
Anti-Hyperlipidemic Effect of Fucoidan Fractions Prepared from Iceland Brown Algae Ascophyllum nodosum in an Hyperlipidemic Mice Model
Abstract
:
1. Introduction
2. Results
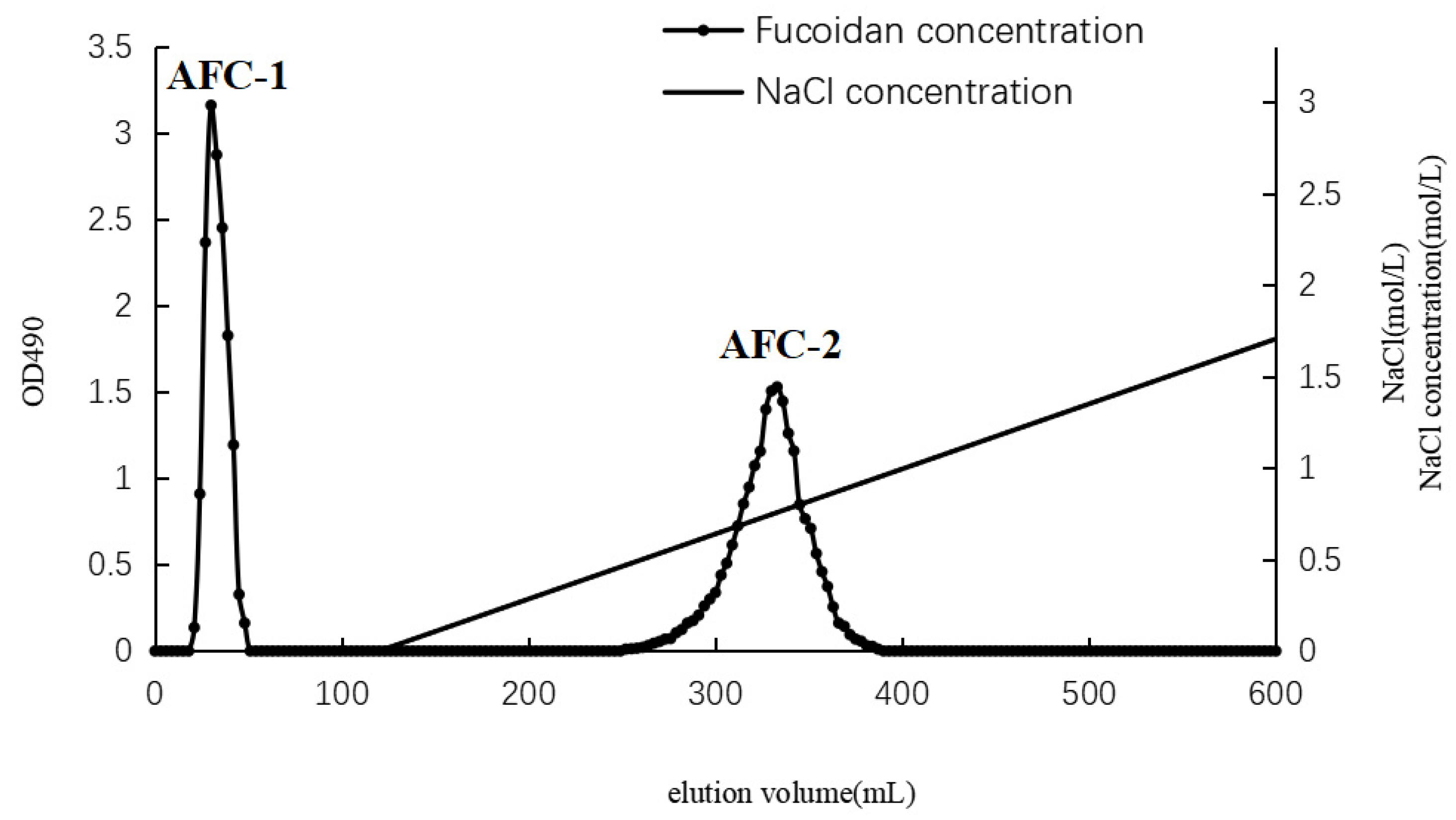
2.1. Separation and Compositional Analysis of A. nodosum Fucoidans (AN-FUC)
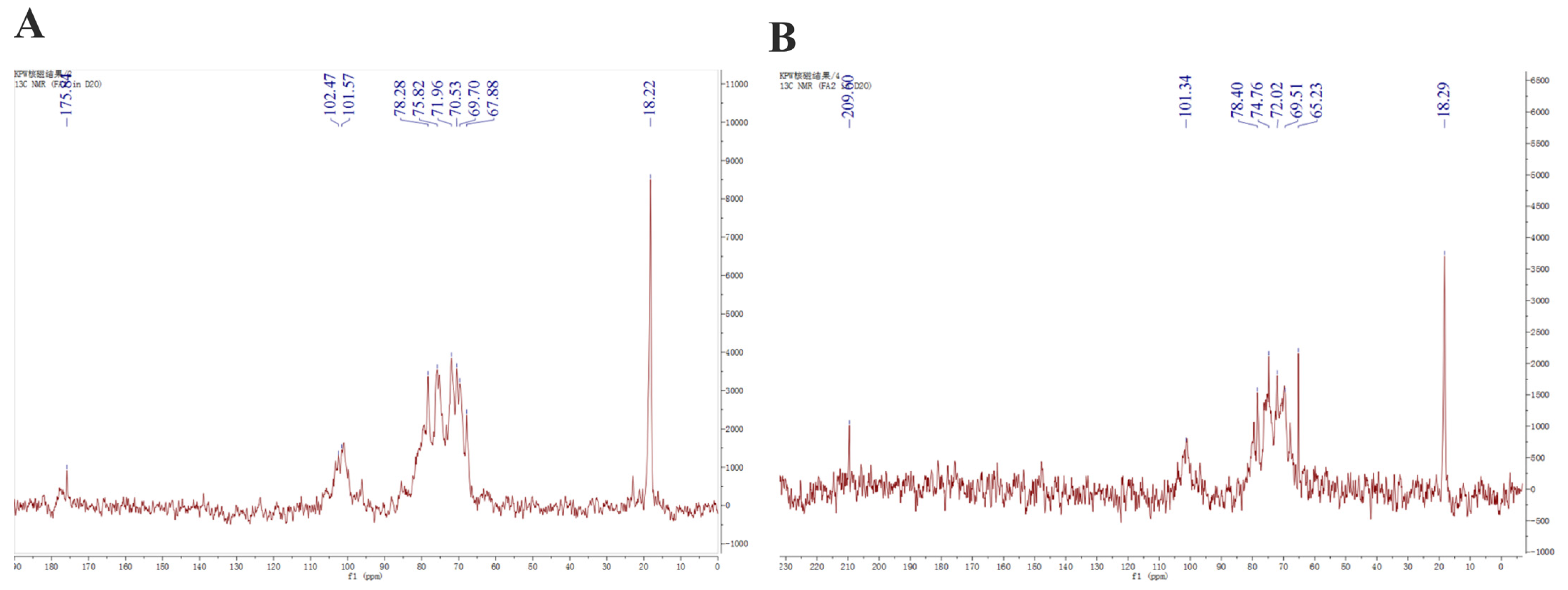
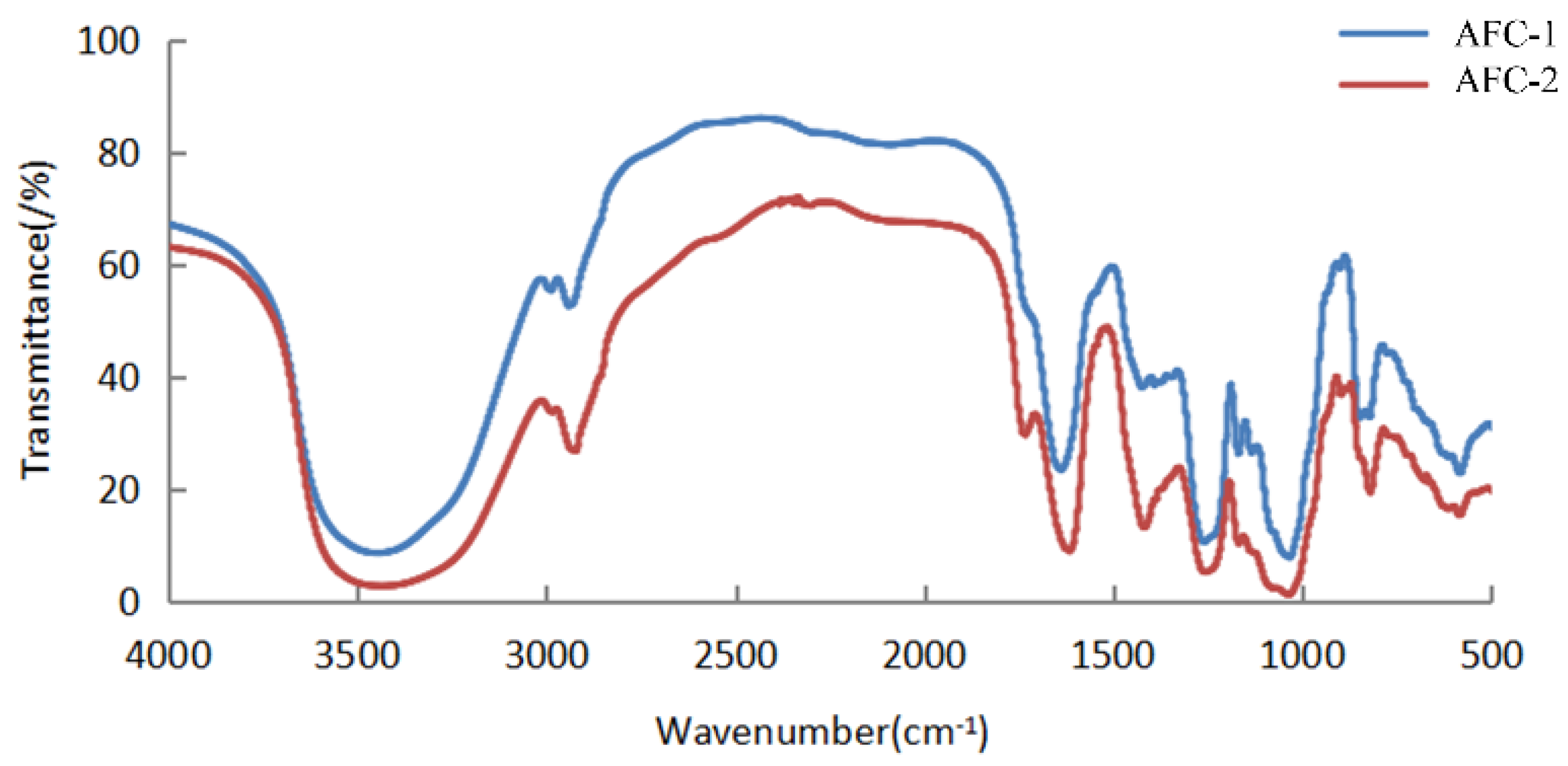
2.2. Structural Features of Fucoidans from A. nodosum
2.3. Effect of Fucoidan Administration on Physiological Parameters and Blood Glucose Level
2.4. Effect of Fucoidan Administration on Lipid Profile
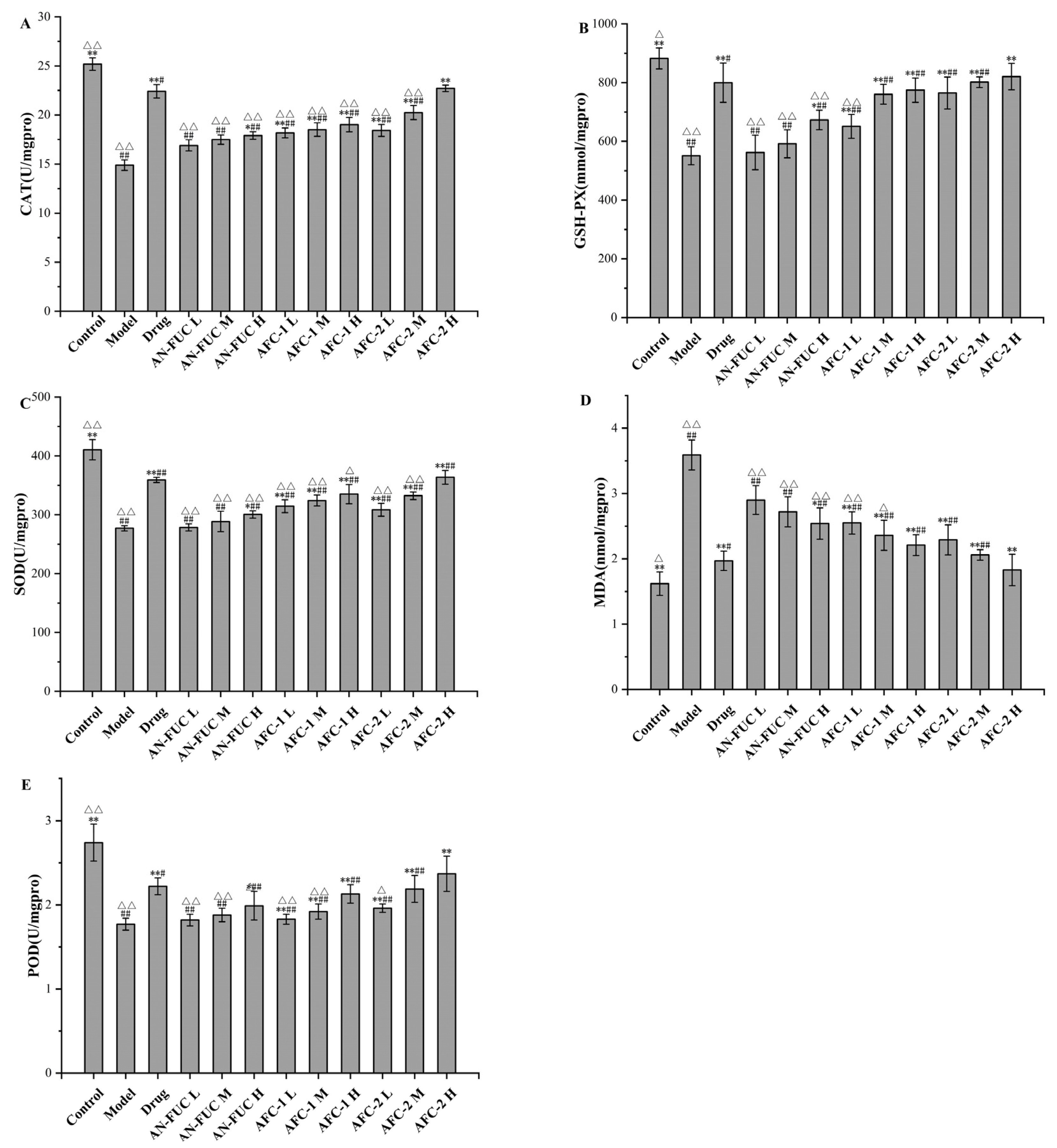
2.5. Effect of Fucoidan Administration on Oxidative-Stress-Relevant Enzyme Activities
2.6. Effect of Fucoidan Administration on Lipoprotein-Metabolism-Related Enzyme Activities
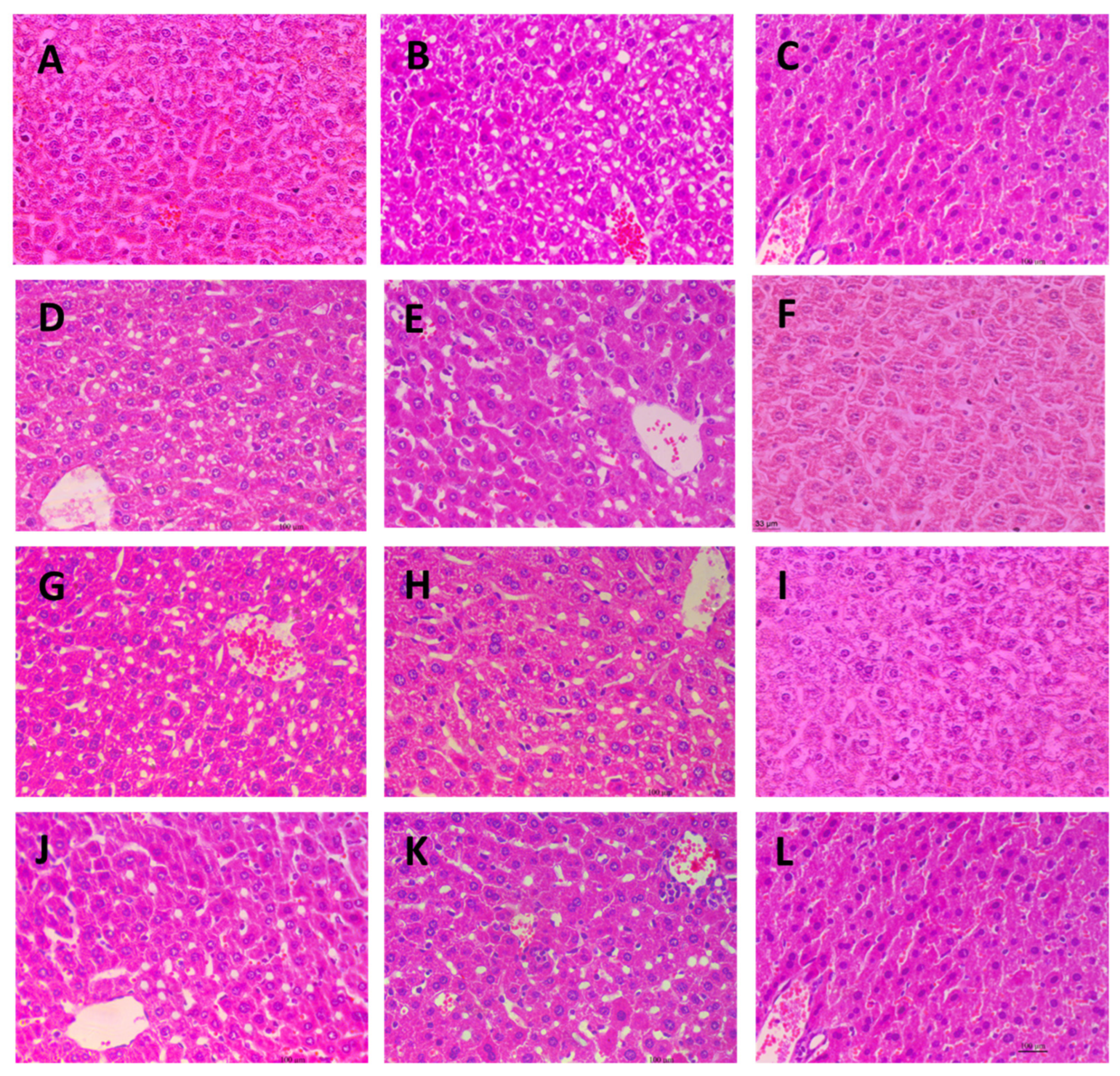
2.7. Histological Observation of Liver Tissues
2.8. Effect of Fucoidan Administration on Gut Flora Composition
3. Discussion
4. Materials and Methods
4.1. Biological Material and Reagents
4.2. Isolation, Purification, and Compositional Analysis of A. nodosum Fucoidan
4.3. Animal Experiments
4.4. Enzyme Activity Assay
4.5. Histological Observation of Liver Tissues
4.6. 16S rRNA High-Throughput Sequencing of the Gut Flora
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pereira, L.; Morrison, L.; Shukla, P.S.; Critchley, A.T. A concise review of the brown macroalga Ascophyllum nodosum (Linnaeus) Le Jolis. J. Appl. Phycol. 2020, 32, 3561–3584. [Google Scholar] [CrossRef]
