


Leaf Morphological and Epidermal Traits Variability along Environmental Gradients in Ten Natural Populations of Pistacia lentiscus
 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
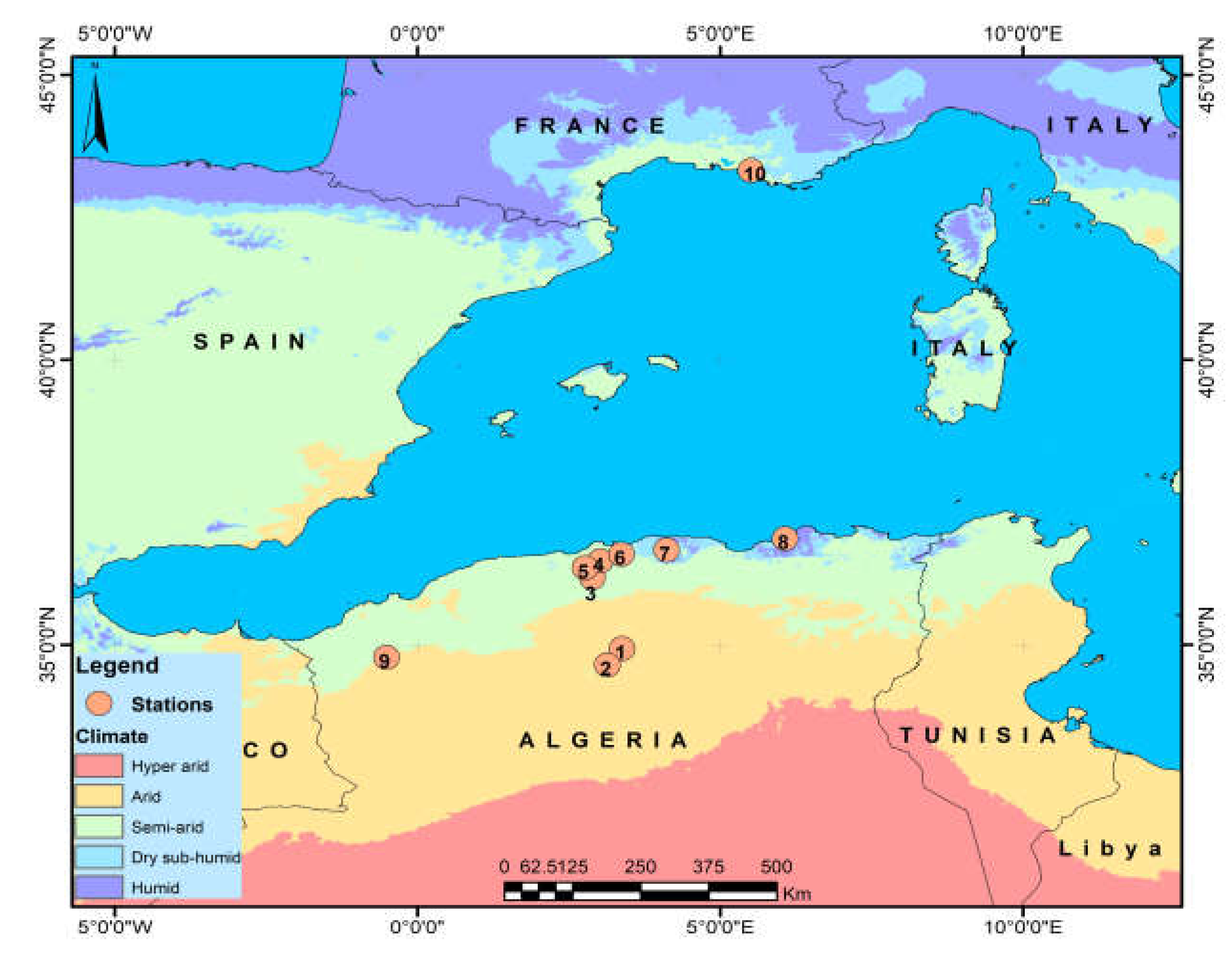
2.1. Sampling
2.2. Morphological and Ultrastructural Study of Leaves
2.2.1. Leaf Macro-Morphology
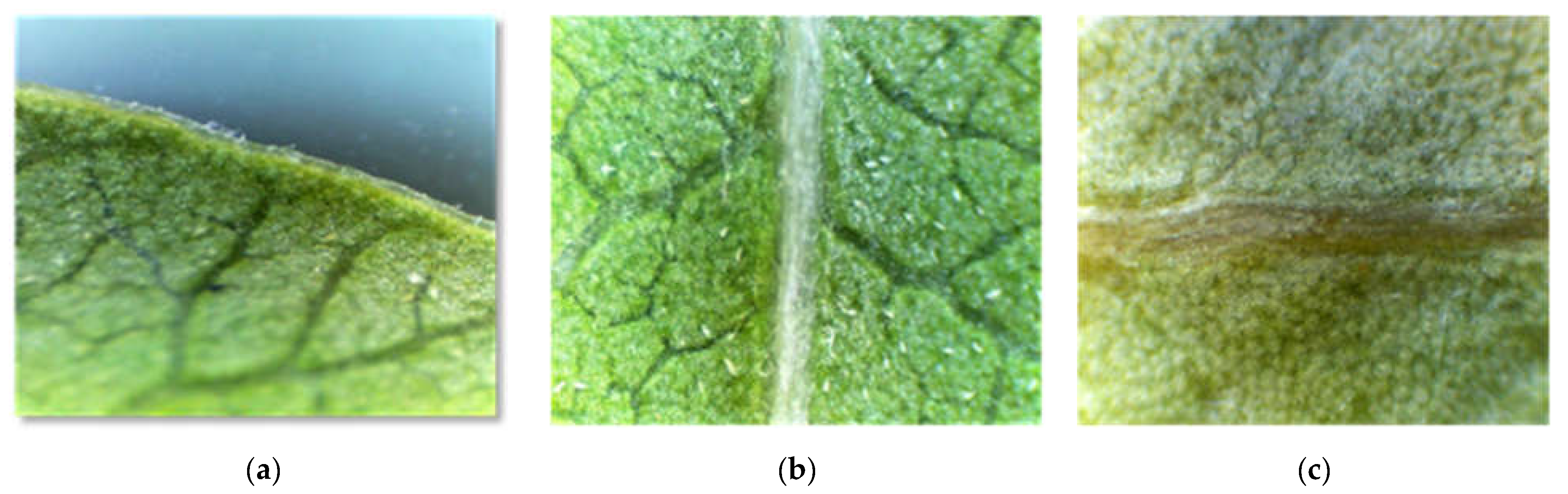
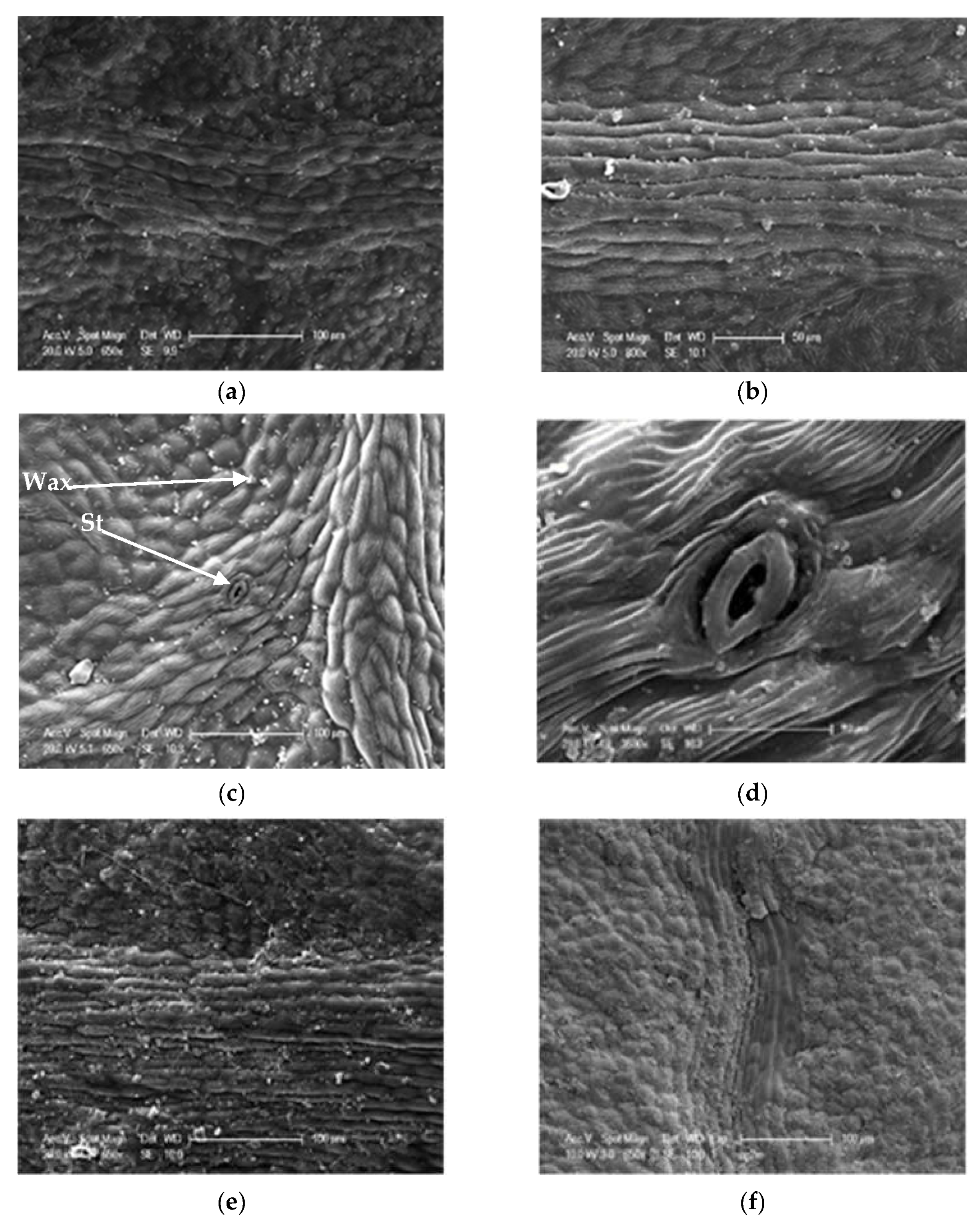
2.2.2. Epidermis, Stomata, and Trichomes
- −
- Qualitative characteristics: shape, distribution and position of stomata on the epidermis, repartition and density of trichomes (leaf margins, adaxial and abaxial surfaces, along the veins), and types of trichomes.
