An Immunohistochemical Study of Gastric Mucosa and Critical Review Indicate That the Subepithelial Telocytes Are Prelymphatic Endothelial Cells

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
3. Results
3.1. Gastric Histology Was Accurately Recognised
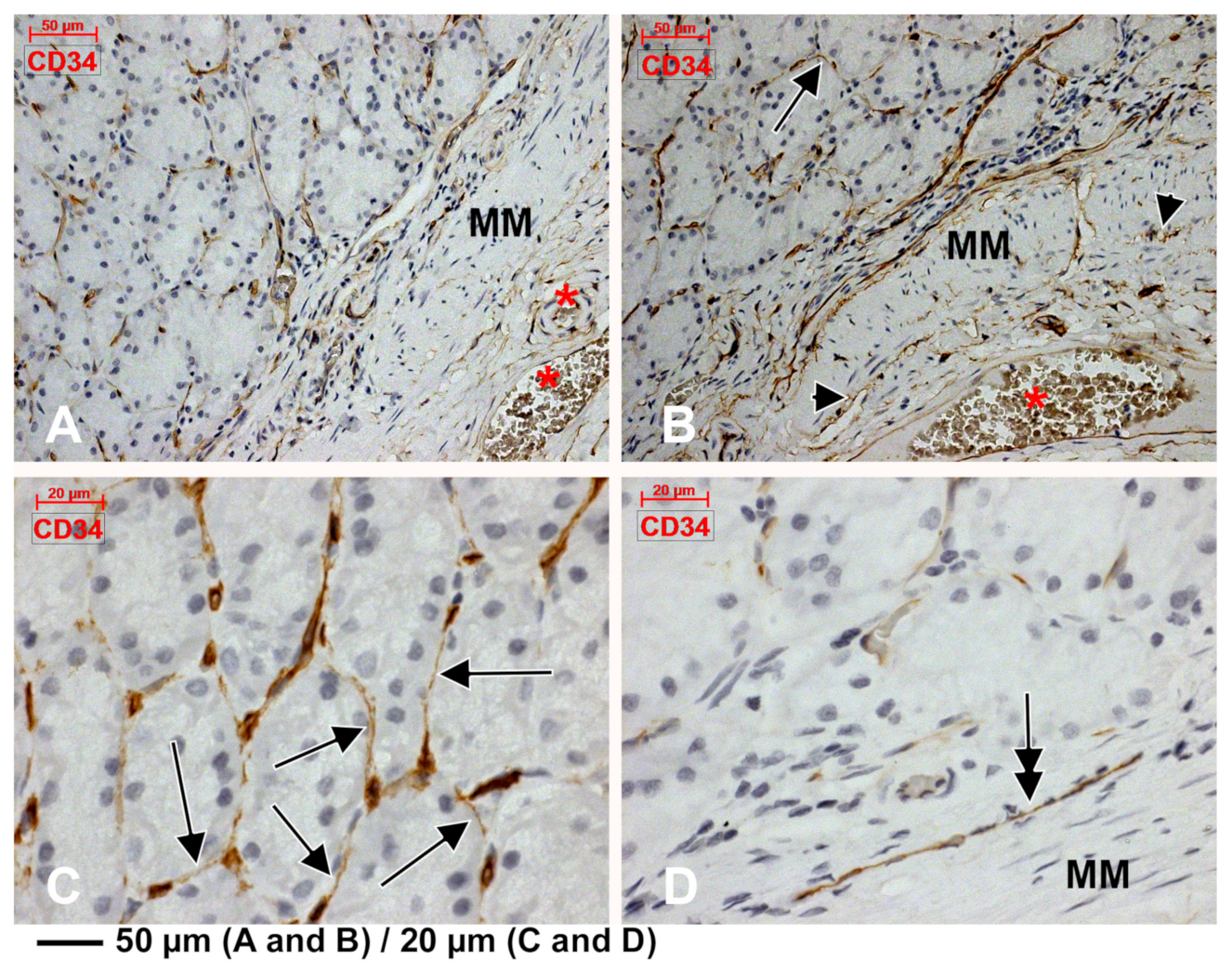
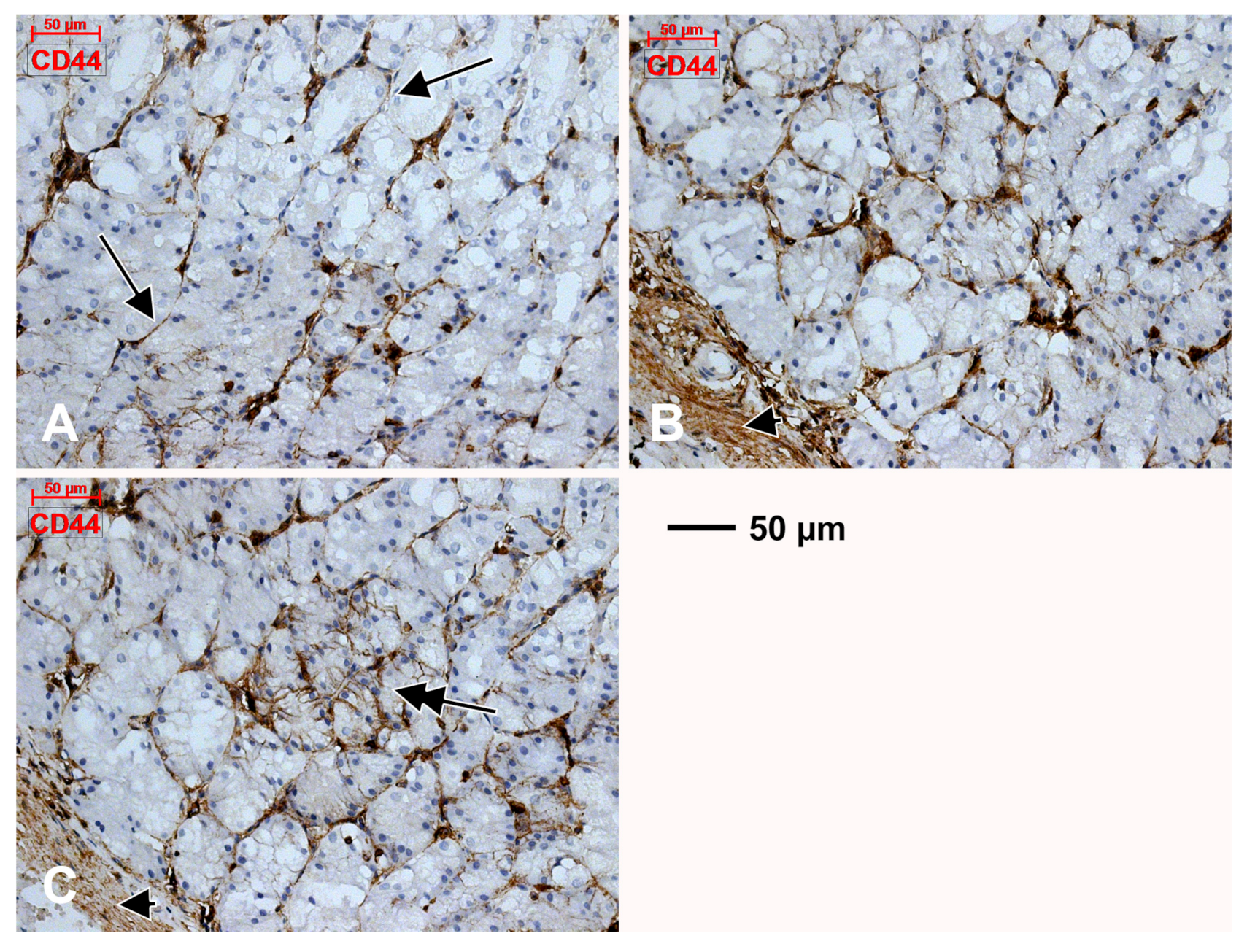
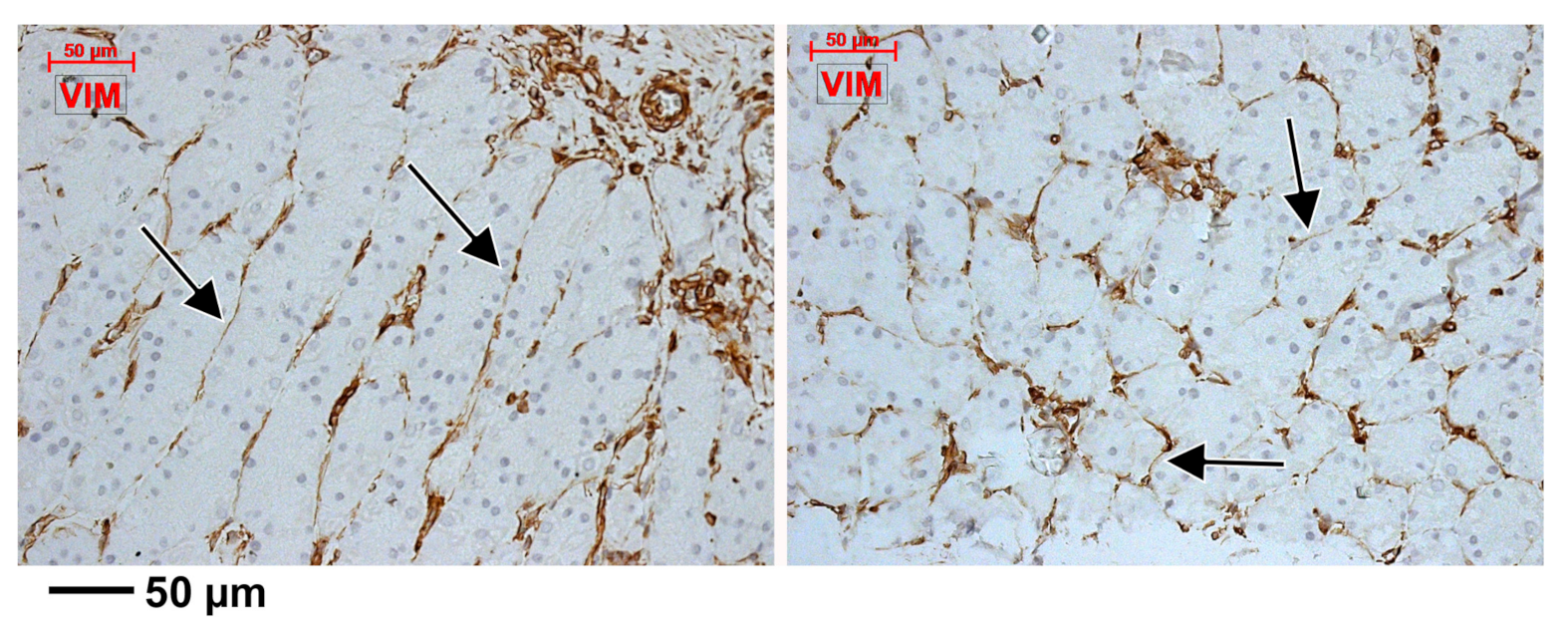
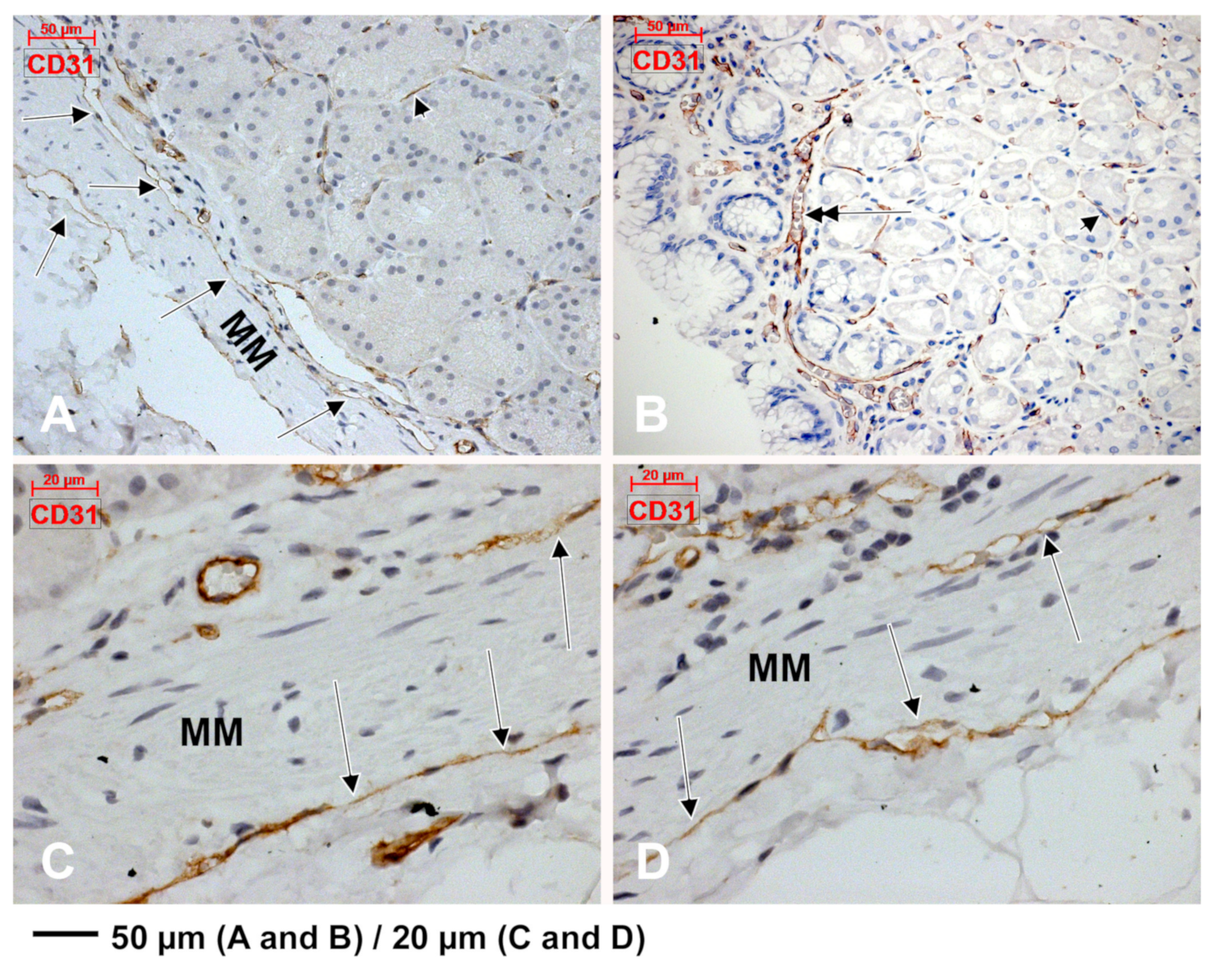
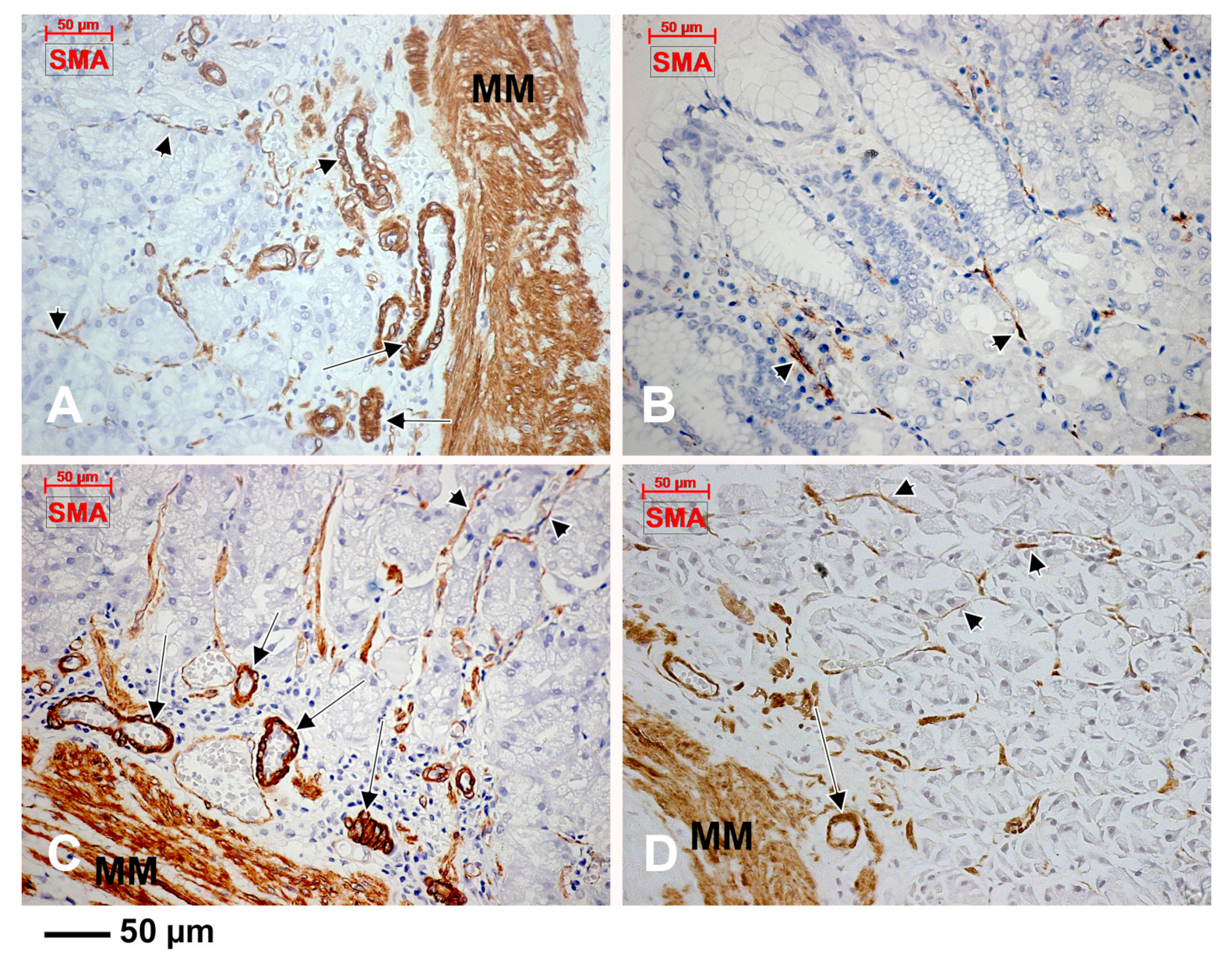
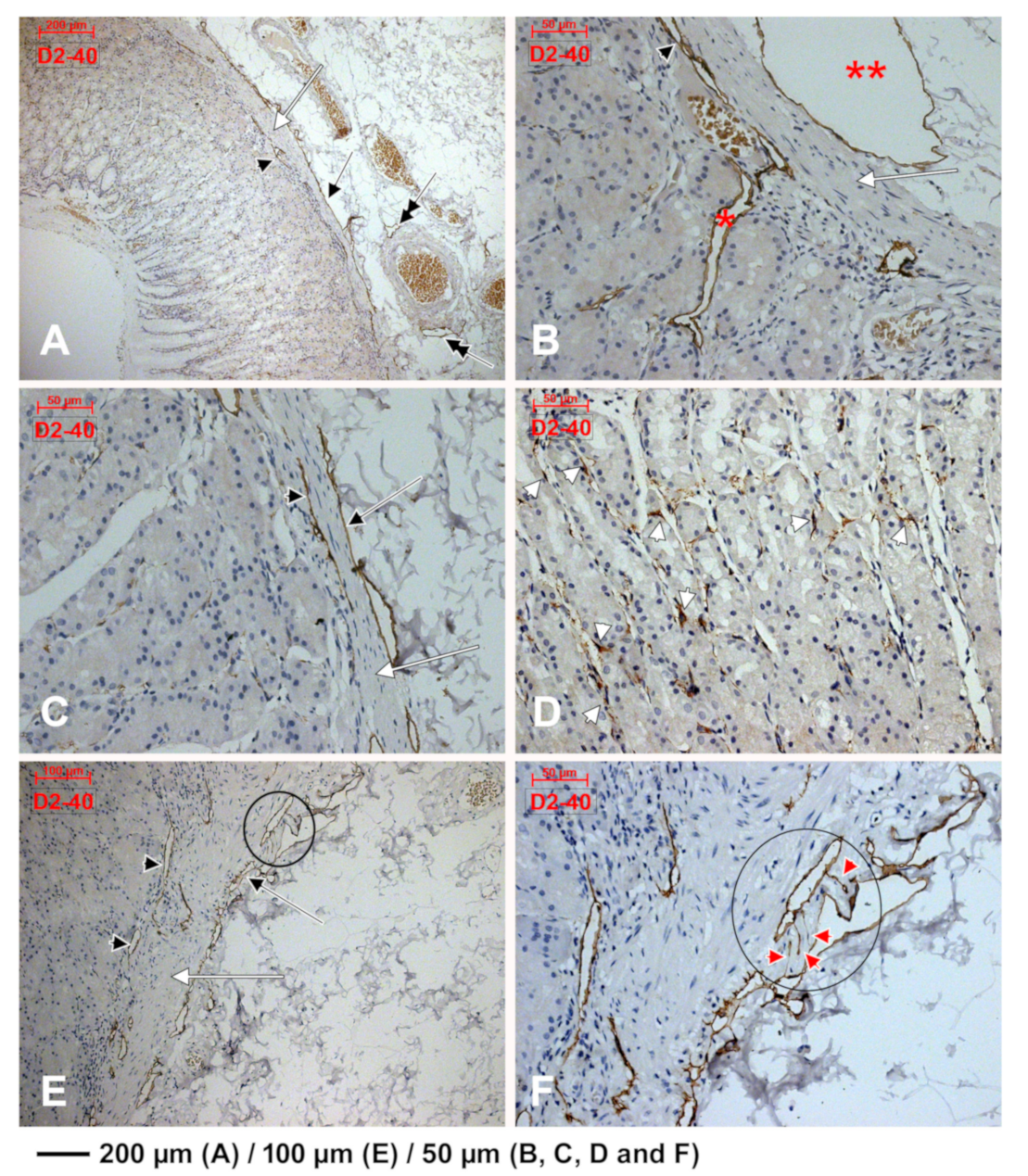
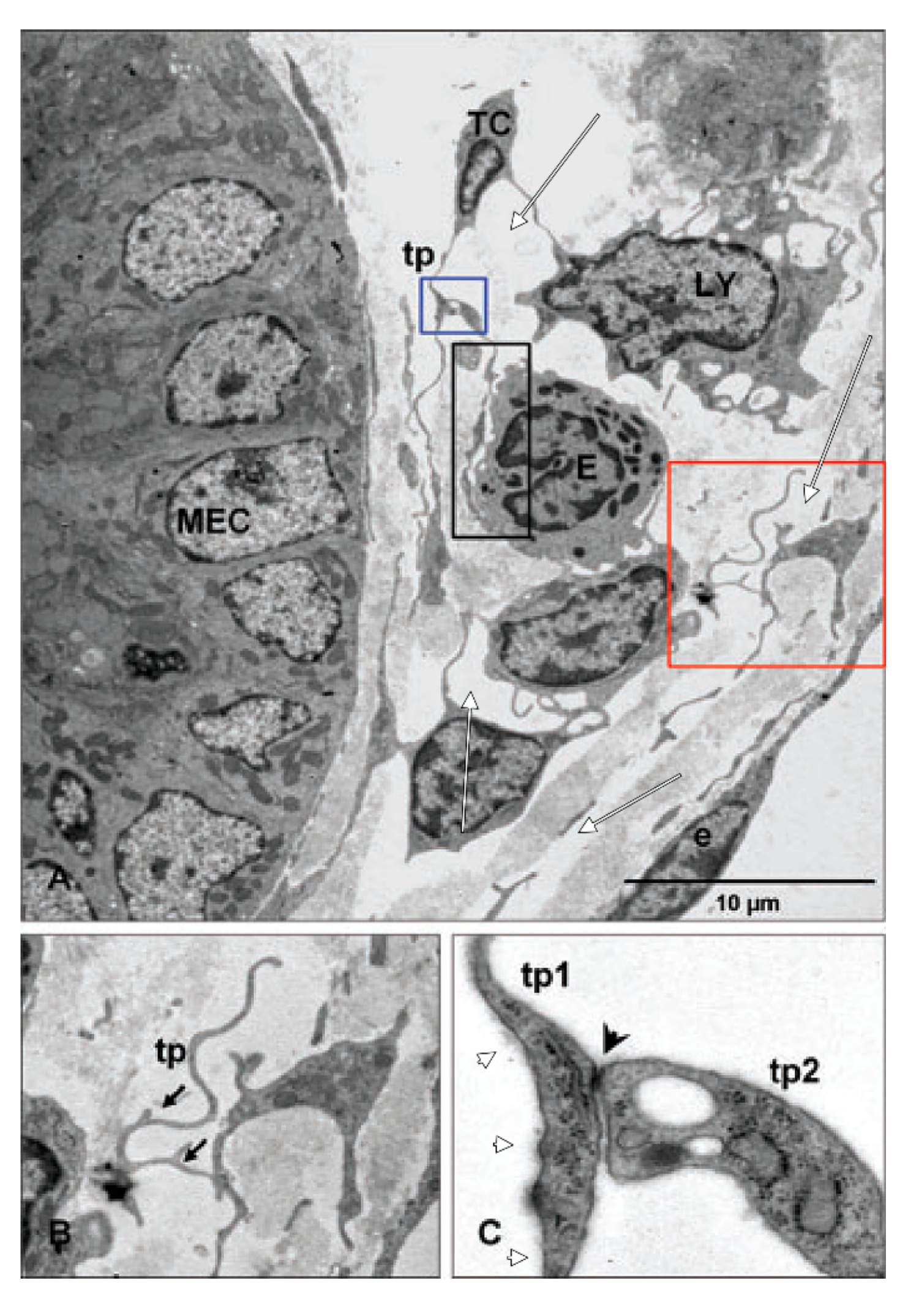
3.2. Subepithelial Telocytes (TCs) Are a Mixture of Blood and Lymphatic Endothelial Cells
4. Discussion
4.1. The Lymphatics of the Gastrointestinal Tract Were Poorly Documented
4.2. Expression of CD44 in the Gastric Mucosa
4.3. Subepithelial ‘Telocytes’ Were Previously Studied as ‘Fibrocytes’ and as ‘Pericryptal Fibroblasts’
4.4. The Markers Used on Gut Subepithelial Cells Are Not Enough for a Positive Diagnosis of Telocytes
4.5. Critical Mini-Review of the Studies Used to Assess Subepithelial Telocytes
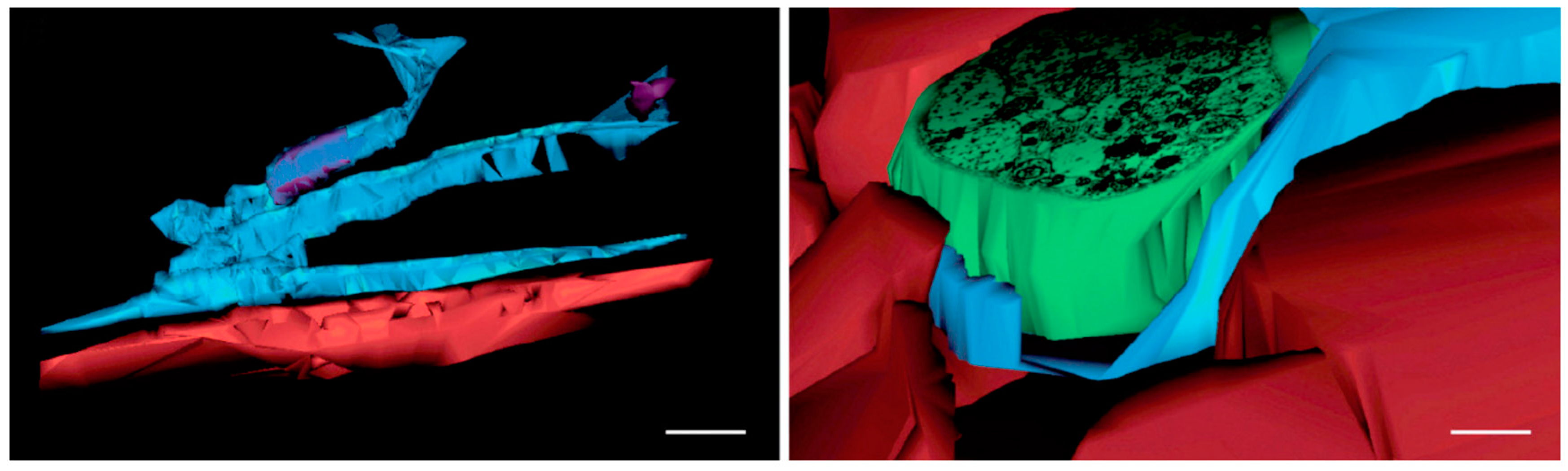
4.6. The Interstitial Organ: TC-Like Cells Limit Prelymphatic Spaces
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CSC | crypt stromal cell |
| Cx43 | Connexin 43 |
| FOXL1 | winged helix transcriptional factor |
| LEC | lymphatic endothelial cell |
| PDGFR | platelet-derived growth factor receptor |
| PF | pericryptal fibroblast |
| SETC | subepithelial telocyte |
| TC | telocyte |
