
Investigation of Zebrafish Embryo Membranes at Epiboly Stage through Electrorotation Technique
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Polarization and Rotation of Eggs
2.2. ROT Chamber
2.3. Zebrafish Embryos Collection, Maintenance and Exposure
2.4. Electrorotational (ROT) Assays
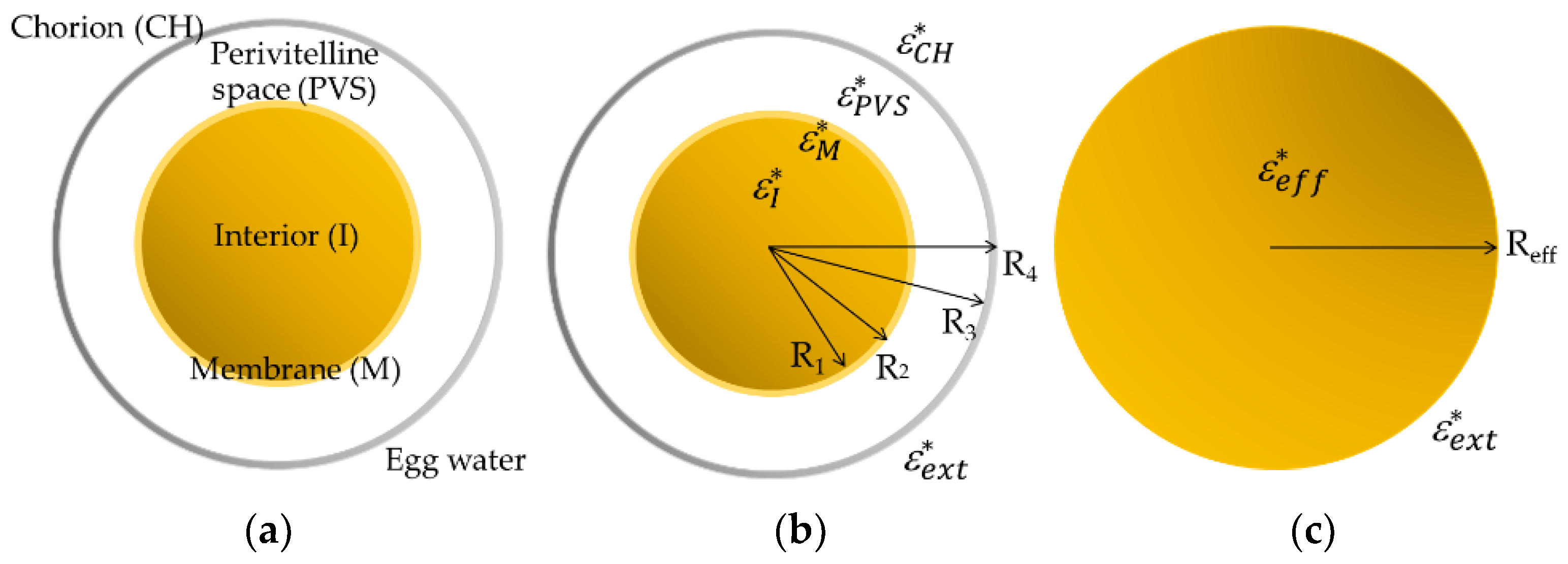
2.5. Dielectric Modelling of Zebrafish Embryos
Multi-Layer Spherical Model
3. Results
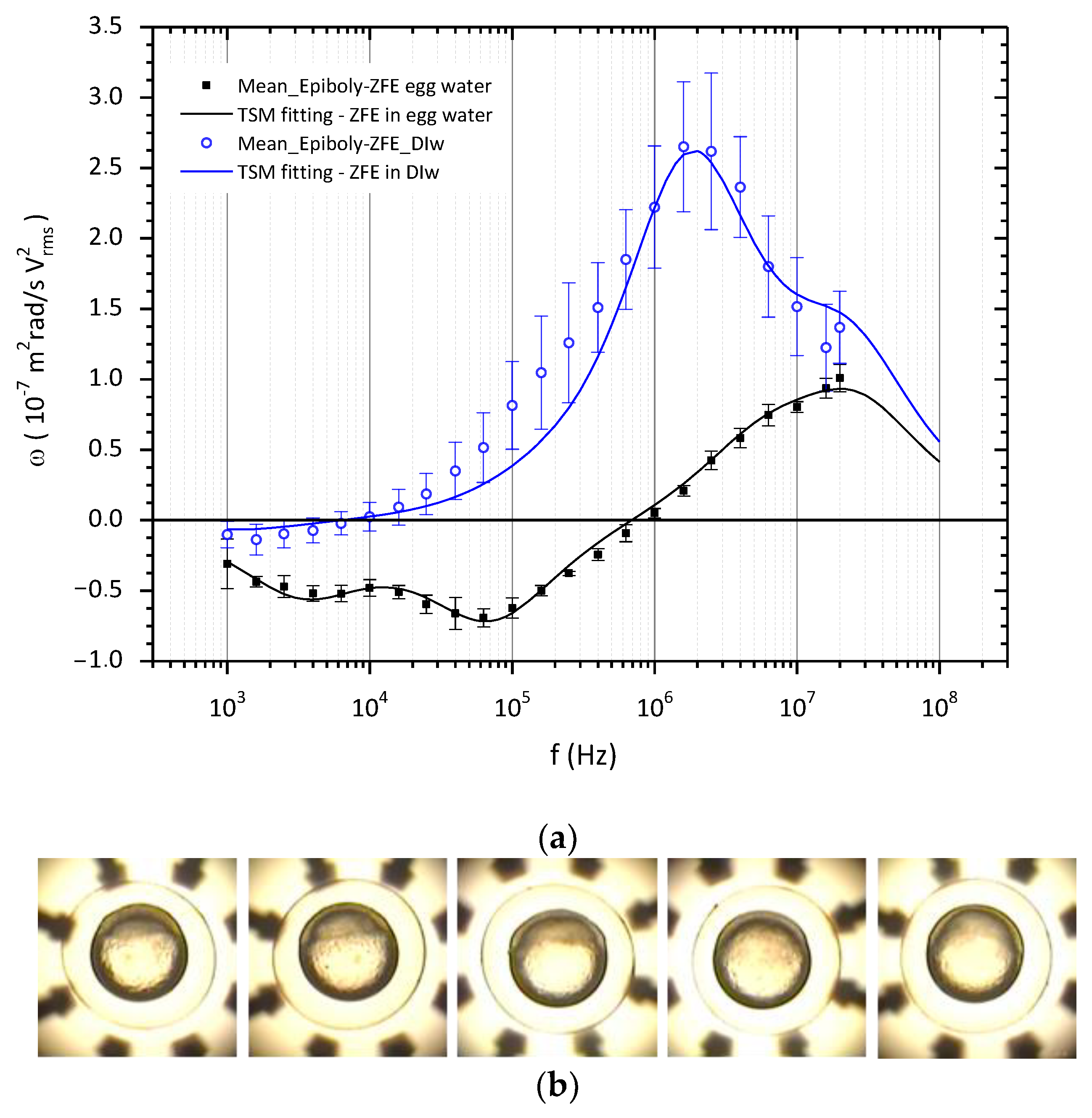
3.1. ROT Spectra and Dielectric Characterization of ZFE
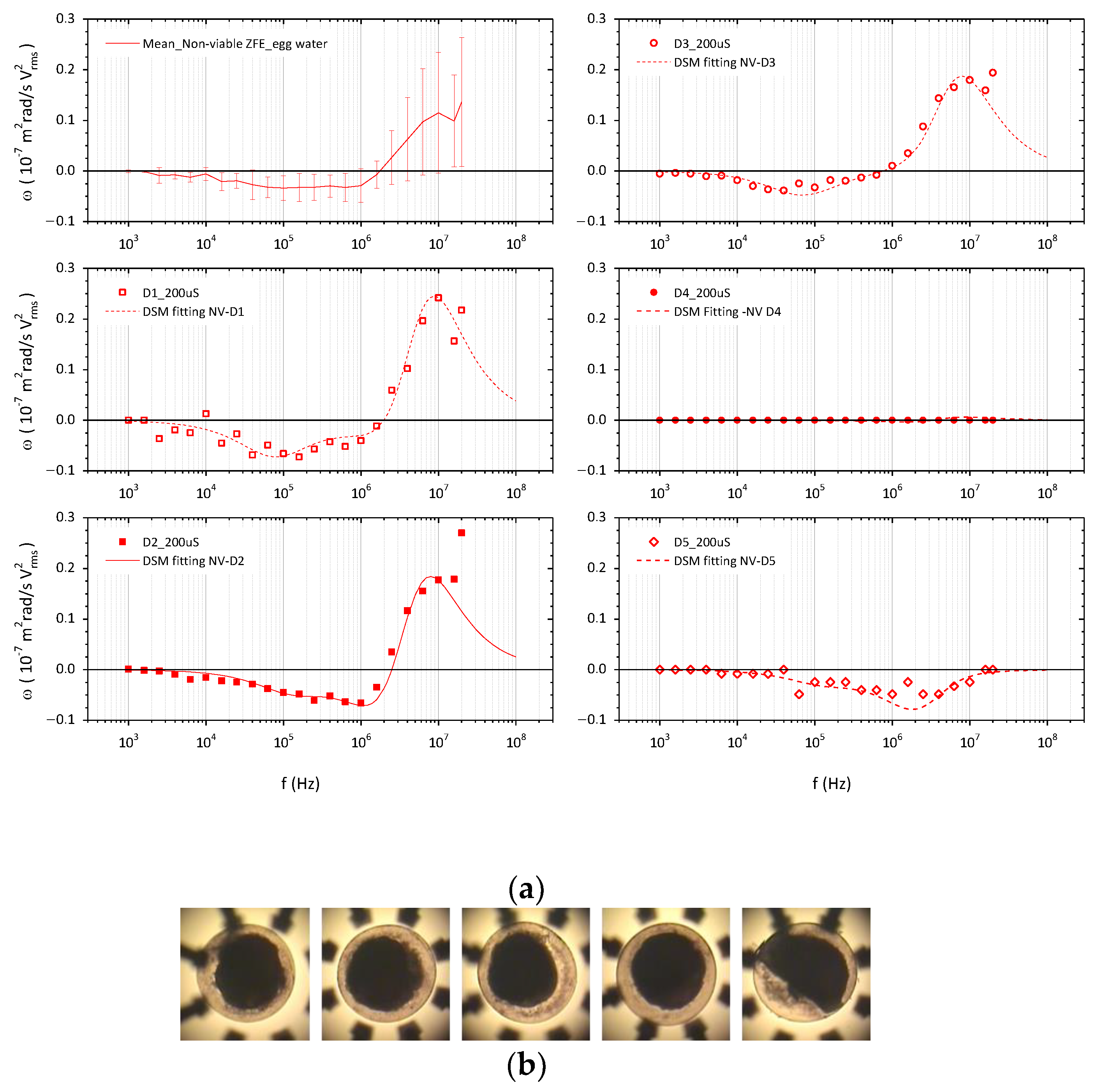
3.2. ROT Spectra and Dielectric Characterization of Non-Viable ZFE
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: Zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Bauer, B.; Mally, A.; Liedtke, D. Zebrafish embryos and larvae as alternative animal models for toxicity testing. Int. J. Mol. Sci. 2021, 22, 13417. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, L.A.; Cooper, M.S. Morphogenetic domains in the yolk syncytial layer of axiating zebrafish embryos. Dev. Dyn. 2001, 222, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Nilén, G.; Obamwonyi, O.S.; Liem-Nguyen, V.; Engwall, M.; Larsson, M.; Keiter, S.H. Observed and predicted embryotoxic and teratogenic effects of organic and inorganic environmental pollutants and their mixtures in zebrafish (Danio rerio). Aquat. Toxicol. 2022, 248, 106175. [Google Scholar] [CrossRef]
- Hamm, J.T.; Ceger, P.; Allen, D.; Stout, M.; Maull, E.A.; Baker, G.; Zmarowski, A.; Padilla, S.; Perkins, E.; Planchart, A.; et al. Characterizing sources of variability in zebrafish embryo screening protocols. Altex 2019, 36, 103–120. [Google Scholar] [CrossRef]
- Finn, R.N. The physiology and toxicology of salmonid eggs and larvae in relation to water quality criteria. Aquat. Toxicol. 2007, 81, 337–354. [Google Scholar] [CrossRef]
