1. Introduction
The gut microbiota have the capacity to interact with human cells, including specific immune cells. These interactions yield different health benefits in the host, including regulation of GIT motility, destroying toxins and mutagens, transforming bile acid and steroids, producing vitamins, absorbing minerals, and modulating mucosal and systemic immunity. Lactic acid bacteria, possessing potential probiotic characteristics and isolated from fermented food products, play a crucial role in improving the quality of the gut microbiota [
1,
2]. Probiotics are defined as “live microorganisms which, when administered in adequate amounts, confer a health benefit on the host” [
3]. Based on this definition, a microorganism is labeled as probiotic only when there is scientific evidence proving its potential health benefits to the host [
4]. The International Scientific Association for Probiotics and Prebiotics (ISAPP) states that for a microorganism to be described as probiotic, it should first go through a series of human or intended user trials to ensure safety and to prove at least one of the health benefits that the microorganism is claimed to provide for the host [
3]. In general, the most common microorganisms added to food products or supplements for their probiotic abilities include members of the lactic acid bacteria (LAB) and bifidobacteria [
1]. The nonviable (killed/dead) cells of probiotics have also been reported to exhibit health benefits [
5,
6]. This encouraged the ISAPP to issue a consensus on the definition of postbiotics (nonviable cells) [
7]. Thus, a postbiotic is defined as a “preparation of inanimate microorganisms and/or their components that confers a health benefit on the host” [
7].
The capacity of bacteria to survive through the GI system, to reach either the small or large intestine in sufficient numbers, and to interact with and/or attach to and colonize the host must be confirmed before probiotics can be added to foods and supplements for their possible health advantages [
1]. Several factors have a deleterious effect on probiotics, including the stomach’s high acidity (pH 1.5–3.0), bile salts, and digestive enzymes. Additionally, prior to consumption, the probiotics must maintain viability throughout culture manufacture and storage, product or supplement manufacture, and product shelf-life [
1].
Clinical studies have demonstrated various health effects associated with consumption of these microorganisms (live and killed), such as reduction in duration and occurrences of diarrhea, alleviation of symptoms of lactose intolerance, reduced incidences of pathogenic infection, and stimulation of the immune system and regulation of the inflammatory response [
8,
9]. The present study aimed to investigate the potential probiotic characteristics, i.e., tolerance to acid and bile, cholesterol removing ability, bile salt hydrolysis, resistance against lysozyme and antibiotics, production of exopolysaccharides (EPS), antimicrobial and hemolytic activities, and cell surface characteristics (auto-aggregation, co-aggregation and hydrophobicity), of some selected LAB isolated from vegetable products [
1].
Fruits and vegetables are one of the main dietary requirements in the adult diet and, according to the Dietary Guidelines for Americans, it is recommended to consume half a plate of fruits or vegetables in all meals [
10]. In terms of nutritional composition, fruits and vegetables are considered highly nutritious foods as they provide high amounts of vitamins, such as vitamin C and A; minerals, specifically electrolytes; and phytochemicals, specific antioxidants that fight free radicals [
10]. Thus, the current study aimed to isolate LAB from fresh vegetables and characterize these isolates as potential probiotics according to different properties, including (1) gastrointestinal tolerance to (a) in vitro digestion, (b) bile salts, and (c) lysozyme; (2) physiological properties such as (a) auto-aggregation, (b) co-aggregation, (c) hydrophobicity, (d) adhesion to HT-29 cells, and (e) cholesterol reduction; (3) production of desirable substances such as (a) bile salt hydrolase, (b) antimicrobials, and (c) EPS; and (4) bioactivities such as immunomodulation and sensitivity to antibiotics.
2. Materials and Methods
2.1. Sample Collection
Samples (140) of fresh fruits and vegetables (various types, namely tomato, cucumber, strawberry, peach, lettuce, parsley, and cabbage) were collected from local markets (Sharjah, United Arab Emirates) and transported in an icebox to the food microbiology lab of the University of Sharjah, Sharjah, United Arab Emirates, where isolation was carried out on De Man, Rogosa, and Sharpe (MRS) agar. Characterization of the LAB isolates as potential probiotics was carried out in the food microbiology lab of United Arab Emirates University (UAEU). Unless otherwise mentioned, the isolation and characterization were completed using Sigma-Aldrich chemicals (St. Louis, MO, USA).