- −
- Quantitative characteristics: length and width of stomata on the abaxial face (µm) and stomatal density on both abaxial and adaxial faces (number of st/mm2).
2.3. Statistical Data Processing
3. Results
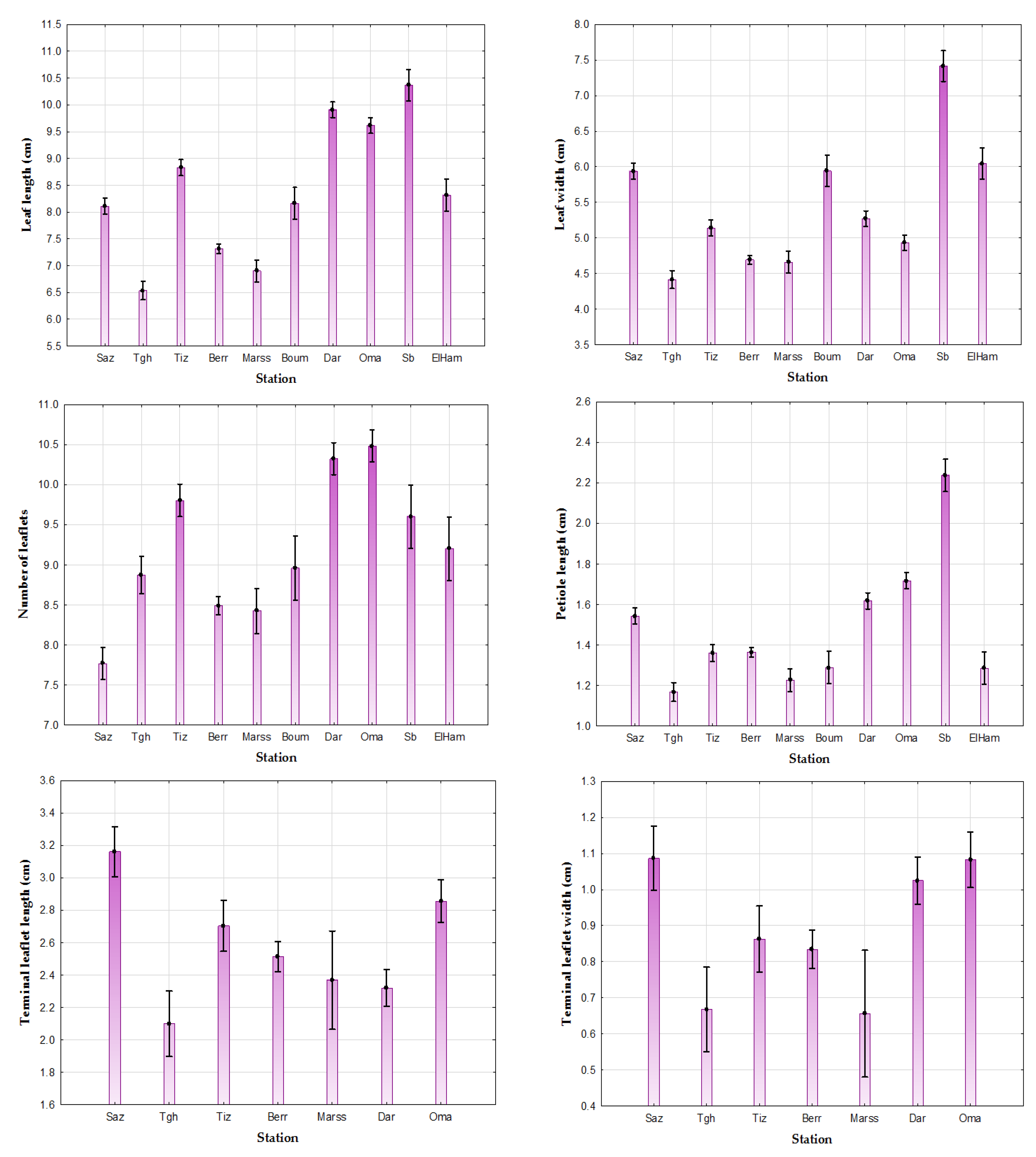
3.1. Inter-Population Variability of Quantitative Macro-Morphological Traits of the Leaf and Terminal Leaflet
3.1.1. Provenance Effect on the Leaf
- Seven distinct groups for the length of the leaf: Group 1: Saz, Boum, and ElHam; Group 2: Oma and Dar; Group 3: Tgh; Group 4: Marss; Group 5: Berr; Group 6: Tiz; and Group 7: Sb.
