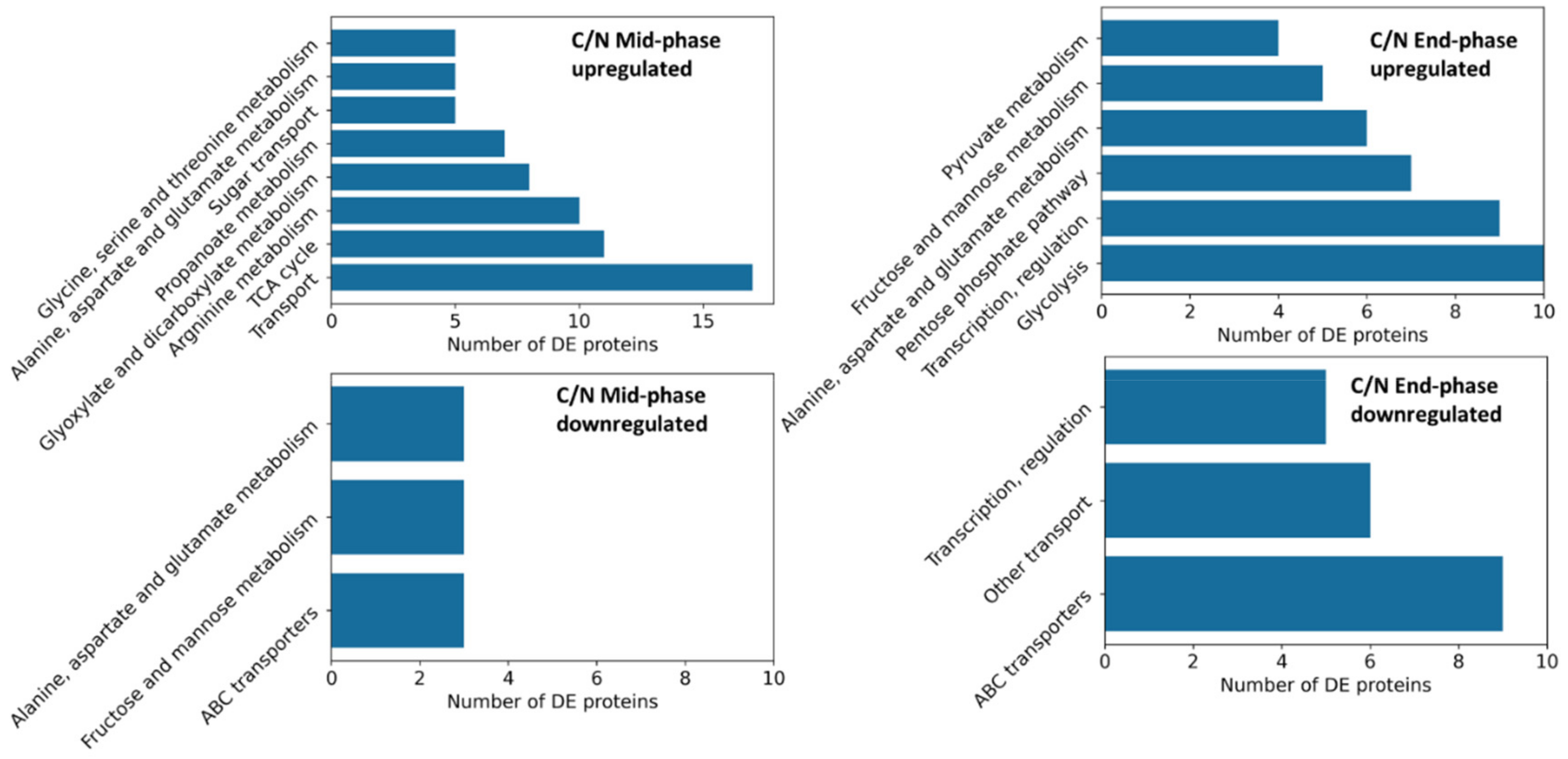
3.4.1. Identification of Differentially Expressed Proteins
3.4.2. Time Course Expression Changes of 2′-FL Production and TCA Related Proteins
3.5. Proteins from the 2′-FL Production Pathways Were Not Significantly Affected by CO2 Enrichment
3.6. CO2 Enrichment Increased TCA Cycle Protein Expression
3.7. Enzymes Involved in Carboxylation and Decarboxylation Reactions
3.8. Nitrogen Uptake Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CO2 Enrichment | Nutrient Limitation | Number of Replicate Fermentations | Number of Fermentations Sampled for Proteomics | Timepoints for Proteomics Sampling |
|---|---|---|---|---|
| Control (0%) | C/N | 4 | 3 | 6 h |
| 30 h | ||||
| 120 h | ||||
| Control (0%) | C | 3 | 2 | 6 h |
| 30 h | ||||
| 80 h | ||||
| 120 h | ||||
| 15% enriched | C | 2 | 2 | 6 h |
| 30 h | ||||
| 80 h | ||||
| 120 h | ||||
| 10% enriched | C/N | 1 | 1 | 30 h |
| 80 h | ||||
| 120 h | ||||
| 15% enriched | C/N | 3 | 2 | 6 h |
| 30 h | ||||
| 80 h | ||||
| 120 h | ||||
| 20% enriched | C/N | 2 | 1 | 6 h |
| 30 h | ||||
| 80 h | ||||
| 100 h | ||||
| 30% enriched | C/N | 2 | n/a | n/a |
Appendix B
| TCA Cycle/Pyruvate Metabolism | 2′-FL Production | |||
|---|---|---|---|---|
| Protein | Function | Protein | Function | |
| AceB | malate synthase A | Gmd | GDP-mannose 4,6-dehydratase | |
| AceK | isocitrate dehydrogenase kinase/isocitrate dehydrogenase phosphatase | LacY | lactose permease | |
| AcnA | aconitate hydratase A | ManA | mannose-6-phosphate isomerase | |
| AcnB | bifunctional aconitate hydratase B and 2-methylisocitrate dehydratase | ManB | phosphomannomutase | |
| FumA | fumarase A | ManC | mannose-1-phosphate guanyltransferase | |
| FumC | Fumarate hydratase | PtsG | glucose-specific PTS enzyme IIBC component | |
| GltA | citrate synthase | FutC | fucosytransferase | |
| Icd | Isocitrate dehydrogenase | WcaG/Fcl | GDP-L-fucose synthase | |
| Lpd | lipoamide dehydrogenase | Decarboxylation/Carboxylation | ||
| MaeA | NAD+-dependent malate dehydrogenase | Protein | Function | |
| Mdh | malate dehydrogenase | Ppc | Phosphoenolpyruvate carboxylase | |
| Ppc | Phosphoenolpyruvate carboxylase | AceF | Acetyltransferase, component of pyruvate dehydrogenase complex | |
| SdhA | succinate:quinone oxidoreductase, FAD binding protein | PyrF | Orotidine 5′-phosphate decarboxylase | |
| SdhB | succinate:quinone oxidoreductase, iron-sulfur cluster binding protein | NadC | Nicotinate-nucleotide pyrophosprolyase, decarboxylating | |