| TEM | transmission electron microscopy |
References
- Rusu, M.C.; Nicolescu, M.I.; Jianu, A.M.; Lighezan, R.; Manoiu, V.S.; Păduraru, D. Esophageal telocytes and hybrid morphologies. Cell Boil. Int. 2012, 36, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Petrea, C.; Craitoiu, S.; Vrapciu, A.; Mănoiu, V.; Rusu, M. The telopode- and filopode-projecting heterogeneous stromal cells of the human sclera niche. Ann. Anat. Anat. Anz. 2018, 218, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, J.; Schroppel, B.; Dass, M.; Zimmermann, H.; Wolburg, H.; Fallier-Becker, P.; Gevaert, T.; Burkhardt, C.J.; Do, H.M.; Stolzenburg, J.U. 3D-electron microscopic characterization of interstitial cells in the human bladder upper lamina propria. Neurourol. Urodyn. 2018, 37, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Varga, I.; Kyselovič, J.; Danišovič, Ľ.; Gálfiová, P.; Kachlík, D.; Polák, Š.; Klein, M. Recently discovered interstitial cells termed telocytes: distinguishing cell-biological and histological facts from fictions. Biologia 2018, 74, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Rusu, M.C.; Hostiuc, S. Critical review: Cardiac telocytes vs cardiac lymphatic endothelial cells. Ann. Anat. 2018, 222, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Iancu, C.B.; Rusu, M.C.; Mogoanta, L.; Hostiuc, S.; Grigoriu, M. Myocardial telocyte-like cells: A review including new evidence. Cells Tissues Organs 2018, 206, 16–25. [Google Scholar] [CrossRef]
