
Role of Tumor-Associated Macrophages in Cervical Cancer: Integrating Classical Perspectives with Recent Technological Advances
 ,
, {kind=link}
Abstract
:1. Introduction
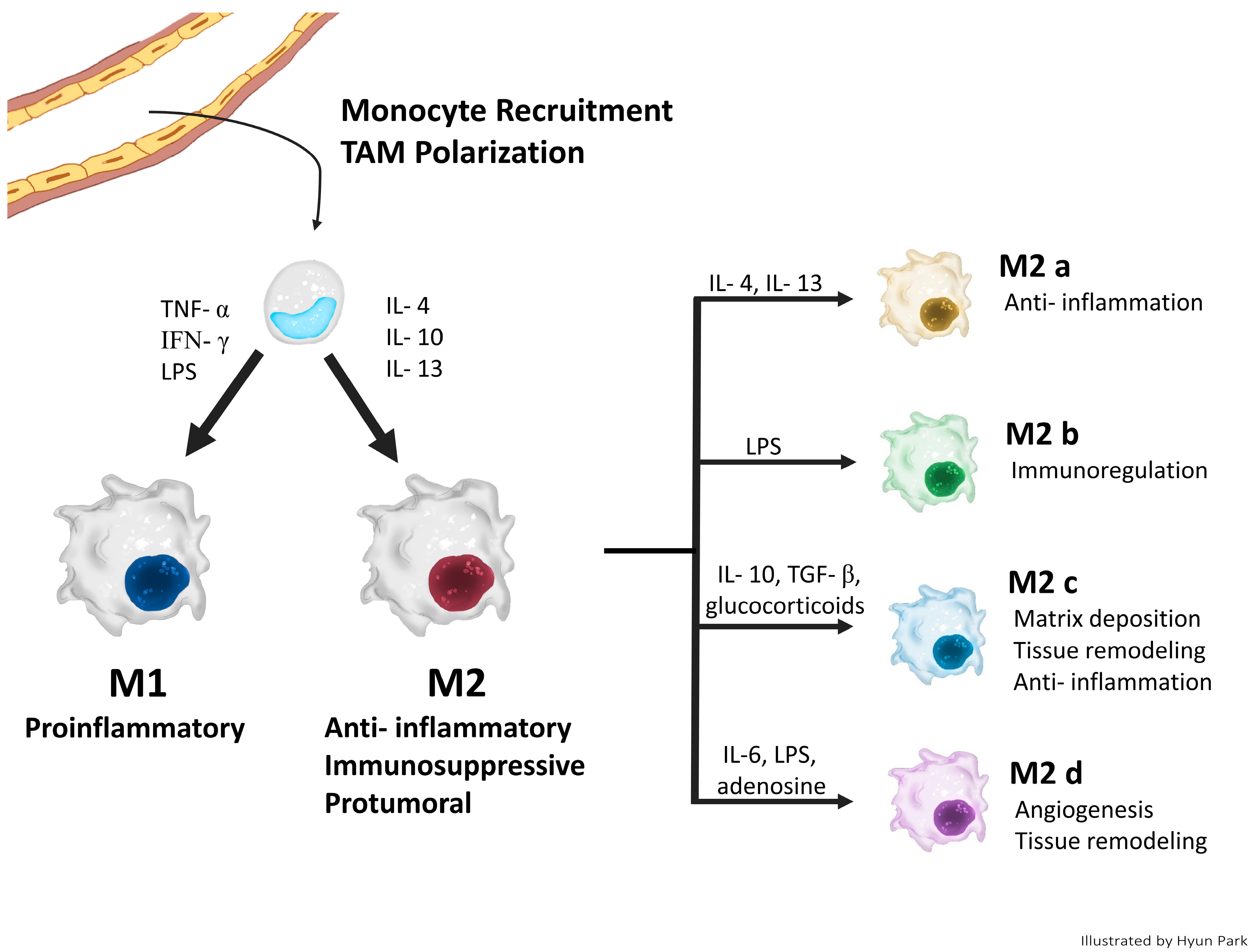
2. Origin and Polarization of TAMs
2.1. Origin of TAMs
2.2. Classical Concept of TAM Polarization
3. The Role of TAMs in CC Progression
3.1. Inflammation/Initiation
3.2. Angiogenesis
3.3. Invasion/Migration/Metastasis
3.4. Immunosuppression
3.5. Cancer Stem Cells
3.6. Prognosis
4. TAM-Targeting Therapy
4.1. Depletion of TAMs
4.2. Inhibiting Monocyte/Macrophage Recruitment
4.3. Reprogramming/Re-Education of TAMs into M1-like Phenotype
4.4. Other Methods
5. Novel Findings of TAMs in the TME with Recent Technologies: Multi-Omics, SC RNA-Seq, and SP RNA-Seq
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sica, A.; Saccani, A.; Mantovani, A. Tumor-Associated Macrophages: A Molecular Perspective. Int. Immunopharmacol. 2002, 2, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Bae, J.S. Tumor-Associated Macrophages and Neutrophils in Tumor Microenvironment. Mediat. Inflamm. 2016, 2016, 6058147. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Steger, A.; Mahner, S.; Jeschke, U.; Heidegger, H. The Formation and Therapeutic Update of Tumor-Associated Macrophages in Cervical Cancer. Int. J. Mol. Sci. 2019, 20, 3310. [Google Scholar] [CrossRef] [PubMed]
- Petty, A.J.; Yang, Y. Tumor-Associated Macrophages: Implications in Cancer Immunotherapy. Immunotherapy 2017, 9, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Sioud, M. Tumor-Associated Macrophage Subsets: Shaping Polarization and Targeting. Int. J. Mol. Sci. 2023, 24, 7493. [Google Scholar] [CrossRef]
- Mantovani, A.; Schioppa, T.; Porta, C.; Allavena, P.; Sica, A. Role of Tumor-Associated Macrophages in Tumor Progression and Invasion. Cancer Metastasis Rev. 2006, 25, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Locati, M. Tumor-Associated Macrophages as a Paradigm of Macrophage Plasticity, Diversity, and Polarization Lessons and Open Questions. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Chanmee, T.; Ontong, P.; Konno, K.; Itano, N. Tumor-Associated Macrophages as Major Players in the Tumor Microenvironment. Cancers 2014, 6, 1670–1690. [Google Scholar] [CrossRef]
- Krishnan, V.; Schaar, B.; Tallapragada, S.; Dorigo, O. Tumor Associated Macrophages in Gynecologic Cancers. Gynecol. Oncol. 2018, 149, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Zeng, F.; Liao, S.; Cao, L.; Zhou, Y. Effects of Glycolysis on the Polarization and Function of Tumor-Associated Macrophages (Review). Int. J. Oncol. 2023, 62, 70. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K.; Allavena, P.; Mantovani, A. Tumor-Associated Macrophages: Functional Diversity, Clinical Significance, and Open Questions. Semin. Immunopathol. 2013, 35, 585–600. [Google Scholar] [CrossRef] [PubMed]
- Hourani, T.; Holden, J.A.; Li, W.; Lenzo, J.C.; Hadjigol, S.; O’Brien-Simpson, N.M. Tumor Associated Macrophages: Origin, Recruitment, Phenotypic Diversity, and Targeting. Front. Oncol. 2021, 11, 788365. [Google Scholar] [CrossRef] [PubMed]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage Plasticity, Polarization, and Function in Health and Disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, G.; Maciejewski, T.M.; Szukiewicz, D. Current Progress in the Inflammatory Background of Angiogenesis in Gynecological Cancers. Inflamm. Res. 2019, 68, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Yu, S.; Zhang, J.; Wu, S. Dysregulated Tumor-Associated Macrophages in Carcinogenesis, Progression and Targeted Therapy of Gynecological and Breast Cancers. J. Hematol. Oncol. 2021, 14, 181. [Google Scholar] [CrossRef]
