
Integrated Computational Approaches for Inhibiting Sex Hormone-Binding Globulin in Male Infertility by Screening Potent Phytochemicals
 , ,
, ,
Abstract
:1. Introduction
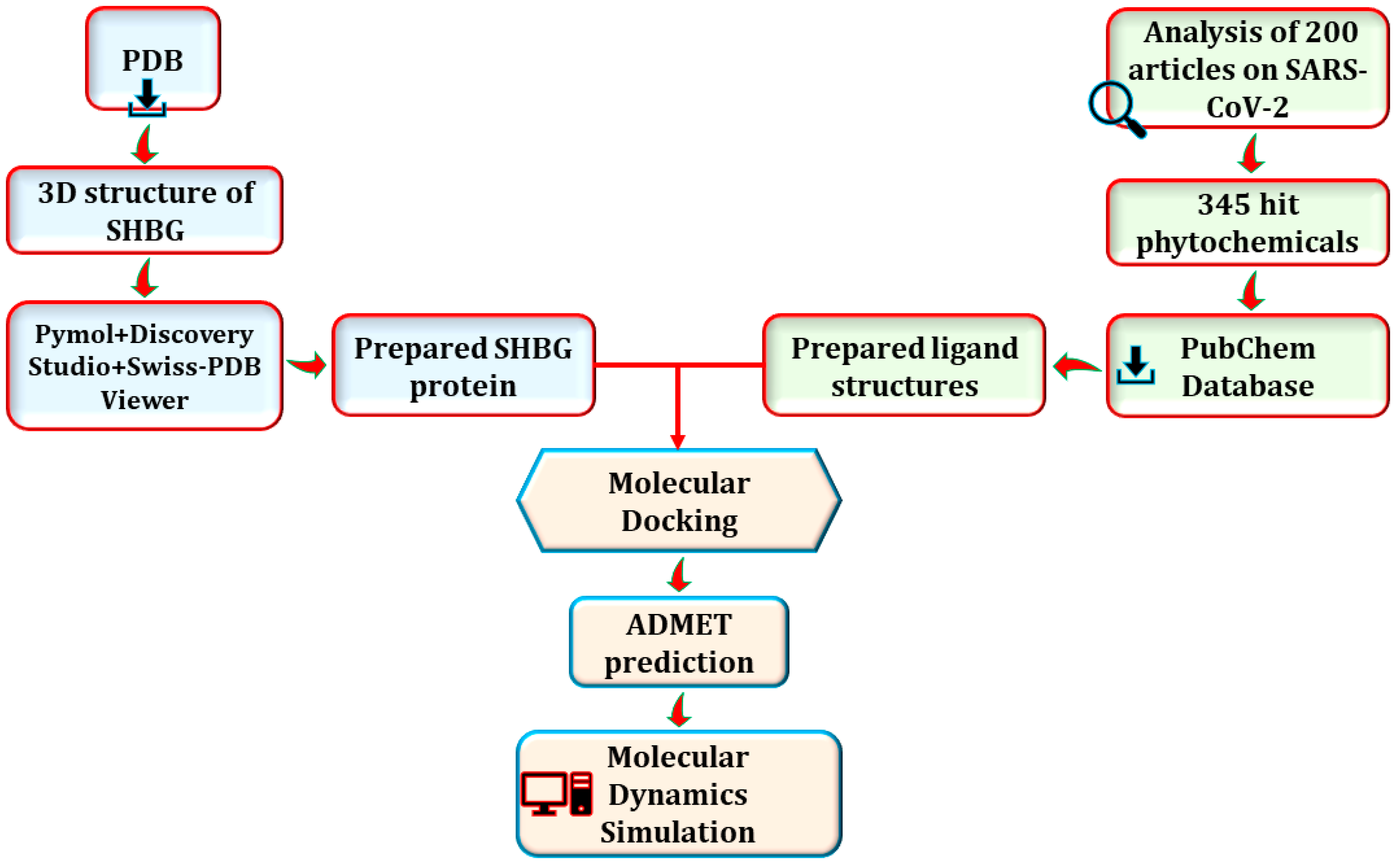
2. Materials and Methods
2.1. Protein Preparation
2.2. Ligand Preparation
2.3. Molecular Docking
2.4. ADMET Prediction
2.5. MD Simulation
3. Results
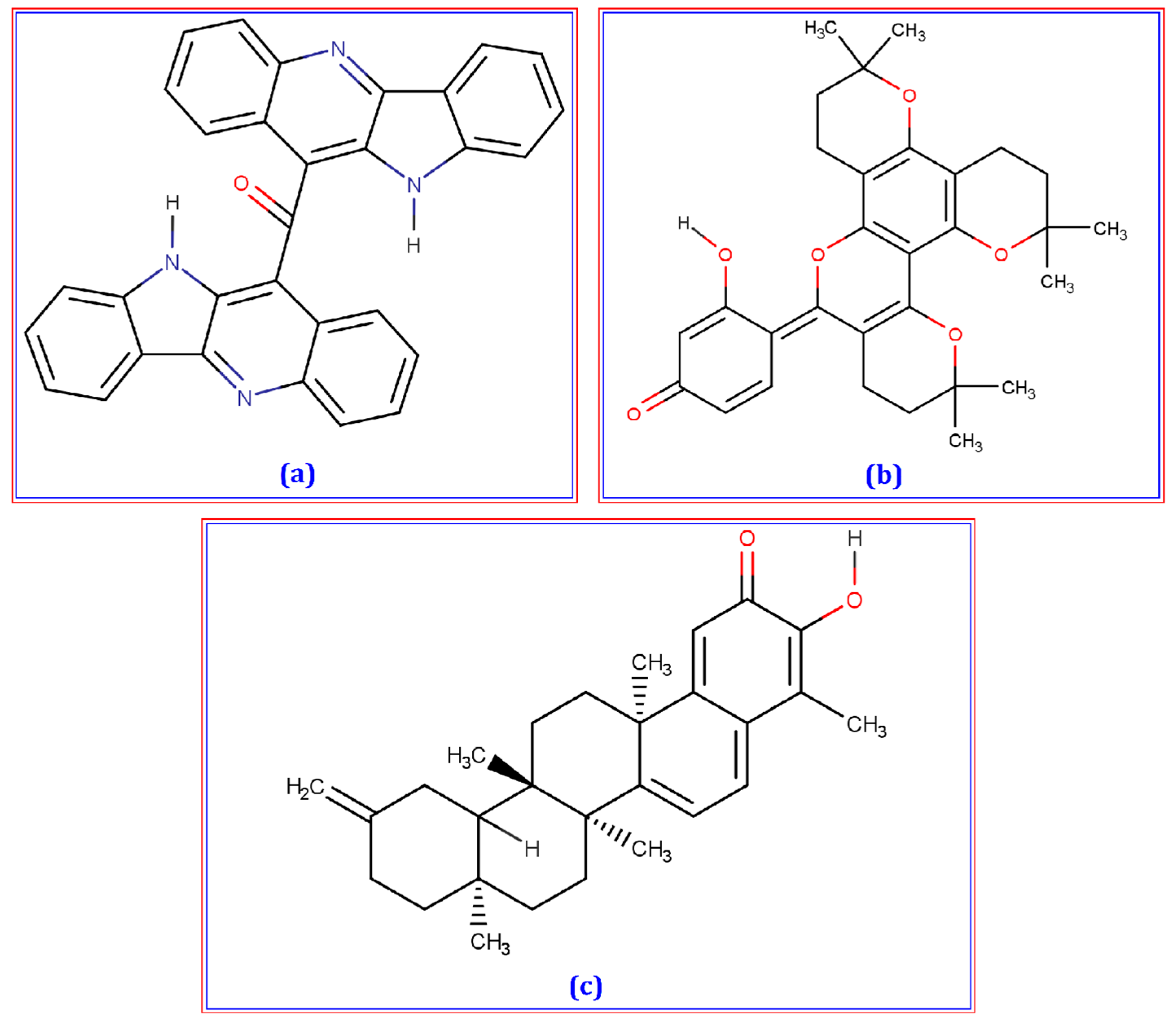
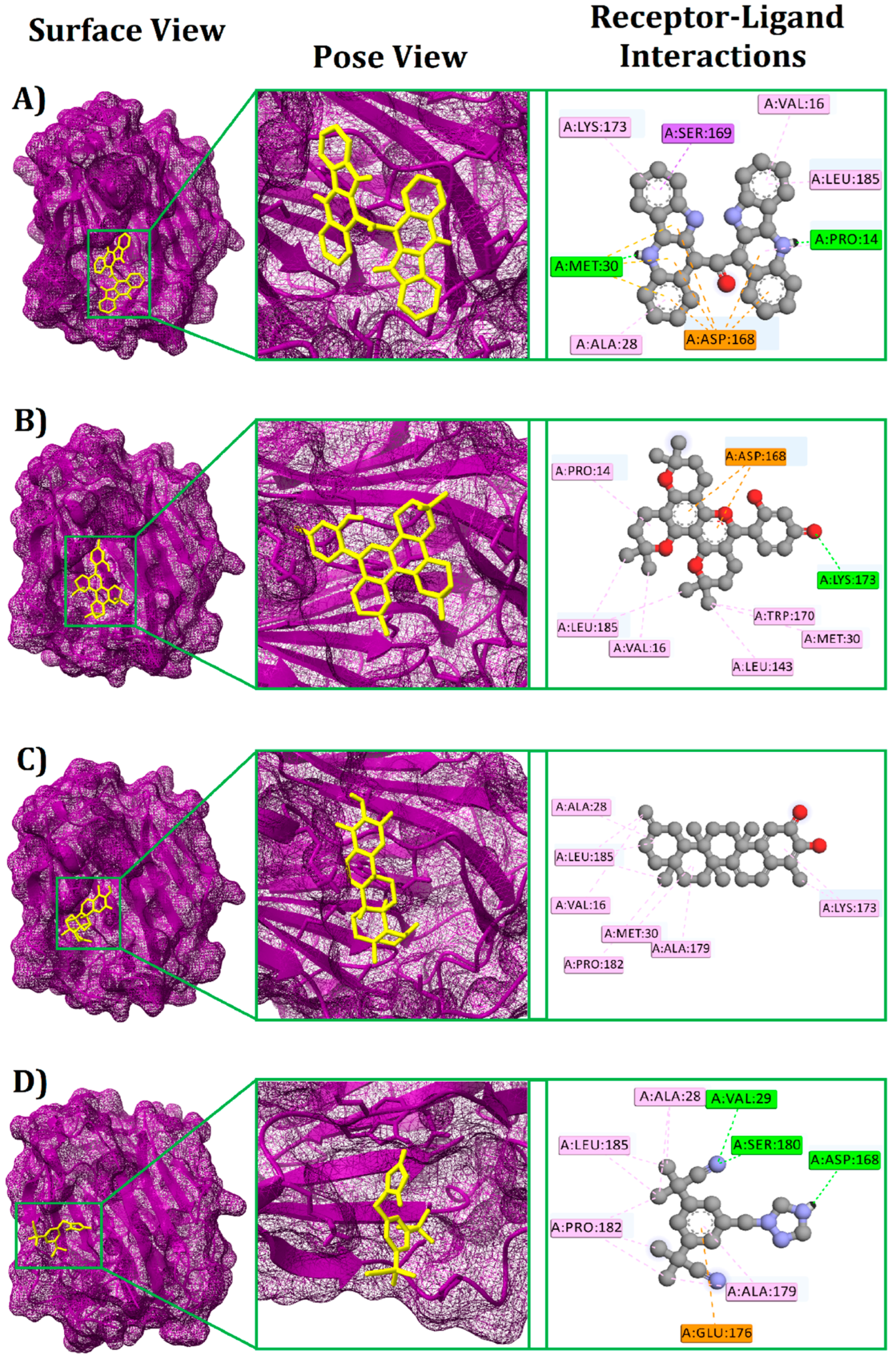
3.1. Molecular Docking
3.2. ADMET Prediction
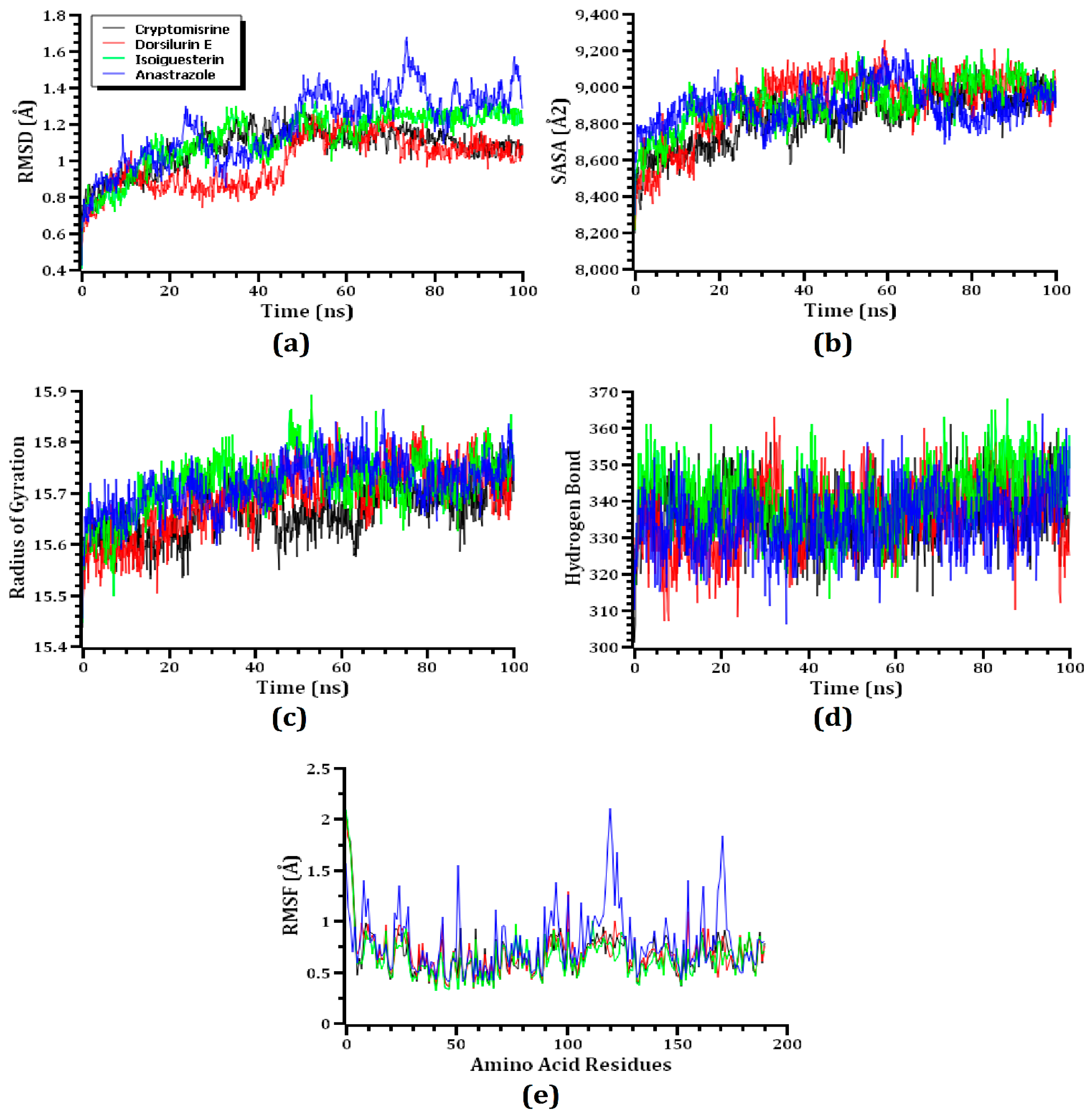
3.3. MD Simulation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kumar, N.; Singh, A. Trends of male factor infertility, an important cause of infertility: A review of literature. J. Hum. Reprod. Sci. 2015, 8, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Zegers-Hochschild, F.; Adamson, G.D.; De Mouzon, J.; Ishihara, O.; Mansour, R.; Nygren, K.; Sullivan, E.; Vanderpoel, S. International Committee for Monitoring Assisted Reproductive Technology (ICMART) and the World Health Organization (WHO) revised glossary of ART terminology, 2009*. Fertil. Steril. 2009, 92, 1520–1524. [Google Scholar] [CrossRef] [PubMed]
- Boivin, J.; Bunting, L.; Collins, J.A.; Nygren, K.G. Reply: International estimates on infertility prevalence and treatment seeking: Potential need and demand for medical care. Hum. Reprod. 2009, 24, 2380–2383. [Google Scholar] [CrossRef]
- Vander Borght, M.; Wyns, C. Fertility and infertility: Definition and epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef]
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 1–9. [Google Scholar] [CrossRef]
- Ombelet, W.; Cooke, I.; Dyer, S.; Serour, G.; Devroey, P. Infertility and the provision of infertility medical services in developing countries. Hum. Reprod. Update 2008, 14, 605–621. [Google Scholar] [CrossRef]
- Cao, X.W.; Lin, K.; Li, C.Y.; Yuan, C.W. A review of WHO Laboratory Manual for the Examination and Processing of Human Semen. Zhonghua Nan Ke Xue 2011, 17, 1059–1063. [Google Scholar]
- Lotti, F.; Maggi, M. Ultrasound of the male genital tract in relation to male reproductive health. Hum. Reprod. Update 2015, 21, 56–83. [Google Scholar] [CrossRef]
- Esther, M.Y.J.; Subramaniyan, V.; Kumar, A.P.; Subramanian, M.; Palani, M. Molecular docking, ADMET analysis and dynamics approach to potent natural inhibitors against sex hormone binding globulin in male infertility. Pharmacogn. J. 2017, 9, s35–s43. [Google Scholar] [CrossRef]
- Anwar, S.; Anwar, A. Infertility: A review on causes, treatment and management. Womens Heal. Gynecol. 2016, 5, 2–5. [Google Scholar]
- Nath, S.; Deb, B. Survey on the effect of plant extract on reproducti ve parameters of mammals: A review. Int. J. Pure App. Biosci. 2015, 3, 216–223. [Google Scholar]