- Varga, I.; Polák, Š.; Kyselovič, J.; Kachlík, D.; Danišovič, Ľ.; Klein, M. Recently Discovered Interstitial Cell Population of Telocytes: Distinguishing Facts from Fiction Regarding Their Role in the Pathogenesis of Diverse Diseases Called “Telocytopathies.”. Medicina 2019, 55, 56. [Google Scholar] [CrossRef]
- Felisbino, S.L.; Sanches, B.D.A.; Delella, F.K.; Scarano, W.R.; Dos Santos, F.C.A.; Vilamaior, P.S.L.; Taboga, S.R.; Justulin, L.A. Prostate telocytes change their phenotype in response to castration or testosterone replacement. Sci. Rep. 2019, 9, 3761. [Google Scholar] [CrossRef]
- Pellegrini, M.-S.F.; Popescu, L.M. Telocytes. Biomol. Concepts 2011, 2, 481–489. [Google Scholar] [CrossRef]
- Vannucchi, M.-G.; Traini, C.; Manetti, M.; Ibba-Manneschi, L.; Faussone-Pellegrini, M.-S.; Ibba-Manneschi, L.; Faussone-Pellegrini, M. Telocytes express PDGFRα in the human gastrointestinal tract. J. Cell. Mol. Med. 2013, 17, 1099–1108. [Google Scholar] [CrossRef]
- Petre, N.; Rusu, M.C.; Pop, F.; Jianu, A.M. Telocytes of the mammary gland stroma. Folia Morphol. 2016, 75, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Rusu, M.C.; Cretoiu, D.; Vrapciu, A.D.; Hostiuc, S.; Dermengiu, D.; Manoiu, V.S.; Cretoiu, S.M.; Mirancea, N. Telocytes of the human adult trigeminal ganglion. Cell Boil. Toxicol. 2016, 32, 199–207. [Google Scholar] [CrossRef]