- Song, Y.; Wang, Q.; Wang, Q.; He, Y.; Ren, D.; Liu, S.; Wu, L. Structural characterization and antitumor effects of fucoidans from brown algae Kjellmaniella crassifolia farmed in northern China. Int. J. Biol. Macro-Mol. 2018, 119, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Neves, N.M.; Reis, R.L.; Martins, A.; Silva, T.H. A review on fucoidan antitumor strategies: From a biological active agent to a structural component of fucoidan-based systems. Carbohydr. Polym. 2020, 239, 116131. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.-M.; Sun, Z.-X.; Makale, E.C.; Du, G.-K.; Long, W.-F.; Huang, H.-R. Antitumor activity of fucoidan: A systematic review and meta-analysis. Transl. Cancer Res. 2021, 10, 5390–5405. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, B.; Nayak, R.; Patra, S.; Bhuyan, P.P.; Behera, P.K.; Mandal, A.K.; Behera, C.; Ki, J.-S.; Adhikary, S.P.; MubarakAli, D.; et al. A state-of-the-art review on fucoidan as an antiviral agent to combat viral infections. Carbohydr. Polym. 2022, 291, 119551. [Google Scholar] [CrossRef]
- Sun, Q.-L.; Li, Y.; Ni, L.-Q.; Li, Y.-X.; Cui, Y.-S.; Jiang, S.-L.; Xie, E.-Y.; Du, J.; Deng, F.; Dong, C.-X. Structural characterization and antiviral activity of two fucoidans from the brown algae Sargassum henslowianum. Carbohydr. Polym. 2019, 229, 115487. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Song, Y.; He, Y.; Ren, D.; Kow, F.; Qiao, Z.; Liu, S.; Yu, X. Structural characterisation of algae Costaria costata fucoidan and its effects on CCl4-induced liver injury. Carbohydr. Polym. 2014, 107, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, Q.; He, Y.; Ren, D.; Kow, F.; Li, J.; Liu, S.; Cong, H. The positive effects of fucoidans extracted from the brown seaweed Saccharina japonica on protection against CCl4-induced liver injury. J. Appl. Phycol. 2017, 29, 2077–2087. [Google Scholar] [CrossRef]