- Safarinejad, M.R.; Shafiei, N.; Safarinejad, S. Association of the (TAAAA)n repeat and Asp327Asn polymorphisms in the sex hormone-binding globulin (SHBG) gene with idiopathic male infertility and relation to serum SHBG concentrations. J. Steroid Biochem. Mol. Biol. 2011, 123, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Akingbemi, B.T. Estrogen regulation of testicular function. Reprod. Biol. Endocrinol. 2005, 3, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlovich, C.P.; King, P.; Goldstein, M.; Schlegel, P.N. Evidence of a treatable endocrinopathy in infertile men. J. Urol. 2001, 165, 837–841. [Google Scholar] [CrossRef]
- Hammoud, A.O.; Gibson, M.; Peterson, C.M.; Meikle, A.W.; Carrell, D.T. Impact of male obesity on infertility: A critical review of the current literature. Fertil. Steril. 2008, 90, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Rosner, W.; Hryb, D.J.; Kahn, S.M.; Nakhla, A.M.; Romas, N.A. Interactions of sex hormone-binding globulin with target cells. Mol. Cell. Endocrinol. 2010, 316, 79–85. [Google Scholar] [CrossRef]
- Hammond, G.L. Molecular properties of corticosteroid binding globulin and the sex-steroid binding proteins. Endocr. Rev. 1990, 11, 65–79. [Google Scholar] [CrossRef]
- Siiteri, P.K.; Murai, J.T.; Hammond, G.L.; Nisker, J.A.; Raymoure, W.J.; Kuhn, R.W. The Serum Transport of Steroid Hormones. In Proceedings of the 1981 Laurentian Hormone Conference; Academic Press: Cambridge, MA, USA, 1982; Volume 38, pp. 457–510, ISBN 0125711387. [Google Scholar]
- Hong, H.; Branham, W.S.; Ng, H.W.; Moland, C.L.; Dial, S.L.; Fang, H.; Perkins, R.; Sheehan, D.; Tong, W. Human sex hormone-binding globulin binding affinities of 125 structurally diverse chemicals and comparison with their binding to androgen receptor, estrogen receptor, and α-Fetoprotein. Toxicol. Sci. 2015, 143, 333–348. [Google Scholar] [CrossRef]
- Vos, M.J.; Mijnhout, G.S.; Rondeel, J.M.M.; Baron, W.; Groeneveld, P.H.P. Sex hormone binding globulin deficiency due to a homozygous missense mutation. J. Clin. Endocrinol. Metab. 2014, 99, E1798–E1802. [Google Scholar] [CrossRef]
- Wan, Q.; Xie, Y.; Zhou, Y.; Shen, X. Research progress on the relationship between sex hormone-binding globulin and male reproductive system diseases. Andrologia 2021, 53, 1–8. [Google Scholar] [CrossRef]
- Hammond, G.L.; Bocchinfuso, W.P. Sex hormone-binding globulin: Gene organization and structure/function analyses. Horm. Res. Paediatr. 1996, 45, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Avvakumov, G.V.; Cherkasov, A.; Muller, Y.A.; Hammond, G.L. Structural analyses of sex hormone-binding globulin reveal novel ligands and function. Mol. Cell. Endocrinol. 2010, 316, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Grishkovskaya, I.; Avvakumov, G.V.; Sklenar, G.; Dales, D.; Hammond, G.L.; Muller, Y.A. Crystal structure of human sex hormone-binding globulin: Steroid transport by a laminin G-like domain. EMBO J. 2000, 19, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Hammond, G.L. Diverse roles for sex hormone-binding globulin in reproduction. Biol. Reprod. 2011, 85, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Grishkovskaya, I.; Avvakumov, G.V.; Hammond, G.L.; Catalano, M.G.; Muller, Y.A. Steroid ligands bind human sex hormone-binding globulin in specific orientations and produce distinct changes in protein conformation. J. Biol. Chem. 2002, 277, 32086–32093. [Google Scholar] [CrossRef]