- Diaz-Flores, L.; Gutierrez, R.; Garcia, M.P.; Saez, F.J.; Diaz-Flores, L., Jr.; Valladares, F.; Madrid, J.F. CD34+ stromal cells/fibroblasts/fibrocytes/telocytes as a tissue reserve and a principal source of mesenchymal cells. Location, morphology, function and role in pathology. Histol. Histopathol. 2014, 29, 831–870. [Google Scholar]
- Cimini, M.; Cannatá, A.; Pasquinelli, G.; Rota, M.; Goichberg, P. Phenotypically heterogeneous podoplanin-expressing cell populations are associated with the lymphatic vessel growth and fibrogenic responses in the acutely and chronically infarcted myocardium. PLoS ONE 2017, 12, e0173927. [Google Scholar] [CrossRef] [PubMed]
- Varga, I.; Kachlík, D.; Žišková, M.; Miko, M. Lymphatic lacunae of the mucosal folds of human uterine tubes — A rediscovery of forgotten structures and their possible role in reproduction. Ann. Anat. Anat. Anz. 2018, 219, 121–128. [Google Scholar] [CrossRef]
- Podgrabinska, S.; Braun, P.; Velasco, P.; Kloos, B.; Pepper, M.S.; Jackson, D.G.; Skobe, M.; Falter-Braun, P. Molecular characterization of lymphatic endothelial cells. Proc. Natl. Acad. Sci. USA 2002, 99, 16069–16074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lokmic, Z.T.; Ng, E.S.; Burton, M.; Stanley, E.G.; Penington, A.J.; Elefanty, A.G. Isolation of Human Lymphatic Endothelial Cells by Multi-parameter Fluorescence-activated Cell Sorting. J. Vis. Exp. 2015, 99, e52691. [Google Scholar] [CrossRef]