2.2. Isolation of Lactic Acid Bacteria
The pour-plate technique was performed using MRS agar, and the plates were incubated anaerobically at 37 °C for 24 h in a CO2 incubator (Binder C 170, Tuttlingen, Germany). The Gram-positive and catalase-negative isolates were subcultured in MRS broth, and then the working stocks were prepared using 50 mL:50 mL glycerol:water. The stocks were stored at −80 °C. Overnight activation at 37 °C was carried out to investigate the potential probiotic characteristics of the isolates.
2.3. Tolerance to Stimulated Digestion Conditions Using INFOGEST2.0
Isolates grown overnight were centrifuged, and then the pellets were resuspended in 1 mL of 0.1 mM sodium phosphate buffer (pH 7.0). The in vitro digestion was done as per Brodkorb [
8]. Briefly, 1 mL of culture was mixed with 1 mL of oral stimulated salivary fluids (amylase 75 U/mL, salivary fluid SSF pH 7.0, 0.3 M CaCl
2, total volume 2 mL) for 2 min. Afterward, the 2 mL of oral bolus was mixed with 2 mL simulated gastric fluids (pepsin 2000 U/mL, gastric lipase using rabbit gastric extract RGE 60 U/mL (Lipolytech, Marseille, France), stimulated gastric fluids (SGF) pH 3.0, 0.3 M CaCl
2, total volume 4 mL) for 120 min at 37 °C. Finally, the 4 mL of gastric chyme was mixed with 4 mL stimulated intestinal fluids (pancreatin 100 U/mL, bile 10 mmol/L, stimulated intestinal fluids (SIF) pH 7.0, 0.3 M CaCl
2, total volume 24 mL) for 120 min at 37 °C. After the digestion steps were completed, a 1 mL sample of the digest was taken aseptically and serial dilutions were made. The bacterial enumeration was carried out on MRS agar using the pour-plate technique. After anaerobic incubation at 37 °C for 48 h, the plates were counted using a colony counter (Interscience San 1200; New York, NY, USA).
2.4. Bile Tolerance
The bile tolerance test was carried out according to Liong and Shah [
6] by adding cholic acid (0.30%), taurocholic acid (1.0%), and oxgall (1.0%) to the overnight-activated isolates in MRS broth at 37 °C. The absorbance was then measured at 620 nm after 0, 3, and 6 h using a microplate spectrophotometer (Epoch-2, BioTek; Winooski, VT, USA). The activated cultures in MRS broth without bile salt were employed as positive controls to calculate the percentage of growth according to the following equation:
2.5. Bile Salt Hydrolase (BSH) Activity
BSH activity was examined as described by Ayyash [
11] by detecting the amino acids released from the conjugated bile salts (6 mM sodium glycochenodeoxycholic, taurocholic, taurochenodeoxycholic, and taurodeoxycholic acids) by measuring the absorbance of ninhydrin at 570 nm using an Epoch-2 Microplate Spectrophotometer (BioTek, Rancho Cordova, CA, USA).
2.6. Cholesterol Removal
The cholesterol removal ability of the isolates was determined as per Shivangi [
12]. The activated cultures were inoculated in MRS broth supplemented with 100 µg/mL water-soluble cholesterol (Sigma, St. Louis, MI, USA), followed by incubation at 37 °C for 24 h. The uninoculated tube was employed as a control. After incubation, the cells were removed by centrifuging at 6000×
g for 15 min. The spent broth was collected in clean, dry tubes. From the collected spent broth, 0.5 mL was placed in a test tube and 3 mL of 95% ethanol was added followed by 2 mL of 50% KOH. After mixing thoroughly, the contents were heated at 60 °C for 10 min in a water bath and subsequently cooled. To each tube, 5 mL of n-hexane was added and mixed, followed by a further 3 mL of distilled water which was also mixed. The tubes were allowed to stand for 15 min at room temperature to separate the phases. After phase separation, 2.5 mL of the upper hexane layer was separated and placed in a clean, dry test tube. The hexane was evaporated at 60 °C under the flow of nitrogen gas. To each tube, 4 mL of o-phthaldehyde reagent was added and allowed to stand at room temperature for 10 min. To each tube, 2 mL of concentrated sulfuric acid was added slowly to the side of tube, mixed, and allowed to stand for 10 min. The color developed was measured at 550 nm using an Epoch-2 Microplate Spectrophotometer (Agilent Technologies, Santa Clara, CA, USA).