- Shanthi, N.; Arumugam, P.; Murugan, M.; Sudhakar, M.P.; Arunkumar, K. Extraction of Fucoidan from Turbinaria decurrens and the Synthesis of Fucoidan-Coated AgNPs for Anticoagulant Application. ACS Omega 2021, 6, 30998–31008. [Google Scholar] [CrossRef]
- Ganapathy, S.; Lingappa, S.; Naidu, K.; Selvaraj, U.; Ramachandiran, S.; Ponnusamy, S.; Somasundaram, S.T. Isolation and Bioactive Potential of Fucoidan from Marine Macroalgae Turbinaria conoides. ChemistrySelect 2019, 4, 14114–14119. [Google Scholar] [CrossRef]
- Ahmad, T.; Eapen, M.S.; Ishaq, M.; Park, A.Y.; Karpiniec, S.S.; Stringer, D.N.; Sohal, S.S.; Fitton, J.H.; Guven, N.; Caruso, V.; et al. Anti-Inflammatory Activity of Fucoidan Extracts In Vitro. Mar. Drugs 2021, 19, 702. [Google Scholar] [CrossRef] [PubMed]
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Katsarov, P.; Nikolova, M.; Iliev, I.; Peychev, L.; Trica, B.; Oancea, F.; Delattre, C.; et al. Immunomodulatory and Anti-Inflammatory Effects of Fucoidan: A Review. Polymers 2020, 12, 2338. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Wang, Q.; Yang, Y.; Hu, Y.; Song, Y.; He, Y.; Liu, S.; Wu, L. Hypolipidemic effects of fucoidan fractions from Saccharina sculpera (Laminariales, Phaeophyceae). Int. J. Biol. Macromol. 2019, 140, 188–195. [Google Scholar] [CrossRef]
- Sirisena, A.; Okeahialam, B. Association of obesity anthropometric indices with hypertension, diabetes mellitus and hypertri-glyceridemia in apparently healthy adult Nigerian population. World J. Cardiol. 2022, 14, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Ma, W.; Wen, H. Study on effects of polysaccharides from Camellia oleifera Abel. on decreasing blood sugar. Sci. Technol. Food Ind. 2014, 35, 337. [Google Scholar]