- Mäkinen, T.; Adams, R.H.; Bailey, J.; Lu, Q.; Ziemiecki, A.; Alitalo, K.; Klein, R.; Wilkinson, G.A.; Klein, R. PDZ interaction site in ephrinB2 is required for the remodeling of lymphatic vasculature. Genome Res. 2005, 19, 397–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoshkes-Carmel, M.; Wang, Y.J.; Wangensteen, K.J.; Toth, B.; Kondo, A.; Massasa, E.E.; Itzkovitz, S.; Kaestner, K.H.; Massassa, E.E. Subepithelial telocytes are an important source of Wnts that supports intestinal crypts. Nature 2018, 557, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Buttler, K.; Becker, J.; Pukrop, T.; Wilting, J.; Becker, J. Maldevelopment of dermal lymphatics in Wnt5a-knockout-mice. Dev. Boil. 2013, 381, 365–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muley, A.; Odaka, Y.; Lewkowich, I.P.; Vemaraju, S.; Yamaguchi, T.P.; Shawber, C.; Dickie, B.H.; Lang, R.A. Myeloid Wnt ligands are required for normal development of dermal lymphatic vasculature. PLoS ONE 2017, 12, e0181549. [Google Scholar] [CrossRef]
- Nicenboim, J.; Malkinson, G.; Lupo, T.; Asaf, L.; Sela, Y.; Mayseless, O.; Gibbs-Bar, L.; Senderovich, N.; Hashimshony, T.; Shin, M.; et al. Lymphatic vessels arise from specialized angioblasts within a venous niche. Nature 2015, 522, 56–61. [Google Scholar] [CrossRef]
- Greicius, G.; Virshup, D.M. Stromal control of intestinal development and the stem cell niche. Differentiation 2019. (E-pub ahead of print). [Google Scholar] [CrossRef]
- Nicolescu, M.I.; Bucur, A.; Dinca, O.; Rusu, M.C.; Popescu, L.M. Telocytes in parotid glands. Anat. Rec. (Hoboken) 2012, 295, 378–385. [Google Scholar] [CrossRef]