- Six distinct groups for the width of the leaf: Group 1: Saz, Boum, and ElHam; Group 2: Marss and Berr; Group 3: Oma and Tiz; Group 4: Tiz and Dar; Group 5: Tgh; and Group 6: Sb.
- Six distinct groups for the number of leaflets: Group 1: Marss, Berr, and Tgh; Group 2: Berr, Tgh, and Boum; Group 3: Tgh, Boum, and ElHam; Group 4: Sb and Tiz; Group 5: Dar and Oma; and Group 6: Saz.
- Six distinct groups for the length of the petiole: Group 1: ElHam, Boum, Tiz, and Berr; Group 2: ElHam, Boum, and Marss; Group 3: Marss and Tgh; Group 4: Saz and (Dar); Group 5: Oma; and Group 6: Sb (Table 3).
3.1.2. Provenance Effect on the Terminal Leaflet
- Five distinct groups for the length of the terminal leaflet: Group 1: Tgh, Dar, and Marss; Group 2: Dar, Marss, and Berr; Group 3: Berr and Tiz; Group 4: Tiz and Oma; and Group 5: Saz.
- Three groups for the width of the terminal leaflet: Group 1: Dar, Oma, and Saz; Group 2: Marss and Tgh; and Group 3: Berr and Tiz.
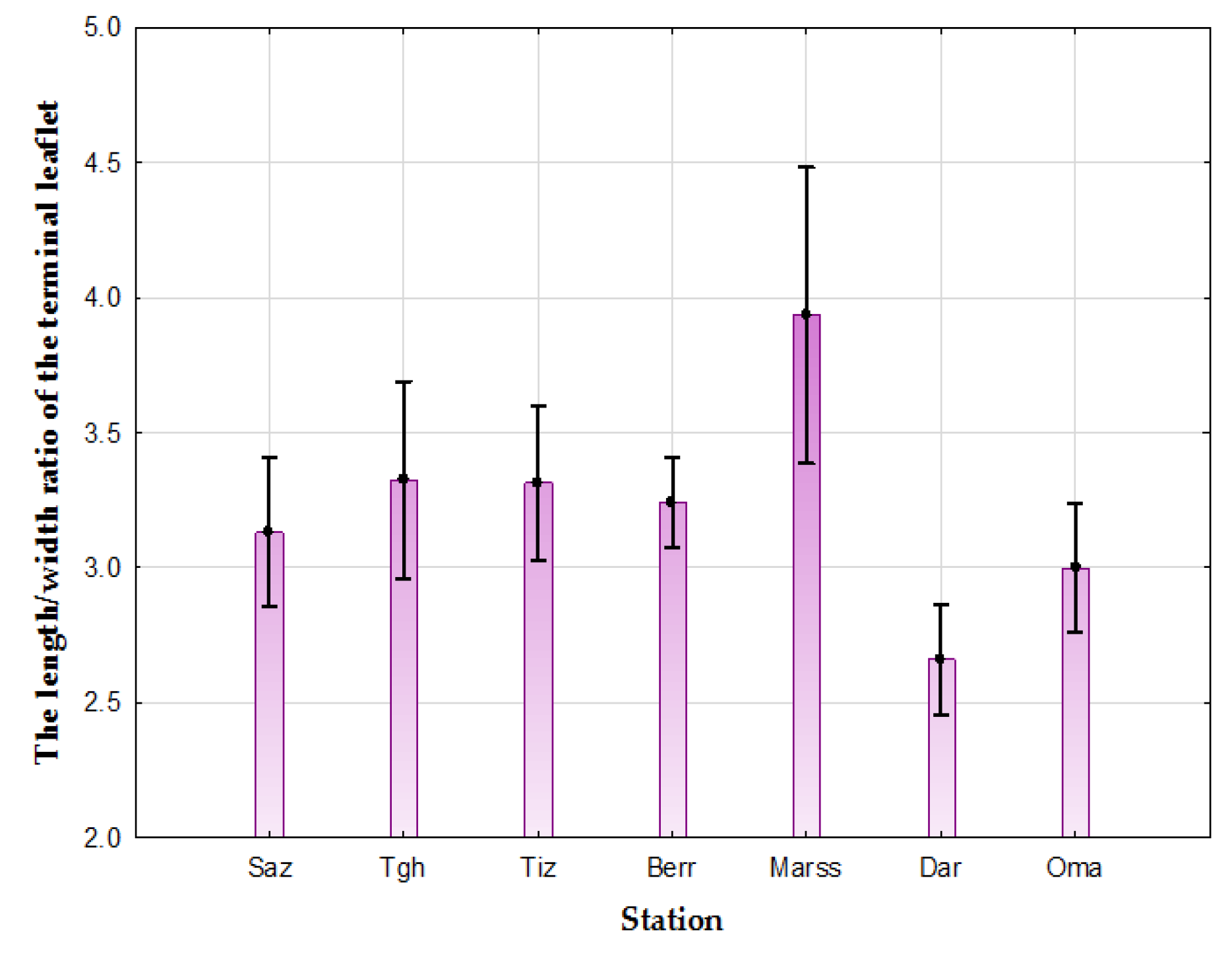
- Three groups for the length/width ratio of the terminal leaflet: Group 1: Oma, Saz, Berr, Tiz, and Tgh; Group 2: Dar, Oma, Saz, and Berr; and Group 3: Marss. (Table 2).