- Gallagher, E.J.; Zelenko, Z.; Neel, B.A.; Antoniou, I.M.; Rajan, L.; Kase, N.; Leroith, D. Elevated tumor LDLR expression ac-celerates LDL cholesterol-mediated breast cancer growth in mouse models of hyperlipidemia. Oncogene 2017, 36, 6462–6471. [Google Scholar] [CrossRef]
- Hurt-Camejo, E. ANGPTL3, PCSK9, and statin therapy drive remarkable reductions in hyperlipidemia and atherosclerosis in a mouse model. J. Lipid Res. 2020, 61, 272–274. [Google Scholar] [CrossRef]
- Li, Y.; Ma, Q.; Wang, J.; Li, P.; Cheng, L.; An, Y.; Duan, Y.; Dai, H.; Wang, T.; Zhao, B. Relationship between hyperlipidemia and the gut microbiome of rats, characterized using high-throughput sequencing. J. Tradit. Chin. Med. Sci. 2020, 7, 154–161. [Google Scholar] [CrossRef]
- Wang, Y.; Han, C.; Cheng, J.; Wang, Z.; Liu, L.; Huang, H.; Liang, Q.; Liu, R.; Ran, B.; Li, W. Fermented Cerasus humilis fruits protect against high-fat diet induced hyperlipidemia which is associated with alteration of gut microbiota. J. Sci. Food Agric. 2023, 103, 2554–2563. [Google Scholar] [CrossRef]
- Li, N.; Fu, X.; Xiao, M.; Wei, X.; Yang, M.; Liu, Z.; Mou, H. Enzymatic preparation of a low-molecular-weight polysaccharide rich in uronic acid from the seaweed Laminaria japonica and evaluation of its hypolipidemic effect in mice. Food Funct. 2020, 11, 2395–2405. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, W.; Chen, X. Effect of Laminaria Polysaccharide on Blood Lipids, Nitric Oxide and Nitric Oxide Synthase in Experimental Atherosclerosis Rats. Genom. Appl. Biol. 2018, 37, 1761–1767. [Google Scholar]
