
Novel Natural Compounds and Their Anatomical Distribution in the Stinging Fireworm Hermodice carunculata (Annelida)
 , ,
, ,
Abstract
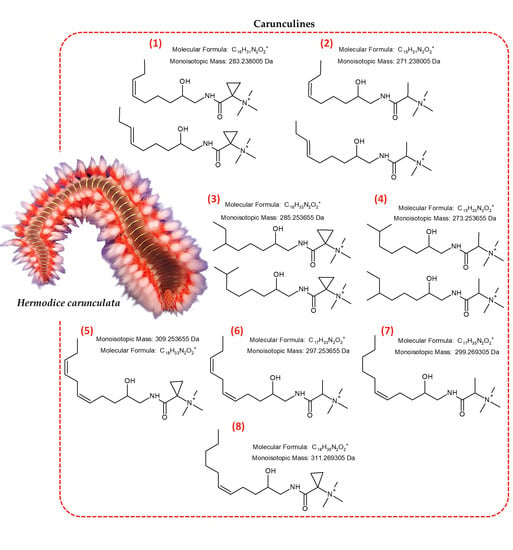
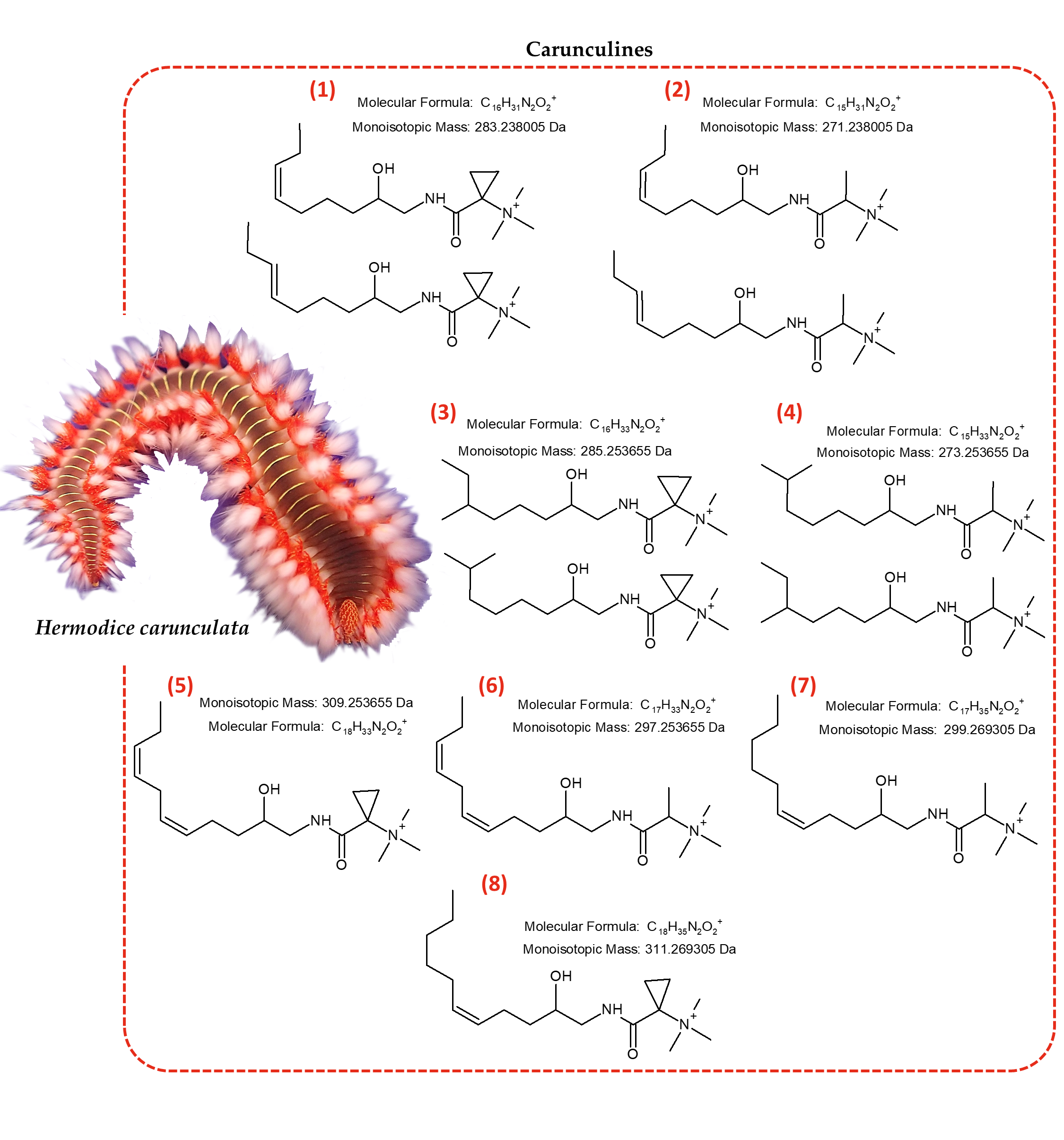
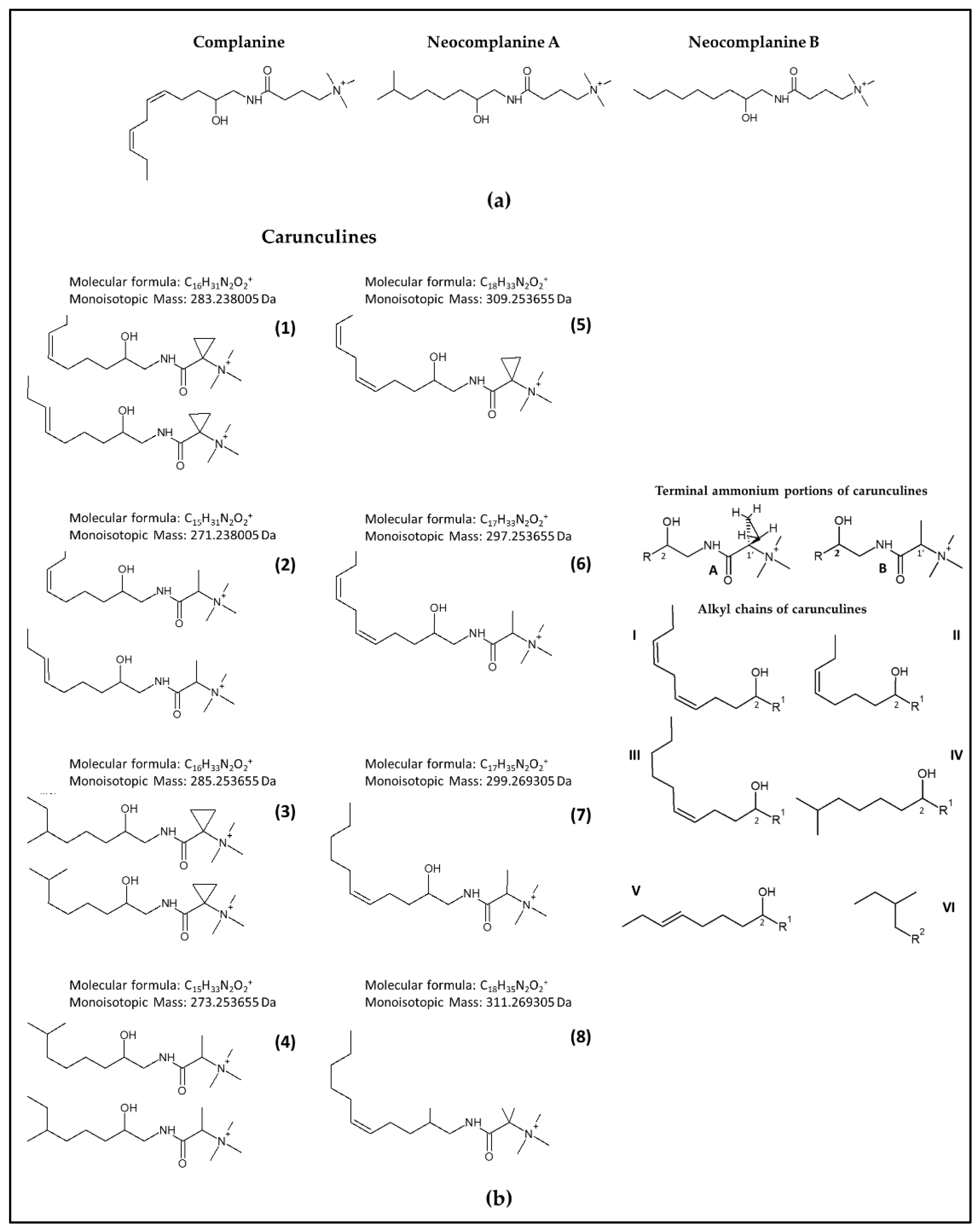
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
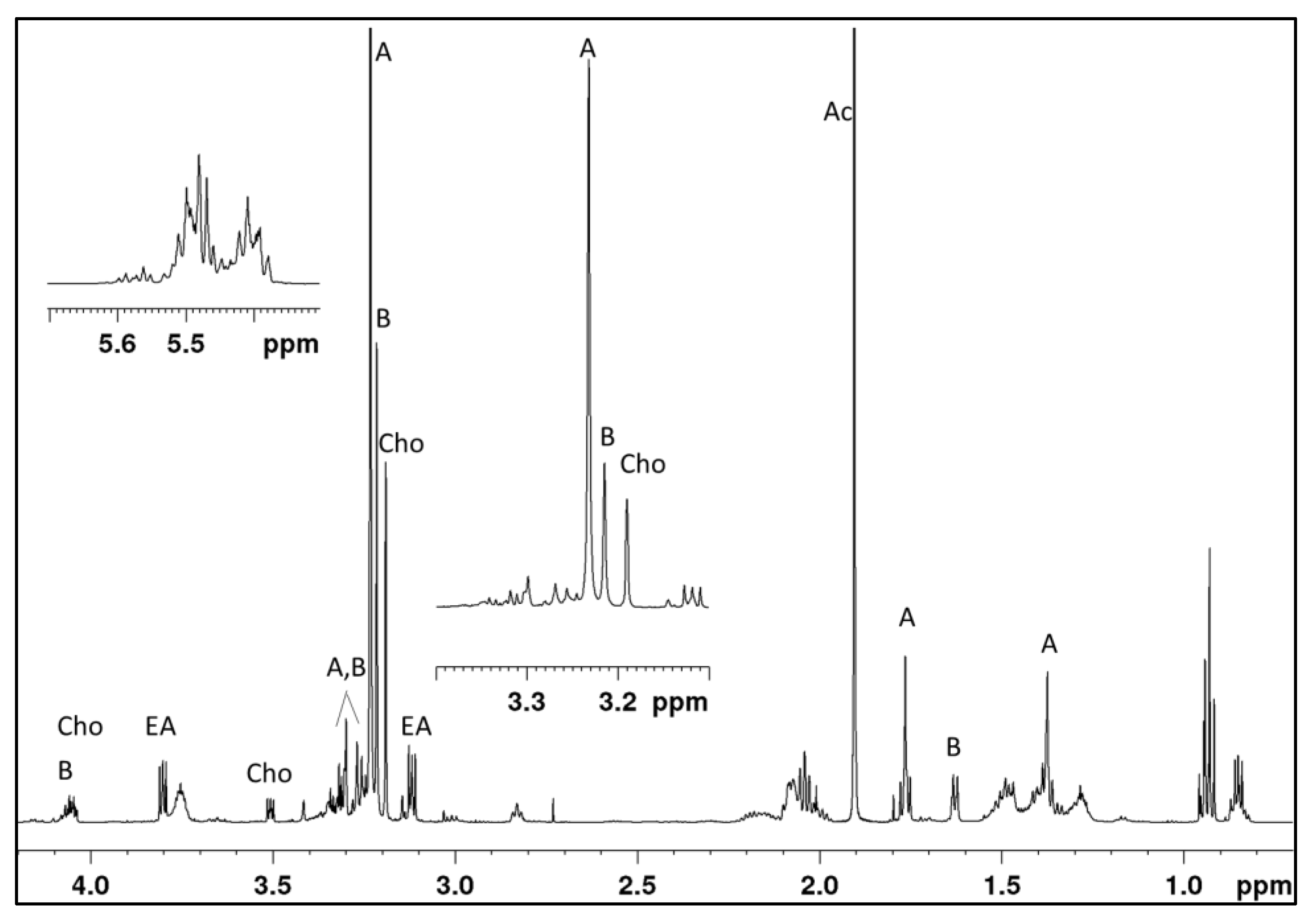
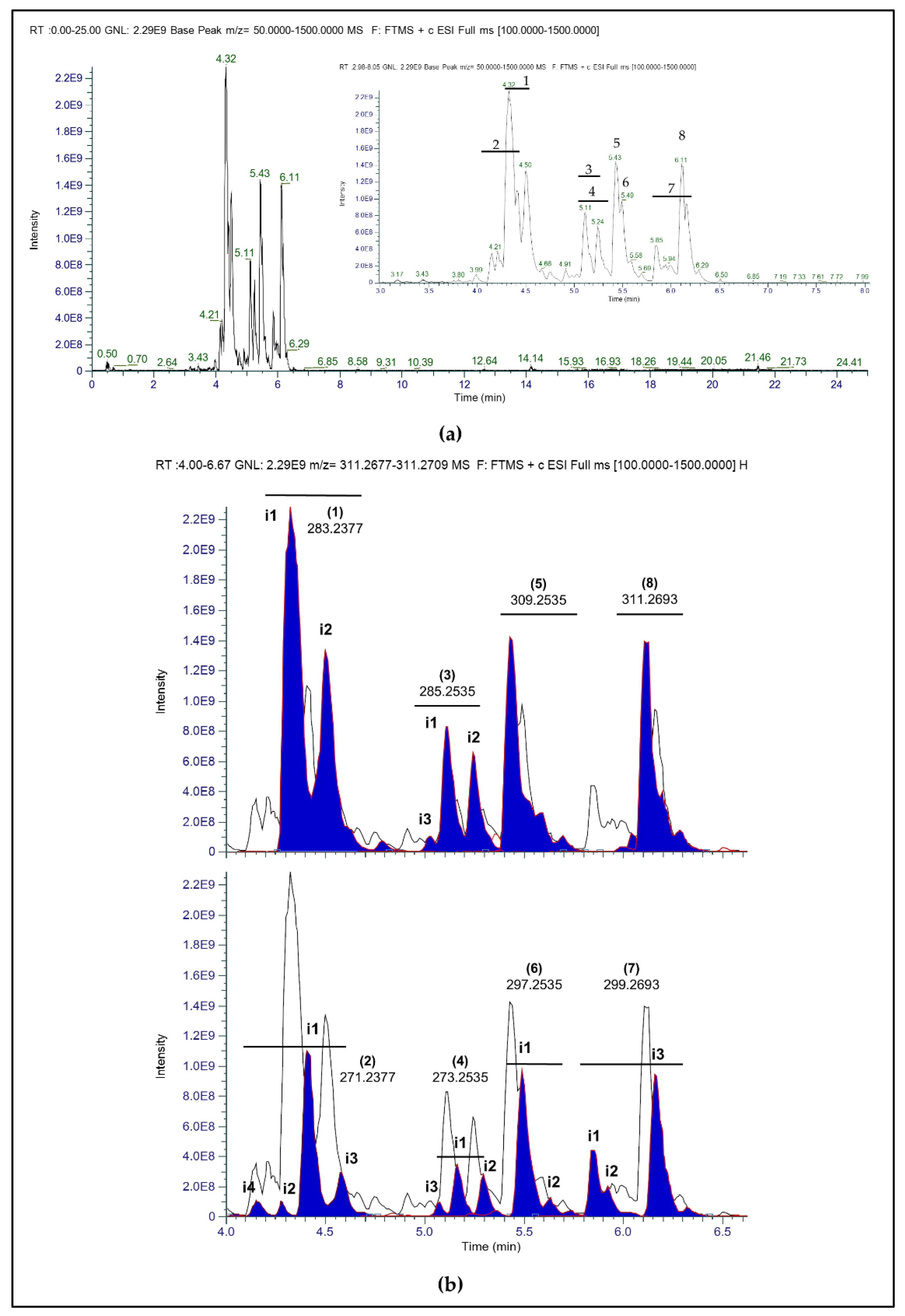
2.1. Identification of Novel Compounds by Mass Spectrometry and NMR Spectroscopy
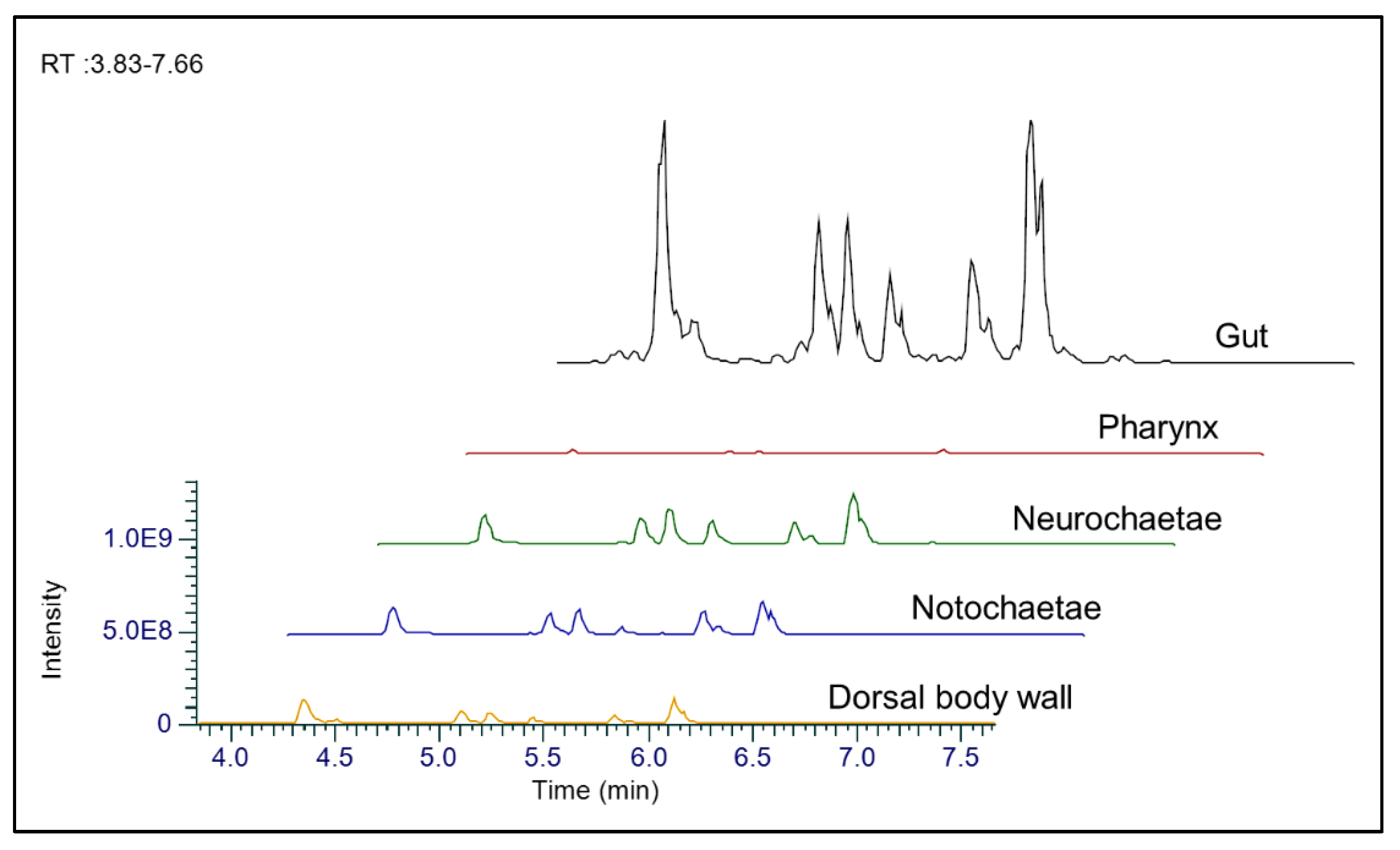
2.2. Anatomical Distribution of Carunculines in H. carunculata
2.3. Preliminary Screening for the Occurrence of Carunculines in Other Marine Invertebrates
3. Discussion
4. Materials and Methods
4.1. Animal Collection
4.2. Isolation of Natural Compounds from H. carunculata
4.3. General Description of Analytical Techniques
4.4. Anatomical Distribution of Fireworm Toxins
4.5. Occurrence of Carunculines in Other Marine Invertebrate Taxa
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Costantino, V.; Fattorusso, E.; Menna, M.; Taglialatela-Scafati, O. Chemical Diversity of Bioactive Marine Natural Products: An Illustrative Case Study. Curr. Med. Chem. 2004, 11, 1671–1692. [Google Scholar] [CrossRef]
- Firn, R.D.; Jones, C.G. Natural products? A simple model to explain chemical diversity. Nat. Prod. Rep. 2003, 20, 382. [Google Scholar] [CrossRef] [PubMed]
- Pawlik, J.R. Antipredatory defensive roles of natural products from marine invertebrates. In Handbook of Marine Natural Products; Fattorusso, E., Gerwick, W.H., Taglialatela-Scafati, O., Eds.; Springer: Dordrecht, the Netherlands, 2012; pp. 677–710. [Google Scholar] [CrossRef]
- Puglisi, M.P.; Sneed, J.M.; Ritson-Williams, R.; Young, R. Marine chemical ecology in benthic environments. Nat. Prod. Rep. 2019, 36, 410–429. [Google Scholar] [CrossRef] [PubMed]