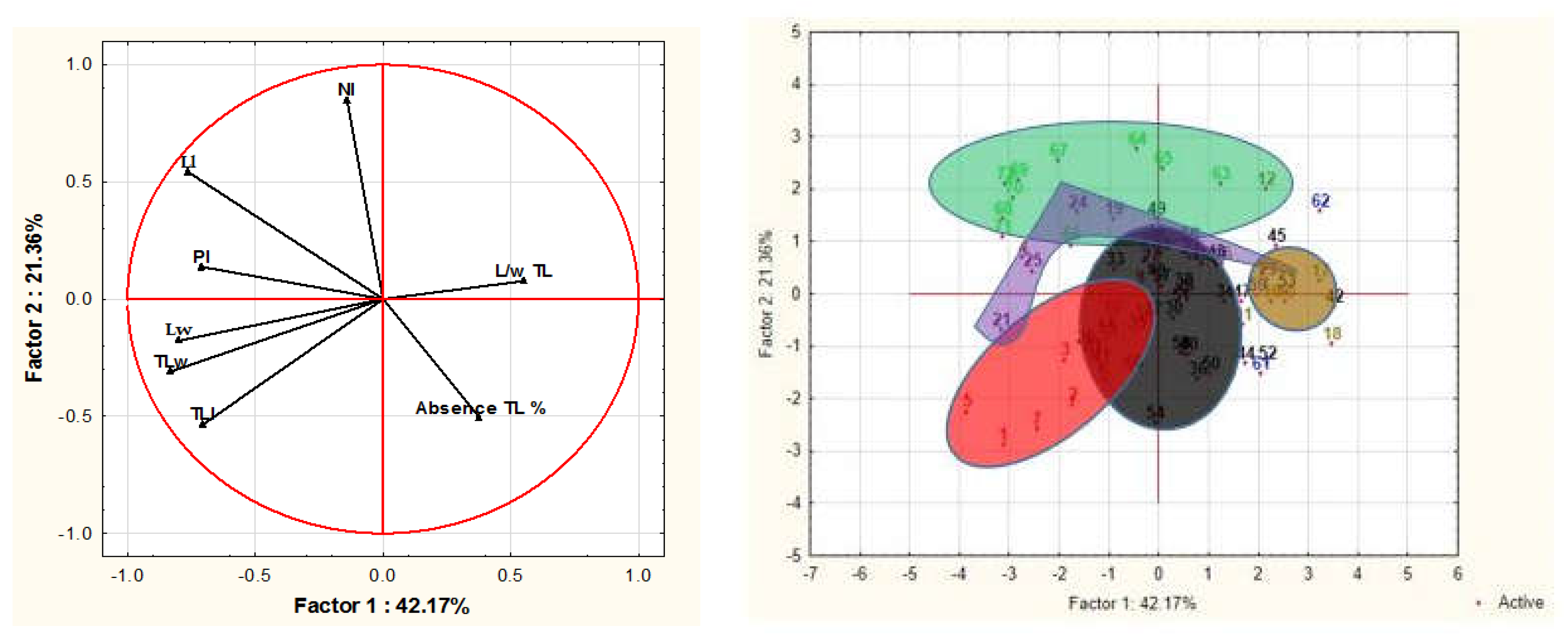
3.1.3. Principal Component Analysis (PCA)
3.2. Inter-Population Variability of Qualitative Macro-Morphological Traits of the Leaf and Terminal Leaflet
3.2.1. Leaf
3.2.2. The Terminal Leaflet
3.3. Micromorphological Traits of the Leaf
3.3.1. The Trichomes
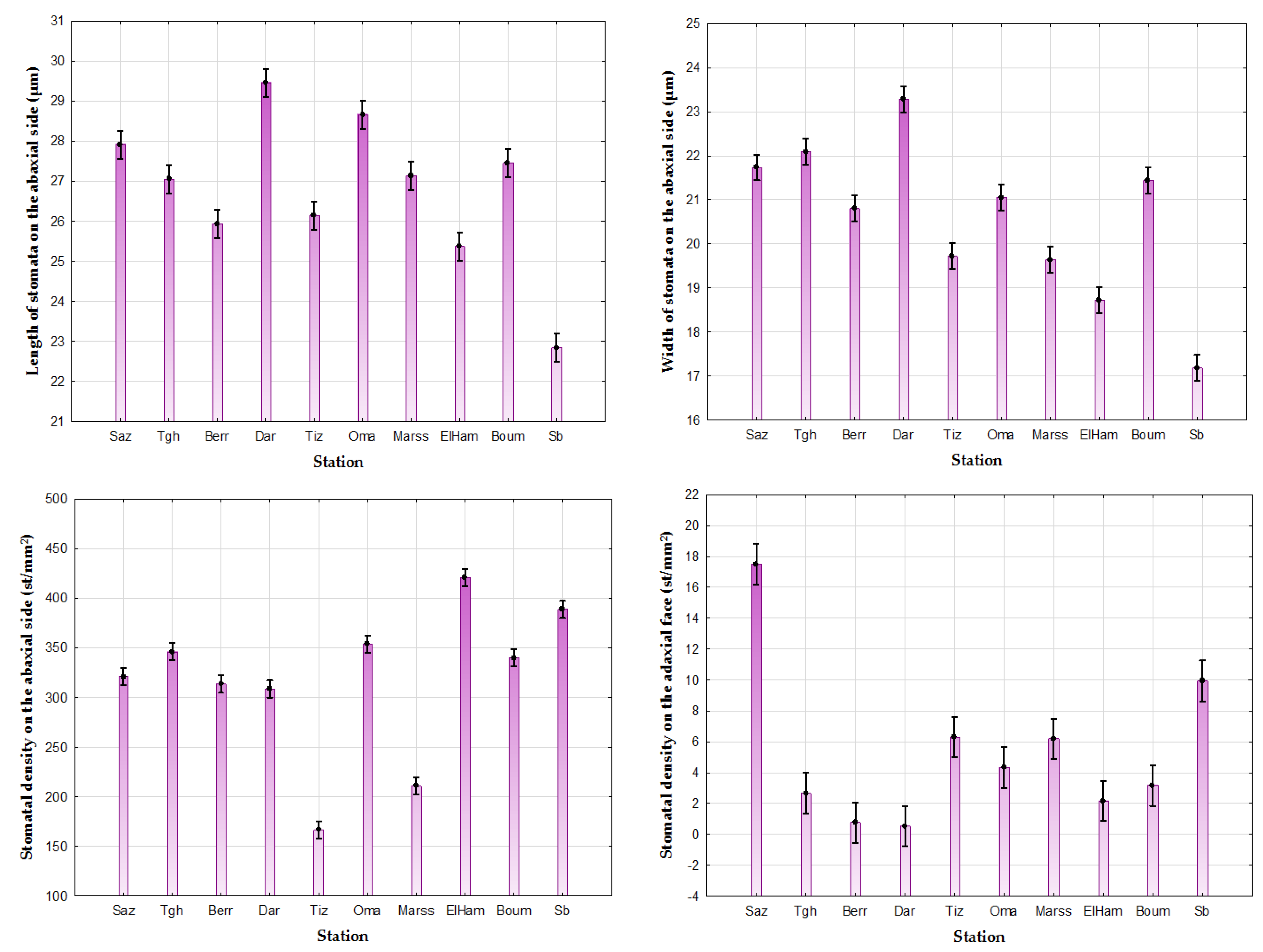
3.3.2. The Stomata
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Codification | Value | |
|---|---|---|
| Leaf rachis wing (Lrw) | 0 | Absent |
| 1 | Present in leaf rachis only | |
| 2 | Present in both leaf rachis and petiole | |
| Presence of terminal leaflet (TL) | 0 | Absent (even pinnate leaf) |
| 1 | Present (odd pinnate leaf) | |
| Size of terminal leaflet relative to basal leaflets (STL) | 1 | Smaller than lateral ones |
| 2 | As large as lateral ones | |
| 3 | Larger than lateral ones | |
| Shape of the terminal leaflet (ShTL) | 1 | Lanceolat |
| 2 | Ovate | |
| 3 | Ovate-oblong | |
| 4 | Oblone | |
| 5 | Elliptic | |
| 6 | Narrow elliptic | |
| 99 | Other | |
| Form of the apex of the terminal leaflet (FA) | 1 | Mucronulate |
| 2 | Acuminte | |
| 3 | Mucronate | |
| 4 | Caudate | |
| 5 | Cuspidate | |
| 6 | Acute | |
| 7 | Obtuse | |
| 8 | Retuse | |
| 9 | Emarginate | |
| Leaf margin (Lm) | 1 | Leathery |
| 2 | Membranaceous | |
| Form of the petiole (Fp) | 1 | Flattznzd |
| 2 | Rounded | |
| 3 | Rounded straight adaxially | |
| Leaf color (Lc) | 1 | Light green |
| 2 | Grenn | |
| 3 | Dark grenn | |
| 99 | Other |
References
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of Earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Rosenthal, J.P.; Kotanen, P.M. Terrestrial plant tolerance to herbivory. Trends Ecol. Evol. 1994, 9, 145–148. [Google Scholar] [CrossRef]
- Zohary, M. A monographical study of the genus Pistacia. Palest. J. Bot. 1952, 5, 187–228. [Google Scholar]
- Belhadj, S. Etude Eco-Botanique de P. atlantica Desf. (Anacardiaceae) en Algérie Préalable à la Conservation des Ressources Génétiques de L’espèce et Sa Valorisation. Ph.D. Thesis, Université Mouloud Mammeri de Tizi Ouzou, Tizi Ouzou, Algeria, 2007. [Google Scholar]
- Bellhadj, S.; Derridj, A.; Aigouy, T.; Gers, C.; Gauquelin, T.; Mevy, J.P. Comparative morphology of leaf epidermis in eight populations of Atlas pistachio (Pistacia atlantica Desf., Anacardiaceae). Microsc. Res. Tech. 2007, 70, 837–846. [Google Scholar] [CrossRef]
- Belhadj, S.; Derridj, A.; Auda, Y.; Gers, C.; Gauquelin, T. Analyse de la variabilité morphologique chez huit populations spontanées de Pistacia atlantica en Algérie. Botany 2008, 86, 520–532. [Google Scholar] [CrossRef]
- Belhadj, S.; Derridj, A.; Moriana, A.; Gijon, M.D.C.; Mevy, J.P.; Gauquelin, T. Comparative analysis of stomatal characters in eight wild atlas pistachio populations (Pistacia atlantica Desf.; Anacardiaceae). Int. Res. J. Plant Sci. 2011, 2, 060–069. [Google Scholar]
- Ait Said, S.; Fernandez, C.; Greff, S.; Derridj, A.; Gauquelin, T.; Mevy, J.P. Inter population variability of leaf morpho-anatomical and terpenoid patterns of Pistacia atlantica Desf ssp atlantica growing along an aridity gradient in Algeria. Flora Morphol. Distrib. Funct. Ecol. Plants 2011, 206, 397–405. [Google Scholar] [CrossRef]
- Tirse, M. Etude microphytodermique des folioles de Pistacia et des genres affiliés à la famille des Anacardiaceae en région Nord-occidentale oranaise. Ph.D. Thesis, Université de Sidi Bel Abbes, Sidi Bel Abbes, Algeria, 2016. [Google Scholar]
- IPGRI. Descriptors for Pistacia spp. (Excluding P. vera); International Plant Genetic Resources Institute: Rome, Italy, 1998. [Google Scholar]
- Bouzillé, J.B. Gestion des Habitats Naturels et Biodiversité; Lavoisier: Paris, France, 2007; 330p. [Google Scholar]