- Coviello, A.D.; Haring, R.; Wellons, M.; Vaidya, D.; Lehtimäki, T.; Keildson, S.; Lunetta, K.L.; He, C.; Fornage, M.; Lagou, V.; et al. A genome-wide association meta-analysis of circulating sex hormone-binding globulin reveals multiple loci implicated in sex steroid hormone regulation. PLoS Genet. 2012, 8, e1002805. [Google Scholar] [CrossRef]
- Laurent, M.R.; Hammond, G.L.; Blokland, M.; Jardí, F.; Antonio, L.; Dubois, V.; Khalil, R.; Sterk, S.S.; Gielen, E.; Decallonne, B.; et al. Sex hormone-binding globulin regulation of androgen bioactivity in vivo: Validation of the free hormone hypothesis. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Goudas, V.T.; Dumesic, D.A. Polycystic ovary syndrome. Endocrinol. Metab. Clin. North Am. 1997, 26, 893–912. [Google Scholar] [CrossRef]
- Haring, R.; Baumeister, S.E.; Völzke, H.; Dörr, M.; Felix, S.B.; Kroemer, H.K.; Nauck, M.; Wallaschofski, H. Prospective association of low total testosterone concentrations with an adverse lipid profile and increased incident dyslipidemia. Eur. J. Prev. Cardiol. 2011, 18, 86–96. [Google Scholar] [CrossRef]
- Alkan, C.; Coe, P.; Eichler, E. Sex Hormone–Binding Globulin and Risk of Type 2 Diabetes in Women and Men. N. Engl. J. Med. 2011, 23, 1–7. [Google Scholar] [CrossRef]
- Thompson, D.J.; Healey, C.S.; Baynes, C.; Kalmyrzaev, B.; Ahmed, S.; Dowsett, M.; Folkerd, E.; Luben, R.N.; Cox, D.; Ballinger, D.; et al. Identification of common variants in the SHBG gene affecting sex hormone-binding globulin levels and breast cancer risk in postmenopausal women. Cancer Epidemiol. Biomark. Prev. 2008, 17, 3490–3498. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Cai, M.; Chen, J.; Huang, J.; Liu, F.; Jiang, C.; Gao, Y. Hypobaric hypoxia causes deleterious effects on spermatogenesis in rats. Reproduction 2010, 139, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, I.A.; Turki, R.F.; Abuzenadah, A.M.; Damanhouri, G.A.; Beg, M.A. Endocrine disruption: Computational perspectives on human sex hormone-binding globulin and phthalate plasticizers. PLoS ONE 2016, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Déchaud, H.; Ravard, C.; Claustrat, F.; De La Perrière, A.B.; Pugeat, M. Xenoestrogen interaction with human sex hormone-binding globulin (hSHBG). Steroids 1999, 64, 328–334. [Google Scholar] [CrossRef]