| SdhC | succinate:quinone oxidoreductase, membrane protein | HemE | Uroporghyrinogen decarboxylase | |
| SdhD | succinate:quinone oxidoreductase, membrane protein | Psd | Phosphatidylserine decarboxylase proenzyme | |
| SucA | 2-oxoglutarate decarboxylase, thiamine-requiring | SucB | succinyltransferase component of 2-oxoglutarate dehydrogenase | |
| SucB | dihydrolipoyltranssuccinylase | SucA | 2-oxoglutarate decarboxylase | |
| SucD | succinyl-CoA synthetase subunit α | LpdA | lipoamide dehydrogenase | |
| Icd | isocitrate dehydrogenase | |||
| PoxB | Pyruvate oxidase | |||
| MaeA | malate dehydrogenase, oxaloacetate-decarboxylating and NAD+ dependent | |||
| MaeB | malate dehydrogenase, oxaloacetate-decarboxylating and NADP+ dependent | |||
Appendix C

Appendix D

Appendix E

References
- Sakanaka, M.; Hansen, M.E.; Gotoh, A.; Katoh, T.; Yoshida, K.; Odamaki, T.; Yachi, H.; Sugiyama, Y.; Kurihara, S.; Hirose, J.; et al. Evolutionary adaptation in fucosyllactose uptake systems supports bifidobacteria-infant symbiosis. Sci. Adv. 2019, 5, eaaw7696. [Google Scholar] [CrossRef] [Green Version]
- Brand-Miller, J.C.; McVeagh, P.; McNeil, Y.; Messer, M. Digestion of human milk oligosaccharides by healthy infants evaluated by the lactulose hydrogen breath test. J. Pediatr. 1998, 133, 95–98. [Google Scholar] [CrossRef]
- Hester, S.N.; Hustead, D.S.; Mackey, A.D.; Singhal, A.; Marriage, B.J. Is the Macronutrient Intake of Formula-Fed Infants Greater Than Breast-Fed Infants in Early Infancy? J. Nutr. Metab. 2012, 2012, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soyyılmaz, B.; Mikš, M.H.; Röhrig, C.H.; Matwiejuk, M.; Meszaros-Matwiejuk, A.; Vigsnæs, L.K. The Mean of Milk: A Review of Human Milk Oligosaccharide Concentrations throughout Lactation. Nutrients 2021, 13, 2737. [Google Scholar] [CrossRef]
- Ammann, R. Achieving the impossible: Jennewein Biotechnologie is dedicated to the production of human milk oligosaccharides. Eur. Dairy Mag. 2017, 5, 30–31. [Google Scholar]
- Bych, K.; Mikš, M.H.; Johanson, T.; Hederos, M.J.; Vigsnæs, L.K.; Becker, P. Production of HMOs using microbial hosts—From cell engineering to large scale production. Curr. Opin. Biotechnol. 2018, 56, 130–137. [Google Scholar] [CrossRef]
- Larsson, G.; Törnkvist, E.; Ståhl Wernersson, C.; Trägårdh, H.; Noorman, H.; Enfors, S.-O. Substrate gradients in bioreactors: Origin and consequences. Bioprocess Biosyst. Eng. 1996, 14, 281–289. [Google Scholar] [CrossRef]