- Alyafi, J. Approches Systématiques et Ecologiques du Genre Pistacia dans la Région Méditerranéenne. Ph.D. Thesis, Faculté des Sciences et Techniques de Saint-Jérôme, Marseille, France, 1979; p. 123. [Google Scholar]
- Barboni, D.; Harrison, S.P.; Bartlein, P.J.; Jalut, G.; New, M.; Prentice, I.C.; Stevenson, A.C. Relationships between plant traits and climate in the Mediterranean region: A pollen data analysis. J. Veg. Sci. 2004, 15, 635–646. [Google Scholar] [CrossRef]
- Fahn, A. Plant Anatomy; Pergamon Press: Exeter, UK, 1967. [Google Scholar]
- Kofidis, G.; Bosabalidis, A.M.; Moustakas, M. Combined effects of altitude and season on leaf characteristics of Clinopodium vulgare L. (Labiatae). Environ. Exp. Bot. 2007, 60, 69–76. [Google Scholar] [CrossRef]
- Taib, N.; Sitayeb, T. Effect of environmental conditions on morphological variability of leaves and fruits of five populations of Pistacia atlantica Desf. in North Algeria. Biodiv. Res. Conserv 2020, 58, 1–12. [Google Scholar]
- Werker, E. Function of essential oil-secreting glandular hairs in aromatic plans of Lamiacea—A review. Flavour Fragr. J. 1993, 8, 249–255. [Google Scholar] [CrossRef]
- Bosabalidis, A.M. Glandular trichomes in Satureja thymbra leaves. Ann. Bot. 1990, 65, 71–78. [Google Scholar] [CrossRef]
- Monjauze, A. Connaissance du « betoum » Pistacia at lant ica Desf. Biologie et forêt. Rev. For. Fran. 1980, 4, 357–363. [Google Scholar]
- Behboodi, B.S. Pistacia atlantica Desf. 1800 in Iran. FAOCIHEAM–NUCIS-Newsl. 2004, 12, 27–29. [Google Scholar]
- El Beyrouthy, M.; Kafrouny, M.; Arnold, N.A.; Al-Hejin, A.; Siddig, L. Macroscopic, microscopic and DNA fingerprinting to fight adulteration of Banafsaj (Viola odorata L.) sold at the Lebanese herbal shops. Eur. J. Sci. Res. 2013, 104, 642–651. [Google Scholar]
- Bosabalidis, A.M.; Tsekos, I. Ultrastructure of the essential oil secretion in glandular scales of Origanum dictamnus L. leaves. In Aromatic Plants; Springer: Dordrecht, The Netherlands, 1982; pp. 3–12. [Google Scholar]
- Bosabalidis, A.; Tsekos, I. Glandular scale development and essential oil secretion in Origanum dictamnus L. Planta 1982, 156, 496–504. [Google Scholar] [CrossRef]
- Kofidis, G.; Bosabalidis, A.M.; Moustakas, M. Contemporary seasonal and altitudinal variations of leaf structural features in oregano (Origanum vulgare L.). Ann. Botany 2003, 92, 635–645. [Google Scholar] [CrossRef]
- Steyn, H.M.; Van Wyk, A.E. Taxonomic significance of trichomes in the genus Acanthopsis Harv. (Acanthaceae, tribe Acantheae). Adansonia 2021, 43, 163–176. [Google Scholar] [CrossRef]
- Böcher, T.W. Xeromorphic leaf types. Evolutionary strategies and tentative semophyletic sequences. Biol. Skr. 1979, 22, 1–71. [Google Scholar]
- Fahn, A.; Shimony, C. Glandular trichomes of Fagonia L. (Zygophyllaceae) species: Structure, development and secreted materials. Ann. Bot. 1996, 77, 25–34. [Google Scholar] [CrossRef]
- Poriazis, D.; Balkwill, K. Developmental variation in a species of Isoglossa (Acanthaceae: Ruellioideae) over a season. Bothalia 2008, 38, 131–140. [Google Scholar] [CrossRef]
- Jürgens, N. Psammophorous plants and other adaptations to desert ecosystems with high incidence of sandstorms. Feddes Repert. 1996, 107, 345–359. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, X.; Lu, C.; Zeng, X.; Li, Y.; Fu, D.; Wu, G. Non-specific lipid transfer proteins in plants: Presenting new advances and an integrated functional analysis. J. Exp. Bot. 2015, 66, 5663–5681. [Google Scholar] [CrossRef]
- Sarret, G.; Harada, E.; Choi, Y.E.; Isaure, M.P.; Geoffroy, N.; Fakra, S.; Marcus, M.A.; Birschwilks, M.; Clemens, S.; Manceau, A. Trichomes of tobacco excrete zinc as zincsubstituted calcium carbonate and other zinc-containing compounds. Plant Physiol. 2006, 141, 1021–1034. [Google Scholar] [CrossRef] [PubMed]