- Jury, H.H.; Zacharewski, T.R.; Hammond, G.L. Interactions between human plasma sex hormone-binding globulin and xenobiotic ligands. J. Steroid Biochem. Mol. Biol. 2000, 75, 167–176. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- DeLano, W.L. The PyMOL Molecular Graphics System; Version 2.3; Schrödinger LLC.: New York, NY, USA, 2020. [Google Scholar]
- Studio, D. Dassault Systemes BIOVIA, Discovery Studio Modelling Environment; Release 4.5; Accelrys Softw. Inc.: San Diego, CA, USA, 2015. [Google Scholar]
- Kaplan, W.; Littlejohn, T.G. Swiss-PDB Viewer (Deep View). Brief. Bioinform. 2001, 2, 195–197. [Google Scholar] [CrossRef]
- Kim, S.; Thiessen, P.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and Compound databases. Nucleic Acids Res. 2019, 47, D1202–D1213. [Google Scholar] [CrossRef]
- Halgren, T.A. Performance of MMFF94*. Scope Parameterization J. Comput. Chem. 1996, 17, 490–519. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.E.V.; Blundell, T.L.; Ascher, D.B. pkCSM: Predicting small-molecule pharmacokinetic and toxicity properties using graph-based signatures. J. Med. Chem. 2015, 58, 4066–4072. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, F.; Li, W.; Zhou, Y.; Shen, J.; Wu, Z.; Liu, G.; Lee, P.W.; Tang, Y. AdmetSAR: A comprehensive source and free tool for assessment of chemical ADMET properties. J. Chem. Inf. Model. 2012, 52, 3099–3105. [Google Scholar] [CrossRef]
- Land, H.; Humble, M.S. YASARA: A tool to obtain structural guidance in biocatalytic investigations. Methods Mol. Biol. 2018, 1685, 43–67. [Google Scholar] [CrossRef]
- Krieger, E.; Vriend, G.; Spronk, C. YASARA–Yet Another Scientific Artificial Reality Application. Yasara 2013, 993, 51–78. [Google Scholar]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and testing of a general Amber force field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef]
- Mahmud, S.; Mita, M.A.; Biswas, S.; Paul, G.K.; Promi, M.M.; Afrose, S.; Hasan, R.; Shimu, S.S.; Zaman, S.; Uddin, S.; et al. Molecular docking and dynamics study to explore phytochemical ligand molecules against the main protease of SARS-CoV-2 from extensive phytochemical datasets. Expert Rev. Clin. Pharmacol. 2021, 14, 1305–1315. [Google Scholar] [CrossRef]
- Harrach, M.F.; Drossel, B. Structure and dynamics of TIP3P, TIP4P, and TIP5P water near smooth and atomistic walls of different hydroaffinity. J. Chem. Phys. 2014, 140, 174501. [Google Scholar] [CrossRef]
- Mahmud, S.; Paul, G.K.; Biswas, S.; Afrose, S.; Mita, M.A.; Hasan, M.R.; Shimu, M.S.S.; Hossain, A.; Promi, M.M.; Ema, F.K.; et al. Prospective Role of Peptide-Based Antiviral Therapy Against the Main Protease of SARS-CoV-2. Front. Mol. Biosci. 2021, 8, 628585. [Google Scholar] [CrossRef]
- Biswas, S.; Mahmud, S.; Mita, M.A.; Afrose, S.; Hasan, M.R.; Sultana Shimu, M.S.; Saleh, M.A.; Mostafa-Hedeab, G.; Alqarni, M.; Obaidullah, A.J.; et al. Molecular Docking and Dynamics Studies to Explore Effective Inhibitory Peptides Against the Spike Receptor Binding Domain of SARS-CoV-2. Front. Mol. Biosci. 2022, 8, 1–10. [Google Scholar] [CrossRef]
