
Xylella fastidiosa: A Glimpse of the Portuguese Situation
 , ,
, , 
Abstract
:1. Introduction
2. Distribution in Europe
3. Current Distribution in Portugal
4. Transmission
5. Hosts
6. Inoculation of the Bacteria by the Insect Vector
7. Bacterial Action in the Xylem
8. Movement and Distribution of the Bacteria in the Plant
9. Symptoms
10. Absence of Symptoms
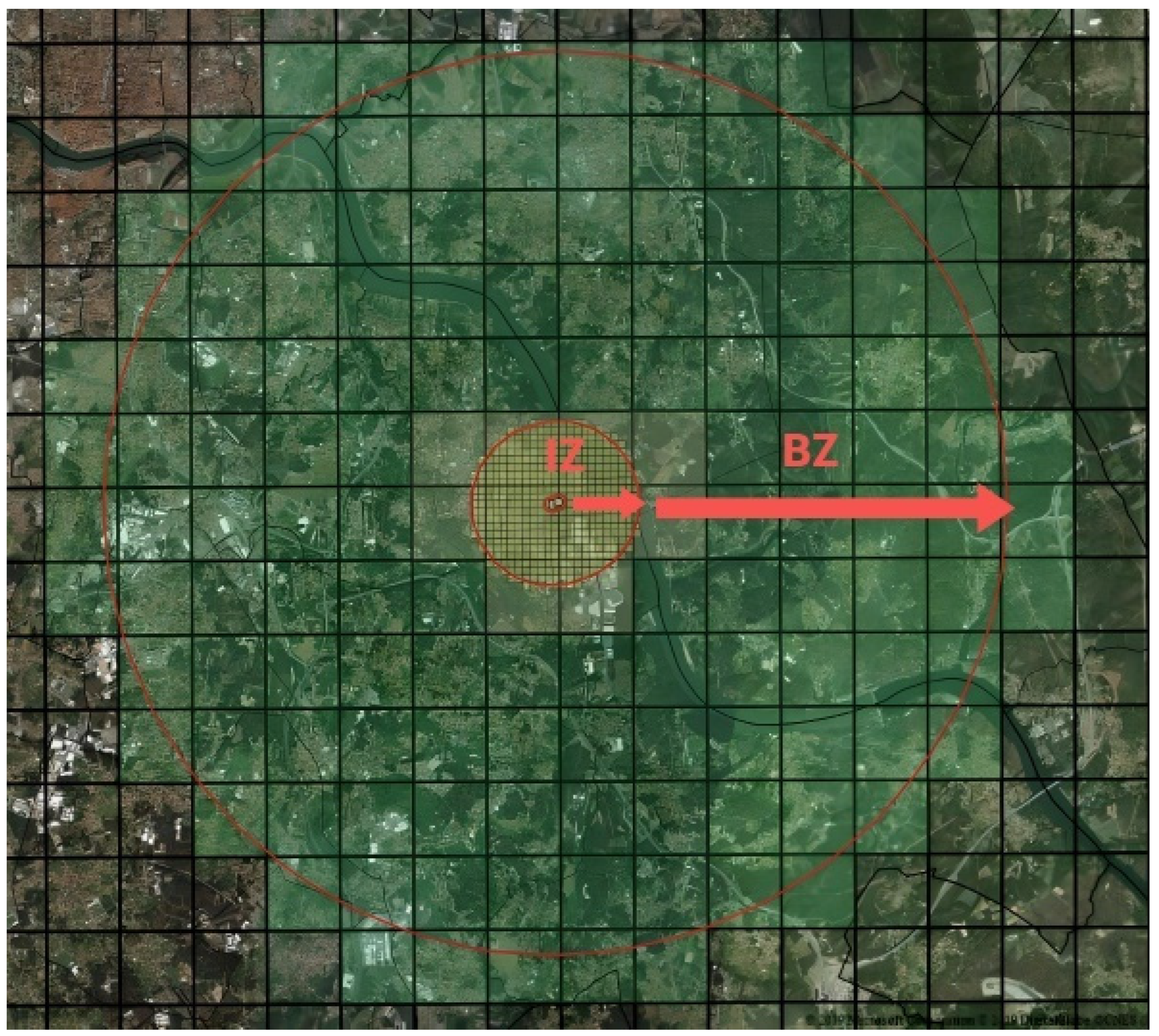
11. Disease Control Measures
- (a)
- The control of infected plants;
- (b)
- The use of tolerant cultivars;
- (c)
- The use of products that affect bacterial development;
- (d)
- The control of insect vectors.