- Gong, Y.; Guo, Y.; Liu, Y. Study on in Vitro Antioxidant, Hypoglycemic and Lipid-lowering Activities of Cherry Kernel Ethanol Extracts. J. Qingdao Univ. Sci. Technol. (Nat. Sci. Ed.) 2017, 38, 15–17. [Google Scholar]
- Yokota, T.; Nomura, K.; Nagashima, M.; Kamimura, N. Fucoidan alleviates high-fat diet-induced dyslipidemia and athero-sclerosis in ApoEshl mice deficient in apolipoprotein E expression. J. Nutr. Biochem. 2016, 32, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Zha, S.; Zhao, Q.; Zhao, B.; Ouyang, J.; Mo, J.; Chen, J. Molecular weight controllable degradation of Laminaria japonica polysaccharides and its antioxidant properties. J. Ocean Univ. China 2016, 15, 637–642. [Google Scholar] [CrossRef]
- Jia, X.; Xu, W.; Zhang, L.; Li, X.; Wang, R.; Wu, S. Impact of Gut Microbiota and Microbiota-Related Metabolites on Hyperlipidemia. Front. Cell. Infect. Microbiol. 2021, 11, 634780. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, Y.; Zhu, J.; Zhang, M.; Ho, C.-T.; Huang, Q.; Cao, J. Metagenomics Analysis of Gut Microbiota in a High Fat Diet-Induced Obesity Mouse Model Fed with (−)-Epigallocatechin 3-O-(3-O-Methyl) Gallate (EGCG3″Me). Mol. Nutr. Food Res. 2018, 62, e1800274. [Google Scholar] [CrossRef]
- Singh, D.P.; Singh, J.; Boparai, R.K.; Zhu, J.; Mantri, S.; Khare, P.; Khardori, R.; Kondepudi, K.K.; Chopra, K.; Bishnoi, M. Isomalto-oligosaccharides, a prebiotic, functionally augment green tea effects against high fat diet-induced metabolic alterations via preventing gut dysbacteriosis in mice. Pharmacol. Res. 2017, 123, 103–113. [Google Scholar] [CrossRef]