- Sandoval-Basurto, E.A.; Gosset, G.; Bolívar, F.; Ramírez, O.T. Culture of Escherichia coliunder dissolved oxygen gradients simulated in a two-compartment scale-down system: Metabolic response and production of recombinant protein. Biotechnol. Bioeng. 2004, 89, 453–463. [Google Scholar] [CrossRef]
- Lara, A.R.; Leal, L.; Flores, N.; Gosset, G.; Bolívar, F.; Ramírez, O.T. Transcriptional and metabolic response of recombinant Escherichia coli to spatial dissolved oxygen tension gradients simulated in a scale-down system. Biotechnol. Bioeng. 2005, 93, 372–385. [Google Scholar] [CrossRef]
- Cortés, J.T.; Flores, N.; Bolívar, F.; Lara, A.R.; Ramírez, O.T. Physiological effects of pH gradients on Escherichia coliduring plasmid DNA production. Biotechnol. Bioeng. 2015, 113, 598–611. [Google Scholar] [CrossRef]
- Crater, J.S.; Lievense, J.C. Scale-up of industrial microbial processes. FEMS Microbiol. Lett. 2018, 365, fny138. [Google Scholar] [CrossRef]
- Junker, B.H. Scale-up methodologies for Escherichia coli and yeast fermentation processes. J. Biosci. Bioeng. 2004, 97, 347–364. [Google Scholar] [CrossRef]
- McIntyre, M.; McNeil, B. Effects of elevated dissolved CO2 levels on batch and continuous cultures of Aspergillus niger A60: An evaluation of experimental methods. Appl. Environ. Microbiol. 1997, 63, 4171–4177. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.M.; Goyal, A.; Rank, D.L.; Gupta, S.K.; Boom, T.V.; Lee, S.S. Effects of Elevated pCO2 and Osmolality on Growth of CHO Cells and Production of Antibody-Fusion Protein B1: A Case Study. Biotechnol. Prog. 2008, 21, 70–77. [Google Scholar] [CrossRef]
- Sander, R. Compilation of Henry’s law constants (version 4.0) for water as solvent. Atmospheric Chem. Phys. 2015, 15, 4399–4981. [Google Scholar] [CrossRef] [Green Version]
- Lara, A.R.; Taymaz-Nikerel, H.; Mashego, M.R.; van Gulik, W.M.; Heijnen, J.J.; Ramírez, O.T.; van Winden, W.A. Fast dynamic response of the fermentative metabolism of Escherichia colito aerobic and anaerobic glucose pulses. Biotechnol. Bioeng. 2009, 104, 1153–1161. [Google Scholar] [CrossRef]
- Simen, J.D.; Löffler, M.; Jäger, G.; Schäferhoff, K.; Freund, A.; Matthes, J.; Müller, J.; Takors, R.; Team, R.N. Transcriptional response of Escherichia colito ammonia and glucose fluctuations. Microb. Biotechnol. 2017, 10, 858–872. [Google Scholar] [CrossRef]
- Baez, A.; Flores, N.; Bolívar, F.; Ramírez, O.T. Simulation of dissolved CO2 gradients in a scale-down system: A metabolic and transcriptional study of recombinant Escherichia coli. Biotechnol. J. 2011, 6, 959–967. [Google Scholar] [CrossRef]