- Rocha, J.; Peixe, L.; Gomes, N.C.M.; Calado, R. Cnidarians as a source of new marine bioactive compounds—An overview of the last decade and future steps for bioprospecting. Mar. Drugs 2011, 9, 1860–1886. [Google Scholar] [CrossRef]
- Kumar, M.S.; Pandita, N.S.; Pal, A.K. LC-MS/MS as a tool for identification of bioactive compounds in marine sponge Spongosorites halichondriodes (Dendy 1905). Toxicon 2012, 60, 1135–1147. [Google Scholar] [CrossRef]
- Cheney, K.L.; White, A.; Mudianta, I.W.; Winters, A.E.; Quezada, M.; Capon, R.J.; Mollo, E.; Garson, M.J. Choose your weaponry: Selective storage of a single toxic compound, Latrunculin A, by closely related nudibranch molluscs. PLoS ONE 2016, 11, e0145134. [Google Scholar] [CrossRef]
- Dìaz-Castaeda, V.; Reish, D.J. Polychaetes in environmental studies. In Annelids in Modern Biology; Shain, D.H., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009; pp. 203–227. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef]
- Coutinho, M.C.L.; Teixeira, V.L.; Santos, C.S.G. A review of “Polychaeta” chemicals and their possible ecological role. J. Chem. Ecol. 2018, 44, 72–94. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.; Nieto, R.M.; Blanco, M.; Valeriote, F.A.; Jiménez, C.; Crews, P. Thelepamide: An unprecedented ketide-amino acid from Thelepus crispus, a marine annelid worm. Org. Lett. 2014, 16, 464–467. [Google Scholar] [CrossRef]
- Calabro, K.; Jennings, L.K.; Lasserre, P.; Doohan, R.; Rodrigues, D.; Reyes, F.; Ramos, C.; Thomas, O.P. Nebulosins: Trisub-stituted thiolane natural products from the Northeastern Atlantic annelid Eupolymnia nebulosa. J. Org. Chem. 2020, 85, 14026–14041. [Google Scholar] [CrossRef]