- Simón, L.E. Variations des caractères foliaires chez Chenopodium subg. Ambrosia sect. Adenois (Chenopodiaceae) en Amérique du Sud: Valeur taxonomique et évolutive. Adansonia 1997, 19, 293–320. [Google Scholar]
- Gindel, I. Stomatal number and size as related to soil moisture in tree xerophytes. Ecology 1969, 50, 263–267. [Google Scholar] [CrossRef]
- Al-Saghir, M.; Porter, D.M. Stomatal distribution in Pistacia species (Anacardiaceae). Int. J. Bot. 2005, 1, 183–187. [Google Scholar] [CrossRef]
- Mott, K.A.; Gibson, A.C.; O’Leary, J.W. The adaptive significance of amphistomatic leaves. Plant Cell Environ. 1982, 5, 455–460. [Google Scholar] [CrossRef]
- Barale, G.; Azar, D. Cuticules végétales dans le Crétacé inférieur du Sud du Liban. Comptes Rendus Palevol 2004, 3, 119–124. [Google Scholar] [CrossRef]
- Scienza, A.; Boselli, M. Fréquence et caractéristiques biométriques des stomates de certains porte-greffes de vigne. Vitis 1981, 20, 281–292. [Google Scholar]
- De Micco, V.; Aronne, G. Morpho-anatomical traits for plant adaptation to drought. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Springer: Berlin/Heidelberg, Germany, 2012; pp. 37–61. [Google Scholar] [CrossRef]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons, 2nd ed.; Clarendon Press: Oxford, UK, 1979; Volume 1. [Google Scholar]









| Region | P (mm) | T° Min | T° Max | Station | GPS Coordinates | Altitude | IA |
|---|---|---|---|---|---|---|---|
| Djelfa | 274.5 | 9.03 | 22.11 | Dar chioukh (Dar) | 34°56′1.58″ N | 1350 | 0.18 |
| 3°23′3.22″ E | |||||||
| Senalba (Sb) | 34°38′40.91″ N | 1296 | 0.2 | ||||
| 3°8′18.41″ N | |||||||
| Medea | 731 | 9.8 | 16.5 | Berrouaghia (Berr) | 36°10′36.18″ N | 1169 | 0.39 |
| 2°53′24.25″ E | |||||||
| El Omaria (Oma) | 36°28′20.1″ N | 779 | 0.35 | ||||
| 03°01′17.8″ E | |||||||
| Elhamdania (ElHam) | 36°21′1.33″ N | 398 | 0.37 | ||||
| 2°45′59.81″ E | |||||||
| Boumerdes | 613 | 12.2 | 21.56 | Elkahla (Boum) | 36°35′26″ N | 495 | 0.45 |
| 3°22′17″ E | |||||||
| Tizi Ouzou | 822 | 11.3 | 20.9 | Taksebt (Tiz) | 36°39′54.30″ N | 209 | 0.5 |
| 4° 6′58.86″ E | |||||||
| Jijel | 1020 | 11.1 | 19.6 | Sidi-abdelaziz (Saz) | 36°51′45.18″ N | 24 | 0.65 |
| 6° 4′21.59″ E | |||||||
| Sidi Bel Abbès | 347 | 8.5 | 20.6 | Telagh (Tgh) | 34°46′41.21″ N | 963 | 0.18 |
| 0°30′40.00″ E | |||||||
| Marseille (France) | 640 | 7.8 | 17 | Montespin (Marss) | 43°19′24.98″ N | 216 | 0.43 |
| 5°30′28.93″ E |
| Character | Avg ± SD; Extent (C.V.) | ||||||
|---|---|---|---|---|---|---|---|
| Pistacia lentiscus | |||||||
| Leaf Length (cm) | Leaf Width (cm) | Number of Leaflets | Terminal Leaflet Length (cm) | Terminal Leaflet Width (cm) | The Length/Width Ratio of the Terminal Leaflet | Petiole Length (cm) | |
| Jijel (Saz) | 8.1 a*** ± 1.33 5.1–11.5 (16.4) | 6.93 a*** ± 1.02 3.7–9.7 (17.2) | 7.76 f*** ± 1.6 4–12 (21.07) | 3.16 e*** ± 0.63 1.83–4.8 (20.4) | 1.08 a*** ± 0.35 0.45–2 (32.67) | 3.13 ab*** ± 0.86 1.64–4.87 (27.7) | 1.54 d*** ± 0.3 0.82–2.5 (20.2) |
| Tlagh (Tgh) | 6.53 c*** ± 1.1 4.5–9.7 (17.49) | 4.4 e*** ± 1.01 2.6–8.5 (23.07) | 8.8 abc*** ± 1.4 6–12 (16.78) | 2.09 a*** ± 0.54 1.35–3.3 (25.7) | 0.66 b*** ± 0.2 0.33–1.2 (37.5) | 3.32 a*** ± 0.6 2.27–4.7 (20.3) | 1.16 c*** ± 0.2 0.6–1.9 (19.34) |
| Tizi Ouzou (Tiz) | 8.82 f*** ± 1.6 5–14.5 (18.83) | 5.13 cd*** ± 1.1 2.2–8.6 (20.88) | 9.8 d*** ± 1.61 5–15 (16.48) | 2.7 cd*** ± 0.63 1.1–4 (23.57) | 0.86 c*** ± 0.27 0.2–1.6 (32.41) | 3.31 a*** ± 0.87 1.5–6 (26.26) | 1.36 a*** ± 0.3 0.5–2.8 (27.85) |
| Berrouaghia (Berr) | 7.31 e*** ± 1.3 3.2–13.5 (17.94) | 4.69 b*** ± 0.6 1.3–7.3 (20.46) | 8.49 ab*** ± 1.8 4–16 (21.99) | 2.51 bc*** ± 0.5 1.2–4.1 (20.29) | 0.83 c*** ± 0.28 0.2–2.3 (34.1) | 3.24 ab*** ± 0.9 1.34–7.5 (28.02) | 1.36 a*** ± 0.3 0.5–3 (23.6) |
| Boumerdes (Boum) | 8.16 a*** ± 1.6 5.2–12.4 (19.6) | 5.94 a*** ± 1.03 3.8–8.4 (17.46) | 8.96 bc*** ± 1.4 6–12 (15.72) | 1.28 ab*** ± 0.2 0.9–2 (19.18) | |||