2.7. Auto-Aggregation
Auto-aggregation of the activated cultures was determined according to Gao [
13], and the absorbance was measured at 0, 4, and 24 h at 600 nm. Auto-aggregation was estimated according to the following equation:
where A
t is absorbance at time t, and A
0 is absorbance at time 0.
2.8. Hydrophobicity
Hydrophobicity of the isolates to various hydrocarbons, namely xylene, hexadecane, and octane, was evaluated according to Abushelaibi [
14], and the final absorbance was measured at 600 nm using an Epoch-2 Microplate Spectrophotometer. The reduced absorbance in the aqueous phase was taken as measure of cell surface hydrophobicity. The hydrophobicity was calculated as per the following equation:
where A is the initial absorbance at 600 nm and A
0 is the final absorbance.
2.9. Adhesion to HT-29 Cells
To determine the adhesion ability, HT-29 cells were seeded on to 24-well tissue culture plates at a concentration of 10
5 cells/well and incubated at 37 °C for 24 h in a 5% CO
2 and 95% air atmosphere. The isolate cells from 20 h cultures were washed twice with Dulbecco’s phosphate-buffered saline (DPBS) and then resuspended in nonsupplemented DMEM at a concentration of 10
7–10
8 CFU/mL. These bacterial suspensions were added to the monolayers of the HT-29 cells. After 2 h of incubation at 37 °C in a 5% CO
2 and 95% air atmosphere, the wells were washed three times with PBS to remove the bacterial suspensions and nonadherent cells and treated with 1% Triton X-100 (Sigma–Aldrich, St. Louis, MO, USA) solution to detach the adherent bacteria. The final viable LAB cell counts (adherent LAB) were enumerated on MRS agar [
15]. Adhesion percentage was then estimated using the following equation:
where A
t is the number of the adhered cells (log CFU/mL) after incubation, and A
0 is the initial cell number (log CFU/mL).
2.10. Co-Aggregation
Co-aggregation was examined according to Abushelaibi [
14] using four pathogenic bacteria, namely
Escherichia coli 0157:H7 1934,
Staphylococcus aureus ATCC 25923,
Salmonella Typhimurium 02-8423, and
Listeria monocytogenes DSM 20649. The results were expressed as co-aggregation percentages utilizing the following equation:
2.11. Antimicrobial Production
Antimicrobial activity was determined using a cell-free supernatant as per Ayyash [
11]. Briefly, the MRS agar plates were overlaid with 7 mL of soft MRS agar inoculated with 20 µL of overnight-activated culture of indicator strains. Different wells were made in the agar. Wells were filled with 50 Al cell-free broth of 24 h old cultures obtained by centrifuging the culture broth at 5000 rpm for 15 min. The broth was neutralized to pH 6.5 and it was also inoculated into wells. The diameter of the zone of inhibition extending laterally around the well was measured and a clear zone of 1 mm or more was considered positive inhibition.
2.12. Lysozyme Activity
Evaluation of LAB isolates’ tolerance to lysozyme over 90 min of incubation at 37 °C was carried out as per Ayyash [
11]. The bacterial enumeration was carried out on MRS agar using the pour-plate technique after 90 min of incubation at 37 °C. After anaerobic incubation at 37 °C for 48 h, the plates were counted using a colony counter (Interscience San 1200; Park Woburn, MA, USA).
2.13. Antibiotic Susceptibility Test
Antibiotic resistance testing was performed according to Shivangi [
12] with slight modifications, as MRS and M17 agar plates were used for the respective isolates. The susceptibility of the isolates was tested against penicillin (PEN, 10 mg), clindamycin (CLI, 2 mg), vancomycin (VAN, 30 mg), and erythromycin (ERY, 15 mg). The interpretations of zones as resistant (R), moderately susceptible (MS), and susceptible (S) were assigned according to Charteris [
16].