- Rusu, M.C.; Pop, F.; Hostiuc, S.; Manta, L.; Maru, N.; Grigoriu, M. Transdifferentiations and heterogeneity in the stromal niches of uterine leiomyomas. Rom. J. Morphol. Embryol. 2018, 59, 663–672. [Google Scholar]
- Manta, L.; Rusu, M.; Pop, F. What podoplanin tells us about cells with telopodes. Ann. Anat. Anat. Anz. 2018, 218, 124–128. [Google Scholar] [CrossRef]
- Listrom, M.B.; Fenoglio-Preiser, C.M. Lymphatic distribution of the stomach in normal, inflammatory, hyperplastic, and neoplastic tissue. Gastroenterol. 1987, 93, 506–514. [Google Scholar] [CrossRef]
- Ji, R.; Kato, S. Enzyme-Histochemical Study on Postnatal Development of Rat Stomach Lymphatic Vessels. Microvasc. Res. 1997, 54, 1–12. [Google Scholar] [CrossRef]
- Ji, R.-C.; Kato, S. Lymphatic Network and Lymphangiogenesis in the Gastric Wall. J. Histochem. Cytochem. 2003, 51, 331–338. [Google Scholar] [CrossRef]
- Standring, S. Gray’s Anatomy: The Anatomical Basis of Clinical Practice, 41 ed.; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Fengcai, T.; Yunxiang, W.; Mingda, H.; Yulan, L.; Lisi, H. Observation of the corrosion casts of stomach lymphatics in the rabbit. Acta Anat. Sin. 1992. Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-JPXB199204001.htm (accessed on 21 March 2019).
- Guangzhi, S. Fine distribution of the intramural lymphatics in the stomach of the Whister rats. J. Qiqihar Med. 1996. Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-QQHB199603001.htm (accessed on 21 March 2019).