| Marseille (Marss) | 6.9 d*** ± 0.97 3.6–10.2 (14.09) | 4.65 b*** ± 0.83 2.9–6.5 (17.9) | 8.42 a*** ± 1.84 5–13 (21.88) | 2.36 ab*** ± 0.65 1.2–3.5 (27.64) | 0.65 b*** ± 0.26 0.2–1.1 (40.87) | 3.93 c*** ± 1.19 2.45–7 (30.38) | 1.2 bc*** ± 0.2 0.7–1.9 (20.88) |
| Darchyoukh (Dar) | 9.9 b*** ± 0.85 7.8–12.1 (8.61) | 5.27 d*** ± 0.82 3.5–10.1 (15.56) | 10.32 e*** ± 2.03 4–16 (19.7) | 2.31 ab*** ± 0.66 0.7–4.4 (28.82) | 1.02 a*** ± 0.44 0.1–2.1 (43.59) | 2.66 b*** ± 1.61 1.16–17 (60.55) | 1.6 d*** ± 0.44 0.7–3.3 (27.3) |
| El Omaria (Oma) | 9.61 b*** ± 1.48 5.7–13.1 (15.46) | 4.93 c*** ± 0.9 2.8–7.7 (20.2) | 10.4 e*** ± 1.7 6–14 (16.46) | 2.85 d*** ± 0.7 0.6–3.9 (25.46) | 1.08 a*** ± 0.44 0.1–2.6 (41.38) | 2.99 ab*** ± 1.12 1.34–8 (37.38) | 1.7 e*** ± 0.4 0.7–4 (28.02) |
| ElHamdania (ElHam) | 8.3 a*** ± 1.06 5.6–11.2 (12.77) | 6.1 a*** ± 0.7 4.6–9.3 (12.47) | 9.2 c*** ± 1.31 6–12(14.29) | 1.28 ab*** ± 0.2 0.9–2.2 (15.85) | |||
| Senalba (Sb) | 10.3 g*** ± 1.01 7.9–12.8 (9.77) | 7.4 f*** ± 0.7 5.9–10.2 (10.63) | 9.6 d*** ± 1.5 6–12 (15.7) | 2.23 f*** ± 0.4 1.2–3.5 (19.87) | |||
| Average | 8.16 ± 1.73 3.2–14.5 (21.21) | 5.09 ± 1.14 1.3–10.2 (22.42) | 9.07 ± 1.96 4–16 (21.68) | 2.58 ± 0.67 0.6–4.81 (26.24) | 0.92 ± 0.38 0.1–2.6 (41.02) | 3.1 ± 1.14 1.16–17 (36.91) | 1.44 ± 0.41 0.5–4 (28.32) |
| Variables | Factor 1 | Factor 2 |
|---|---|---|
| Leaf length (Ll) | −0.75 | 0.53 |
| Leaf width (Lw) | −0.80 | −0.17 |
| Number of leaflet Pairs (Nl) | 0.14 | 0.84 |
| Terminal leaflet length (TLl) | −0.70 | −0.53 |
| Terminal leaflet width (TLw) | −0.82 | −0.30 |
| Terminal leaflet length/width Ratio (L/w TL) | 0.55 | 0.07 |
| Petiole length (Pl) | −0.70 | 0.13 |
| Absence terminal leaflet (TL) % | 0.37 | −0.5 |
| Provenance | P. lentiscus | Average | |||||
|---|---|---|---|---|---|---|---|
| Jijel (Saz) | Tlagh (Tgh) | Tizi-Ouzou (Tiz) | Berrouaghia (Berr) | Marseille (Marss) | |||
| Leaf rachis wing (Lrw) | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 1 | 1 | 17.56 | 0 | 0 | 0 | 2.18 | |
| 2 | 98.99 | 82.43 | 100 | 100 | 100 | 97.82 | |
| Presence of terminal leaflet (TL) | 0 | 78.93 | 82.88 | 79.58 | 81.33 | 89.33 | 81.51 |
| 1 | 21.07 | 17.12 | 20.41 | 18.66 | 10.66 | 18.49 | |
| Size of terminal leaflet relative to basal leaflets (STL) | 1 | 12.70 | 8.55 | 13.49 | 13.44 | 6 | 11.18 |
| 2 | 8.36 | 5.4 | 2.07 | 5.22 | 0.66 | 4.89 | |
| 3 | 6.02 | 3.15 | 4.84 | 0 | 4 | 2.52 | |
| Shape of the terminal leaflet (ShTL) | 1 | 0.66 | 0 | 1.03 | 0.23 | 0.66 | 0.43 |
| 2 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 3 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 4 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 5 | 0.66 | 0 | 0 | 0.78 | 0.66 | 0.53 | |
| 6 | 19.73 | 17.12 | 19.37 | 17.56 | 9.33 | 17.74 | |
| 99 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Form of the apex of the terminal leaflet (FA) | 1 | 15.05 | 14.41 | 19.03 | 14.66 | 8.66 | 14.89 |
| 2 | 0 | 0 | 0 | 0.11 | 0.66 | 0.12 | |
| 3 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 4 | 0 | 0 | 0 | 0 | 1.33 | 0.18 | |
| 5 | 2.34 | 2.7 | 0 | 0 | 0 | 0.69 | |
| 6 | 3.01 | 0 | 0 | 1.22 | 0 | 1.07 | |
| 7 | 0.33 | 0 | 0 | 2.22 | 0 | 1.12 | |
| 8 | 0 | 0 | 1.38 | 0.33 | 0 | 0.37 | |
| 9 | 0.33 | 0 | 0 | 0 | 0 | 0.06 | |
| Leaf margin (Lm) | 1 | 100 | 100 | 100 | 100 | 100 | 100 |
| 2 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Form of the petiole (Fp) | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 2 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 3 | 100 | 100 | 100 | 100 | 100 | 100 | |
| Leaf color (Lc) | 1 | 1 | 0.45 | 0 | 6.11 | 0 | 3.17 |
| 2 | 25.75 | 99.54 | 58.47 | 32.11 | 60 | 45.48 | |
| 3 | 73.24 | 0 | 41.52 | 59.33 | 40 | 50.16 | |
| 99 | 0 | 0 | 0 | 2.44 | 0 | 1.18 | |
| TrichomeMargin | 0 | 10.03 | 22.52 | 64.70 | 95 | 94.66 | 67.95 |