- Baez, A.; Flores, N.; Bolívar, F.; Ramírez, O.T. Metabolic and transcriptional response of recombinant Escherichia colito elevated dissolved carbon dioxide concentrations. Biotechnol. Bioeng. 2009, 104, 102–110. [Google Scholar] [CrossRef]
- Castan, A.; Näsman, A.; Enfors, S.-O. Oxygen enriched air supply in Escherichia coli processes: Production of biomass and recombinant human growth hormone. Enzym. Microb. Technol. 2002, 30, 847–854. [Google Scholar] [CrossRef]
- Knoll, A.; Bartsch, S.; Husemann, B.; Engel, P.; Schroer, K.; Ribeiro, B.; Stöckmann, C.; Seletzky, J.; Büchs, J. High cell density cultivation of recombinant yeasts and bacteria under non-pressurized and pressurized conditions in stirred tank bioreactors. J. Biotechnol. 2007, 132, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Zhang, F.; Hu, X.; Liao, X. Effects of high-pressure carbon dioxide on proteins and DNA in Escherichia coli. Microbiology 2011, 157, 709–720. [Google Scholar] [CrossRef] [Green Version]
- Ballestra, P.; DA Silva, A.A.; Cuq, J. Inactivation of Escherichia coli by Carbon Dioxide under Pressure. J. Food Sci. 1996, 61, 829–831. [Google Scholar] [CrossRef]
- Oulé, M.K.; Tano, K.; Bernier, A.-M.; Arul, J. Escherichia coli inactivation mechanism by pressurized CO2. Can. J. Microbiol. 2006, 52, 1208–1217. [Google Scholar] [CrossRef] [PubMed]
- Eklund, T. The effect of carbon dioxide on bacterial growth and on uptake processes in bacterial membrane vesicles. Int. J. Food Microbiol. 1984, 1, 179–185. [Google Scholar] [CrossRef]
- Zanghi, J.A.; Schmelzer, A.E.; Mendoza, T.P.; Knop, R.H.; Miller, W.M. Bicarbonate concentration and osmolality are key determinants in the inhibition of CHO cell polysialylation under elevated pCO2 or pH. Biotechnol. Bioeng. 1999, 65, 182–191. [Google Scholar] [CrossRef]
- Dixon, N.M.; Kell, D.B. The inhibition by CO2 of the growth and metabolism of micro-organisms. J. Appl. Bacteriol. 1989, 67, 109–136. [Google Scholar] [CrossRef]
- Amoabediny, G.; Büchs, J. Determination of CO2 sensitivity of micro-organisms in shaken bioreactors. I. Novel method based on the resistance of sterile closure. Biotechnol. Appl. Biochem. 2010, 57, 157–166. [Google Scholar] [CrossRef]
- Pedersen, M.; Papadakis, M. Nucleic Acid Construct for In Vitro and In Vivo Gene Expression. Denamrk Patent WO2019123324A1, 27 June 2019. [Google Scholar]