- Tan, J.; Yang, L.; Zhao, H.; Ai, Y.; Ren, L.; Zhang, F.; Dong, W.; Shi, R.; Sun, D.; Feng, Y. The Role of NFATc1/c-Myc/PKM2/IL-10 Axis in Activating Cervical Cancer Tumor-Associated M2 Macrophage Polarization to Promote Cervical Cancer Progression. Exp. Cell Res. 2022, 413, 113052. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Reyes, K.; Bravo-Cuellar, A.; Hernández-Flores, G.; Lerma-Díaz, J.M.; Jave-Suárez, L.F.; Gómez-Lomelí, P.; De Celis, R.; Aguilar-Lemarroy, A.; Domínguez-Rodríguez, J.R.; Ortiz-Lazareno, P.C. Cervical Cancer Cell Supernatants Induce a Phenotypic Switch from U937-Derived Macrophage-Activated M1 State into M2-like Suppressor Phenotype with Change in Toll-like Receptor Profile. Biomed Res. Int. 2014, 2014, 683068. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, G.; Zhang, S. Associations between Intratumoral and Peritumoral M2 Macrophage Counts and Cervical Squamous Cell Carcinoma Invasion Patterns. Int. J. Gynecol. Obstet. 2017, 139, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, L.; Li, Y.; Zhao, X. Research Progress on Tumor-Associated Macrophages and Inflammation in Cervical Cancer. Biomed Res. Int. 2020, 2020, 6842963. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Song, Y.; Du, W.; Gong, L.; Chang, H.; Zou, Z. Tumor-Associated Macrophages: An Accomplice in Solid Tumor Progression. J. Biomed. Sci. 2019, 26, 78. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Yi, M.; Wu, Y.; Dong, B.; Wu, K. Roles of Tumor-Associated Macrophages in Tumor Progression: Implications on Therapeutic Strategies. Exp. Hematol. Oncol. 2021, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Allavena, P.; Sica, A.; Solinas, G.; Porta, C.; Mantovani, A. The Inflammatory Micro-Environment in Tumor Progression: The Role of Tumor-Associated Macrophages. Crit. Rev. Oncol. Hematol. 2008, 66, 1–9. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Malekghasemi, S.; Majidi, J.; Baghbanzadeh, A.; Abdolalizadeh, J.; Baradaran, B.; Aghebati-Maleki, L. Tumor-Associated Macrophages: Protumoral Macrophages in Inflammatory Tumor Microenvironment. Adv. Pharm. Bull. 2020, 10, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Utrera-Barillas, D.; Castro-Manrreza, M.; Castellanos, E.; Gutiérrez-Rodríguez, M.; Arciniega-Ruíz de Esparza, O.; García-Cebada, J.; Velazquez, J.R.; Flores-Reséndiz, D.; Hernández-Hernández, D.; Benítez-Bribiesca, L. The Role of Macrophages and Mast Cells in Lymphangiogenesis and Angiogenesis in Cervical Carcinogenesis. Exp. Mol. Pathol. 2010, 89, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Steinberger, K.J.; Eubank, T.D. The Underexplored Landscape of Hypoxia-Inducible Factor 2 Alpha and Potential Roles in Tumor Macrophages: A Review. Oxygen 2023, 3, 45–76. [Google Scholar] [CrossRef] [PubMed]
- Henze, A.; Mazzone, M.; Henze, A.; Mazzone, M. The Impact of Hypoxia on Tumor-Associated Macrophages Find the Latest Version: The Impact of Hypoxia on Tumor-Associated Macrophages. J. Clin. Investig. 2016, 126, 3672–3679. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.; Li, Y.; Jian, L.; Yang, Y.; Zhao, L.; Wei, M. The Hypoxia-Driven Crosstalk between Tumor and Tumor-Associated Macrophages: Mechanisms and Clinical Treatment Strategies. Mol. Cancer 2022, 21, 177. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Yu, Y.; Wang, X.; Zhang, T. Tumor-Associated Macrophages in Tumor Immunity. Front. Immunol. 2020, 11, 583084. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Zhang, S. Tumor-Associated Macrophages and Their Functional Transformation in the Hypoxic Tumor Microenvironment. Front. Immunol. 2021, 12, 741305. [Google Scholar] [CrossRef] [PubMed]
- Baradaran, A.; Asadzadeh, Z.; Hemmat, N.; Baghbanzadeh, A.; Shadbad, M.A.; Khosravi, N.; Derakhshani, A.; Alemohammad, H.; Afrashteh Nour, M.; Safarpour, H.; et al. The Cross-Talk between Tumor-Associated Macrophages and Tumor Endothelium: Recent Advances in Macrophage-Based Cancer Immunotherapy. Biomed. Pharmacother. 2022, 146, 112588. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Wu, S.; Yan, R.M.; Fan, L.S.; Yu, L.; Zhang, Y.M.; Wei, W.F.; Zhou, C.F.; Wu, X.G.; Zhong, M.; et al. The Role of the Hypoxia-Nrp-1 Axis in the Activation of M2-like Tumor-Associated Macrophages in the Tumor Microenvironment of Cervical Cancer. Mol. Carcinog. 2019, 58, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Mazibrada, J.; Rittà, M.; Mondini, M.; De Andrea, M.; Azzimonti, B.; Borgogna, C.; Ciotti, M.; Orlando, A.; Surico, N.; Chiusa, L.; et al. Interaction between Inflammation and Angiogenesis during Different Stages of Cervical Carcinogenesis. Gynecol. Oncol. 