| 1 | 89.63 | 77.02 | 35.3 | 5 | 5.33 | 31.93 | |
| 2 | 0.33 | 0.45 | 0 | 0 | 0 | 0.1 | |
| Trichome Midrib | 0 | 19.06 | 13.51 | 70.58 | 72.33 | 91.33 | 58.01 |
| 1 | 80.93 | 86.03 | 29.41 | 27.66 | 8.66 | 41.93 | |
| 2 | 0 | 0.45 | 0 | 0 | 0 | 0.05 | |
| TrichomeAdaxial F | 0 | 23.07 | 15.76 | 88.93 | 69 | 93.33 | 60.32 |
| 1 | 76.25 | 84.23 | 11.07 | 31 | 6.66 | 39.56 | |
| 2 | 0.66 | 0 | 0 | 36.33 | 0 | 0.1 | |
| Trichome Abaxial F. | 0 | 18.06 | 11.71 | 94.46 | 59.22 | 90.66 | 54.94 |
| 1 | 81.94 | 88.29 | 5.53 | 40.77 | 9.33 | 45.05 | |
| 2 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Character | Avg ± SD; Extent (C.V.) | |||
|---|---|---|---|---|
| Pistacia lentiscus | ||||
| Length of Stomata on the Abaxial Side (µm) | Width of Stomata on the Abaxial Side (µm) | Stomatal Density on the Abaxial Side (st/mm2) | Stomatal Density on the Adaxial Face (st/mm2) | |
| Jijel (Saz) | 27.91 c*** ± 3.22 20.08–37.72 (11.5) | 21.73 de*** ± 2.97 12.6–28.86 (13.6) | 320.66 a*** ± 68.69 125–541.66 (21.4) | 17.5 f*** ± 25.99 0–125 (148.5) |
| Tlagh (Tgh) | 27.05 a*** ± 2.77 19.1–35.3 (10.23) | 22.09 e*** ± 2.39 14.66–27.5 (10.8) | 345.83 b*** ± 79.58 166.6–583.3 (23.01) | 2.66 abc*** ± 10.87 0–83.33 (407.97) |
| Berrouaghia (Berr) | 25.93 b*** ± 3.17 15.49–39.44 (12.2) | 20.8 b*** ± 3.04 14.4–37.74 (14.6) | 313.7 a*** ± 80.22 125–558.3 (25.6) | 0.76 ab*** ± 2.73 0–16.66 (357.15) |
| Dar chioukh (Dar) | 29.46 g*** ± 4.15 12.7–42.67 (14.08) | 23.27 h*** ± 2.76 14.98–31.09 (11.9) | 308.53 a*** ± 76.98 125–508.3 (24.95) | 0.53 a*** ± 2.41 0–16.66 (453.3) |
| Tizi Ouzou (Tiz) | 26.14 b*** ± 2.47 17.49–31.21 (9.44) | 19.71 a*** ± 2.23 13.22–29.15 (11.3) | 166.4 c*** ± 55.57 75–375 (33.39) | 6.3 d*** ± 7.39 0–25 (117.39) |
| Omaria (Om) | 28.66 f*** ± 1.34 18.97–32.01 (4.69) | 21.05 bc*** ± 1.45 16.70–24.55 (6.92) | 353.5 b*** ± 91.04 100–591.6 (25.73) | 4.33 cd*** ± 7.91 0–50 (182.5) |
| Marseille (Marss) | 27.13 a*** ± 1.37 19.46–31.63 (5.07) | 19.63 a*** ± 1.22 15.89–23.36 (6.22) | 210.83 d*** ± 53.04 100–383.3 (25.16) | 6.2 bd*** ± 7.79 0–25 (125.65) |
| Elhamdania (ElHam) | 25.36 e*** ± 2.43 19.25–31.88 (9.61) | 18.72 g*** ± 2.10 13.97–25.67 (11.22) | 420.5 f*** ± 67.7 283.3–591.6 (16.1) | 2.16 abc*** ± 4.35 0–25 (201.14) |
| Boumerdes (Boum) | 27.44 ac*** ± 3.30 20.67–36.64 (12.04) | 21.43 cd*** ± 2.57 15.52–29.79 (12.00) | 339.7 b*** ± 53.68 175–483.3 (15.8) | 3.16 bc*** ± 5.94 0–41.6 (187.8) |
| Senalba (Sb) | 22.84 d*** ± 2.87 16.16–34.57 (12.59) | 17.18 f*** ± 2.27 11.55–24.37 (13.21) | 388.6 e*** ± 61.46 258.3–558.3 (15.8) | 9.93 be*** ± 9.3 0–41.6 (93.7) |
| Average | 26.88 ± 3.38 12.71–42.67 (12.6) | 20.71 ± 2.96 11.55–37.74 (14.3) | 319.05 ± 99.52 75–600 (31.2) | 5.14 ± 11.27 0–125 (219.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doghbage, A.; Belhadj, S.; Belhouadjeb, F.A.; Boukerker, H.; Mevy, J.P.; Gauquelin, T.; Tonetto, A.; Merdas, S.; Touati, B.; Saimi, F.; et al. Leaf Morphological and Epidermal Traits Variability along Environmental Gradients in Ten Natural Populations of Pistacia lentiscus. Life 2023, 13, 1617. https://doi.org/10.3390/life13071617
Doghbage A, Belhadj S, Belhouadjeb FA, Boukerker H, Mevy JP, Gauquelin T, Tonetto A, Merdas S, Touati B, Saimi F, et al. Leaf Morphological and Epidermal Traits Variability along Environmental Gradients in Ten Natural Populations of Pistacia lentiscus. Life. 2023; 13(7):1617. https://doi.org/10.3390/life13071617
Chicago/Turabian StyleDoghbage, Abdelghafour, Safia Belhadj, Fathi Abdellatif Belhouadjeb, Hassen Boukerker, Jean Philippe Mevy, Thierry Gauquelin, Alain Tonetto, Saifi Merdas, Bakria Touati, Fethi Saimi, and et al. 2023. "Leaf Morphological and Epidermal Traits Variability along Environmental Gradients in Ten Natural Populations of Pistacia lentiscus" Life 13, no. 7: 1617. https://doi.org/10.3390/life13071617