- Mahmud, S.; Hasan, M.R.; Biswas, S.; Paul, G.K.; Afrose, S.; Mita, M.A.; Sultana Shimu, M.S.; Promi, M.M.; Hani, U.; Rahamathulla, M.; et al. Screening of Potent Phytochemical Inhibitors Against SARS-CoV-2 Main Protease: An Integrative Computational Approach. Front. Bioinforma. 2021, 1, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, S.; Biswas, S.; Paul, G.K.; Mita, M.A.; Promi, M.M.; Afrose, S.; Hasan, M.R.; Zaman, S.; Uddin, M.S.; Dhama, K.; et al. Plant-based phytochemical screening by targeting main protease of sars-cov-2 to design effective potent inhibitors. Biology 2021, 10, 589. [Google Scholar] [CrossRef]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef] [Green Version]
- Krieger, E.; Nielsen, J.E.; Spronk, C.A.E.M.; Vriend, G. Fast empirical pKa prediction by Ewald summation. J. Mol. Graph. Model. 2006, 25, 481–486. [Google Scholar] [CrossRef]
- Krieger, E.; Vriend, G. New ways to boost molecular dynamics simulations. J. Comput. Chem. 2015, 36, 996–1007. [Google Scholar] [CrossRef]
- Mahmud, S.; Paul, G.K.; Afroze, M.; Islam, S.; Gupt, S.B.R.; Razu, M.H.; Biswas, S.; Zaman, S.; Uddin, M.S.; Khan, M.; et al. Efficacy of phytochemicals derived from avicennia officinalis for the management of covid-19: A combined in silico and biochemical study. Molecules 2021, 26, 2210. [Google Scholar] [CrossRef] [PubMed]
- Kumar Paul, G.; Mahmud, S.; Aldahish, A.A.; Afroze, M.; Biswas, S.; Briti Ray Gupta, S.; Hasan Razu, M.; Zaman, S.; Salah Uddin, M.; Nahari, M.H.; et al. Computational screening and biochemical analysis of Pistacia integerrima and Pandanus odorifer plants to find effective inhibitors against Receptor-Binding domain (RBD) of the spike protein of SARS-Cov-2. Arab. J. Chem. 2022, 15, 103600. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, S.; Rafi, O.; Paul, G.K.; Promi, M.M.; Sharmin, M.; Shimu, S.; Biswas, S.; Emran, T.; Dhama, K. Designing a multi-epitope vaccine candidate to combat MERS—CoV by employing an immunoinformatics approach. Sci. Rep. 2021, 11, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, S.; Afrose, S.; Biswas, S.; Nagata, A.; Paul, G.K.; Mita, M.A.; Hasan, M.R.; Shimu, M.S.S.; Zaman, S.; Uddin, M.S.; et al. Plant-derived compounds effectively inhibit the main protease of SARS-CoV-2: An in silico approach. PLoS ONE 2022, 17, e0273341. [Google Scholar] [CrossRef]
- Mahmud, S.; Biswas, S.; Kumar Paul, G.; Mita, M.A.; Afrose, S.; Robiul Hasan, M.; Sharmin Sultana Shimu, M.; Uddin, M.A.R.; Salah Uddin, M.; Zaman, S.; et al. Antiviral peptides against the main protease of SARS-CoV-2: A molecular docking and dynamics study. Arab. J. Chem. 2021, 14, 103315. [Google Scholar] [CrossRef] [PubMed]
- Mahfuz, A.M.U.; Khan, A.; Biswas, S.; Afrose, S.; Mahmud, S.; Mohammed Bahadur, N.; Ahmed, F. In search of inhibitors of anti-cancer drug target Fibroblast Growth Factor Receptors: Insights from virtual screening, molecular docking, and molecular dynamics study. Arab. J. Chem. 2022, 15, 103882. [Google Scholar] [CrossRef]