11.1. Control of Infected Plants
11.2. Use of Tolerant Cultivars
11.3. Use of Products That Affect the Development of the Bacteria
11.4. Control of Insect Vectors
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wells, J.M.; Raju, B.C.; Hung, H.Y.; Weisburg, W.G.; Mandelco-Paul, L.; Brenner, D.J. Xylella fastidiosa gen. nov., sp. nov: Gram-Negative, xylem-limited, fastidious plant bacteria related to Xanthomonas spp. Int. J. Syst. Evol. Microbiol. 1987, 37, 136–143. [Google Scholar] [CrossRef]
- Montilon, V.; De Stradis, A.; Saponari, M.; Kubaa, R.A.; Giampetruzzi, A.; D’Attoma, G.; Saldareli, P. Xylella fastidiosa subsp. pauca ST53 exploits pit membranes of susceptible olive cultivars to spread systemically in the xylem. Plant Pathol. 2023, 72, 144–153. [Google Scholar] [CrossRef]
- Pereira, P.S. Xylella fastidiosa—A new menace for Portuguese agriculture and forestry. Rev. Ciências Agrárias 2015, 38, 149–154. [Google Scholar]
- Petit, G.; Bleve, G.; Gallo, A.; Mita, G.; Montanaro, G.; Nuzzo, V.; Zambonini, D.; Pitacco, A. Susceptibility to Xylella fastidiosa and functional xylem anatomy in Olea europaea: Revisiting a tale of plant-pathogen interaction. AoB Plants 2021, 13, plab027. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.; van der Werf, W.; Cendoya, M.; Lasink, A.O. Impact of Xylella fastidiosa subspecies pauca in European olives. Proc. Natl. Acad. Sci. USA 2020, 117, 9250–9259. [Google Scholar] [CrossRef] [PubMed]
- Trkulja, V.; Tomić, A.; Iličić, R.; Nožinić, M.; Milovanović, T.P. Xylella fastidiosa in Europe: From the introduction to the current status. Plant Pathol. J. 2022, 38, 551. [Google Scholar] [CrossRef] [PubMed]
- EPPO. PM 7/24 (4) Xylella fastidiosa. EPPO Bull. 2019, 49, 175–227. [Google Scholar] [CrossRef]
- Schaad, N.W.; Postnikova, E.; Lacy, G.; Fatmi, M.; Chang, C.J. Xylella fastidiosa subspecies: X. fastidiosa subsp. piercei, subsp. nov., X. fastidiosa subsp. multiplex subsp. nov., and X. fastidiosa subsp. pauca subsp. nov. Syst. Appl. Microbiol. 2004, 27, 290–300. [Google Scholar] [CrossRef]
- Schuenzel, E.L.; Scally, M.; Stouthamer, R.; Nunney, L. A multigene phylogenetic study of clonal diversity and divergence in North American strains of the plant pathogen Xylella fastidiosa. Appl. Environ. Microbiol. 2005, 71, 3832. [Google Scholar] [CrossRef]
- Nunney, L.; Schuenzel, E.L.; Scally, M.; Bromley, R.E.; Stouthamerc, R. Large-scale intersubspecific recombination in the plant-pathogenic bacterium Xylella fastidiosa is associated with the host shift to mulberry. Appl. Environ. Microbiol. 2014, 80, 3025–3033. [Google Scholar] [CrossRef]
- Randall, J.J.; Goldberg, N.P.; Kemp, J.D.; Radionenko, M.; French, J.M.; Olsen, M.W.; Hanson, S.F. Genetic analysis of a novel Xylella fastidiosa subspecies found in the Southwestern United States. Appl. Environ. Microbiol. 2009, 75, 5631–5638. [Google Scholar] [CrossRef] [PubMed]
- Janse, J.D.; Obradovic, A. Xylella fastidiosa: Its biology, diagnosis, control and risks on JSTOR. J. Plant Pathol. 2010, 92, 1.35–1.48. Available online: https://www.jstor.org/stable/41998754 (accessed on 15 April 2023).
- Sicard, A.; Zeilinger, A.R.; Vanhove, M.; Schartel, T.E.; Beal, D.J.; Daugherty, M.P.; Almeida, R.P.P. Xylella fastidiosa: Insights into an emerging plant pathogen. Annu. Rev. Phytopathol. 2018, 56, 181–202. [Google Scholar] [CrossRef] [PubMed]
- Baù, A.; Delbianco, A.; Stancanelli, G.; Tramontini, S. Susceptibility of Olea europaea L. varieties to Xylella fastidiosa subsp. pauca ST53: Systematic literature search up to 24 March 2017. EFSA J. 2017, 15, e04772. [Google Scholar] [CrossRef]
- Martelli, G.P.; Boscia, D.; Porcelli, F.; Saponari, M. The olive quick decline syndrome in south-east Italy: A threatening phytosanitary emergency. Eur. J. Plant Pathol. 2016, 144, 235–243. [Google Scholar] [CrossRef]
- Delbianco, A.; Gibin, D.; Pasinato, L.; Boscia, D.; Morelli, M. Update of the Xylella spp. host plant database—Systematic literature search up to 30 June 2022. EFSA J. 2023, 21, e07726. [Google Scholar] [CrossRef]
- Martinetti, D.; Soubeyrand, S. Identifying lookouts for epidemio-surveillance: Application to the emergence of Xylella fastidiosa in France. Phytopathology 2019, 109, 265–276. [Google Scholar] [CrossRef]
- Hernández, O.G.; García, L.V. Incidencia de Xylella fastidiosa en las Islas Baleares y distribución potencial en la península ibérica. Investig. Geográficas 2018, 69, 55–72. [Google Scholar] [CrossRef]
- Gilioli, G.; Simonetto, A.; Colturato, M.; Bazarra, N.; Férnandez, J.R.; Naso, M.G.; Donato, B.; Bosco, D.; Dongiovanni, C.; Maiorano, A.; et al. An eco-epidemiological model supporting rational disease management of Xylella fastidiosa. An application to the outbreak in Apulia (Italy). Ecol. Modell. 2023, 476, 110226. [Google Scholar] [CrossRef]
- Olmo, D.; Nieto, A.; Borràs, D.; Montesinos, M.; Adrover, F.; Pascual, A.; Gost, P.A.; Quetglas, B.; Urbano, A.; García, J.D.; et al. Landscape epidemiology of Xylella fastidiosa in the Balearic Islands. Agronomy 2021, 11, 473. [Google Scholar] [CrossRef]
- DGAV. Plano de Contingência Xylella fastidiosa e Seus Vetores; Direção-Geral da Alimentação e Veterinária: Lisbon, Portugal, 2022. [Google Scholar]
- DGAV. Xylella fastidiosa. 2023. Available online: https://www.dgav.pt/plantas/conteudo/sanidade-vegetal/inspecao-fitossanitaria/informacao-fitossanitaria/xylella-fastidiosa/ (accessed on 25 March 2023).