- Webb, S.E.; Miller, A.L. Ca2+ signaling and early embryonic patterning during the Blastula and Gastrula Periods of Zebrafish and Xenopus development. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2006, 1763, 1192–1208. [Google Scholar] [CrossRef] [PubMed]
- Paudel, S.; Sindelar, R.; Saha, M. Calcium Signaling in Vertebrate Development and Its Role in Disease. Int. J. Mol. Sci. 2018, 19, 3390. [Google Scholar] [CrossRef]
- Hernández-Vega, A.; Marsal, M.; Pouille, P.-A.; Tosi, S.; Colombelli, J.; Luque, T.; Navajas, D.; Pagonabarraga, I.; Martín-Blanco, E. Polarized cortical tension drives zebrafish epiboly movements. EMBO J. 2017, 36, 25–41. [Google Scholar] [CrossRef]
- Marsal, M.; Hernández-Vega, A.; Martin-Blanco, E. Contractility, differential tension and membrane removal lead zebrafish epiboly biomechanics. Cell Cycle 2017, 16, 1328–1335. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Nuccitelli, R. The electrical changes accompanying fertilization and cortical vesicle secretion in the medaka egg. Dev. Biol. 1980, 76, 483–498. [Google Scholar] [CrossRef]
- Schwan, H.P. Dielectric-Spectroscopy and Electro-Rotation of Biological Cells. Ferroelectrics 1988, 86, 205–223. [Google Scholar] [CrossRef]
- Muñoz, S.; Sebastián, J.L.; Antoranz, P.; García-Cambero, J.P.; Sanchis-Otero, A. Toxicity assessment of biological suspensions using the dielectric impedance spectroscopy technique. Int. J. Radiat. Biol. 2018, 94, 944–950. [Google Scholar] [CrossRef] [PubMed]
- Asami, K.; Irimajiri, A.; Hanai, T. Development of Medaka Eggs as Monitored by their Dielectric Behavior. Bull. Inst. Chem. Res. 1986, 64, 339–343. [Google Scholar]
- Asami, K.; Irimajiri, A. Dielectrospectroscopic monitoring of early embryogenesis in single frog embryos. Phys. Med. Biol. 2000, 45, 3285. [Google Scholar] [CrossRef] [PubMed]
- Arnold, W.; Zimmermann, U. Rotating-field-induced rotation and measurement of the membrane capacitance of single mesophyll cells of Avena sativa. Z. Naturforsch. C 1982, 37, 908–915. [Google Scholar] [CrossRef]