- Kicklighter, C.; Kubanek, J.; Barsby, T.; Hay, M. Palatability and defense of some tropical infaunal worms: Alkylpyrrole sulfamates as deterrents to fish feeding. Mar. Ecol. Prog. Ser. 2003, 263, 299–306. [Google Scholar] [CrossRef]
- Kicklighter, C.E.; Kubanek, J.; Hay, M.E. Do brominated natural products defend marine worms from consumers? Some do, most don’t. Limnol. Oceanogr. 2004, 49, 430–441. [Google Scholar] [CrossRef]
- Von Reumont, B.M.; Campbell, L.I.; Richter, S.; Hering, L.; Sykes, D.; Hetmank, J.; Jenner, R.A.; Bleidorn, C. A Polychaete’s powerful punch: Venom gland transcriptomics of Glycera reveals a complex cocktail of toxin homologs. GBE 2014, 6, 2406–2423. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, M.; Ramos, Í.; Martins, C.; Costa, P.M. An investigation into the toxicity of tissue extracts from two distinct marine polychaeta. Toxicon X 2022, 14, 100116. [Google Scholar] [CrossRef]
- Simonini, R.; Maletti, I.; Righi, S.; Fai, S.; Prevedelli, D. Laboratory observations on predator–prey interactions between the bearded fireworm (Hermodice carunculata) and Mediterranean benthic invertebrates. Mar. Freshw. Behav. Physiol. 2018, 51, 145–158. [Google Scholar] [CrossRef]
- Simonini, R.; Maggioni, F.; Zanetti, F.; Fai, S.; Forti, L.; Prevedelli, D.; Righi, S. Synergy between mechanical injury and toxins triggers the urticating system of marine fireworms. J. Exp. Mar. Biol. Ecol. 2021, 534, 151487. [Google Scholar] [CrossRef]
- Righi, S.; Savioli, M.; Prevedelli, D.; Simonini, R.; Malferrari, D. Unravelling the ultrastructure and mineralogical compo-sition of fireworm stinging bristles. Zoology 2021, 144, 125851. [Google Scholar] [CrossRef]