- Islam, S.; Pramanik, M.J.; Biswas, S.; Moniruzzaman, M.; Biswas, J. Biological Efficacy of Compounds from Stingless Honey and Sting Honey against Two Pathogenic Bacteria: An In Vitro and In Silico Study. Molecules 2022, 27, 6536. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Kumar, R.; Consortium, O.S.D.D.; Lynn, A. g _ mmpbsa—A GROMACS tool for MM-PBSA and its optimization for high-throughput binding energy calculations. J. Chem. Inf. Model. 2014, 54, 1951–1962. [Google Scholar] [CrossRef] [PubMed]
- Gohlke, H.; Case, D.A. Converging Free Energy Estimates: MM-PB(GB)SA Studies on the Protein-Protein Complex Ras-Raf. J. Comput. Chem. 2004, 25, 238–250. [Google Scholar] [CrossRef]
- Åqvist, J.; Medina, C.; Samuelsson, J.E. A new method for predicting binding affinity in computer-aided drug design. Protein Eng. 2010, 7, 1–7. [Google Scholar] [CrossRef]
- Kuhn, B.; Gerber, P.; Schulz-Gasch, T.; Stahl, M. Validation and use of the MM-PBSA approach for drug discovery. J. Med. Chem. 2005, 48, 4040–4048. [Google Scholar] [CrossRef]
- Dash, R.; Junaid, M.; Mitra, S.; Arifuzzaman, M.; Hosen, S.M.Z. Structure-based identification of potent VEGFR-2 inhibitors from in vivo metabolites of a herbal ingredient. J. Mol. Model. 2019, 25, 98. [Google Scholar] [CrossRef]
- Lipinski, C.; Lipinski, C.A. Lead- and drug-like compounds: The rule-of-five revolution. Drug Discov. Today Technol. 2004, 1, 337–341. [Google Scholar] [CrossRef]
- Mahmud, S.; Paul, G.K.; Biswas, S.; Kazi, T.; Mahbub, S.; Mita, M.A.; Afrose, S.; Islam, A.; Ahaduzzaman, S.; Hasan, R.; et al. Phytochemdb: A Platform for Virtual Screening and Computer-Aided Drug Designing. Database 2022, 2022, 1–14. [Google Scholar] [CrossRef]
- Savojardo, C.; Manfredi, M.; Martelli, P.L.; Casadio, R. Solvent Accessibility of Residues Undergoing Pathogenic Variations in Humans: From Protein Structures to Protein Sequences. Front. Mol. Biosci. 2021, 7, 626363. [Google Scholar] [CrossRef]
- Islam, S.; Mahmud, L.; Almalki, W.H.; Biswas, S.; Islam, A. Cell-Free Supernatants ( CFSs ) from the Culture of Bacillus subtilis Inhibit Pseudomonas sp. Biofilm Formation. Microorganisms 2022, 10, 2105. [Google Scholar] [CrossRef]
- Kamiński, M.; Ding, M.; Truccolo, W.A.; Bressler, S.L. Evaluating causal relations in neural systems: Granger causality, directed transfer function and statistical assessment of significance. Biol. Cybern. 2001, 85, 145–157. [Google Scholar] [CrossRef]
- Del Mar Grasa, M.; Gulfo, J.; Camps, N.; Alcalá, R.; Monserrat, L.; Moreno-Navarrete, J.M.; Ortega, F.J.; Esteve, M.; Remesar, X.; Fernández-López, J.A.; et al. Modulation of SHBG binding to testosterone and estradiol by sex and morbid obesity. Eur. J. Endocrinol. 2017, 176, 393–404. [Google Scholar] [CrossRef] [Green Version]