2008, 108, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Schoppmann, S.F.; Birner, P.; Stöckl, J.; Kalt, R.; Ullrich, R.; Caucig, C.; Nagy, K.; Alitalo, K.; Kerjaschki, D. Tumor-Associated Macrophages Express Lymphatic Endothelial Growth Factors and Are Related to Peritumoral Lymphangiogenesis. Am. J. Pathol. 2002, 161, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Cai, J.; Mao, M.; Fang, Y.; Huang, Z.; Jia, J.; Li, T.; Xu, L.; Wang, J.; Zhou, J.; et al. Tumor-Associated Macrophages Induce Lymphangiogenesis in Cervical Cancer via Interaction with Tumor Cells. Apmis 2014, 122, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Wei, W.F.; Wang, Z.C.; Wang, N.; Guo, C.H.; Zhou, C.F.; Liang, L.J.; Wu, S.; Liang, L.; Wang, W. A Novel Lymphatic Pattern Promotes Metastasis of Cervical Cancer in a Hypoxic Tumour-Associated Macrophage-Dependent Manner. Angiogenesis 2021, 24, 549–565. [Google Scholar] [CrossRef] [PubMed]
- Aras, S.; Raza Zaidi, M. TAMeless Traitors: Macrophages in Cancer Progression and Metastasis. Br. J. Cancer 2017, 117, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.Z.; Chan, M.K.K.; Chan, A.S.W.; Leung, K.T.; Jiang, X.; To, K.F.; Wu, Y.; Tang, P.M.K. Tumour-Associated Macrophages: Versatile Players in the Tumour Microenvironment. Front. Cell Dev. Biol. 2023, 11, 1261749. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, L.; Peng, X.; Zhan, X. Progress of Tumor-Associated Macrophages in the Epithelial-Mesenchymal Transition of Tumor. Front. Oncol. 2022, 12, 911410. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Ye, M.; Zhang, W. E6/E7 Oncoproteins of High Risk HPV-16 Upregulate MT1-MMP, MMP-2 and MMP-9 and Promote the Migration of Cervical Cancer Cells. Int. J. Clin. Exp. Pathol. 2015, 8, 4981–4989. [Google Scholar] [PubMed]
- Hu, W.; Li, X.; Zhang, C.; Yang, Y.; Jiang, J.; Wu, C. Tumor-Associated Macrophages in Cancers. Clin. Transl. Oncol. 2016, 18, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Sun, P.L.; He, Y.; Yao, M.; Gao, H. Immune Stromal Features in Cervical Squamous Cell Carcinoma Are Prognostic Factors for Distant Metastasis: A Retrospective Study. Pathol. Res. Pract. 2020, 216, 152751. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Yang, Y.; Fang, M.; Li, X.; Yuan, X.; Yuan, J. Co-Evolution of Tumor-Associated Macrophages and Tumor Neo-Vessels during Cervical Cancer Invasion. Oncol. Lett. 2016, 12, 2625–2631. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Deng, Y.R.; Wang, Z.C.; Wei, W.F.; Zhou, C.F.; Zhang, Y.M.; Yan, R.M.; Liang, L.J.; Zhong, M.; Liang, L.; et al. Hypoxia-Induced ZEB1 Promotes Cervical Cancer Progression via CCL8-Dependent Tumour-Associated Macrophage Recruitment. Cell Death Dis. 2019, 10, 508. [Google Scholar] [CrossRef] [PubMed]
- Ruffell, B.; Affara, N.I.; Coussens, L.M. Differential Macrophage Programming in the Tumor Microenvironment Macrophages in Solid Malignancies. Trends Immunol. 2012, 33, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Takeya, M.; Komohara, Y. Role of Tumor-Associated Macrophages in Human Malignancies: Friend or Foe? Pathol. Int. 2016, 66, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Petty, A.J.; Li, A.; Wang, X.; Dai, R.; Heyman, B.; Hsu, D.; Huang, X.; Yang, Y. Hedgehog Signaling Promotes Tumor-Associated Macrophage Polarization to Suppress Intratumoral CD8+ T Cell Recruitment. J. Clin. Investig. 2019, 129, 5151–5162. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Feng, Y.C.; Zhao, G.; Zhang, R.; Cheng, Z.Z.; Kong, W.N.; Wu, H.L.; Xu, B.; Lv, X.; Ma, X.M. Tumor-Associated CD163+ M2 Macrophage Infiltration Is Highly Associated with PD-L1 Expression in Cervical Cancer. Cancer Manag. Res. 2020, 12, 5831–5843. [Google Scholar] [CrossRef] [PubMed]