- Nakamura, K.; Tachikawa, Y.; Kitamura, M.; Ohno, O.; Suganuma, M.; Uemura, D. Complanine, an inflammation-inducing substance isolated from the marine fireworm Eurythoe complanata. Org. Biomol. Chem. 2008, 6, 2058. [Google Scholar] [CrossRef]
- Nakamura, K.; Tachikawa, Y.; Ohno, O.; Kitamura, M.; Suganuma, M.; Uemura, D. Neocomplanines A and B, a complanine family isolated from the marine fireworm. J. Nat. Prod. 2010, 73, 303–305. [Google Scholar] [CrossRef]
- Borda, E.; Yáñez-Rivera, B.; Ochoa, G.M.; Kudenov, J.D.; Sanchez-Ortiz, C.; Schulze, A.; Rouse, G.W. Revamping Amphinomidae (Annelida: Amphinomida), with the inclusion of Notopygos. Zool. Scr. 2015, 44, 324–333. [Google Scholar] [CrossRef]
- Garrido, L.; Zubía, E.; Ortega, M.J.; Naranjo, S.; Salvá, J. Obscuraminols, new unsaturated amino alcohols from the tunicate pseudodistoma obscurum: Structure and absolute configuration. Tetrahedron 2001, 57, 4579–4588. [Google Scholar] [CrossRef]
- Ciavatta, M.L.; Manzo, E.; Nuzzo, G.; Villani, G.; Varcamonti, M.; Gavagnin, M. Crucigasterins A–E, antimicrobial amino alcohols from the mediterranean colonial ascidian Pseudodistoma crucigaster. Tetrahedron 2010, 66, 7533–7538. [Google Scholar] [CrossRef]
- Aiello, A.; Fattorusso, E.; Giordano, A.; Menna, M.; Navarrete, C.; Muñoz, E. Clavaminols A–F, novel cytotoxic 2-amino-3-alkanols from the ascidian Clavelina phlegraea. Bioorg. Med. Chem. 2007, 15, 2920–2926. [Google Scholar] [CrossRef] [PubMed]
- Aiello, A.; Fattorusso, E.; Giordano, A.; Menna, M.; Navarrete, C.; Muñoz, E. Clavaminols G–N, six new marine sphingoids from the mediterranean ascidian Clavelina phlegraea. Tetrahedron 2009, 65, 4384–4388. [Google Scholar] [CrossRef]
- Jiménez, C.; Crews, P. Novel marine sponge amino acids, 10. Xestoaminoh from Xestaspongia sp. J. Nat. Prod. 1990, 53, 978–982. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.J.; Garson, M.J.; Hooper, J.N.A. Antifungal alkyl amino alcohols from the tropical marine sponge Haliclona n. sp. J. Nat. Prod. 2001, 64, 1568–1571. [Google Scholar] [CrossRef]