- Lai, Z.-L.; Tseng, C.-H.; Ho, H.J.; Cheung, C.K.Y.; Lin, J.-Y.; Chen, Y.-J.; Cheng, F.-C.; Hsu, Y.-C.; Lin, J.-T.; El-Omar, E.M.; et al. Fecal microbiota transplantation confers beneficial metabolic effects of diet and exercise on diet-induced obese mice. Sci. Rep. 2018, 8, 15625. [Google Scholar] [CrossRef]
- Sun, S.S.; Wang, K.; Ma, K.; Bao, L.; Liu, H.W. An insoluble polysaccharide from the sclerotium of Poria cocos improves hyperglycemia, hyperlipidemia and hepatic steatosis in ob/ob mice via modulation of gut microbiota. Chin. J. Nat. Med. 2019, 17, 3–14. [Google Scholar] [CrossRef]
- Alexandros, T.; Ronan, L.; Ioannis, Z. Inflammation, not Cholesterol, Is a Cause of Chronic Disease. Nutrients 2018, 10, 604. [Google Scholar]
- Conde, T.; Zabetakis, I.; Tsoupras, A.; Medina, I.; Costa, M.; Silva, J.; Neves, B.; Domingues, P.; Domingues, M. Microalgal Lipid Extracts Have Potential to Modulate the Inflammatory Response: A Critical Review. Int. J. Mol. Sci. 2021, 22, 9825. [Google Scholar] [CrossRef] [PubMed]
- Ren, R.; Gong, J.; Zhao, Y.; Zhuang, X.; Ye, Y.; Lin, W. Sulfated polysaccharides from Enteromorpha prolifera suppress SREBP-2 and HMG-CoA reductase expression and attenuate non-alcoholic fatty liver disease induced by a high-fat diet. Food Funct. 2017, 8, 1899–1904. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, Q.; Li, P.; He, Y.; Song, Y.; Hou, D.; Hou, L. Separating Fucoidan from Laminaria japonica Using Electrodialysis Technology. Prog. Fish. Sci. 2014, 35, 121–127. [Google Scholar]
- Ye, Y.; Ji, D.; You, L.; Zhou, L.; Zhao, Z.; Brennan, C. Structural properties and protective effect of Sargassum fusiforme poly-saccharides against ultraviolet B radiation in hairless Kun Ming mice. J. Funct. Foods 2018, 43, 8–16. [Google Scholar] [CrossRef]