- Bruderer, R.; Bernhardt, O.M.; Gandhi, T.; Miladinović, S.M.; Cheng, L.-Y.; Messner, S.; Ehrenberger, T.; Zanotelli, V.R.T.; Butscheid, Y.; Escher, C.; et al. Extending the Limits of Quantitative Proteome Profiling with Data-Independent Acquisition and Application to Acetaminophen-Treated Three-Dimensional Liver Microtissues. Mol. Cell. Proteom. 2015, 14, 1400–1410. [Google Scholar] [CrossRef] [Green Version]
- Löffler, M.; Simen, J.D.; Müller, J.; Jäger, G.; Laghrami, S.; Schäferhoff, K.; Freund, A.; Takors, R. Switching between nitrogen and glucose limitation: Unraveling transcriptional dynamics in Escherichia coli. J. Biotechnol. 2017, 258, 2–12. [Google Scholar] [CrossRef]
- Bernal, V.; Cerezo, S.C.; Cánovas, M. Acetate metabolism regulation in Escherichia coli: Carbon overflow, pathogenicity, and beyond. Appl. Microbiol. Biotechnol. 2016, 100, 8985–9001. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Jahic, M.; Enfors, S.-O. Modeling of Overflow Metabolism in Batch and Fed-Batch Cultures of Escherichia coli. Biotechnol. Prog. 1999, 15, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Turner, C.; Gregory, M.E.; Turner, M.K. A study of the effect of specific growth rate and acetate on recombinant protein production of Escherichia coli JM107. Biotechnol. Lett. 1994, 16, 891–896. [Google Scholar] [CrossRef]
- Luli, G.W.; Strohl, W.R. Comparison of growth, acetate production, and acetate inhibition of Escherichia coli strains in batch and fed-batch fermentations. Appl. Environ. Microbiol. 1990, 56, 1004–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinhal, S.; Ropers, D.; Geiselmann, J.; de Jong, H. Acetate Metabolism and the Inhibition of Bacterial Growth by Acetate. J. Bacteriol. 2019, 201, e00147-19. [Google Scholar] [CrossRef] [Green Version]
- Trček, J.; Mira, N.P.; Jarboe, L.R. Adaptation and tolerance of bacteria against acetic acid. Appl. Microbiol. Biotechnol. 2015, 99, 6215–6229. [Google Scholar] [CrossRef] [PubMed]
- Chubukov, V.; Desmarais, J.J.; Wang, G.; Chan, L.J.G.; Baidoo, E.E.K.; Petzold, C.J.; Keasling, J.D.; Mukhopadhyay, A. Engineering glucose metabolism of Escherichia coli under nitrogen starvation. NPJ Syst. Biol. Appl. 2017, 3, 16035. [Google Scholar] [CrossRef]
- Morita, T.; Fukuoka, T.; Imura, T.; Kitamoto, D. Accumulation of cellobiose lipids under nitrogen-limiting conditions by two ustilaginomycetous yeasts, Pseudozyma aphidis and Pseudozyma hubeiensis. FEMS Yeast Res. 2013, 13, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Fukamachi, T.; Saito, H.; Kobayashi, H. Carbon dioxide increases acid resistance in Escherichia coli. Lett. Appl. Microbiol. 2005, 40, 397–400. [Google Scholar] [CrossRef]