- Shimoda, H.; Kato, S.; Kudo, T. Enzyme-Histochemical Demonstration of the Intramural Lymphatic Network in the Monkey Jejunum. Arch. Histol. Cytol. 1997, 60, 215–224. [Google Scholar] [CrossRef]
- Ohtani, O. Three-dimensional organization of lymphatics and its relationship to blood vessels in rat small intestine. Cell Tissue Res. 1987, 248, 365–374. [Google Scholar] [CrossRef]
- Hirai, T.; Nimura, Y.; Sakai, H. The three-dimensional microstructure of intramural lymphatics in the canine large intestine. Gastroenterol. Jpn. 1990, 25, 169–174. [Google Scholar] [CrossRef]
- Alexander, J.; Ganta, V.C.; Jordan, P.; Witte, M.H. Gastrointestinal lymphatics in health and disease. Pathophysiology 2010, 17, 315–335. [Google Scholar] [CrossRef] [Green Version]
- Lehnert, T.; A Erlandson, R.; Decosse, J.J. Lymph and blood capillaries of the human gastric mucosa. A morphologic basis for metastasis in early gastric carcinoma. Gastroenterology 1985, 89, 939–950. [Google Scholar] [CrossRef]
- Petrea, C.; Rusu, M.; Mănoiu, V.; Vrapciu, A. Telocyte-like cells containing Weibel–Palade bodies in rat lamina fusca. Ann. Anat. Anat. Anz. 2018, 218, 88–94. [Google Scholar] [CrossRef]
- Rusu, M.; Mănoiu, V.; Cretoiu, D.; Creţoiu, S.; Vrapciu, A. Stromal cells/telocytes and endothelial progenitors in the perivascular niches of the trigeminal ganglion. Ann. Anat. Anat. Anz. 2018, 218, 141–155. [Google Scholar] [CrossRef]
- Baluk, P.; McDonald, D.M. Markers for microscopic imaging of lymphangiogenesis and angiogenesis. Ann. N. Y. Acad. Sci. 2008, 1131, 1–12. [Google Scholar] [CrossRef]
- Nakayama, H.; Enzan, H.; Yasui, W. Expression of podoplanin/D2-40 in pericryptal stromal cells in superficial colorectal epithelial neoplasia. Med. Mol. Morphol. 2013, 46, 20–23. [Google Scholar] [CrossRef]
- Terpe, H.J.; Stark, H.; Prehm, P.; Günthert, U. CD44 variant isoforms are preferentially expressed in basal epithelial of non-malignant human fetal and adult tissues. Histochemistry 1994, 101, 79–89. [Google Scholar] [CrossRef]
- Mahtab, E.A.; Wijffels, M.C.; Akker, N.M.V.D.; Hahurij, N.D.; Wisse, L.J.; DeRuiter, M.C.; Uhrin, P.; Zaujec, J.; Binder, B.R.; Schalij, M.J.; et al. Cardiac malformations and myocardial abnormalities inpodoplanin knockout mouse embryos: Correlation with abnormal epicardial development. Dev. Dyn. 2008, 237, 847–857. [Google Scholar] [CrossRef]
- Deane, H.W. Some electron microscopic observations on the lamina propria of the gut, with comments on the close association of macrophages, plasma cells, and eosinophils. Anat. Rec. 1964, 149, 453–473. [Google Scholar] [CrossRef]
- Pascal, R.R.; Kaye, G.I.; Lane, M.N. Colonic Pericryptal Fibroblast Sheath: Replication, Migration, and Cytodifferentiation of a Mesenchymal Cell System in Adult Tissue. Gastroenterology 1968, 54, 835–851. [Google Scholar] [CrossRef]
- Kaye, G.; Pascal, R.R.; Lane, N. The colonic pericryptal fibroblast sheath: replication, migration, and cytodifferentiation of a mesenchymal cell system in adult tissue. 3. Replication and differentiation in human hyperplastic and adenomatous polyps. Gastroenterology 1971, 60, 515–536. [Google Scholar] [CrossRef]
- Barth, P.J.; Westhoff, C.C. CD34+ fibrocytes: morphology, histogenesis and function. Curr. Stem Cell. Res. Ther. 2007, 2, 221–227. [Google Scholar] [CrossRef]
- Vannucchi, M.G.; Traini, C. Interstitial cells of Cajal and telocytes in the gut: twins, related or simply neighbor cells? Biomol. Concepts 2016, 7, 93–102. [Google Scholar] [CrossRef]
- Holzer, T.R.; Reising, L.O.; Credille, K.M.; Schade, A.E.; Oakley, G.J. Variability in Platelet-Derived Growth Factor Receptor Alpha Antibody Specificity May Impact Clinical Utility of Immunohistochemistry Assays. J. Histochem. Cytochem. 2016, 64, 785–810. [Google Scholar] [CrossRef] [Green Version]
- Iancu, C.B.; Rusu, M.C.; Mogoantă, L.; Hostiuc, S.; Toader, O.D. The telocytes in the subepicardial niche. Appl. Sci. 2019, 9, 1615. [Google Scholar] [CrossRef]
- Stzepourginski, I.; Nigro, G.; Jacob, J.M.; Dulauroy, S.; Sansonetti, P.J.; Eberl, G.; Peduto, L. CD34+ mesenchymal cells are a major component of the intestinal stem cells niche at homeostasis and after injury. Proc. Natl. Acad. Sci. USA 2017, 114, E506–E513. [Google Scholar] [CrossRef]
- Popescu, L.M.; Faussone-Pellegrini, M.S. TELOCYTES - A case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to TELOCYTES. J. Cell. Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef]
- Ogasawara, R.; Hashimoto, D.; Kimura, S.; Hayase, E.; Ara, T.; Takahashi, S.; Ohigashi, H.; Yoshioka, K.; Tateno, T.; Yokoyama, E.; et al. Intestinal Lymphatic Endothelial Cells Produce R-Spondin3. Sci. Rep. 2018, 8, 10719. [Google Scholar] [CrossRef]
- Greenbaum, L.E.; Wells, R.G. The role of stem cells in liver repair and fibrosis. Int. J. Biochem. Cell. Biol. 2011, 43, 222–229. [Google Scholar] [CrossRef] [Green Version]
- Köhn-Gaone, J.; Gogoi-Tiwari, J.; Ramm, G.A.; Olynyk, J.; Tirnitz-Parker, J.E.E. The role of liver progenitor cells during liver regeneration, fibrogenesis and carcinogenesis. Am. J. Physiol. Liver Physiol. 2016, 310, 143–154. [Google Scholar] [CrossRef]
- Hatta, K.; Huang, M.-L.; Weisel, R.D.; Li, R.-K. Culture of rat endometrial telocytes. J. Cell. Mol. Med. 2012, 16, 1392–1396. [Google Scholar] [CrossRef]
- Kanady, J.D.; Simon, A.M. Lymphatic communication: Connexin junction, what’s your function? Lymphology 2011, 44, 95–102. [Google Scholar]
- Cha, B.; Geng, X.; Mahamud, M.R.; Zhang, J.Y.; Chen, L.; Kim, W.; Jho, E.-H.; Kim, Y.; Choi, D.; Dixon, J.B.; et al. Complementary Wnt Sources Regulate Lymphatic Vascular Development via PROX1-Dependent Wnt/β-Catenin Signaling. Cell Rep. 2018, 25, 571–584.e5. [Google Scholar] [CrossRef]