- Zhang, P.; Jia, J.; Jiang, P.; Zheng, W.; Li, X.; Song, S.; Ai, C. Polysaccharides from edible brown seaweed Undaria pinnatifida are effective against high-fat diet-induced obesity in mice through the modulation of intestinal microecology. Food Funct. 2022, 13, 2581–2593. [Google Scholar] [CrossRef]
- Wang, S.-H.; Huang, C.-Y.; Chen, C.-Y.; Chang, C.-C.; Huang, C.-Y.; Dong, C.-D.; Chang, J.-S. Isolation and purification of brown algae fucoidan from Sargassum siliquosum and the analysis of anti-lipogenesis activity. Biochem. Eng. J. 2021, 165, 107798. [Google Scholar] [CrossRef]
- Yang, J.; Zhao, Z.; Guo, W.; Guo, J. Effects of Deoxynivalenol on Intestinal Microbiota of Mice Analyzed by Illumina-MiSeq High-Throughput Sequencing Technology. Chin. J. Anim. Nutr. 2017, 29, 158–167. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Carbohydrate Content (%) | Sulfate Content(%) | Purity (%) | Monosaccharide Composition (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Fuc | Gal | Man | Glu | Rha | Xyl | Glu-UA | Gal-UA | ||||
| AN-FUC | 62.80 ± 1.21 | 20.48 ± 0.72 | 83.28 | 58.98 | 5.08 | 5.03 | 2.04 | 6.29 | 16.15 | 6.43 | N.D |
| AFC-1 | 57.77 ± 1.44 | 27.84 ± 0.81 | 85.61 | 54.36 | 6.10 | 5.67 | 1.50 | 6.23 | 19.25 | 6.89 | N.D |
| AFC-2 | 76.18 ± 1.02 | 17.60 ± 0.56 | 93.78 | 57.25 | 7.03 | 6.25 | N.D | 6.49 | 16.76 | 6.22 | N.D |
| Group | The Initial Body Weight (g) | The Final Body Weight (g) | Liver Index (mg/g) | Blood Glucose Level (mmol/L) |
|---|---|---|---|---|
| Control | 18.80 ± 1.06 | 25.49 ± 1.99 ** | 3.62 ± 0.16 ** △ | 4.48 ± 0.61 ** △ |
| Model | 19.50 ± 0.96 | 32.09 ± 3.23 ## △ | 4.55 ± 0.12 ## △△ | 7.92 ± 0.62 ## △△ |
| Drug | 19.26 ± 1.22 | 25.94 ± 1.22 ** | 3.97 ± 0.10 ** # | 5.82 ± 0.71 ** ## |