- Godefroid, M.; Morente, M.; Schartel, T.; Cornara, D.; Purcell, D.; Gallego, A.; Moreno, A.; Pereira, J.A.; Fereres, A. Climate tolerances of Philaenus spumarius should be considered in risk assessment of disease outbreaks related to Xylella fastidiosa. J. Pest Sci. 2004, 95, 855–868. [Google Scholar] [CrossRef]
- EFSA; Vos, S.; Camilleri, M.; Diakaki, M.; Lázaro, E.; Parnell, S.; Schenck, M.; Schrader, G.; Vincent, A. Pest survey card on Xylella fastidiosa. EFSA Support. Publ. 2019, 16, 1667E. [Google Scholar] [CrossRef]
- Di Serio, F.; Bodino, N.; Cavalieri, V.; Demichelis, S.; Di Carolo, M.; Dongiovanni, C.; Fumarola, G.; Gilioli, G.; Guerrieri, E.; Picciotti, U.; et al. Collection of data and information on biology and control of vectors of Xylella fastidiosa. EFSA J. 2019, 16, 1–102. [Google Scholar] [CrossRef]
- Purcell, A.H.; Saunders, S.R.; Hendson, M.; Grebus, M.E.; Henry, M.J. Causal role of Xylella fastidiosa in oleander leaf scorch disease. Phytopathology 2007, 89, 53–58. [Google Scholar] [CrossRef] [PubMed]
- EFSA; Lázaro, E.; Parnell, S.; Civera, A.V.; Schans, J.; Schenck, M.; Schrader, G.; Abrahantes, J.C.; Zancanaro, G.; Vos, S. Guidelines for statistically sound and risk-based surveys of Xylella fastidiosa. EFSA Support. Publ. 2020, 17, EN-1873. [Google Scholar] [CrossRef]
- Almeida, R.P.P. (Ed.) Chapter 12: Xylella fastidiosa vector transmission biology. In Vector-Mediated Transmission of Plant Pathogens; The American Phytopathological Society: Saint Paul, MN, USA, 2016; pp. 165–173. [Google Scholar]
- Chuche, J.; Sauvion, N.; Thiéry, D. Mixed xylem and phloem sap ingestion in sheath-feeders as normal dietary behavior: Evidence from the leafhopper Scaphoideus titanus. J. Insect Physiol. 2017, 102, 62–72. [Google Scholar] [CrossRef]
- Pompon, J.; Quiring, D.; Goyer, C.; Giordanengo, P.; Pelletier, Y. A phloem-sap feeder mixes phloem and xylem sap to regulate osmotic potential. J. Insect Physiol. 2011, 57, 1317–1322. [Google Scholar] [CrossRef]
- Cavalieri, V.; Altamura, G.; Fumarola, G.; di Carolo, M.; Saponari, M.; Cornara, D.; Bosco, D.; Dongiovanni, C. Transmission of Xylella fastidiosa subspecies pauca sequence type 53 by different insect species. Insects 2019, 10, 324. [Google Scholar] [CrossRef]
- Li, W.B.; Pria, W.D.; Lacava, P.M.; Qin, X.; Hartung, J.S. Presence of Xylella fastidiosa in sweet orange fruit and seeds and its transmission to seedlings. Phytopathology 2003, 93, 953–958. [Google Scholar] [CrossRef]
- Dalla, W.; Magalhães, P.L.; Li, W.; Miranda, V.S.; Costa, P.I.; Farias, P.R.S.; Hartung, V.S.; Pereira, E.O.; Francischini, F.J.B. Xylella fastidiosa em frutos e sementes de laranja-doce afetados pela clorose dos variegada dos citros. Laranja 2002, 23, 183–202. [Google Scholar]
- Cordeiro, A.B.; Sugahara, V.H.; Stein, B.; Leite Junior, R.P. Evaluation by PCR of Xylella fastidiosa subsp. pauca transmission through citrus seeds with special emphasis on lemons (Citrus limon (L.) Burm. f). Crop Prot. 2014, 62, 86–92. [Google Scholar] [CrossRef]
- Regulation (EU) 2016/2031 of the European Parliament and of the Council on protective measures against pests of plants, amending Regulations (EU) No. 228/2013, (EU) No. 652/2014 and (EU) No. 1143/2014 of the European Parliament and of the Council and repealing Council Directives 69/464/EEC, 74/647/EEC, 93/85/EEC, 98/57/EC, 2000/29/EC, 2006/91/EC and 2007/33/EC. Available online: https://eur-lex.europa.eu/legal-content/PT/TXT/PDF/?uri=CELEX:32020R1201&from=EN (accessed on 6 November 2022).