- Cuadros, R.; Montejo de Garcini, E.; Wandosell, F.; Faircloth, G.; Fernández-Sousa, J.M.; Avila, J. The marine compound spisulosine, an inhibitor of cell proliferation, promotes the disassembly of actin stress fibers. Cancer Lett. 2000, 152, 23–29. [Google Scholar] [CrossRef]
- Dean, L.J.; Prinsep, M.R. The chemistry and chemical ecology of nudibranchs. Nat. Prod. Rep. 2017, 34, 1359–1390. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2017, 34, 235–294. [Google Scholar] [CrossRef]
- Verdes, A.; Simpson, D.; Holford, M. Are fireworms venomous? Evidence for the convergent evolution of toxin homologs in three species of fireworms (Annelida, Amphinomidae). GBE 2018, 10, 249–268. [Google Scholar] [CrossRef]
- Polat, A. The importance of betaine and some attractive substances as fish feed additives. In Feed Manufacturing in the Mediterranean Region: Recent Advances in Research and Technology; Brufau, J., Tacon, A., Eds.; CIHEAM: Zaragoza, Spain, 1999; pp. 217–220. [Google Scholar]
- Yancey, P.H.; Blake, W.R.; Conley, J. Unusual organic osmolytes in deep-sea animals: Adaptations to hydrostatic pressure and other perturbants. Comp. Biochem. Physiol. Mol. Amp. Integr. Physiol. 2002, 133, 667–676. [Google Scholar] [CrossRef]
- McNeil, S.D.; Nuccio, M.L.; Hanson, A.D. Betaines and related osmoprotectants. targets for metabolic engineering of stress resistance. Plant Physiol. 1999, 120, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Antonsen, S.G. Total synthesis as a tool for structural elucidation of some marine lipid natural products. Ph. D. Thesis, Norwegian University of Life Sciences, Ås, Norway, February 2017. [Google Scholar]