- Corvol, P.L.; Chrambach, A.; Rodbard, D.; Bardin, C.W. Physical Properties and Binding Capacity of Testosterone-Estradiol-binding Globulin in Human Plasma, Determined by Polyacrylamide Gel Electrophoresis. J. Biol. Chem. 1971, 246, 3435–3443. [Google Scholar] [CrossRef]
- Hazarika, J.; Ganguly, M.; Mahanta, R. Molecular interactions of chlorpyrifos and its environmental degradation products with human sex hormone-binding globulin: An in silico study. J. Appl. Toxicol. 2019, 39, 1002–1011. [Google Scholar] [CrossRef]
- Borquaye, L.S.; Gasu, E.N.; Ampomah, G.B.; Kyei, L.K.; Amarh, M.A.; Mensah, C.N.; Nartey, D.; Commodore, M.; Adomako, A.K.; Acheampong, P.; et al. Alkaloids from Cryptolepis sanguinolenta as Potential Inhibitors of SARS-CoV-2 Viral Proteins: An in Silico Study. Biomed Res. Int. 2020, 2020, 1–14. [Google Scholar] [CrossRef]
- Jiménez-Avalos, G.; Vargas-Ruiz, A.P.; Delgado-Pease, N.E.; Olivos-Ramirez, G.E.; Sheen, P.; Fernández-Díaz, M.; Quiliano, M.; Zimic, M.; Agurto-Arteaga, A.; Antiparra, R.; et al. Comprehensive virtual screening of 4.8 k flavonoids reveals novel insights into allosteric inhibition of SARS-CoV-2 MPRO. Sci. Rep. 2021, 11, 1–19. [Google Scholar] [CrossRef]
- Gyebi, G.A.; Ogunyemi, O.M.; Ibrahim, I.M.; Ogunro, O.B.; Adegunloye, A.P.; Afolabi, S.O. SARS-CoV-2 host cell entry: An in silico investigation of potential inhibitory roles of terpenoids. J. Genet. Eng. Biotechnol. 2021, 19, 1–22. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | Pubchem CID | Docking Score (kcal/mole) |
|---|---|---|
| Cryptomisrine | 10600127 | −9.2 |
| Dorsilurin E | 15478906 | −9 |
| Isoiguesterin | 11373102 | −8.8 |
| (-)-Catechin gallate | 6419835 | −8.6 |
| 2,3-dihydroamentoflavone | 16066857 | −8.4 |
| Epicatechin 3,5-di-O-gallate | 14284594 | −8.4 |
| Theaflavine | 135403798 | −8.2 |
| Fortunellin | 5317385 | −8.1 |
| Cassigarol G | 10005549 | −8.1 |
| Pseudojervine | 16398499 | −8 |
| Compound Name | Pubchem CID | Docking Score | Residues in Contact | Interaction Type | Distance in Å |
|---|---|---|---|---|---|
| Cryptomisrine | 10600127 | −9.2 | MET30 | Conventional Hydrogen Bond | 2.8018 |
| PRO14 | Conventional Hydrogen Bond | 1.78914 | |||
| ASP168 | Electrostatic (Pi-Anion) | 4.64313 | |||
| SER169 | Hydrophobic (Pi-Sigma) | 2.88827 | |||
| LYS173 | Hydrophobic (Pi-Alkyl) | 4.70796 | |||
| ALA28 | Hydrophobic (Pi-Alkyl) | 5.006 | |||
| VAL16 | Hydrophobic (Pi-Alkyl) | 4.3933 | |||
| LEU185 | Hydrophobic (Pi-Alkyl) | 4.90543 | |||
| Dorsilurin E | 15478906 | −9 | LYS173 | Conventional Hydrogen Bond | 2.71543 |
| ASP168 | Electrostatic (Pi-Anion) | 3.87237 | |||
| PRO14 | Hydrophobic (Alkyl) | 5.24634 | |||
| LEU185 | Hydrophobic (Alkyl) | 4.233 | |||
| VAL16 | Hydrophobic (Alkyl) | 4.08529 | |||
| MET30 | Hydrophobic (Alkyl) | 5.21275 | |||
| LEU143 | Hydrophobic (Alkyl) | 4.93558 | |||
| TRP170 | Hydrophobic (Pi-Alkyl) | 5.14218 | |||