- Gibin, D.; Pasinato, L.; Delbianco, A. Update of the Xylella spp. host plant database—Systematic literature search up to 31 December 2022. EFSA J. 2023, 21, e08061. [Google Scholar] [CrossRef]
- DGAV. Plano de Ação pra Erradicação de Xylella fastidiosa e controlo dos seus vetores. Zona Demarcada da Área Metropolitana do Porto; Direção-Geral da Alimentação e Veterinária: Lisbon, Portugal, 2022. [Google Scholar]
- Carvalho, C.; Rodrigues, J.; Martins, L. Dispersion of the bacterium Xylella fastidiosa in Portugal. J. Agric. Sci. Technol. A 2022, 12, 35–41. [Google Scholar] [CrossRef]
- Selcuk, Y. On the meadow spittlebug Philaenus spumarius. Turk. J. Zool. 2000, 24, 447–459. [Google Scholar]
- Lago, C.; Cornara, D.; Minutillo, S.A.; Moreno, A.; Fereres, A. Feeding behaviour and mortality of Philaenus spumarius exposed to insecticides and their impact on Xylella fastidiosa transmission. Pest Manag. Sci. 2022, 78, 4841. [Google Scholar] [CrossRef]
- Dietrich, C.H. Keys to the families of Cicadomorpha and subfamilies and tribes of Cicadellidae (Hemiptera: Auchonorrhyncha). Fla. Enthomol. 2005, 88, 502–517. [Google Scholar] [CrossRef]
- Simionato, A.V.C.; Da Silva, D.S.; Lambais, M.R.; Carrilho, E. Characterization of a putative Xylella fastidiosa diffusible signal factor by HRGC-EI-MS. J. Mass Spectrom. 2007, 42, 490–496. [Google Scholar] [CrossRef]
- Killiny, N.; Almeida, R.P.P. Factors affecting the initial adhesion and retention of the plant pathogen Xylella fastidiosa in the foregut of an insect vector. Appl. Environ. Microbiol. 2014, 80, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Newman, K.L.; Almeida, R.P.P.; Purcell, A.H.; Lindow, S.E. Cell-cell signaling controls Xylella fastidiosa interactions with both insects and plants. Proc. Natl. Acad. Sci. USA 2004, 101, 1737–1742. [Google Scholar] [CrossRef] [PubMed]
- Killiny, N.; Prado, S.S.; Almeida, R.P.P. Chitin utilization by the insect-transmitted bacterium Xylella fastidiosa. Appl. Environ. Microbiol. 2010, 76, 6134–6140. [Google Scholar] [CrossRef] [PubMed]
- Redak, R.A.; Purcell, A.H.; Lopes, J.R.S.; Blua, M.J.; Mizell, R.F.; Andersen, P.C. The biology of xylem fluid-feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology. Annu. Rev. Entomol. 2004, 49, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Rapicavoli, J.N.; Blanco-Ulate, B.; Muszynski, A.; Figueroa-Balderas, R.; Morales-Cruz, A.; Azadi, P.; Doburchowska, J.M.; Castro, C.; Cantu, D.; Roper, M.C. Lipopolysaccharide O-antigen delays plant innate immune recognition of Xylella fastidiosa. Nat. Commun. 2018, 9, 390. [Google Scholar] [CrossRef] [PubMed]
- Surano, A.; Abou Kubaa, R.; Nigro, F.; Altamura, G.; Losciale, P.; Saponari, M.; Saldarelli, P. Susceptible and resistant olive cultivars show differential physiological response to Xylella fastidiosa infections. Front. Plant Sci. 2022, 13, 3603. [Google Scholar] [CrossRef] [PubMed]
- Pierce, B.K.; Kirkpatrick, B.C. The PhoP/Q two-component regulatory system is essential for Xylella fastidiosa survival in Vitis vinifera grapevines. Physiol. Mol. Plant Pathol. 2015, 89, 55–61. [Google Scholar] [CrossRef]
- Silva, F.R.; Vettore, A.L.; Kemper, E.L.; Leite, A.; Arruda, P. Fastidian gum: The Xylella fastidiosa exopolysaccharide possibly involved in bacterial pathogenicity. FEMS Microbiol. Lett. 2001, 203, 165–171. [Google Scholar] [CrossRef]
- Killiny, N.; Hernandez Martinez, R.; Korsi Dumenyo, C.; Cooksey, D.A.; Almeida, R.P.P. The exopolysaccharide of Xylella fastidiosa is essential for biofilm formation, plant virulence, and vector transmission. Mol. Plant-Microbe Interact. MPMI 2013, 26, 1044–1053. [Google Scholar] [CrossRef]
- Cursino, L.; Athinuwat, D.; Patel, K.R.; Galvani, C.D.; Zaini, P.A.; Li, Y.; De La Fuente, L.; Hoch, H.C.; Burr, T.J.; Mowery, P. Characterization of the Xylella fastidiosa PD1671 gene encoding eegenerate c-di-GMP GGDEF/EAL domains, and its role in the development of Pierce’s Disease. PLoS ONE 2015, 10, e0121851. [Google Scholar] [CrossRef]
- De Benedictis, M.; De Caroli, M.; Baccelli, I.; Marchi, G.; Bleve, G.; Gallo, A.; Ranaldi, F.; Falco, V.; Pasqualli, V.; Piro, G.; et al. Vessel occlusion in three cultivars of Olea europaea naturally exposed to Xylella fastidiosa in open field. J. Phytopathol. 2017, 165, 589–594. [Google Scholar] [CrossRef]
- Sun, Q.; Sun, Y.; Walker, M.A.; Labavitch, J.M. Vascular occlusions in grapevines with Pierce’s disease make disease symptom development worse. Plant Physiol. 2013, 161, 1529–1541. [Google Scholar] [CrossRef]
- Pérez-Donoso, A.G.; Sun, Q.; Caroline Roper, M.; Carl Greve, L.; Kirkpatrick, B.; Labavitch, J.M. Cell wall-degrading enzymes enlarge the pore size of intervessel pit membranes in healthy and Xylella fastidiosa-infected grapevines. Plant Physiol. 2010, 152, 1748–1759. [Google Scholar] [CrossRef]