| AN-FUC L | 18.88 ± 2.11 | 28.95 ± 3.73 | 4.45 ± 0.16 ## △△ | 6.58 ± 0.86 ** ## |
| AN-FUC M | 19.34 ± 1.26 | 27.52 ± 2.87 | 4.39 ± 0.16 * ## △△ | 7.58 ± 0.99 ## △△ |
| AN-FUC H | 18.74 ± 0.73 | 26.66 ± 2.72 * | 4.35 ± 0.13 * ## △ | 7.08 ± 0.40 ## △ |
| AFC-1 L | 18.66 ± 2.12 | 26.50 ± 2.44 * | 4.34 ± 0.22 * ## △ | 6.67 ± 0.73 * ## |
| AFC-1 M | 20.19 ± 1.71 | 29.08 ± 3.56 | 4.33 ± 0.17 ** ## △ | 6.40 ± 0.91 ** ## |
| AFC-1 H | 19.46 ± 1.01 | 28.81 ± 3.11 | 4.04 ± 0.23 ** ## | 6.23 ± 0.59 ** ## |
| AFC-2 L | 18.93 ± 1.15 | 27.20 ± 3.17 | 4.16 ± 0.22 ** ## | 5.74 ± 0.33 ** ## |
| AFC-2 M | 19.38 ± 1.34 | 26.55 ± 1.99 * | 4.03 ± 0.12 ** ## | 6.02 ± 0.73 ** ## |
| AFC-2 H | 19.20 ± 0.90 | 26.77 ± 1.56 * | 3.87 ± 0.29 ** | 5.60 ± 0.67 ** # |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Li, Y.; Shen, P.; Li, S.; Zhang, L.; Wang, Q.; Ren, D.; Liu, S.; Zhang, D.; Zhou, H. Anti-Hyperlipidemic Effect of Fucoidan Fractions Prepared from Iceland Brown Algae Ascophyllum nodosum in an Hyperlipidemic Mice Model. Mar. Drugs 2023, 21, 468. https://doi.org/10.3390/md21090468
He Y, Li Y, Shen P, Li S, Zhang L, Wang Q, Ren D, Liu S, Zhang D, Zhou H. Anti-Hyperlipidemic Effect of Fucoidan Fractions Prepared from Iceland Brown Algae Ascophyllum nodosum in an Hyperlipidemic Mice Model. Marine Drugs. 2023; 21(9):468. https://doi.org/10.3390/md21090468
Chicago/Turabian StyleHe, Yunhai, Yutong Li, Peili Shen, Shangkun Li, Linsong Zhang, Qiukuan Wang, Dandan Ren, Shu Liu, Demeng Zhang, and Hui Zhou. 2023. "Anti-Hyperlipidemic Effect of Fucoidan Fractions Prepared from Iceland Brown Algae Ascophyllum nodosum in an Hyperlipidemic Mice Model" Marine Drugs 21, no. 9: 468. https://doi.org/10.3390/md21090468