2.14. Exopolysaccharide (EPS) Production
EPS production indication testing (−
ve/+
ve) was conducted as described by Abushelaibi [
14], using milk–ruthenium medium. Overnight cultures were streaked onto the surface of plates containing ruthenium red milk (10%
w/
v, skim milk powder, 1%
w/
v, sucrose, and 0.08 g/L ruthenium red, 1.5%
w/
v agar).
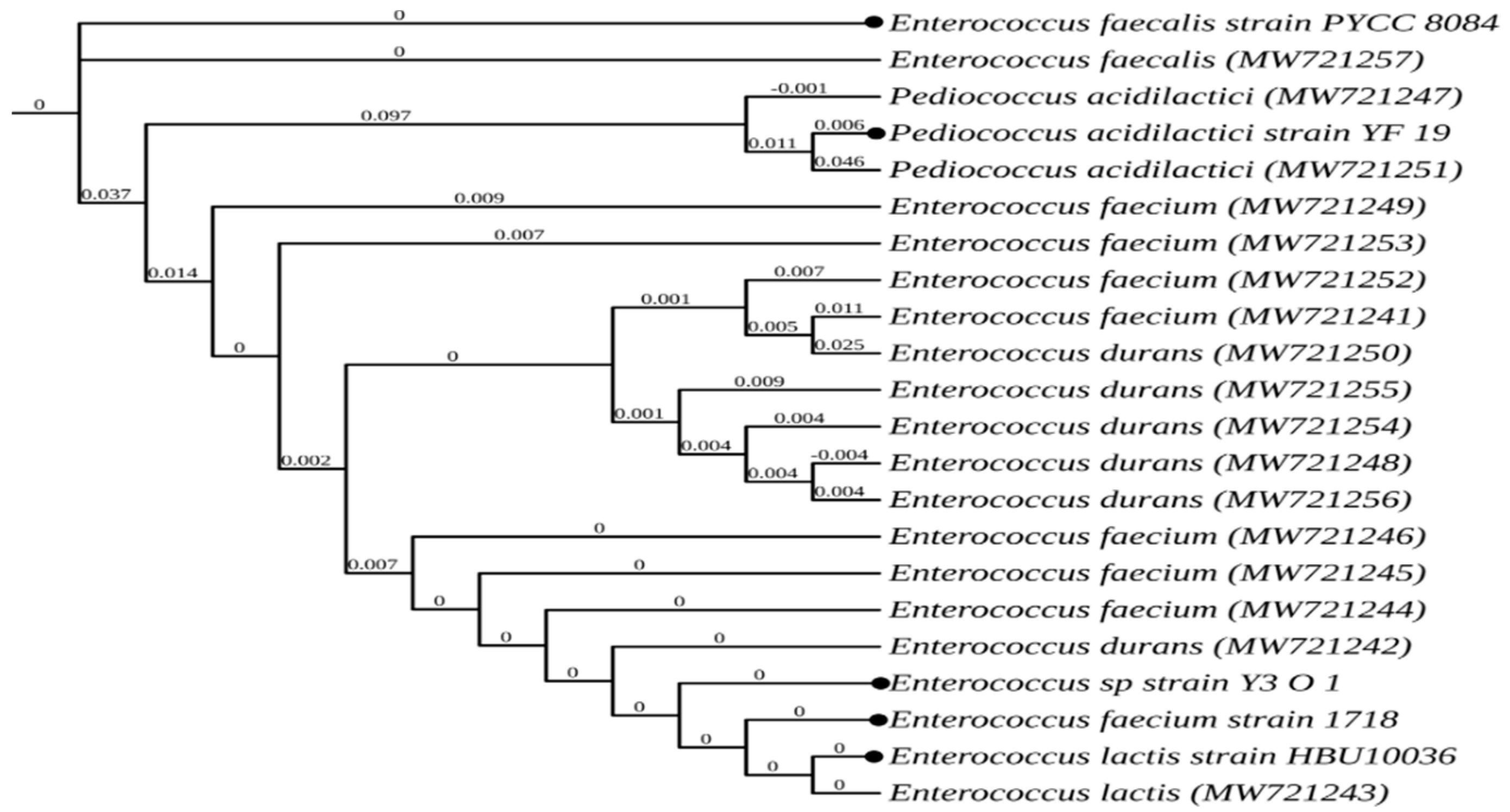
2.15. Identification of the LAB Isolates
The 16S rDNA of the selected isolates was amplified according to Alkalbani [
17], using PCR primers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-TACGGYTACCTTGTTACGACTT-3′), and 16S rDNA sequencing of the PCR product was done by Macrogen Sequencing Facilities (Macrogen, Seoul, Korea). The BLAST algorithm in the NCBI database was used to align the sequences and retrieve the accession number for each isolate from the GenBank. An online tool developed by Lemoine [
18] was used to determine the bacterial species most closely related to the isolates and to create the dendrogram.