- Chatterjee, S.; Wistrom, C.; Lindow, S.E. A cell-cell signaling sensor is required for virulence and insect transmission of Xylella fastidiosa. Proc. Natl. Acad. Sci. USA 2008, 105, 2670–2675. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, M.; Zaini, P.A.; Baccari, C.; Tran, S.; Da Silva, A.M.; Lindow, S.E. Xylella fastidiosa outer membrane vesicles modulate plant colonization by blocking attachment to surfaces. Proc. Natl. Acad. Sci. USA 2014, 111, E3910–E3918. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, M.; Luvisi, A.; Meyer, J.B.; Sabella, E.; De Bellis, L.; Cruz, A.C.; Ampatzidis, Y.; Cherubini, P. Specific fluorescence in situ hybridization (Fish) test to highlight colonization of xylem vessels by Xylella fastidiosa in naturally infected olive trees (Olea europaea L.). Front. Plant Sci. 2018, 9, 431. [Google Scholar] [CrossRef] [PubMed]
- He, C.X.; Li, W.B.; Ayres, A.J.; Hartung, J.S.; Miranda, V.S.; Teixeira, D.C. Distribution of Xylella fastidiosa in citrus rootstocks and transmission of citrus variegated chlorosis between sweet orange plants through natural root grafts. Plant Dis. 2007, 84, 622–626. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.P.P.; Pereira, E.F.; Purcell, A.H.; Lopes, J.R.S. Multiplication and movement of a citrus strain of Xylella fastidiosa within sweet orange. Plant Dis. 2007, 85, 382–386. [Google Scholar] [CrossRef]
- Lechthaler, S.; Kiorapostolou, N.; Pitacco, A.; Anfodillo, T.; Petit, G. The total path length hydraulic resistance according to known anatomical patterns: What is the shape of the root-to-leaf tension gradient along the plant longitudinal axis? J. Theor. Biol. 2020, 502, 110369. [Google Scholar] [CrossRef]
- Gerlin, L.; Cottret, L.; Cesbron, S.; Taghouti, G.; Jacques, M.A.; Genin, S.; Baroukh, C. Genome-scale investigation of the metabolic determinants generating bacterial fastidious growth. mSystems 2020, 5, e00698-19. [Google Scholar] [CrossRef]
- Zhang, Y.; Carmesin, C.; Kaack, L.; Klepsch, M.M.; Kotowska, M.; Matei, T.; Schenk, H.J.; Weber, M.; Walther, P.; Schmidt, V.; et al. High porosity with tiny pore constrictions and unbending pathways characterize the 3D structure of intervessel pit membranes in angiosperm xylem. Plant Cell Environ. 2020, 43, 116–130. [Google Scholar] [CrossRef]
- Van Ieperen, W.; Van Meeteren, U.; Van Gelder, H. Fluid ionic composition influences hydraulic conductance of xylem conduits. J. Exp. Bot. 2000, 51, 769–776. [Google Scholar] [CrossRef]
- Goodwin, P.H.; DeVay, J.E.; Meredith, C.P. Physiological responses of Vitis vinifera cv. ‘Chardonnay’ to infection by the Pierce’s disease bacterium. Physiol. Mol. Plant Pathol. 1988, 32, 17–32. [Google Scholar] [CrossRef]
- Caruso Machado, E. Trocas gasosas e relações hídricas de laranjeira “Valência” enxertada sobre quatro porta-enxertos e submetida à deficiências hídrica. Bragantia 1998, 57, 15–22. [Google Scholar] [CrossRef]
- Purcell, A.H.; Saunders, S.R. Fate of Pierce’s Disease strains of Xylella fastidiosa in common riparian plants in California. Plant Dis. 2007, 83, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Purcell, A. Paradigms: Examples from the bacterium Xylella fastidiosa. Annu. Rev. Phytopathol. 2013, 51, 339–356. [Google Scholar] [CrossRef] [PubMed]
- Bragard, C.; Dehnen-Scmutz, K.; Di Serio, F.; Gonthier, P.; Jacques, M.-A.; Miret, J.A.J.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; Milonas, P.; et al. Update of the scientific ppinion on the risks to plant health posed by Xylella fastidiosa in the EU territory. EFSA J. 2019, 17, e05665. [Google Scholar] [CrossRef] [PubMed]
- Purcell, A.H.; Hopkins, D.L. Fastidious xylem-limited bacterial plant pathogens. Annu. Rev. Phytopathol. 2003, 34, 131–151. [Google Scholar] [CrossRef]
- Román-Écija, M.; Navas-Cortés, J.A.; Velasco-Amo, M.P.; Arias-Giraldo, L.F.; Gómez, L.M.; Fuente, L.; Landa, B.B. Two Xylella fastidiosa subsp. multiplex strains isolated from almond in Spain differ in plasmid content and virulence traits. Phytopathology 2022, 113, 960–974. [Google Scholar] [CrossRef]
- McElrone, A.J.; Sherald, J.L. Effects of water stress on symptomatology and growth of Parthenocissus quinquefolia infected by Xylella fastidiosa. Plant Dis. 2001, 85, 1160–1164. [Google Scholar] [CrossRef]
- Queiroz-Voltan, R.B.; Perosin Cabral, L.; Paradela Filho, O. Severidade do sintoma da bactéria Xylella fastidiosa em cultivares de cafeeiro. Bragantia 2004, 63, 395–404. [Google Scholar] [CrossRef]
- Hopkins, D.L. Biological control of Pierce’s Disease in the vineyard with strains of Xylella fastidiosa benign to grapevine. Plant Dis. 2005, 89, 1348–1352. [Google Scholar] [CrossRef]
- Kandel, P.P.; Almeida, R.P.P.; Cobine, P.A.; De La Fuente, L. Natural competence rates are variable among Xylella fastidiosa strains and homologous recombination occurs in vitro between subspecies fastidiosa and multiplex. Mol. Plant-Microbe Interact.–MPMI 2017, 30, 589–600. [Google Scholar] [CrossRef]
- Girelli, C.R.; Hussain, M.; Verweire, D.; Oehl, M.; Massana-Codina, J.; Avendaño, S.; Migoni, D.; Scortichini, M.; Fanizzi, F.P. Agro-active endo-therapy treated Xylella fastidiosa subsp. pauca-infected olive trees assessed by the first 1H-NMR-based metabolomic study. Scient. Rep. 2022, 12, 5973. [Google Scholar] [CrossRef] [PubMed]
- DRE. Decreto-Lei n.o 67/2020, de 15 de Setembro. Available online: https://dre.pt/dre/detalhe/decreto-lei/67-2020-142870334 (accessed on 23 April 2023).