- Arnold, W.M.; Zimmermann, U. Rotation of an isolated cell in a rotating electric field. Naturwissenschaften 1982, 69, 297–298. [Google Scholar] [CrossRef]
- Fuhr, G.; Geissler, F.; Müller, T.; Hagedorn, R.; Torner, H. Differences in the rotation spectra of mouse oocytes and zygotes. BBA-Mol. Cell Res. 1987, 930, 65–71. [Google Scholar] [CrossRef]
- Arnold, W.M.; Schmutzler, R.K.; Schmutzler, A.G.; van der Ven, H.; Al-Hasani, S.; Krebs, D.; Zimmermann, U. Electro-rotation of mouse oocytes: Single-cell measurements of zona-intact and zona-free cells and of the isolated zona pellucida. Biochim. Biophys. Acta-Biomembr. 1987, 905, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Dalton, C.; Goater, A.D.; Smith, H.V. Fertilization state of Ascaris suum determined by electrorotation. J. Helminthol. 2006, 80, 25–31. [Google Scholar] [CrossRef]
- Shirakashi, R.; Mischke, M.; Fischer, P.; Memmel, S.; Krohne, G.; Fuhr, G.R.; Zimmermann, H.; Sukhorukov, V.L. Changes in the dielectric properties of medaka fish embryos during development, studied by electrorotation. Biochem. Biophys. Res. Commun. 2012, 428, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Abou-Ali, G.; Kaler, K.V.I.S.; Paul, R.; Björklund, N.K.; Gordon, R. Electrorotation of axolotl embryos. Bioelectromagnetics 2002, 23, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, R.Y.; Bao, Q.Y.; Rawson, D.M. Development of a new rapid measurement technique for fish embryo membrane permeability studies using impedance spectroscopy. Theriogenology 2006, 66, 982–988. [Google Scholar] [CrossRef]
- Zhang, C.; Frazier, J.M.; Chen, H.; Liu, Y.; Lee, J.A.; Cole, G.J. Molecular and morphological changes in zebrafish following transient ethanol exposure during defined developmental stages. Neurotoxicol. Teratol. 