- Ring, K.L.; Yemelyanova, A.V.; Soliman, P.T.; Frumovitz, M.M.; Jazaeri, A.A. Potential Immunotherapy Targets in Recurrent Cervical Cancer. Gynecol. Oncol. 2017, 145, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Morales, V.A.; Chávez-Sánchez, L.; Rocha-Zavaleta, L.; Espíndola-Garibay, S.; Monroy-García, A.; Castro-Manrreza, M.E.; Fajardo-Orduña, G.R.; Apresa-García, T.; Gutiérrez-de la Barrera, M.; Mayani, H.; et al. Mesenchymal Stem/Stromal Cells Derived from Cervical Cancer Promote M2 Macrophage Polarization. Cells 2023, 12, 1047. [Google Scholar] [CrossRef] [PubMed]
- Heeren, A.M.; Punt, S.; Bleeker, M.C.; Gaarenstroom, K.N.; Van Der Velden, J.; Kenter, G.G.; De Gruijl, T.D.; Jordanova, E.S. Prognostic Effect of Different PD-L1 Expression Patterns in Squamous Cell Carcinoma and Adenocarcinoma of the Cervix. Mod. Pathol. 2016, 29, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Yang, G.; Ye, P.; Cao, N.; Chi, X.; Yang, W.H.; Yan, X. Macrophages Are a Double-Edged Sword: Molecular Crosstalk between Tumor-Associated Macrophages and Cancer Stem Cells. Biomolecules 2022, 12, 850. [Google Scholar] [CrossRef] [PubMed]
- Jinushi, M.; Baghdadi, M.; Chiba, S.; Yoshiyama, H. Regulation of Cancer Stem Cell Activities by Tumor-Associated Macrophages. Am. J. Cancer Res. 2012, 2, 529–539. [Google Scholar] [PubMed]
- Chen, Y.; Tan, W.; Wang, C. Tumor-Associated Macrophage-Derived Cytokines Enhance Cancer Stem-like Characteristics through Epithelial–Mesenchymal Transition. Onco Targets Ther. 2018, 11, 3817–3826. [Google Scholar] [CrossRef] [PubMed]
- Sainz, B.; Carron, E.; Vallespinós, M.; Machado, H.L. Cancer Stem Cells and Macrophages: Implications in Tumor Biology and Therapeutic Strategies. Mediat. Inflamm. 2016, 2016, 9012369. [Google Scholar] [CrossRef] [PubMed]
- Radharani, N.N.V.; Yadav, A.S.; Nimma, R.; Kumar, T.V.S.; Bulbule, A.; Chanukuppa, V.; Kumar, D.; Patnaik, S.; Rapole, S.; Kundu, G.C. Tumor-Associated Macrophage Derived IL-6 Enriches Cancer Stem Cell Population and Promotes Breast Tumor Progression via Stat-3 Pathway. Cancer Cell Int. 2022, 22, 122. [Google Scholar] [CrossRef] [PubMed]
- Raonic, J.; Lopicic, M.; Vuckovic, L.; Vucinic, J. Immunohistochemical Analysis of CD68, CD4, CD8 and CD20 Expression in Cervical Dysplasia and Its Relationship with HR-HPV Infection. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 7598–7606. [Google Scholar] [CrossRef]
- Davidson, B.; Goldberg, I.; Kopolovic, J. Inflammatory Response in Cervical Intraepithelial Neoplasia and Squamous Cell Carcinoma of the Uterine Cervix. Pathol. Res. Pract. 1997, 193, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Fridman, W.H.; Zitvogel, L.; Sautès-Fridman, C.; Kroemer, G. The Immune Contexture in Cancer Prognosis and Treatment. Nat. Rev. Clin. Oncol. 2017, 14, 717–734. [Google Scholar] [CrossRef] [PubMed]
- Horta, B.; Pereira, T.; Medeiros, R.; Cerqueira, F. Cervical Cancer Outcome and Tumor-Associated Macrophages: Research Evidence. Immuno 2022, 2, 460–468. [Google Scholar] [CrossRef]
- Hammes, L.S.; Tekmal, R.R.; Naud, P.; Edelweiss, M.I.; Kirma, N.; Valente, P.T.; Syrjänen, K.J.; Cunha-Filho, J.S. Macrophages, Inflammation and Risk of Cervical Intraepithelial Neoplasia (CIN) Progression-Clinicopathological Correlation. Gynecol. Oncol. 2007, 105, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, Z.; Gao, A.; Wen, Q.; Sun, Y. The Prognostic Landscape of Tumor-Infiltrating Immune Cells in Cervical Cancer. Biomed. Pharmacother. 2019, 120, 109444. [Google Scholar] [CrossRef] [PubMed]
- Den Boon, J.A.; Pyeon, D.; Wang, S.S.; Horswill, M.; Schiffman, M.; Sherman, M.; Zuna, R.E.; Wang, Z.; Hewitt, S.M.; Pearson, R.; et al. Molecular Transitions from Papillomavirus Infection to Cervical Precancer and Cancer: Role of Stromal Estrogen Receptor Signaling. Proc. Natl. Acad. Sci. USA 2015, 112, E3255–E3264. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, M.; Zannoni, G.F.; Martinelli, E.; Anchora, L.P.; Ferrandina, G.; Tropeano, G.; Fagotti, A.; Scambia, G. Polarisation of Tumor-Associated Macrophages toward M2 Phenotype Correlates with Poor Response to Chemoradiation and Reduced Survival in Patients with Locally Advanced Cervical Cancer. PLoS ONE 2015, 10, e0136654. [Google Scholar] [CrossRef] [PubMed]