- Nakamura, K.; Tachikawa, Y.; Uemura, D. (−)-Complanine, an inflammatory substance of marine fireworm: A synthetic study. Beilstein J. Org. Chem. 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Righi, S. University of Modena and Reggio Emilia, Modena, Italy. Unpublished materials not intended for publication, 2021.
- Righi, S.; Savioli, M.; Prevedelli, D.; Simonini, R.; Malferrari, D. Response to Tilic and Bartolomaeus’s commentary on the original research paper “unravelling the ultrastructure and mineralogical composition of fireworm stinging bristles” (Zoology, 144). Zoology 2021, 144, 125889. [Google Scholar] [CrossRef] [PubMed]
- Carbone, M.; Gavagnin, M.; Haber, M.; Guo, Y.-W.; Fontana, A.; Manzo, E.; Genta-Jouve, G.; Tsoukatou, M.; Rudman, W.B.; Cimino, G.; et al. Packaging and delivery of chemical weapons: A defensive trojan horse stratagem in chromodorid nudibranchs. PLoS ONE 2013, 8, e62075. [Google Scholar] [CrossRef]
- Winters, A.E.; White, A.M.; Dewi, A.S.; Mudianta, I.W.; Wilson, N.G.; Forster, L.C.; Garson, M.J.; Cheney, K.L. Distribution of defensive metabolites in nudibranch molluscs. J. Chem. Ecol. 2018, 44, 384–396. [Google Scholar] [CrossRef]
- Stabili, L.; Schirosi, R.; Di Benedetto, A.; Merendino, A.; Villanova, L.; Giangrande, A. First insights into the biochemistry of Sabella spallanzanii (Annelida: Polychaeta) mucus: A potentially unexplored resource for applicative purposes. J. Mar. Biol. Ass. 2011, 91, 199–208. [Google Scholar] [CrossRef]
- Weigert, A.; Helm, C.; Meyer, M.; Nickel, B.; Arendt, D.; Hausdorf, B.; Santos, S.R.; Halanych, K.M.; Purschke, G.; Bleidorn, C.; et al. Illuminating the base of the annelid tree using transcriptomics. Mol. Biol. Evol. 2014, 31, 1391–1401. [Google Scholar] [CrossRef] [Green Version]