2014, 44, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Bodewein, L.; Schmiedchen, K.; Dechent, D.; Stunder, D.; Graefrath, D.; Winter, L.; Kraus, T.; Driessen, S. Systematic review on the biological effects of electric, magnetic and electromagnetic fields in the intermediate frequency range (300 Hz to 1 MHz). Environ. Res. 2019, 171, 247–259. [Google Scholar] [CrossRef]
- Piccinetti, C.C.; De Leo, A.; Cosoli, G.; Scalise, L.; Randazzo, B.; Cerri, G.; Olivotto, I. Measurement of the 100 MHz EMF radiation in vivo effects on zebrafish D. rerio embryonic development: A multidisciplinary study. Ecotoxicol. Environ. Saf. 2018, 154, 268–279. [Google Scholar] [CrossRef]
- Jones, T.B. Basic theory of dielectrophoresis and electrorotation. Ieee Eng. Med. Biol. Mag. 2003, 22, 33–42. [Google Scholar] [CrossRef]
- Arnold, W.M.; Zimmermann, U. Electro-rotation: Development of a technique for dielectric measurements on individual cells and particles. J. Electrost. 1988, 21, 151–191. [Google Scholar] [CrossRef]
- Sebastián, J.L.; Muñoz, S.; Sancho, M.; Martínez, G.; Alvarez, G. Electromechanical effects on multilayered cells in nonuniform rotating fields. Phys. Rev. E 2011, 84, 011926. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Holzel, R.; Pethig, R.; Wang, X.B. Differences in the AC electrodynamics of viable and non-viable yeast cells determined through combined dielectrophoresis and electrorotation studies. Phys. Med. Biol. 1992, 37, 1499–1517. [Google Scholar] [CrossRef] [PubMed]
- Bonsignorio, D.; Perego, L.; Del Giacco, L.; Cotelli, F. Structure and macromolecular composition of the zebrafish egg chorion. Zygote 1996, 4, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Rawson, D.M.; Zhang, T.; Kalicharan, D.; Jongebloed, W.L. Field emission scanning electron microscopy and transmission electron microscopy studies of the chorion, plasma membrane and syncytial layers of the gastrula-stage embryo of the zebrafish Brachydanio rerio: A consideration of the structural and functional relationships with respect to cryoprotectant penetration. Aquac. Res. 2000, 31, 325–336. [Google Scholar]