- Kawachi, A.; Yoshida, H.; Kitano, S.; Ino, Y.; Kato, T.; Hiraoka, N. Tumor-Associated CD204+ M2 Macrophages Are Unfavorable Prognostic Indicators in Uterine Cervical Adenocarcinoma. Cancer Sci. 2018, 109, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Carus, A.; Ladekarl, M.; Hager, H.; Nedergaard, B.S.; Donskov, F. Tumour-Associated CD66b + Neutrophil Count Is an Independent Prognostic Factor for Recurrence in Localised Cervical Cancer. Br. J. Cancer 2013, 108, 2116–2122. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Han, L.F.; Wu, X.G.; Wei, W.F.; Wu, L.F.; Yi, H.Y.; Yan, R.M.; Bai, X.Y.; Zhong, M.; Yu, Y.H.; et al. Clinical Significance of CD163+ and CD68+ Tumor-Associated Macrophages in High-Risk HPV-Related Cervical Cancer. J. Cancer 2017, 8, 3868–3875. [Google Scholar] [CrossRef]
- Guo, F.; Kong, W.; Zhao, G.; Cheng, Z.; Ai, L.; Lv, J.; Feng, Y.; Ma, X. The Correlation between Tumor-Associated Macrophage Infiltration and Progression in Cervical Carcinoma. Biosci. Rep. 2021, 41, BSR20203145. [Google Scholar] [CrossRef] [PubMed]
- Dimitrova, P.; Vasileva-Slaveva, M.; Shivarov, V.; Hasan, I.; Yordanov, A. Infiltration by Intratumor and Stromal CD8 and CD68 in Cervical Cancer. Medicina 2023, 59, 728. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.; Luo, Y. Targeting Macrophages in Cancer Immunotherapy. Signal Transduct. Target. Ther. 2021, 6, 127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Y.; Song, X.Y.; Li, Y.; Ye, L.L.; Zhou, Q.; Yang, W.B. Tumor-Associated Macrophages: A Promising Target for a Cancer Immunotherapeutic Strategy. Pharmacol. Res. 2020, 161, 105111. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, R.; Gao, Q. The Roles and Targeting of Tumor-Associated Macrophages. Front. Biosci. Landmark 2023, 28, 207. [Google Scholar] [CrossRef] [PubMed]
- Baci, D.; Bosi, A.; Gallazzi, M.; Rizzi, M.; Noonan, D.M.; Poggi, A.; Bruno, A.; Mortara, L. The Ovarian Cancer Tumor Immune Microenvironment (Time) as Target for Therapy: A Focus on Innate Immunity Cells as Therapeutic Effectors. Int. J. Mol. Sci. 2020, 21, 3125. [Google Scholar] [CrossRef] [PubMed]
- Truxova, I.; Cibula, D.; Spisek, R.; Fucikova, J. Targeting Tumor-Associated Macrophages for Successful Immunotherapy of Ovarian Carcinoma. J. Immunother. Cancer 2023, 11, e005968. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Yang, Q. Tumor-Associated Macrophage-Targeted Therapeutics in Ovarian Cancer. Int. J. Cancer 2021, 149, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yang, Y.; Ma, P.; Huang, H.; Tang, Q.; Miao, H.; Fang, Y.; Jiang, N.; Li, Y.; Zhu, Q.; et al. Landscape and Perspectives of Macrophage -Targeted Cancer Therapy in Clinical Trials. Mol. Ther. Oncolytics 2022, 24, 799–813. [Google Scholar] [CrossRef]
- Cao, Y.; Qiao, B.; Chen, Q.; Xie, Z.; Dou, X.; Xu, L.; Ran, H.; Zhang, L.; Wang, Z. Tumor Microenvironment Remodeling via Targeted Depletion of M2-like Tumor-Associated Macrophages for Cancer Immunotherapy. Acta Biomater. 2023, 160, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Jia, L.; Niu, Y.; Qi, H.; Chen, X.; Zhang, Q.; Zhang, J.; Wang, Y.; Dong, L.; Wang, C. Targeted Depletion of Tumour-Associated Macrophages by an Alendronate-Glucomannan Conjugate for Cancer Immunotherapy. Biomaterials 2014, 35, 10046–10057. [Google Scholar] [CrossRef]
- De Nola, R.; Menga, A.; Castegna, A.; Loizzi, V.; Ranieri, G.; Cicinelli, E.; Cormio, G. The Crowded Crosstalk between Cancer Cells and Stromal Microenvironment in Gynecological Malignancies: Biological Pathways and Therapeutic Implication. Int. J. Mol. Sci. 2019, 20, 2401. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Yin, H. Engeletin Suppresses Cervical Carcinogenesis in Vitro and in Vivo by Reducing NF-ΚB-Dependent Signaling. Biochem. Biophys. Res. Commun. 2020, 526, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Almahariq, M.F.; Quinn, T.J.; Kesarwani, P.; Kant, S.; Miller, C.R.; Chinnaiyan, P. Inhibition of Colony-Stimulating Factor-1 Receptor Enhances the Efficacy of Radiotherapy and Reduces Immune Suppression in Glioblastoma. In Vivo 2021, 35, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhang, Q.; Xu, M.; Wang, L.; Chen, X.; Feng, Y.; Li, Y.; Zhang, X.; Cui, W.; Jia, X. CCL2-CCR2 Axis Recruits Tumor Associated Macrophages to Induce Immune Evasion through PD-1 Signaling in Esophageal Carcinogenesis. Mol. Cancer 2020, 19, 41. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Luo, Z.; Li, X.; Han, X.; Shi, S.; Zhang, T. Tumor-Associated Macrophages: Role in Tumorigenesis and Immunotherapy Implications. J. Cancer 2021, 12, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Zou, D.; Guo, M.; Zhou, Q. A Clinical Study of Pegylated Recombinant Human Granulocyte Colony Stimulating Factor (PEG-RhG-CSF) in Preventing Neutropenia during Concurrent Chemoradiotherapy of Cervical Cancer. BMC Cancer 2021, 21, 661. [Google Scholar] [CrossRef] [PubMed]
- Wanderley, C.W.; Colón, D.F.; Luiz, J.P.M.; Oliveira, F.F.; Viacava, P.R.; Leite, C.A.; Pereira, J.A.; Silva, C.M.; Silva, C.R.; Silva, R.L.; et al. Paclitaxel Reduces Tumor Growth by Reprogramming Tumor-Associated Macrophages to an M1 Profile in a TLR4-Dependent Manner. Cancer Res. 2018, 78, 5891–5900. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Marchesi, F.; Garlanda, C. Macrophages as Tools and Targets in Cancer Therapy. Nat. Rev. Drug Discov. 2022, 21, 799–820. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Wei, S.; Hurt, E.M.; Green, M.D.; Zhao, L.; Vatan, L.; Szeliga, W.; Herbst, R.; Harms, P.W.; Fecher, L.A.; et al. Host Expression of PD-L1 Determines Efficacy of PD-L1 Pathway Blockade–Mediated Tumor Regression. J. Clin. Investig. 2018, 128, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Mittman, S.; Rodriguez, R.; Moskalenko, M.; Pacheco-Sanchez, P.; Yang, Y.; Nickles, D.; Cubas, R. Anti-PD-L1 Treatment Results in Functional Remodeling of the Macrophage Compartment. Cancer Res. 2019, 79, 1493–1506. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Ni, T.; Wang, J.; Liu, Y.; Fan, Q.; Wang, Y.; Huang, T.; Chu, Y.; Sun, X.; Wang, Y. CD47 Blockade Inhibits Tumor Progression through Promoting Phagocytosis of Tumor Cells by M2 Polarized Macrophages in Endometrial Cancer. J. Immunol. Res. 2018, 2018, 6156757. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Saito, Y.; Kotani, T.; Matozaki, T. CD47-Signal Regulatory Protein α Signaling System and Its Application to Cancer Immunotherapy. Cancer Sci. 2018, 109, 2349–2357. [Google Scholar] [CrossRef]
- Pan, Y.; Lu, F.; Fei, Q.; Yu, X.; Xiong, P.; Yu, X.; Dang, Y.; Hou, Z.; Lin, W.; Lin, X.; et al. Single-Cell RNA Sequencing Reveals Compartmental Remodeling of Tumor-Infiltrating Immune Cells Induced by Anti-CD47 Targeting in Pancreatic Cancer. J. Hematol. Oncol. 2019, 12, 124. [Google Scholar] [CrossRef]
- Hayat, S.M.G.; Bianconi, V.; Pirro, M.; Jaafari, M.R.; Hatamipour, M.; Sahebkar, A. CD47: Role in the Immune System and Application to Cancer Therapy. Cell. Oncol. 2020, 43, 19–30. [Google Scholar] [CrossRef]
- Locati, M.; Curtale, G.; Mantovani, A. Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 123–147. [Google Scholar] [CrossRef] [PubMed]
- Newick, K.; Moon, E.; Albelda, S.M. Chimeric Antigen Receptor T-Cell Therapy for Solid Tumors. Mol. Ther. Oncolytics 2016, 3, 16006. [Google Scholar] [CrossRef] [PubMed]
- Kankeu Fonkoua, L.A.; Sirpilla, O.; Sakemura, R.; Siegler, E.L.; Kenderian, S.S. CAR T Cell Therapy and the Tumor Microenvironment: Current Challenges and Opportunities. Mol. Ther. Oncolytics 2022, 25, 69–77. [Google Scholar] [CrossRef]
- Klichinsky, M.; Ruella, M.; Shestova, O.; Lu, X.M.; Best, A.; Zeeman, M.; Schmierer, M.; Gabrusiewicz, K.; Anderson, N.R.; Petty, N.E.; et al. Human Chimeric Antigen Receptor Macrophages for Cancer Immunotherapy. Nat. Biotechnol. 2020, 38, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Guo, M.; Xu, J.; Wu, F.; Fan, J.; Huang, Q.; Yang, G.; Lv, Z.; Wang, X.; Jin, Y. Nanoparticles Targeting Macrophages as Potential Clinical Therapeutic Agents against Cancer and Inflammation. Front. Immunol. 2019, 10, 1998. [Google Scholar] [CrossRef] [PubMed]
- Penn, C.A.; Yang, K.; Zong, H.; Lim, J.Y.; Cole, A.; Yang, D.; Baker, J.; Goonewardena, S.N.; Buckanovich, R.J. Therapeutic Impact of Nanoparticle Therapy Targeting Tumor-Associated Macrophages. Mol. Cancer Ther. 2018, 17, 96–106. [Google Scholar] [CrossRef]