- Aguilera, J.; Petit, T.; De Winde, J.H.; Pronk, J.T. Physiological and genome-wide transcriptional responses of Saccharomyces cerevisiae to high carbon dioxide concentrations. FEMS Yeast Res. 2005, 5, 579–593. [Google Scholar] [CrossRef] [Green Version]
- Kai, Y.; Matsumura, H.; Izui, K. Phosphoenolpyruvate carboxylase: Three-dimensional structure and molecular mechanisms. Arch. Biochem. Biophys. 2003, 414, 170–179. [Google Scholar] [CrossRef]
- Lukas, H.; Reimann, J.; Bin Kim, O.; Grimpo, J.; Unden, G. Regulation of Aerobic and Anaerobic d -Malate Metabolism of Escherichia coli by the LysR-Type Regulator DmlR (YeaT). J. Bacteriol. 2010, 192, 2503–2511. [Google Scholar] [CrossRef] [Green Version]










| CO2 Enrichment | Limitation | Number of Replicates | Relative Accumulated Biomass Yield (Yxs, %) * | Relative Accumulated Product Yield (Yps, %) * |
|---|---|---|---|---|
| 0 | C/N | 3 | 100 ± 6.7 | 100 ± 8.3 |
| 10% | C/N | 1 | 105 | 104 |
| 15% | C/N | 3 | 106 ± 7 | 96 ± 2.0 |
| 20% | C/N | 2 | 128–141 | 88–90 |
| 30% | C/N | 2 | n/a ** | n/a ** |
| 0% | C | 3 | 118 ± 5.1 | 85 ± 9.5 |
| 15% | C | 2 | 115–116 | 89–92 |
| Large-scale | C/N | 3 | 109 ± 4.7 | 77± 2.7 |
| C-Limited | C/N-Limited | ||||
|---|---|---|---|---|---|
| Comparison | Fermentation Phase | Number of DE Proteins | Comparison | Fermentation Phase | Number of DE Proteins |
| Control vs. 15% enriched | Early | 22 (9↑,13↓) | Control vs. 15% enriched | Early | N/A |
| Mid | 17 (9↑,8↓) | Mid | 139 (98↑,41↓) | ||
| End | 5 (3↑,2↓) | End | 218 (163↑,55↓) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gecse, G.; Vente, A.; Kilstrup, M.; Becker, P.; Johanson, T. Impact of Elevated Levels of Dissolved CO2 on Performance and Proteome Response of an Industrial 2′-Fucosyllactose Producing Escherichia coli Strain. Microorganisms 2022, 10, 1145. https://doi.org/10.3390/microorganisms10061145
Gecse G, Vente A, Kilstrup M, Becker P, Johanson T. Impact of Elevated Levels of Dissolved CO2 on Performance and Proteome Response of an Industrial 2′-Fucosyllactose Producing Escherichia coli Strain. Microorganisms. 2022; 10(6):1145. https://doi.org/10.3390/microorganisms10061145
Chicago/Turabian StyleGecse, Greta, André Vente, Mogens Kilstrup, Peter Becker, and Ted Johanson. 2022. "Impact of Elevated Levels of Dissolved CO2 on Performance and Proteome Response of an Industrial 2′-Fucosyllactose Producing Escherichia coli Strain" Microorganisms 10, no. 6: 1145. https://doi.org/10.3390/microorganisms10061145