- Liebeke, M.; Bundy, J.G. Biochemical diversity of betaines in earthworms. Biochem. Biophys. Res. Commun. 2013, 430, 1306–1311. [Google Scholar] [CrossRef]
- Wiklund, H.; Nygren, A.; Pleijel, F.; Sundberg, P. The phylogenetic relationships between Amphinomidae, Archinomidae and Euphrosinidae (Amphinomida: Aciculata: Polychaeta), inferred from molecular data. J. Mar. Biol. Ass. 2008, 88, 509–513. [Google Scholar] [CrossRef]
- Borda, E.; Kudenov, J.D.; Bienhold, C.; Rouse, G.W. Towards a revised Amphinomidae (Annelida, Amphinomida): Description and affinities of a new genus and species from the nile deep-sea fan, mediterranean sea: Wood fall amphinomids and systematics. Zool. Scr. 2012, 41, 307–325. [Google Scholar] [CrossRef]
- Kicklighter, C.E.; Hay, M.E. Integrating prey defensive traits: Contrasts of marine worms from temperate and tropical habitats. Ecol. Monogr. 2006, 76, 195–215. [Google Scholar] [CrossRef]
- Rachamim, T.; Morgenstern, D.; Aharonovich, D.; Brekhman, V.; Lotan, T.; Sher, D. The dynamically evolving nematocyst content of an anthozoan, a scyphozoan, and a hydrozoan. Mol. Biol. Evol. 2015, 32, 740–753. [Google Scholar] [CrossRef]
- Jouiaei, M.; Yanagihara, A.; Madio, B.; Nevalainen, T.; Alewood, P.; Fry, B. Ancient venom systems: A review on Cnidaria toxins. Toxins 2015, 7, 2251–2271. [Google Scholar] [CrossRef] [PubMed]
- D’Ambra, I.; Lauritano, C. A review of toxins from Cnidaria. Mar. Drugs 2020, 18, 507. [Google Scholar] [CrossRef]
- Madio, B.; King, G.F.; Undheim, E.A.B. Sea anemone toxins: A structural overview. Mar. Drugs 2019, 17, 325. [Google Scholar] [CrossRef]
- Simonini, R.; Righi, S.; Maletti, I.; Fai, S.; Prevedelli, D. Bearded versus thorny: The fireworm Hermodice carunculata preys on the sea urchin Paracentrotus lividus. Ecology 2017, 98, 2730–2732. [Google Scholar] [CrossRef]