- He, Y.; de Araújo Júnior, R.F.; Cruz, L.J.; Eich, C. Functionalized Nanoparticles Targeting Tumor-Associated Macrophages as Cancer Therapy. Pharmaceutics 2021, 13, 1670. [Google Scholar] [CrossRef]
- Andersen, M.H. The Targeting of Tumor-Associated Macrophages by Vaccination. Cell Stress 2019, 3, 139–140. [Google Scholar] [CrossRef]
- Che, Y.; Yang, Y.; Suo, J.; An, Y.; Wang, X. Induction of Systemic Immune Responses and Reversion of Immunosuppression in the Tumor Microenvironment by a Therapeutic Vaccine for Cervical Cancer. Cancer Immunol. Immunother. 2020, 69, 2651–2664. [Google Scholar] [CrossRef] [PubMed]
- Hiremath, S.B.; Devendrappa, S.L. Safety and Efficacy of Tirapazamine as Anti-Cancer Drug: A Meta-Analysis of Randomized Controlled Trials. Int. J. Basic Clin. Pharmacol. 2018, 7, 783. [Google Scholar] [CrossRef]
- Ou, Z.; Lin, S.; Qiu, J.; Ding, W.; Ren, P.; Chen, D.; Wang, J.; Tong, Y.; Wu, D.; Chen, A.; et al. Single-Nucleus RNA Sequencing and Spatial Transcriptomics Reveal the Immunological Microenvironment of Cervical Squamous Cell Carcinoma. Adv. Sci. 2022, 9, 2203040. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.Y.; Ng, C.W.; Rajapakse, M.P.; Ang, N.; Yeong, J.P.S.; Lau, M.C. The Promise and Challenge of Spatial Omics in Dissecting Tumour Microenvironment and the Role of AI. Front. Oncol. 2023, 13, 1172314. [Google Scholar] [CrossRef] [PubMed]
- Ptacek, J.; Locke, D.; Finck, R.; Cvijic, M.E.; Li, Z.; Tarolli, J.G.; Aksoy, M.; Sigal, Y.; Zhang, Y.; Newgren, M.; et al. Multiplexed Ion Beam Imaging (MIBI) for Characterization of the Tumor Microenvironment across Tumor Types. Lab. Investig. 2020, 100, 1111–1123. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, D.; Wauters, E.; Boeckx, B.; Aibar, S.; Nittner, D.; Burton, O.; Bassez, A.; Decaluwé, H.; Pircher, A.; Van den Eynde, K.; et al. Phenotype Molding of Stromal Cells in the Lung Tumor Microenvironment. Nat. Med. 2018, 24, 1277–1289. [Google Scholar] [CrossRef] [PubMed]
- Laviron, M.; Petit, M.; Weber-Delacroix, E.; Combes, A.J.; Arkal, A.R.; Barthélémy, S.; Courau, T.; Hume, D.A.; Combadière, C.; Krummel, M.F.; et al. Tumor-Associated Macrophage Heterogeneity Is Driven by Tissue Territories in Breast Cancer. Cell Rep. 2022, 39, 110865. [Google Scholar] [CrossRef]
- Yin, W.; Ping, Y.F.; Li, F.; Lv, S.Q.; Zhang, X.N.; Li, X.G.; Guo, Y.; Liu, Q.; Li, T.R.; Yang, L.Q.; et al. A Map of the Spatial Distribution and Tumour-Associated Macrophage States in Glioblastoma and Grade 4 IDH-Mutant Astrocytoma. J. Pathol. 2022, 258, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Peng, C.W.; Yang, G.F.; Hu, W.Q.; Yang, X.J.; Huang, C.Q.; Xiong, B.; Li, Y. Distribution Pattern of Tumor Associated Macrophages Predicts the Prognosis of Gastric Cancer. Oncotarget 2017, 8, 92757–92769. [Google Scholar] [CrossRef] [PubMed]
- Artemova, D.; Vishnyakova, P.; Khashchenko, E.; Elchaninov, A.; Sukhikh, G.; Fatkhudinov, T. Endometriosis and Cancer: Exploring the Role of Macrophages. Int. J. Mol. Sci. 2021, 22, 5196. [Google Scholar] [CrossRef] [PubMed]
- Haque, A.S.M.R.; Moriyama, M.; Kubota, K.; Ishiguro, N.; Sakamoto, M.; Chinju, A.; Mochizuki, K.; Sakamoto, T.; Kaneko, N.; Munemura, R.; et al. CD206+ Tumor-Associated Macrophages Promote Proliferation and Invasion in Oral Squamous Cell Carcinoma via EGF Production. Sci. Rep. 2019, 9, 14611. [Google Scholar] [CrossRef] [PubMed]
- De Logu, F.; Ugolini, F.; Iannone, L.F.; Simi, S.; Maio, V.; de Giorgi, V.; Maria di Giacomo, A.; Miracco, C.; Cossu, A.; Palmieri, G.; et al. Spatial Proximity and Relative Distribution of Tumor-Infiltrating Lymphocytes and Macrophages Predict Survival in Melanoma. Lab. Investig. 2023, 103, 100259. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, X.; Huang, Q.; Zhang, M.; Lei, T.; Wang, F.; Zou, W.; Huang, R.; Hu, X.; Wang, C.; et al. Single-Cell RNA-Sequencing Reveals Radiochemotherapy-Induced Innate Immune Activation and MHC-II Upregulation in Cervical Cancer. Signal Transduct. Target. Ther. 2023, 8, 44. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Qu, X.; Tang, X.; Song, Y.; Wang, J.; Hua, K.; Qiu, J. Spatiotemporally Deciphering the Mysterious Mechanism of Persistent HPV-induced Malignant Transition and Immune Remodelling from HPV-infected Normal Cervix, Precancer to Cervical Cancer: Integrating Single-cell RNA-sequencing and Spatial Transcriptome. Clin. Transl. Med. 2023, 13, e1219. [Google Scholar] [CrossRef]