| Isoiguesterin | 11373102 | −8.8 | ALA28 | Hydrophobic (Alkyl) | 4.55253 |
| MET30 | Hydrophobic (Alkyl) | 5.09204 | |||
| ALA179 | Hydrophobic (Alkyl) | 4.91895 | |||
| LEU185 | Hydrophobic (Alkyl) | 4.57361 | |||
| PRO182 | Hydrophobic (Alkyl) | 4.51348 | |||
| VAL16 | Hydrophobic (Alkyl) | 4.71484 | |||
| LYS173 | Hydrophobic (Alkyl) | 3.86922 | |||
| Anastrazole | 2187 | −7.0 | VAL29 | Conventional Hydrogen Bond | 2.93091 |
| SER180 | Conventional Hydrogen Bond | 2.89776 | |||
| ASP168 | Conventional Hydrogen Bond | 2.82215 | |||
| GLU176 | Electrostatic (Pi-Anion) | 4.86148 | |||
| ALA28 | Hydrophobic (Alkyl) | 4.35855 | |||
| ALA179 | Hydrophobic (Alkyl) | 4.14398 | |||
| PRO182 | Hydrophobic (Alkyl) | 4.48299 | |||
| LEU185 | Hydrophobic (Alkyl) | 4.43503 |
| Parameters | Cryptomisrine | Dorsilurin E | Isoiguesterin |
|---|---|---|---|
| Molecular weight | 462.5 g/mol | 490.6 g/mol | 404.6 g/mol |
| Num. H-bond acceptors | 3 | 6 | 2 |
| Num. H-bond donors | 2 | 1 | 1 |
| TPSA (S) | 74.43 Å2 | 74.22 Å2 | 37.30 Å2 |
| AMES toxicity | No | No | No |
| Human intestinal absorption | 96.507 (% Absorbed) | 93.133 (% Absorbed) | 95.798 (% Absorbed) |
| Skin Sensitization | No | No | No |
| P-glycoprotein substrate | No | No | No |
| CNS permeability | −1.073 | −2.703 | −1.955 |
| Carcinogenicity | Non-carcinogenic | Non-carcinogenic | Non-carcinogenic |
| Acute oral toxicity | No | No | No |
| BBB permeability | 0.7629 | −0.368 | −0.202 |
| Hepatotoxicity | No | No | No |
| Lipinski rule of five | Yes; 1 violation | Yes; 0 violation | Yes; 1 violation |
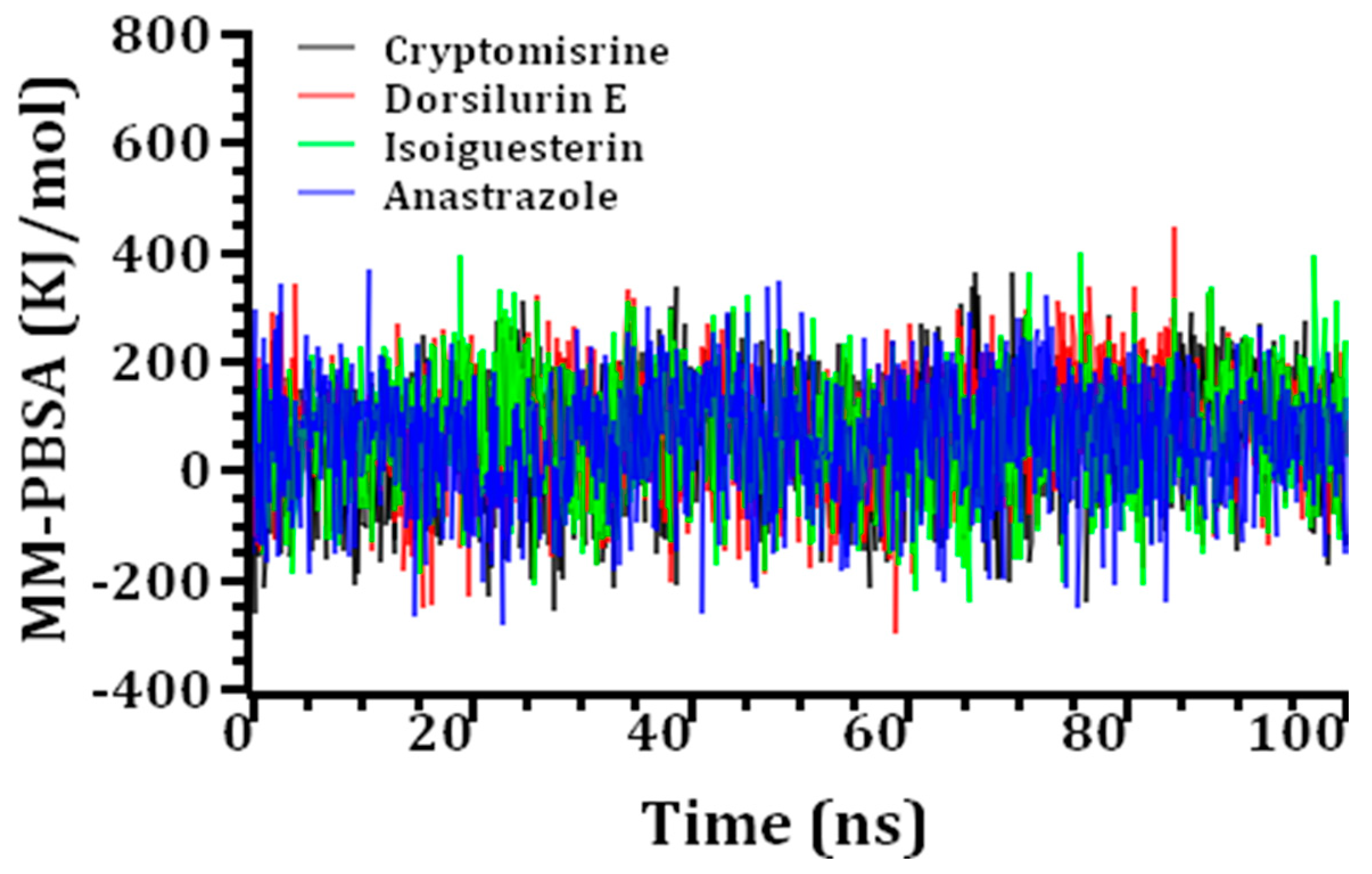
| Compound Name | Average Binding Free Energy (KJ/mol) |
|---|---|
| Cryptomisrine | 67.64 |
| Dorsilurin E | 71.39 |
| Isoiguesterin | 69.13 |
| Anastrazole | 57.38 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biswas, S.; Mita, M.A.; Afrose, S.; Hasan, M.R.; Islam, M.T.; Rahman, M.A.; Ara, M.J.; Chowdhury, M.B.A.; Meem, H.N.; Mamunuzzaman, M.; et al. Integrated Computational Approaches for Inhibiting Sex Hormone-Binding Globulin in Male Infertility by Screening Potent Phytochemicals. Life 2023, 13, 476. https://doi.org/10.3390/life13020476
Biswas S, Mita MA, Afrose S, Hasan MR, Islam MT, Rahman MA, Ara MJ, Chowdhury MBA, Meem HN, Mamunuzzaman M, et al. Integrated Computational Approaches for Inhibiting Sex Hormone-Binding Globulin in Male Infertility by Screening Potent Phytochemicals. Life. 2023; 13(2):476. https://doi.org/10.3390/life13020476
Chicago/Turabian StyleBiswas, Suvro, Mohasana Akter Mita, Shamima Afrose, Md. Robiul Hasan, Md. Tarikul Islam, Md. Ashiqur Rahman, Mst. Jasmin Ara, Md. Bakhtiar Abid Chowdhury, Habibatun Naher Meem, Md. Mamunuzzaman, and et al. 2023. "Integrated Computational Approaches for Inhibiting Sex Hormone-Binding Globulin in Male Infertility by Screening Potent Phytochemicals" Life 13, no. 2: 476. https://doi.org/10.3390/life13020476