- Cretoiu, D.; Cretoiu, S.M.; A Simionescu, A.; Popescu, L.M. Telocytes, a distinct type of cell among the stromal cells present in the lamina propria of jejunum. Histol. Histopathol. 2012, 27, 1067–1078. [Google Scholar]
- Carmona, I.C.; Bartolomé, M.J.L.; Escribano, C.J. Identification of telocytes in the lamina propria of rat duodenum: transmission electron microscopy. J. Cell. Mol. Med. 2011, 15, 26–30. [Google Scholar] [CrossRef]
- Benias, P.C.; Wells, R.G.; Sackey-Aboagye, B.; Klavan, H.; Reidy, J.; Buonocore, D.; Miranda, M.; Kornacki, S.; Wayne, M.; Carr-Locke, D.L.; et al. Structure and Distribution of an Unrecognized Interstitium in Human Tissues. Sci. Rep. 2018, 8, 4947. [Google Scholar] [CrossRef]
- Popescu, L.M.; Gherghiceanu, M.; Suciu, L.C.; Manole, C.G.; Hinescu, M.E. Telocytes and putative stem cells in the lungs: electron microscopy, electron tomography and laser scanning microscopy. Cell Tissue Res. 2011, 345, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Rusu, M.C.; Jianu, A.M.; Mirancea, N.; Didilescu, A.C.; Manoiu, V.S.; Paduraru, D. Tracheal telocytes. J. Cell. Mol. Med. 2012, 16, 401–405. [Google Scholar] [CrossRef]
- Pieri, L.; Vannucchi, M.G.; Faussone-Pellegrini, M.S. Histochemical and ultrastructural characteristics of an interstitial cell type different from ICC and resident in the muscle coat of human gut. J. Cell. Mol. Med. 2008, 12, 1944–1955. [Google Scholar] [CrossRef] [Green Version]
- Vannucchi, M.G.; Traini, C. The telocytes/myofibroblasts 3-D network forms a stretch receptor in the human bladder mucosa. Is this structure involved in the detrusor overactive diseases? Ann. Anat. 2018, 218, 118–123. [Google Scholar] [CrossRef]
- Rusu, M.C.; Mirancea, N.; Manoiu, V.S.; Valcu, M.; Nicolescu, M.I.; Paduraru, D. Skin telocytes. Ann. Anat. 2012, 194, 359–367. [Google Scholar] [CrossRef]
- Marini, M.; Rosa, I.; Guasti, D.; Gacci, M.; Sgambati, E.; Ibba-Manneschi, L.; Manetti, M. Reappraising the microscopic anatomy of human testis: identification of telocyte networks in the peritubular and intertubular stromal space. Sci. Rep. 2018, 8, 14780. [Google Scholar] [CrossRef]
- Manetti, M.; Guiducci, S.; Ruffo, M.; Rosa, I.; Faussone-Pellegrini, M.S.; Matucci-Cerinic, M.; Ibba-Manneschi, L. Evidence for progressive reduction and loss of telocytes in the dermal cellular network of systemic sclerosis. J. Cell. Mol. Med. 2013, 17, 482–496. [Google Scholar] [CrossRef]
- Manetti, M.; Rosa, I.; Messerini, L.; Guiducci, S.; Matucci-Cerinic, M.; Ibba-Manneschi, L.; Matucci-Cerinic, M.; Ibba-Manneschi, L. A loss of telocytes accompanies fibrosis of multiple organs in systemic sclerosis. J. Cell. Mol. Med. 2014, 18, 253–262. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | Intramural Lymphatics | Ref. |
|---|---|---|
| Oesophagus (human) | Lymphatic plexuses in the lamina propria, submucosa and muscularis propria | [30] |
| Stomach (rabbit) | Blind-ending lymphatic capillaries in the deep mucosa; layered lymphatic plexuses in the submucosa, muscularis propria and serosa | [31] |
| Stomach (Wistar rat) | Initial lymphatics in the mucosa, built-up by extremely thin endothelial cells. Submucosa, myenteric and serosal lymphatic networks. | [32] |
| Jejunum (Japanese monkey) | Central lymphatics in the villi are connected to an irregular mucosal lymphatic network consisting of tubular lymphatics. Tubulo-saccular lymphatics build a submucosa mesh-like network. Within the muscularis propria lies a maze-like lymphatic network. The lymphatic networks are independent of the blood vessels. | [33] |
| Small intestine (rat) | Lacteals in the villi fuse and form a wide sinus at the villous base which further drains into a submucosa lymphatic plexus. The coarse myenteric lymphatic plexus and the submucosa one drain through collecting lymphatics towards the mesenteric ones. | [34] |
| Large intestine (dog) | Superficial and deep lymphatic networks in the lamina propria, beneath the blood capillary networks. Thicker lymphatics and denser networks in the submucosa. Two efferent paths to the subserosa collecting trunks, a direct one and a minor one, the later supplemented by the lymphatics of the muscularis propria. | [35] |
| Papers Which Support the Use of the Term ‘Telocytes’ for Subepithelial Mesenchymal Cells | TCs Identified in Light Microscopy (LM) | TCs Identified in Transmission Electron Microscopy (TEM) | TCs Were Differentiated from LECs |
|---|---|---|---|
| ‘Identification of telocytes in the lamina propria of rat duodenum: transmission electron microscopy’ [60] | (+): LM on semithin sections | (+) | (–) |
| ‘Telocytes, a distinct type of cell among the stromal cells present in the lamina propria of jejunum’ [59] | (+): LM on semithin sections | (+) | (–) |
| ‘Telocytes express PDGFRa in the human gastrointestinal tract’ [10] | (+): IF, IHC | (–) | (–) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toader, O.D.; Rusu, M.C.; Mogoantă, L.; Hostiuc, S.; Jianu, A.M.; Ilie, A.C. An Immunohistochemical Study of Gastric Mucosa and Critical Review Indicate That the Subepithelial Telocytes Are Prelymphatic Endothelial Cells. Medicina 2019, 55, 316. https://doi.org/10.3390/medicina55070316
Toader OD, Rusu MC, Mogoantă L, Hostiuc S, Jianu AM, Ilie AC. An Immunohistochemical Study of Gastric Mucosa and Critical Review Indicate That the Subepithelial Telocytes Are Prelymphatic Endothelial Cells. Medicina. 2019; 55(7):316. https://doi.org/10.3390/medicina55070316
Chicago/Turabian StyleToader, Oana D., Mugurel C. Rusu, Laurenţiu Mogoantă, Sorin Hostiuc, Adelina Maria Jianu, and Adrian Cosmin Ilie. 2019. "An Immunohistochemical Study of Gastric Mucosa and Critical Review Indicate That the Subepithelial Telocytes Are Prelymphatic Endothelial Cells" Medicina 55, no. 7: 316. https://doi.org/10.3390/medicina55070316