- DRE. Portaria n.o 243/2020, de 14 de Outubro. Available online: https://dre.pt/dre/detalhe/portaria/243-2020-145359683 (accessed on 23 April 2023).
- Saponari, M.; Giampetruzzi, A.; Loconsole, G.; Boscia, D.; Saldarelli, P. Xylella fastidiosa in olive in Apulia: Where we stand. Phytopathology 2019, 109, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Grebus, M.E.; Henry, J.M. Evaluation of pruning as a method to reduce damage by oleander leaf scorch. Slosson Rep. 1999, 98–99, 1–3. [Google Scholar]
- Beretta, M.J.G.; Rodas, V.; Junior, A.G.; Derrick, K.S. Control of citrus variegated chlorosis by pruning. Int. Organ. Citrus Virol. Conf. Proc. (1957–2010) 1996, 13, 378–379. [Google Scholar] [CrossRef]
- Queiroz-Voltan, R.B.; Cabral, L.P.; Filho, O.P.; Fazuoli, L.C. Eficiência da poda em cafeeiros no controle da Xylella fastidiosa. Bragantia 2006, 65, 433–440. [Google Scholar] [CrossRef]
- Feil, H.; Feil, W.S.; Purcell, A.H. Effects of date of inoculation on the within-plant movement of Xylella fastidiosa and persistence of Pierce’s Disease within field grapevines. Phytopathology 2007, 93, 244–251. [Google Scholar] [CrossRef]
- Alves, E.; Kitajima, E.W.; Leite, B. Interaction of Xylella fastidiosa with different cultivars of Nicotiana tabacum: A comparison of colonization patterns. J. Phytopathol. 2003, 151, 500–506. [Google Scholar] [CrossRef]
- Haviland, D.R.; Viveros, M.A. Surveys for Almond Leaf Scorch in Kern County, CA, and Implications on Pruning as a Tool for Management. Annual Report to the Almond Board of California. 2004. Available online: https://cekern.ucanr.edu/files/98498.pdf (accessed on 25 March 2023).
- Bucci, E.M. Xylella fastidiosa, a new plant pathogen that threatens global farming: Ecology, molecular biology, search for remedies. Biochem. Biophys. Res. Commun. 2018, 502, 173–182. [Google Scholar] [CrossRef]
- Feil, H.; Purcell, A.H. Temperature-dependent growth and survival of Xylella fastidiosa in vitro and in potted grapevines. Plant Dis. 2007, 85, 1230–1234. [Google Scholar] [CrossRef]
- Lieth, J.H.; Meyer, M.M.; Yeo, K.H.; Kirkpatrick, B.C. Modeling cold curing of Pierce’s Disease in Vitis vinifera ‘Pinot Noir’ and ‘Cabernet Sauvignon’ grapevines in California. Phytopathology 2011, 101, 1492–1500. [Google Scholar] [CrossRef]
- Amanifar, N.; Taghavi, M.; Salehi, M. Xylella fastidiosa from almond in Iran: Overwinter recovery and effects of antibiotics. Phytopathol. Mediterr. 2016, 55, 337–345. [Google Scholar] [CrossRef]
- Krivanek, A.F.; Walker, M.A. Vitis resistance to Pierce’s Disease is characterized by differential Xylella fastidiosa populations in stems and leaves. Phytopathology 2005, 95, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Alves De Souza, A.; Aurélio Takita, M.; Morais Do Amaral, A.; Coletta-Filho, H.D.; Machado, M.A. Citrus responses to Xylella fastidiosa infection, the causal agent of citrus variegated chlorosis. Tree Forest. Sci. Biotechnol. 2009, 3, 73–80. Available online: www.fundecitrus.com (accessed on 25 March 2023).
- Boscia, D.; Altamura, G.; Saponari, M.; Tavano, D.; Zicca, S.; Pollastro, P.; Silletti, M.R.; Savino, V.N.; Martelli, G.P.; Delle Done, A.; et al. Incidenza di Xylella in oliveti con disseccamento rapido. L’Informatore Agrar. 2017, 27, 47–51. [Google Scholar]
- Mauricio, F.N.; Soratto, T.A.T.; Diogo, J.A.; Boscariol-Camargo, R.L.; De Souza, A.A.; Coletta-Filho, H.D.; Silva, J.A.A.; Medeiros, A.H.; Machado, M.A.; Cristofani-Yaly, M. Analysis of defense-related gene expression in citrus hybrids infected by Xylella fastidiosa. Phytopathology 2019, 109, 301–306. [Google Scholar] [CrossRef]
- Muranaka, L.S.; Giorgiano, T.E.; Takita, M.A.; Forim, M.R.; Silva, L.F.C.; Coletta-Filho, H.D.; Machado, M.A.; de Souza, A.A. N-Acetylcysteine in agriculture, a novel use for an old molecule: Focus on controlling the plant-pathogen Xylella fastidiosa. PLoS ONE 2013, 8, e72937. [Google Scholar] [CrossRef]
- Lacava, P.T.; Araújo, W.L.; Maccheroni, W.; Azevedo, J.L. RAPD profile and antibiotic susceptibility of Xylella fastidiosa, causal agent of citrus variegated chlorosis. Lett. Appl. Microbiol. 2001, 33, 302–306. [Google Scholar] [CrossRef]
- Blonda, P.; Tarantino, C.; Scortichini, M.; Tarantino, M.; Adamo, M. Monitoring the effectiveness of a bio-fertilizer restoration technique using multi-resolution satellite and meteo-data: The case of Xylella fastidiosa subsp. pauca. Res. Sq. 2023. preprint. [Google Scholar] [CrossRef]