- Cheng, S.; Li, Z.; Gao, R.; Xing, B.; Gao, Y.; Yang, Y.; Qin, S.; Zhang, L.; Ouyang, H.; Du, P.; et al. A Pan-Cancer Single-Cell Transcriptional Atlas of Tumor Infiltrating Myeloid Cells. Cell 2021, 184, 792–809.e23. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.Y.; Black, A.; Qian, B.Z. Macrophage Diversity in Cancer Revisited in the Era of Single-Cell Omics. Trends Immunol. 2022, 43, 546–563. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hua, K. Dissecting the Single-Cell Transcriptome Network of Immune Environment Underlying Cervical Premalignant Lesion, Cervical Cancer and Metastatic Lymph Nodes. Front. Immunol. 2022, 13, 897366. [Google Scholar] [CrossRef] [PubMed]
- Yue, S.; Wang, Q.; Zhang, J.; Hu, Q.; Liu, C. Understanding Cervical Cancer at Single-Cell Resolution. Cancer Lett. 2023, 576, 216408. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Q.; Chen, G.; Luo, D. Multi-Omics Analysis Showed the Clinical Value of Gene Signatures of C1QC+ and SPP1+ TAMs in Cervical Cancer. Front. Immunol. 2021, 12, 694801. [Google Scholar] [CrossRef] [PubMed]
- Khanduri, I.; Maru, D.M.; Parra, E.R. Exploratory Study of Macrophage Polarization and Spatial Distribution in Colorectal Cancer Liver Metastasis: A Pilot Study. Front. Immunol. 2023, 14, 1223864. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lin, J.; Wang, G.; Xu, D. Targeting Proliferating Tumor-Infiltrating Macrophages Facilitates Spatial Redistribution of CD8+ T Cells in Pancreatic Cancer. Cancers 2022, 14, 1474. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Guo, Z.; Wei, X.; Zhao, G.; Han, D.; Zhang, T.; Chen, X.; Cao, F.; Dong, J.; Zhao, L.; et al. Spatial Distribution and Predictive Significance of Dendritic Cells and Macrophages in Esophageal Cancer Treated With Combined Chemoradiotherapy and PD-1 Blockade. Front. Immunol. 2022, 12, 786429. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Yang, Y.C.; An, Z.J.; Zhang, M.H.; Fu, X.H.; Huang, Z.F.; Yuan, Y.; Hou, J. Advances in Spatial Transcriptomics and Related Data Analysis Strategies. J. Transl. Med. 2023, 21, 330. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.G.; Lee, H.J.; Asatsuma, T.; Vento-Tormo, R.; Haque, A. An Introduction to Spatial Transcriptomics for Biomedical Research. Genome Med. 2022, 14, 68. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Lin, K.; Li, X.; Yuan, X.; Xu, P.; Ni, P.; Xu, D. Redefining Tumor-Associated Macrophage Subpopulations and Functions in the Tumor Microenvironment. Front. Immunol. 2020, 11, 1731. [Google Scholar] [CrossRef] [PubMed]
- Gorvel, L.; Panouillot, M.; Rouvière, M.-S.; Sonongbua, J.; Fattori, S.; Boucherit, N.; Amara, A.B.; Quilichini, O.; Granjeaud, S.; Degos, C.; et al. Tertiary Lymphoid Structures Are Associated with Enhanced Macrophage and Dendritic Cell Activation and Proximity to CD8+ T Cells, Which Better Predict the Clinical Outcome of Cervical Cancer Patients. bioRxiv 2023. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, Y.; Lee, D.; Kim, N.Y.; Seo, I.; Park, N.J.-Y.; Chong, G.O. Role of Tumor-Associated Macrophages in Cervical Cancer: Integrating Classical Perspectives with Recent Technological Advances. Life 2024, 14, 443. https://doi.org/10.3390/life14040443
Choi Y, Lee D, Kim NY, Seo I, Park NJ-Y, Chong GO. Role of Tumor-Associated Macrophages in Cervical Cancer: Integrating Classical Perspectives with Recent Technological Advances. Life. 2024; 14(4):443. https://doi.org/10.3390/life14040443
Chicago/Turabian StyleChoi, Yeseul, Donghyeon Lee, Na Young Kim, Incheol Seo, Nora Jee-Young Park, and Gun Oh Chong. 2024. "Role of Tumor-Associated Macrophages in Cervical Cancer: Integrating Classical Perspectives with Recent Technological Advances" Life 14, no. 4: 443. https://doi.org/10.3390/life14040443