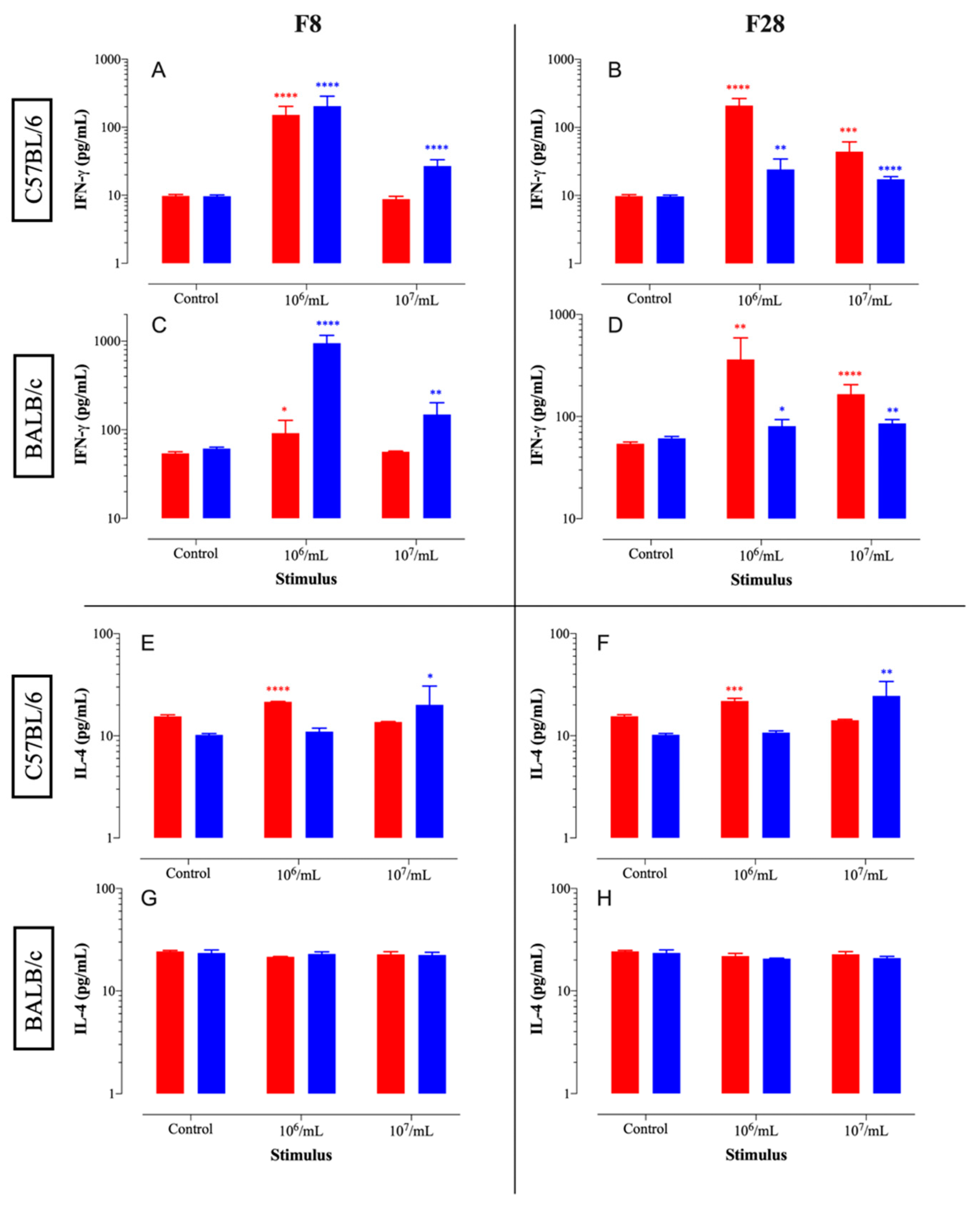
2.16. Immunomodulatory Effects
The immunomodulatory effects of the selected isolates were tested using spleen cells of BALB/c and C57BL/6 mice (Jackson Laboratory, Bar Harbor, ME, USA). The immunomodulatory activities of the selected bacterial isolates were tested by incubating them at different concentrations with aseptically prepared single-cell suspensions of murine spleen cells. Splenocytes of BALB/c and C57BL/6 mice were prepared for in vitro culture following a published protocol [
19]. Viable red blood cell depleted spleen cells were enumerated based on exclusion of trypan blue dye using a hemocytometer. Viable splenocytes (2 × 10
5/well) were co-cultured in 96-well plates with either 2 × 10
4 or 2 × 10
5 CFUs (equivalent to 10
6 or 10
7 CFUs/mL, respectively) of viable or killed LAB isolates (set up in triplicate per group) in RPMI medium supplemented with 10% FBS, penicillin/streptomycin, and gentamicin (Gibco-ThermoFisher Scientific, Waltham, MA, USA). Control wells were also set up in which spleen cells were cultured in medium alone. After 3 days of incubation at 37 °C, cell-free culture supernatants were harvested and kept at −20 °C until they were assayed for cytokines using specific ELISAs, as described previously [
20], utilizing commercially available kits for mouse IFN-γ (ThermoFisher Scientific, USA; cat#88-7314) and IL-4 (OptEIA kit; BD Biosciences, USA, ca# 555232). The sensitivity limits of the IFN-γ and IL-4 ELISAs were ~15 pg/mL and ~10 pg/mL, respectively.
2.17. Statistical Analysis