- Muranaka, L.S.; Takita, M.A.; Olivato, J.C.; Kishi, L.T.; de Souza, A.A. Global expression profile of biofilm resistance to antimicrobial compounds in the plant-pathogenic bacterium Xylella fastidiosa reveals evidence of persister cells. J. Bacteriol. 2012, 194, 4561–4569. [Google Scholar] [CrossRef]
- Lee, M.W.; Tan, C.C.; Rogers, E.E.; Stenger, D.C. Toxin-antitoxin systems mqsR/ygiT and dinJ/relE of Xylella fastidiosa. Physiol. Mol. Plant. Pathol. 2014, 87, 59–68. [Google Scholar] [CrossRef]
- Merfa, M.V.; Niza, B.; Takita, M.A.; De Souza, A.A. The MqsRA toxin-antitoxin system from Xylella fastidiosa plays a key role in bacterial fitness, pathogenicity, and persister cell formation. Front. Microbiol. 2016, 7, 904. [Google Scholar] [CrossRef] [PubMed]
- Ishida, M.L.; Andersen, P.C.; Leite, B. Effect of Vitis vinifera L. cv. Chardonnay xylem fluid on cecropin B activity against Xylella fastidiosa. Physiol. Mol. Plant Pathol. 2004, 64, 73–81. [Google Scholar] [CrossRef]
- Aldrich, T.J.; Rolshausen, P.E.; Roper, M.C.; Reader, J.M.; Steinhaus, M.J.; Rapicavoli, J.; Vosburg, D.A.; Maloney, K.N. Radicinin from Cochliobolus sp. inhibits Xylella fastidiosa, the causal agent of Pierce’s Disease of grapevine. Phytochemistry 2015, 116, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Maddox, C.E.; Laur, L.M.; Tian, L. Antibacterial activity of phenolic compounds against the phytopathogen Xylella fastidiosa. Curr. Microbiol. 2010, 60, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, J.L.; Araújo, W.L.; Lacava, P.T. The diversity of citrus endophytic bacteria and their interactions with Xylella fastidiosa and host plants. Genet. Mol. Biol. 2016, 39, 476–491. [Google Scholar] [CrossRef] [PubMed]
- Dourado, M.N.; Santos, D.S.; Nunes, L.R.; da Costa de Oliveira, R.L.B.; de Oliveira, M.V.; Araújo, W.L. Differential gene expression in Xylella fastidiosa 9a5c during co-cultivation with the endophytic bacterium Methylobacterium mesophilicum SR1.6/6. J. Basic Microbiol. 2015, 55, 1357–1366. [Google Scholar] [CrossRef]
- Ahern, S.J.; Das, M.; Bhowmick, T.S.; Young, R.; Gonzalez, C.F. Characterization of novel virulent broad-host-range phages of Xylella fastidiosa and Xanthomonas. J. Bacteriol. 2014, 196, 459–471. [Google Scholar] [CrossRef]
- Das, M.; Bhowmick, T.S.; Ahern, S.J.; Young, R.; Gonzalez, C.F. Control of Pierce’s Disease by Phage. PLoS ONE 2015, 10, e0128902. [Google Scholar] [CrossRef]
- Baccari, C.; Antonova, E.; Lindow, S. Biological control of Pierce’s disease of grape by an endophytic bacterium. Phytopathology 2019, 109, 248–256. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Dung, N.T.; Suh, H.J.; Kang, S.C. Antibacterial activity of essential oil and extracts of Cleistocalyx operculatus buds against the bacteria of Xanthomonas spp. J. Am. Oil Chem. Soc. 2010, 87, 1341–1349. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential oils as potential alternative biocontrol products against plant pathogens and weeds: A review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [PubMed]
- Santiago, M.B.; Moraes, T.S.; Massuco, J.E.; Silva, L.O.; Lucarini, R.; da Silva, D.F.; Vieira, T.M.; Crotti, A.E.M.; Martins, C.H.G. In vitro evaluation of essential oils for potential antibacterial effects against Xylella fastidiosa. J. Phytopathol. 2018, 166, 790–798. [Google Scholar] [CrossRef]
- Montesinos, L.; Baró, A.; Gascón, B.; Montesinos, E. Bactericidal and plant defense elicitation activities of Eucalyptus oil decrease the severity of infections by Xylella fastidiosa on almond plants. Front. Plant Sci. 2023, 14, 868. [Google Scholar] [CrossRef]
- Meyer, M.M.; Kirkpatrick, B.C. Exogenous applications of abscisic acid increase curing of Pierce’s disease-affected grapevines growing in pots. Plant Dis. 2011, 95, 173–177. [Google Scholar] [CrossRef]
- DGAV. Autorização Excecional de Emergência N.º 2023/12. Art.º 53 do Regulamento (CE) n.º 1107/2009, de 21 de outubro, para utilização de produtos fitofarmacêuticos no controlo de potenciais vetores da bactéria Xylella fastidiosa em plantas hospedeiras, no contexto do Plano de contingência para a Xylella fastidiosa e seus vetores. 3 pp. Available online: https://www.dgav.pt/plantas/conteudo/sanidade-vegetal/inspecao-fitossanitaria/informacao-fitossanitaria/xylella-fastidiosa (accessed on 26 June 2023).