- Glaser, R.; Fuhr, G.R.; Gimsa, J. Rotation of erythrocytes, plantcells, and protoplasts in an outside rotating electric-field. Stud. Biophys. 1983, 96, 11–20. [Google Scholar]
- Fuhr, G.R.; Gimsa, J.; Glaser, R. Interpretation of electrorotation of protoplasts. 1. Theoretical considerations. Stud. Biophys. 1985, 108, 149–164. [Google Scholar]
- Terpitz, U.; Letschert, S.; Bonda, U.; Spahn, C.; Guan, C.; Sauer, M.; Zimmermann, U.; Bamberg, E.; Zimmermann, D.; Sukhorukov, V.L. Dielectric analysis and multi-cell electrofusion of the yeast pichia pastoris for electrophysiological studies. J. Membr. Biol. 2012, 245, 815–826. [Google Scholar] [CrossRef]
- Hart, N.H.; Donovan, M. Fine structure of the chorion and site of sperm entry in the egg of Brachydanio. J. Exp. Zool. 1983, 227, 277–296. [Google Scholar] [CrossRef]
- Hisaoka, K.K. Microscopic Studies of the Teleost Chorion. Trans. Am. Microsc. Soc. 1958, 77, 240–243. [Google Scholar] [CrossRef]
- Bruce, A.E.E. Zebrafish epiboly: Spreading thin over the yolk. Dev. Dyn. 2016, 245, 244–258. [Google Scholar] [CrossRef]
- Solnica-Krezel, L.; Driever, W. Microtubule arrays of the zebrafish yolk cell: Organization and function during epiboly. Development 1994, 120, 2443–2455. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.V.; Trinkaus, J.P. Electrical coupling between embryonic cells by way of extracellular space and specialized junctions. J. Cell Biol. 1970, 44, 592–610. [Google Scholar] [CrossRef] [PubMed]
- McMillan, D.B. Ovulation. In Fish Histology: Female Reproductive Systems, 1st ed.; McMillan, D.B., McMillan, D.B., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 209–284. [Google Scholar]
- Hart, N.H.; Pietri, R.; Donovan, M. The structure of the chorion and associated surface filaments in Oryzias—Evidence for the presence of extracellular tubules. J. Exp. Zool. 1984, 230, 273–296. [Google Scholar] [CrossRef] [PubMed]