One-way ANOVA was applied to determine whether the differences between LAB isolates had a significant influence on the quantitative parameters (p < 0.05). Tukey’s test was used to detect differences between mean values with a p value of <0.05. To calculate the mean values and standard deviations, all tests were performed at least three times. Minitab version 21.0 (Minitab, Ltd., Coventry, UK) was used for all statistical analyses for noncell line studies. For the immunomodulatory effects, statistical significance between control and LAB isolate-stimulated cultures was analyzed using the unpaired two-tailed Student’s t-test. The statistical analyses were performed using GraphPad PRISM 8 software (GraphPad Software, San Diego, CA, USA), and differences with a p value ≤ 0.05 were considered significant.
4. Conclusions
To sum up, in vitro digestion was performed to screen for good probiotic characteristics involved in bile and lysozyme tolerance, cholesterol reduction activity, attachment property including hydrophobicity to hydrocarbons, auto-aggregation, co-aggregation, and attachment to HT-29 cells. Moreover, the selected isolates displayed antimicrobial and salt hydrolysis activities, and antibiotic susceptibility. Isolates F1, F5, F8, F15, F18, F21, F23, F25, F26, F28, F40, F43, and F46 displayed potential probiotic and postbiotic properties. However, F8 and F28 exhibited noticeable immunomodulatory activities. The immunomodulatory activities suggest a promising role of these isolates if used in animal studies and later human trials. Moreover, EPS was produced by all isolates except isolates F1, F5, F21, F23, F25, F26, F43, and F46 (Enterococcus faecium, E. durans, E. lactis, and Pediococcus acidilactici) which can be used to characterize isolates for industrial and medical purposes.


 ,
, 


{kind=link}
{kind=link}