- Simonini, R.; Righi, S.; Zanetti, F.; Fai, S.; Prevedelli, D. Development and catch efficiency of an attracting device to collect and monitor the invasive fireworm Hermodice carunculata in the Mediterranean Sea. Medit. Mar. Sci. 2021, 22, 706–714. [Google Scholar] [CrossRef]
- Righi, S.; Prevedelli, D.; Simonini, R. Ecology, distribution and expansion of a Mediterranean native invader, the fireworm Hermodice carunculata (Annelida). Medit. Mar. Sci. 2020, 21, 558–574. [Google Scholar] [CrossRef]
- Chemspider. Available online: http://www.chemspider.com/Chemical-Structure.25047857.html (accessed on 2 February 2020).
- Crutchfield, C.A.; Clarke, W. High resolution accurate mass (HRAM) mass spectrometry. In Mass Spectrometry for the Clinical Laboratory; Elsevier: Amsterdam, the Netherlands, 2017; pp. 247–259. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Righi, S.; Forti, L.; Simonini, R.; Ferrari, V.; Prevedelli, D.; Mucci, A. Novel Natural Compounds and Their Anatomical Distribution in the Stinging Fireworm Hermodice carunculata (Annelida). Mar. Drugs 2022, 20, 585. https://doi.org/10.3390/md20090585
Righi S, Forti L, Simonini R, Ferrari V, Prevedelli D, Mucci A. Novel Natural Compounds and Their Anatomical Distribution in the Stinging Fireworm Hermodice carunculata (Annelida). Marine Drugs. 2022; 20(9):585. https://doi.org/10.3390/md20090585
Chicago/Turabian StyleRighi, Sara, Luca Forti, Roberto Simonini, Valentina Ferrari, Daniela Prevedelli, and Adele Mucci. 2022. "Novel Natural Compounds and Their Anatomical Distribution in the Stinging Fireworm Hermodice carunculata (Annelida)" Marine Drugs 20, no. 9: 585. https://doi.org/10.3390/md20090585