- Dongiovanni, C.; Di Carolo, M.; Fumarola, G.; Tauro, D.; Altamura, G.; Cavalieri, V. Evaluation of insecticides for the control of juveniles of Philaenus spumarius L., 2015–2017. Arthropod Manag. Tests 2018, 43, tsy073. [Google Scholar] [CrossRef]
- Bethke, J.A.; Blua, M.J.; Redak, R.A. Effect of selected insecticides on Homalodisca coagulata (Homoptera: Cicadellidae) and transmission of oleander leaf scorch in a greenhouse study. J. Econ. Entomol. 2001, 94, 1031–1036. [Google Scholar] [CrossRef] [PubMed]
- Dongiovanni, C.; Cavalieri, V.; Altamura, G.; Di Carolo, M.; Fumarola, G.; Saponari, M.; Porcelli, F. Preliminary results of comparative efficacy evalutation trials against Philaenus spumarius L., vector of Xylella fastidiosa. In Xylella fastidiosa & the Olive Quick Decline Syndrome (OQDS). A Serious Worldwide Challenge for the Safeguard of Olive Trees; D’Onghia, A.M., Brunel, S., Valentini, F., Eds.; Options Méditerranéennes–Série A Séminaires Méditerranéens; CHIEAM: Montpellier, France, 2017; Volume 121, pp. 79–80. Available online: https://om.ciheam.org/article.php?IDPDF=00007219 (accessed on 21 September 2023).
- di Domenico, C.; Ganassi, C.S.; Delfine, S.; Pistillo, M.; Germinara, G.S.; de Cristofaro, A. Biological activities of some essential oils towards Philaenus spumarius adults. IOBC-WPRS Bull. 2019, 141, 88–90. [Google Scholar]
- Lago, C.; Morente, M.; Heras-Bravo, D.; Martí-Campoy, A.; Rodríguez-Ballester, F.; Plaza, M.; Moreno, A.; Fereres, A. Dispersal ability of Neophilaenus campestris, a vector of Xylella fastidiosa, from olive groves to over-summering hosts. J. Appl. Entomol. 2021, 145, 648–659. [Google Scholar] [CrossRef]
- Avosani, S.; Nieri, R.; Mazzoni, V.; Anfora, G.; Hamouche, Z.; Zippari, C.; Vitale, M.L.; Verrastro, V.; Tarasco, E.; D’Isita, I.; et al. Intruding into a conversation: How behavioral manipulation could support management of Xylella fastidiosa and its insect vectors. J. Pest Sci. 2023, 1, 1–17. [Google Scholar] [CrossRef]
- Hoddle, M.S.; Van Driesche, R.G. Biological control of insect pests. In Encyclopedia of Insects, 2nd ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 91–101. [Google Scholar] [CrossRef]
- Stenger, D.C.; Sisterson, M.S.; Krugner, R.; Backus, E.A.; Hunter, W.B. A new Phytoreovirus infecting the glassy-winged sharpshooter (Homalodisca vitripennis). Virology 2009, 386, 469–477. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regions | Number of DZ | DZ Designations |
|---|---|---|
| Northern | 7 | Oporto Metropolitan Area |
| Sabrosa | ||
| Alijó | ||
| Baião | ||
| Mirandela | ||
| Mirandela II | ||
| Bougado | ||
| Central | 8 | Castelo Novo |
| Covilhã | ||
| Fundão | ||
| Gândaras | ||
| Marrazes | ||
| Monte Redondo | ||
| Penamacor | ||
| Póvoa de Midões | ||
| Lisbon and Tagus Valley | 3 | Lisbon Metropolitan Area |
| Colares | ||
| Palmela |
| Disease | Host | Symptoms |
|---|---|---|
| Pierce’s disease (PD) | Vine | Leaf wilting, yellow and red chlorosis with irregular distribution and dieback, green “islands” of healthy tissue, and separation of leaf from petiole |
| Olive quick decline (OQD) | Olive | Leaf scorching and rapid decline in aging olive trees with progressive death from apical to root zone |
| Citrus variegated chlorosis (CVC) | Citrus | Yellowish chlorotic spots with irregular borders beginning in the middle part of the crown and spreading over the entire plant |
| Oleander leaf scorch | Oleander | Yellowing of the leaves, which is followed by characteristic scorching and necrosis of the apical and marginal zones of the leaves |
| Bacterial leaf scorch | Oak | Irregular leaf scorching on oak trees, very evident in late summer and autumn, showing pronounced apical discoloration with a red or yellow halo between burnt and green tissues, with veins standing out yellow in apparently healthy areas |
| Almond leaf scorch | Almond | Irregular patterns of leaf necrosis causing leaf scorch, leading to a clear decrease in productivity, a progressive mortality from the apical branches, and, finally, death of the affected trees |
| Phony peach disease (PPD) | Peach | Branches with shorter internodes, smaller petiole length and leaf area, and, in a more advanced stage of infection, senescence of the more mature leaves occurs, leaving the branch leafless or with a small number of leaves at its apex |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loureiro, T.; Mesquita, M.M.; Dapkevicius, M.d.L.E.; Serra, L.; Martins, Â.; Cortez, I.; Poeta, P. Xylella fastidiosa: A Glimpse of the Portuguese Situation. Microbiol. Res. 2023, 14, 1568-1588. https://doi.org/10.3390/microbiolres14040108
Loureiro T, Mesquita MM, Dapkevicius MdLE, Serra L, Martins Â, Cortez I, Poeta P. Xylella fastidiosa: A Glimpse of the Portuguese Situation. Microbiology Research. 2023; 14(4):1568-1588. https://doi.org/10.3390/microbiolres14040108
Chicago/Turabian StyleLoureiro, Talita, Maria Manuel Mesquita, Maria de Lurdes Enes Dapkevicius, Luís Serra, Ângela Martins, Isabel Cortez, and Patrícia Poeta. 2023. "Xylella fastidiosa: A Glimpse of the Portuguese Situation" Microbiology Research 14, no. 4: 1568-1588. https://doi.org/10.3390/microbiolres14040108