- Cameron, I.L.; Hunter, K.E. Regulation of the permeability of the medaka fish embryo chorion by exogeneous sodium and calcium ions. J. Exp. Zool. 1984, 231, 447–454. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | External Medium | Chorion | PVS | Membrane | Interior | |
|---|---|---|---|---|---|---|
| δ or r (μm) | - | 3 | 210 | 0.1 | 397 | |
| Medium 1 | εr | 78 | 90 | 78 | 50 | 15 |
| σ (mS/m) | 19 | 0.25 | 30 | 1 × 10−4 | 200 | |
| Medium 2 | εr | 78 | 90 | 78 | 50 | 15 |
| σ (mS/m) | 0.115 | 0.35 ± 0.1 | 15 | 1 × 10−4 | 200 |
| NV ZFE in Egg Water | Chorion | Perivitelline Space | Egg Core | |
|---|---|---|---|---|
| D1 | εr | 90 | 40 | 10 |
| σ (mS/m) | 0.35 ± 0.05 | 6 | 18 | |
| D2 | εr | 90 | 53.5 ± 0.5 | 7 |
| σ (mS/m) | 0.55 ± 0.05 | 6 | 14 | |
| D3 | εr | 90 | 29 ± 1 | 7 |
| σ (mS/m) | 0.32 ± 0.02 | 5 | 11.5 ± 0.5 | |
| D4 | εr | 90 | 35 | 13 |
| σ (mS/m) | 30 | 6.5 | 5 | |
| D5 | εr | 90 | 53 | 6.5 ± 0.5 |
| σ (mS/m) | 0.55 ± 0.05 | 6 | 7 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanchis-Otero, A.; Reina-Maldonado, M.T.; Roldán, J.; Barragán, V.M.; Muñoz, S. Investigation of Zebrafish Embryo Membranes at Epiboly Stage through Electrorotation Technique. Membranes 2023, 13, 785. https://doi.org/10.3390/membranes13090785
Sanchis-Otero A, Reina-Maldonado MT, Roldán J, Barragán VM, Muñoz S. Investigation of Zebrafish Embryo Membranes at Epiboly Stage through Electrorotation Technique. Membranes. 2023; 13(9):785. https://doi.org/10.3390/membranes13090785
Chicago/Turabian StyleSanchis-Otero, Aránzazu, María Teresa Reina-Maldonado, José Roldán, Vicenta María Barragán, and Sagrario Muñoz. 2023. "Investigation of Zebrafish Embryo Membranes at Epiboly Stage through Electrorotation Technique" Membranes 13, no. 9: 785. https://doi.org/10.3390/membranes13090785